Estimating Rangeland Forage Production Using Remote Sensing Data from a Small Unmanned Aerial System (sUAS) and PlanetScope Satellite

 ,
, 
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
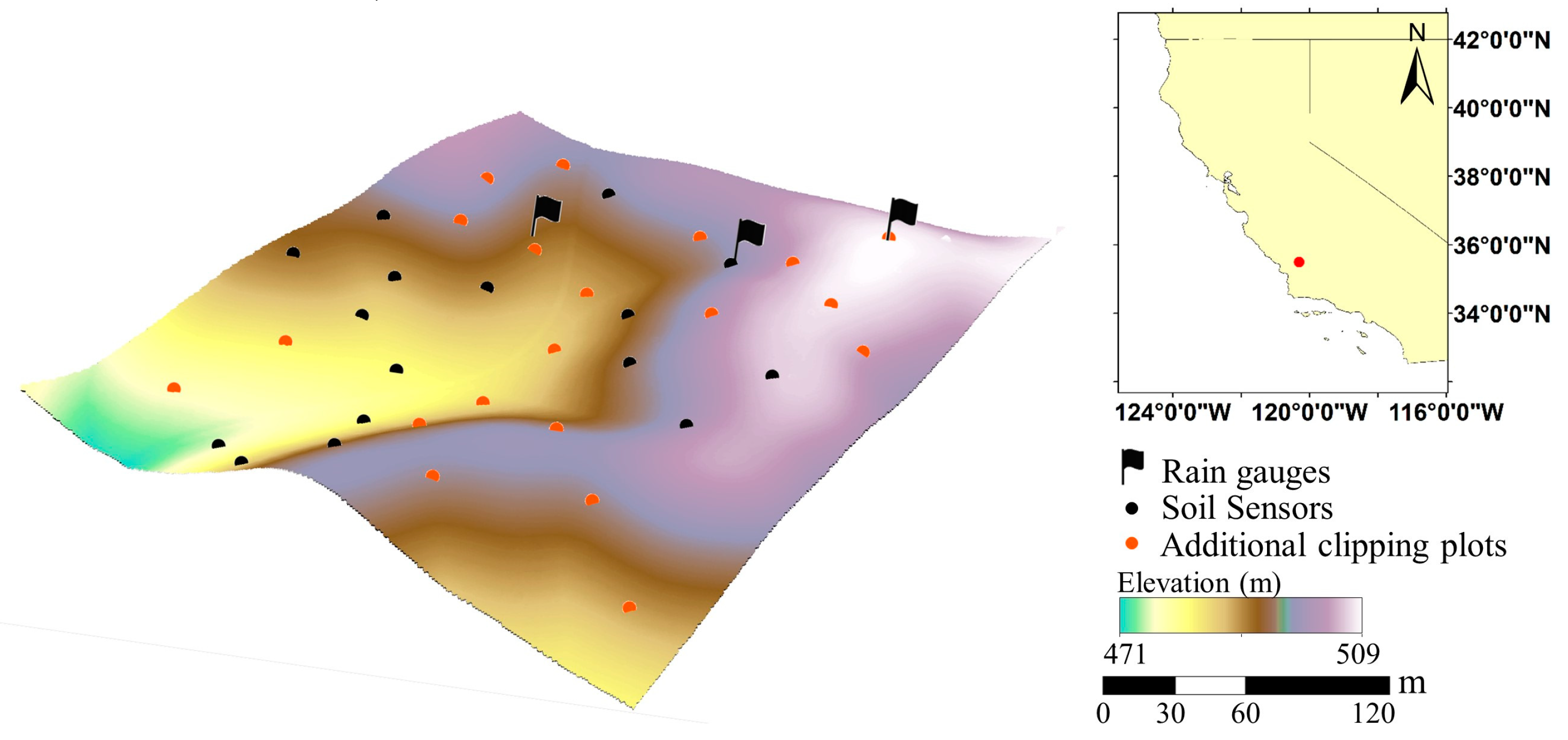
2.1. Study Site
2.2. Field Measurements
2.3. sUAS Flights and Image Preprocessing
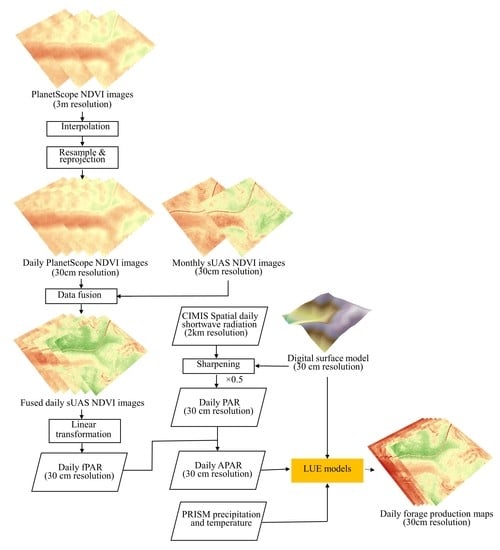
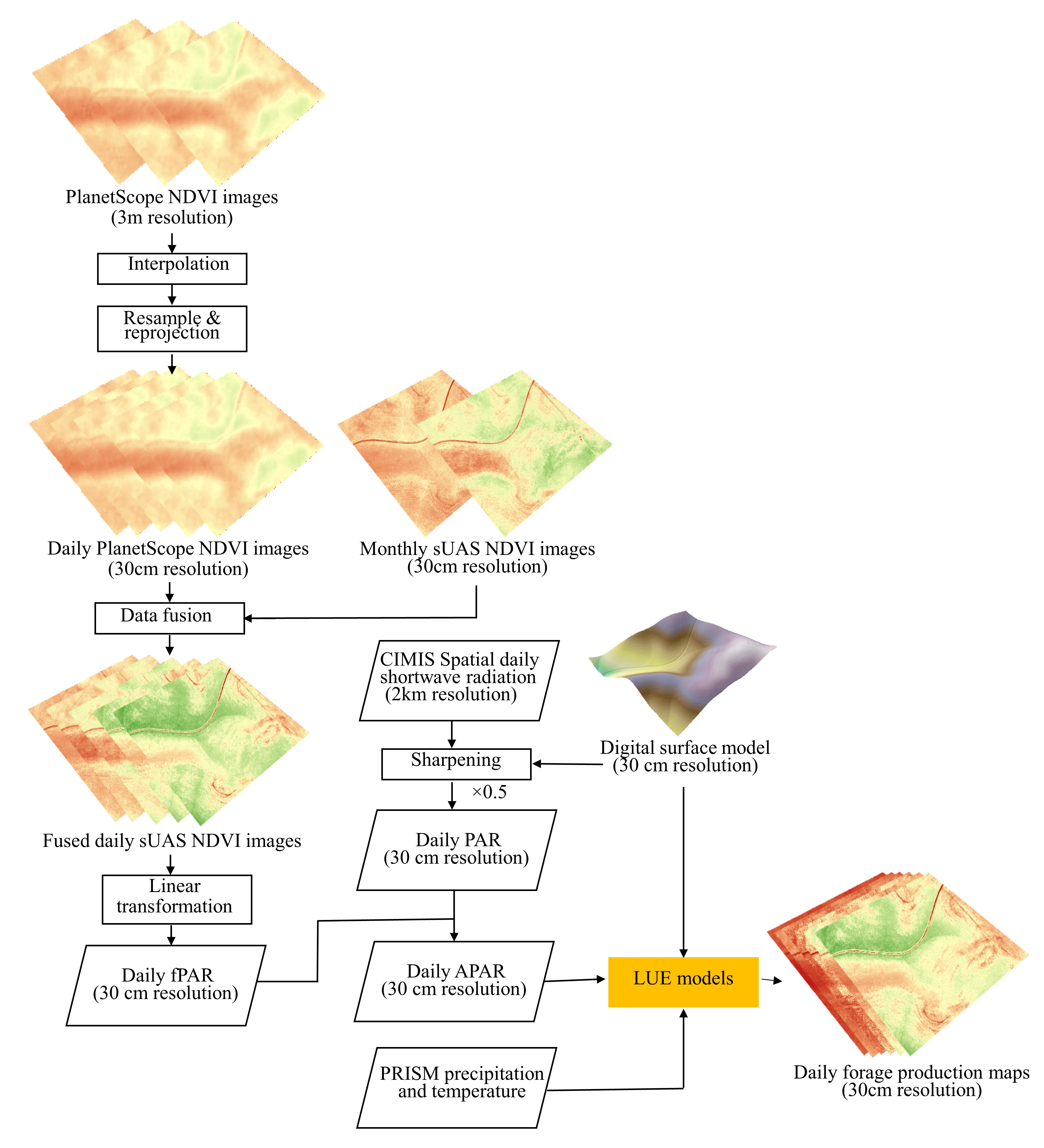
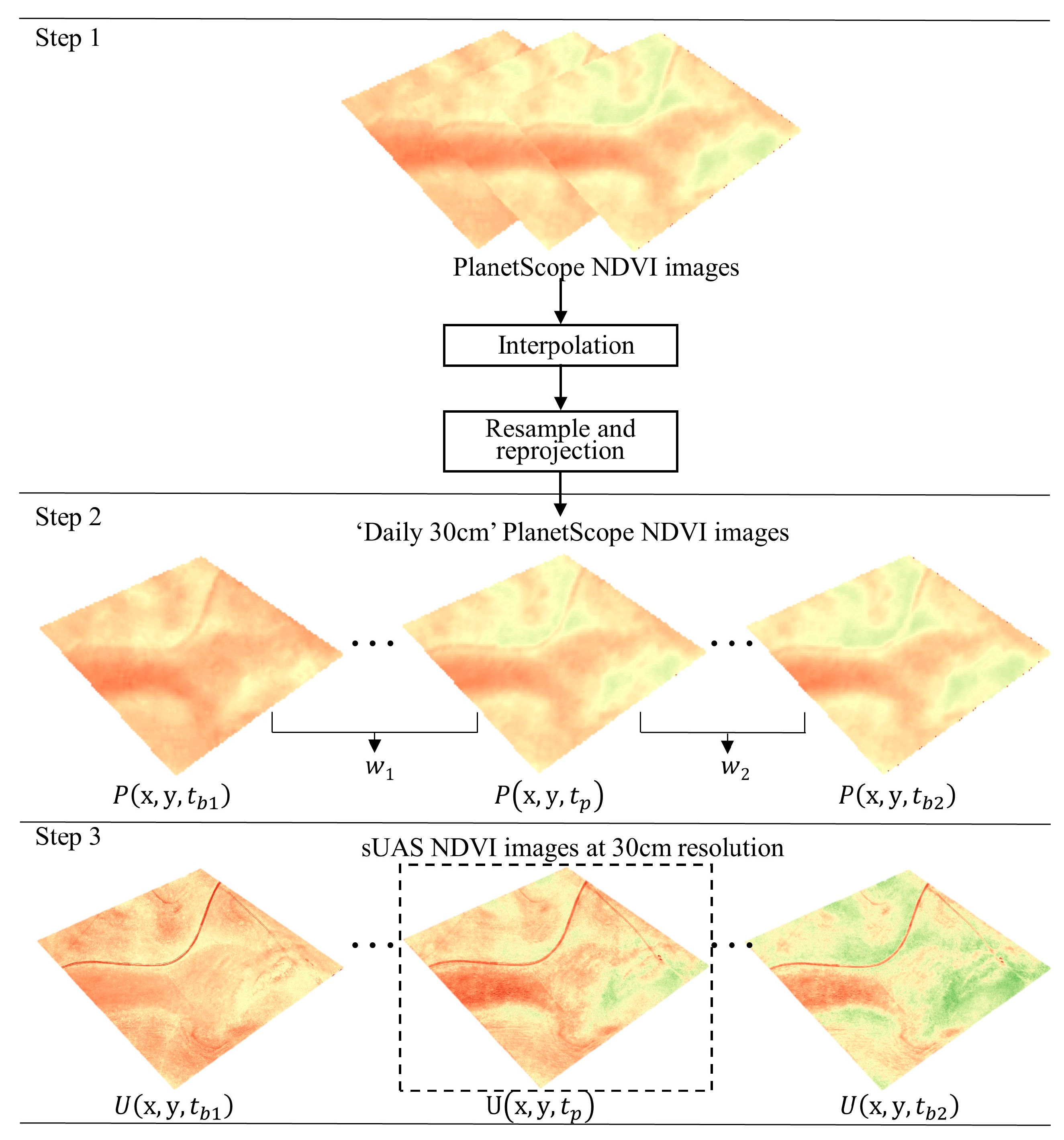
2.4. High Resolution Satellite Imagery and Data Fusion
2.5. Forage Production Estimation Methods and Assessment
2.5.1. Empirical Statistical Analysis
2.5.2. Light Use Efficiency (LUE) Models
2.6. Forage Production Mapping and Patterns
3. Results
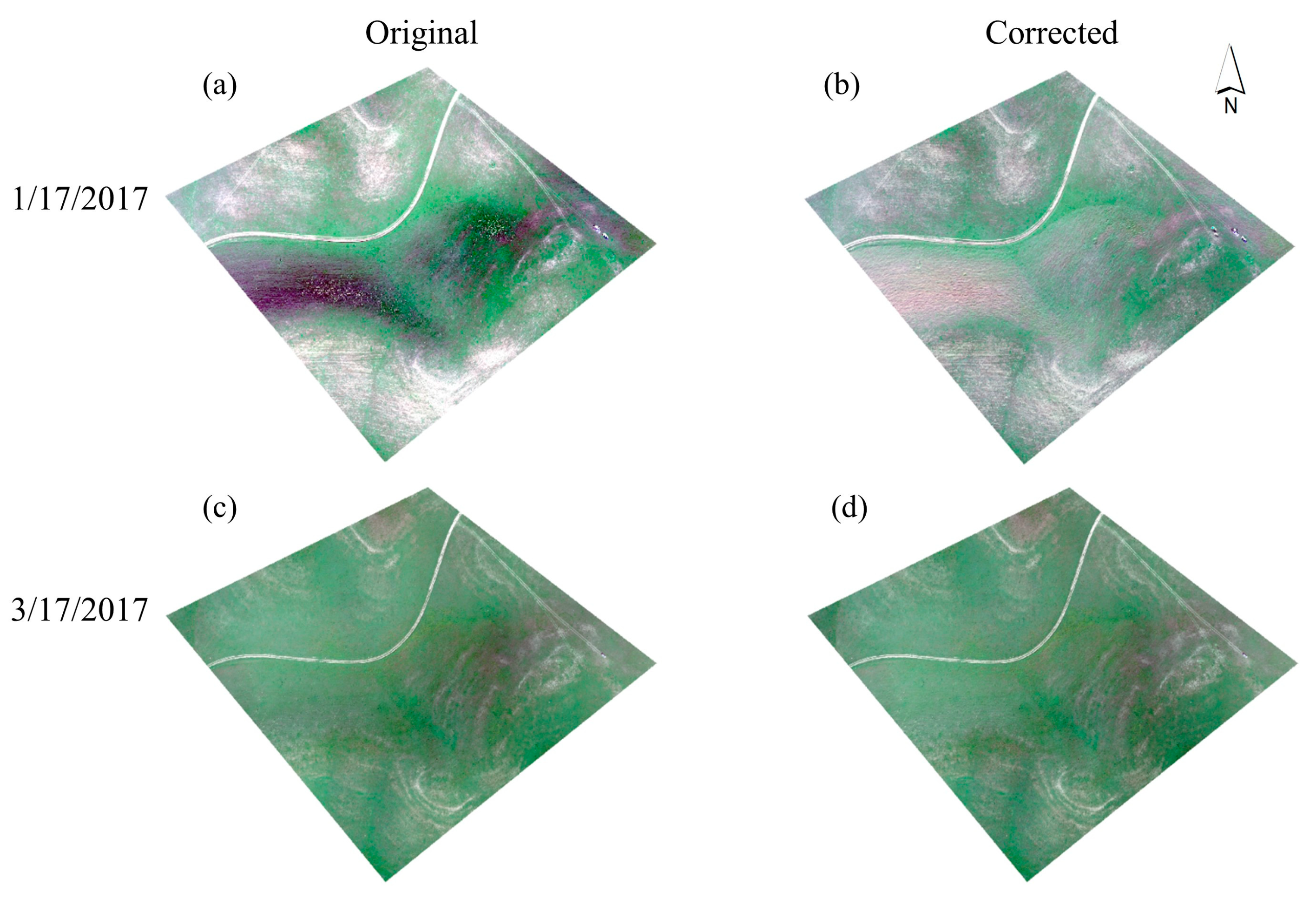
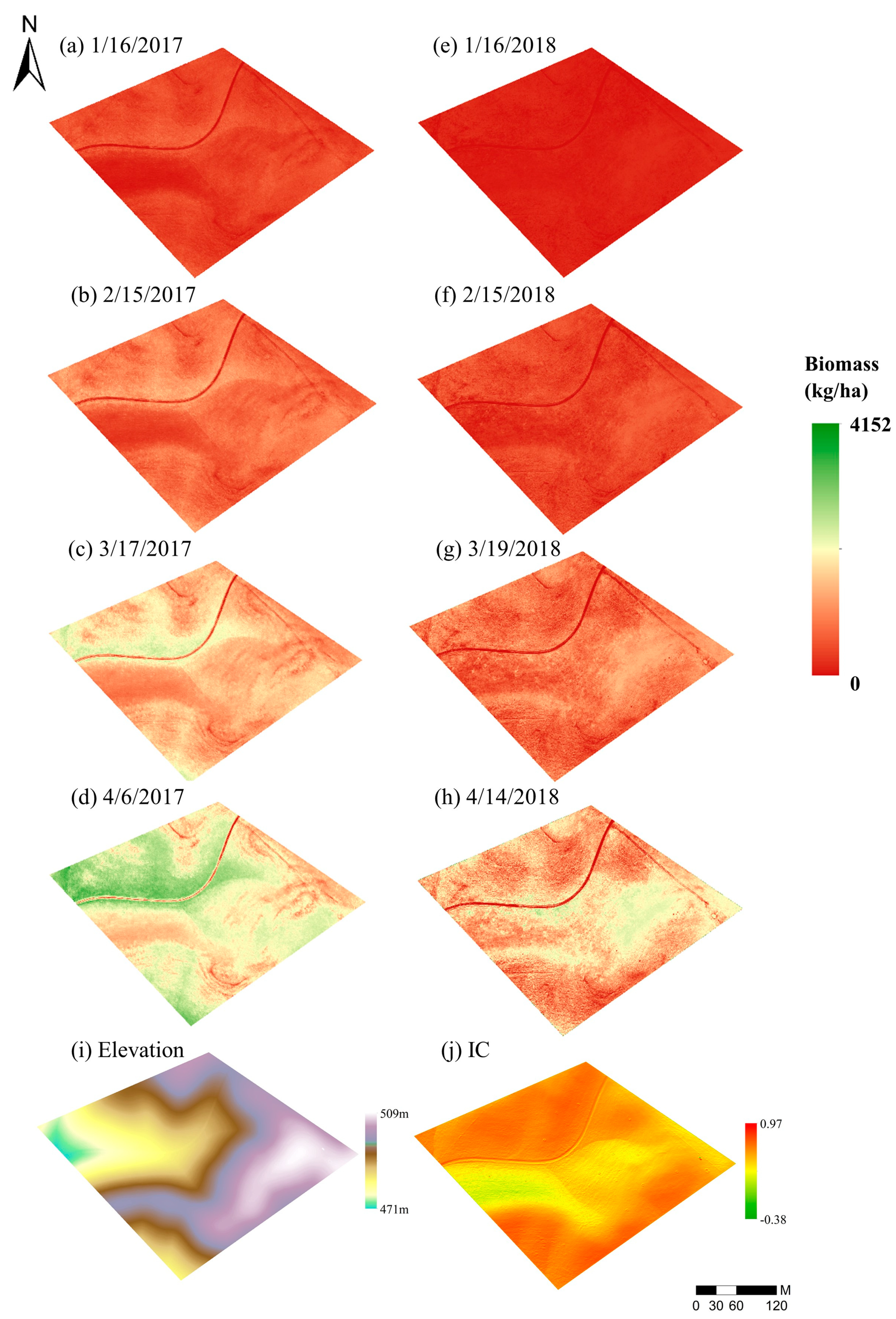
3.1. Terrain Correction
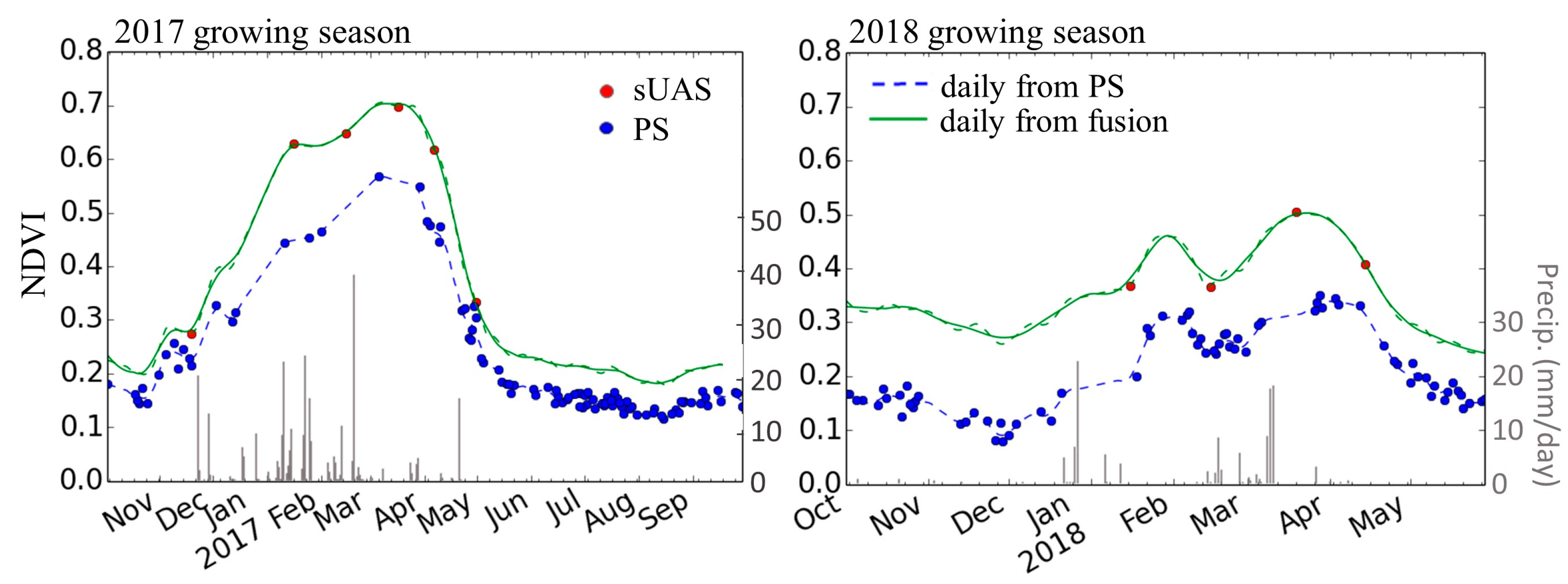
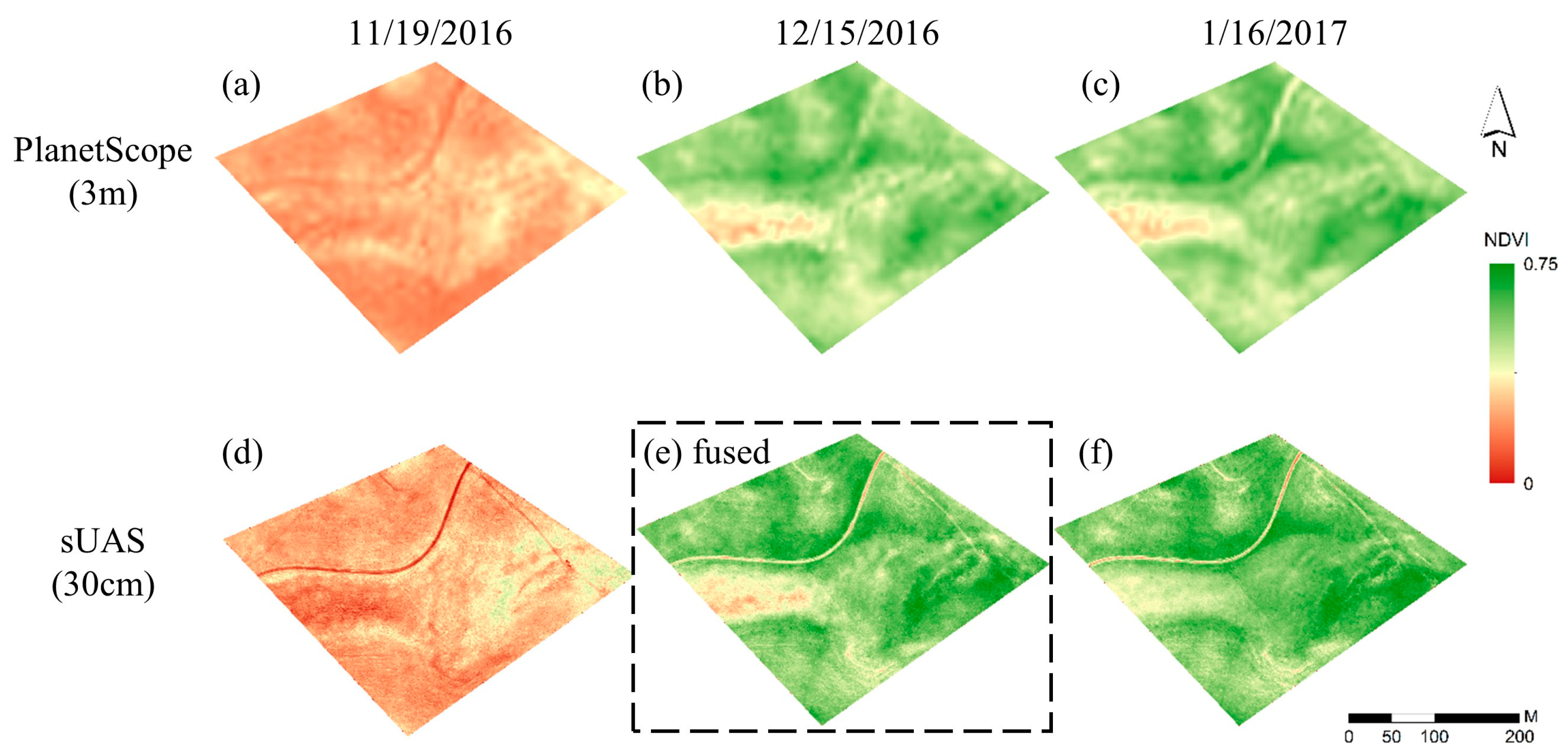
3.2. sUAS and PlanetScope Data Fusion
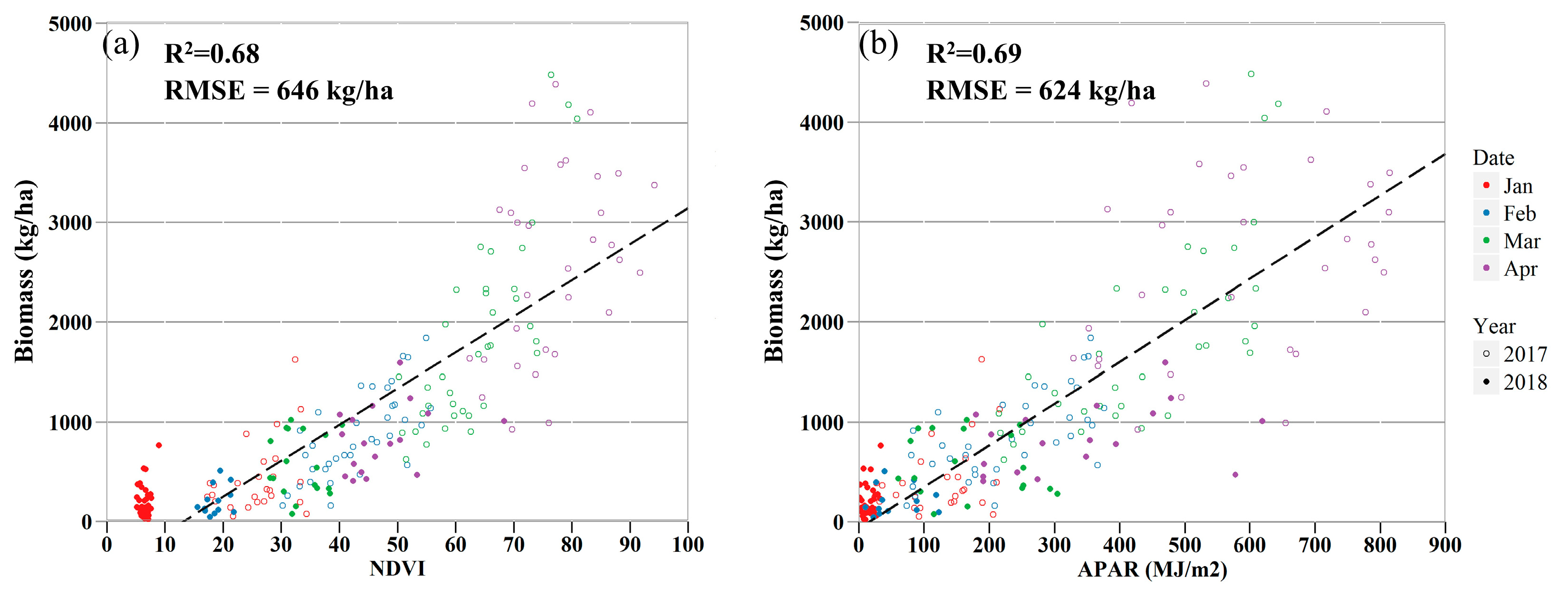
3.3. Relationships Between Remote Sensing Metrics and Biomass Measurements
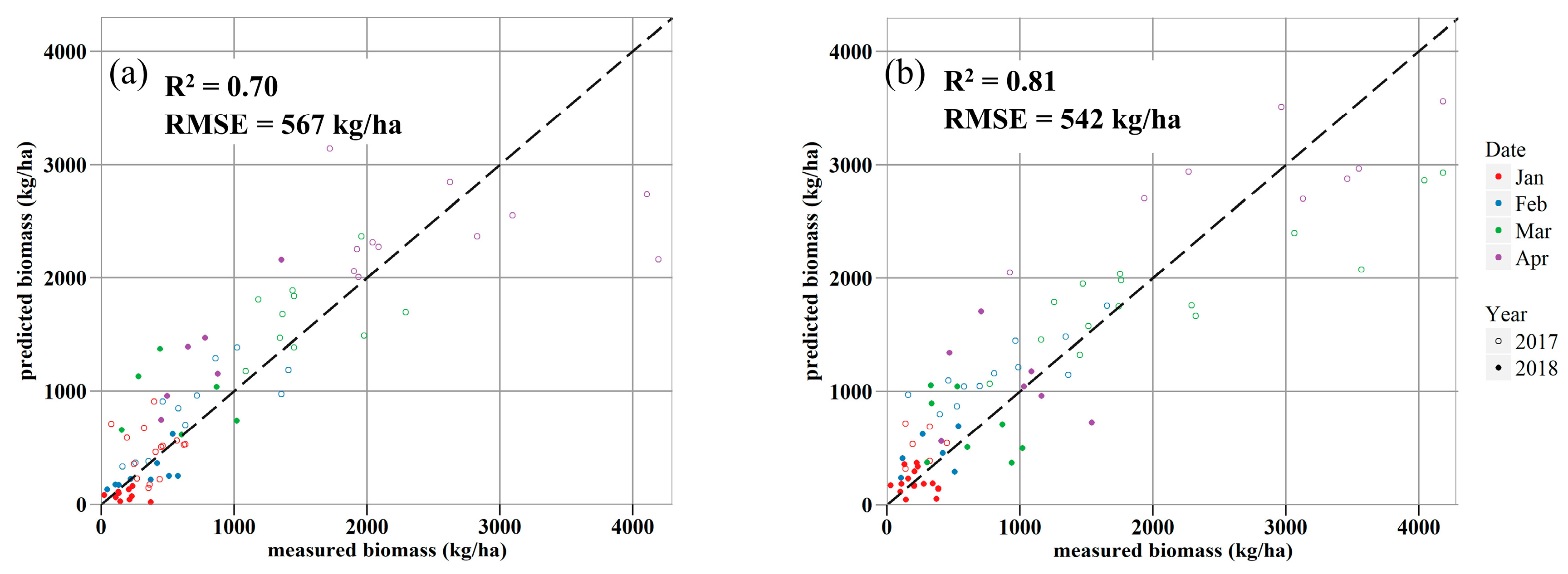
3.4. Forage Production Models
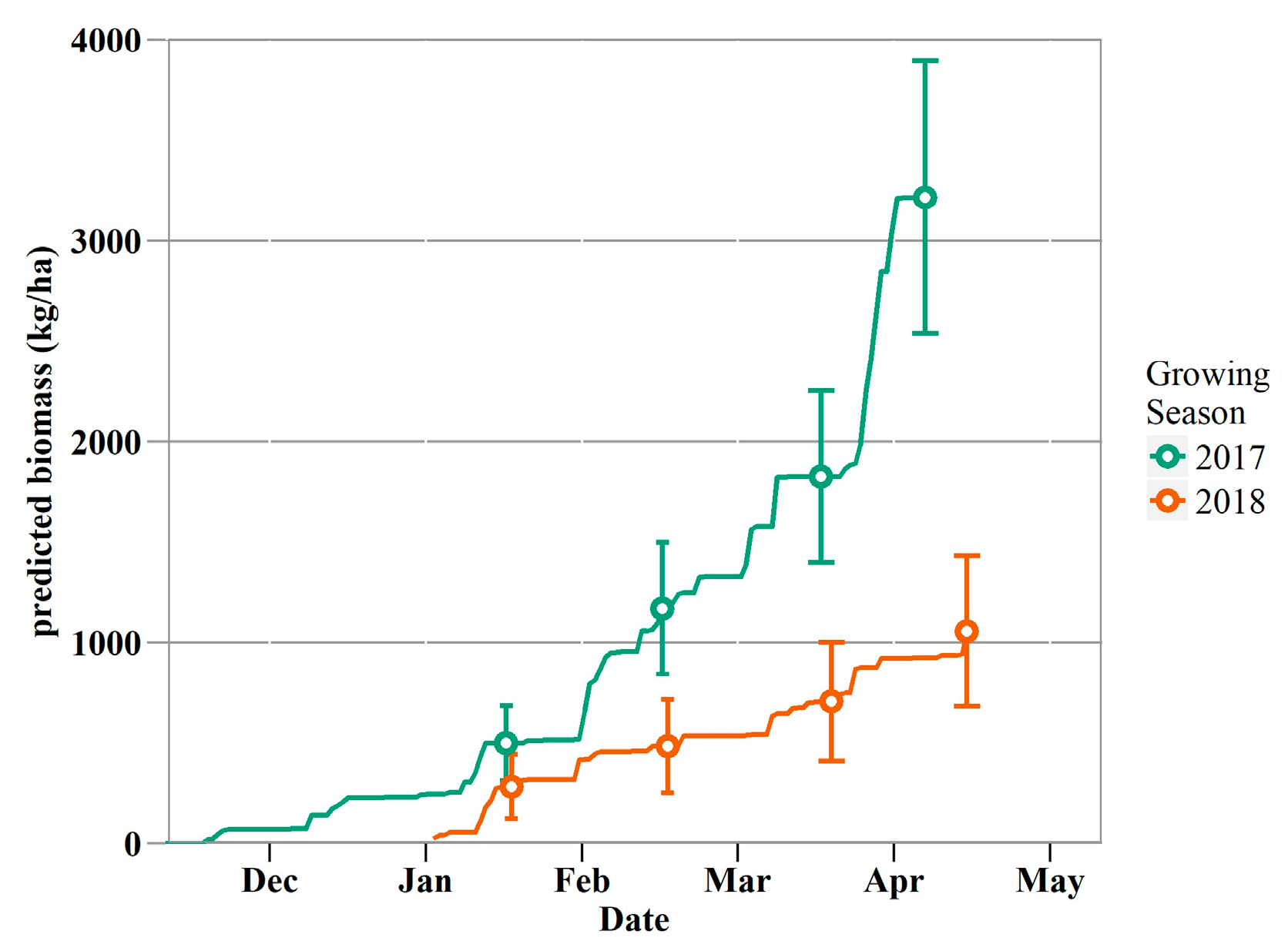
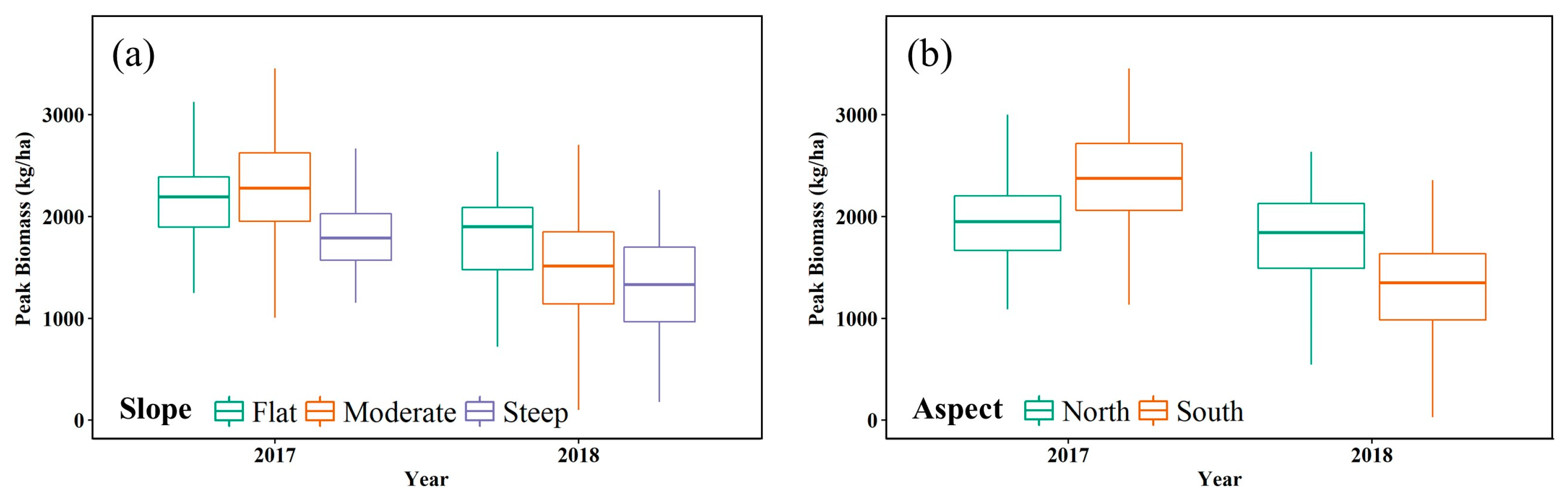
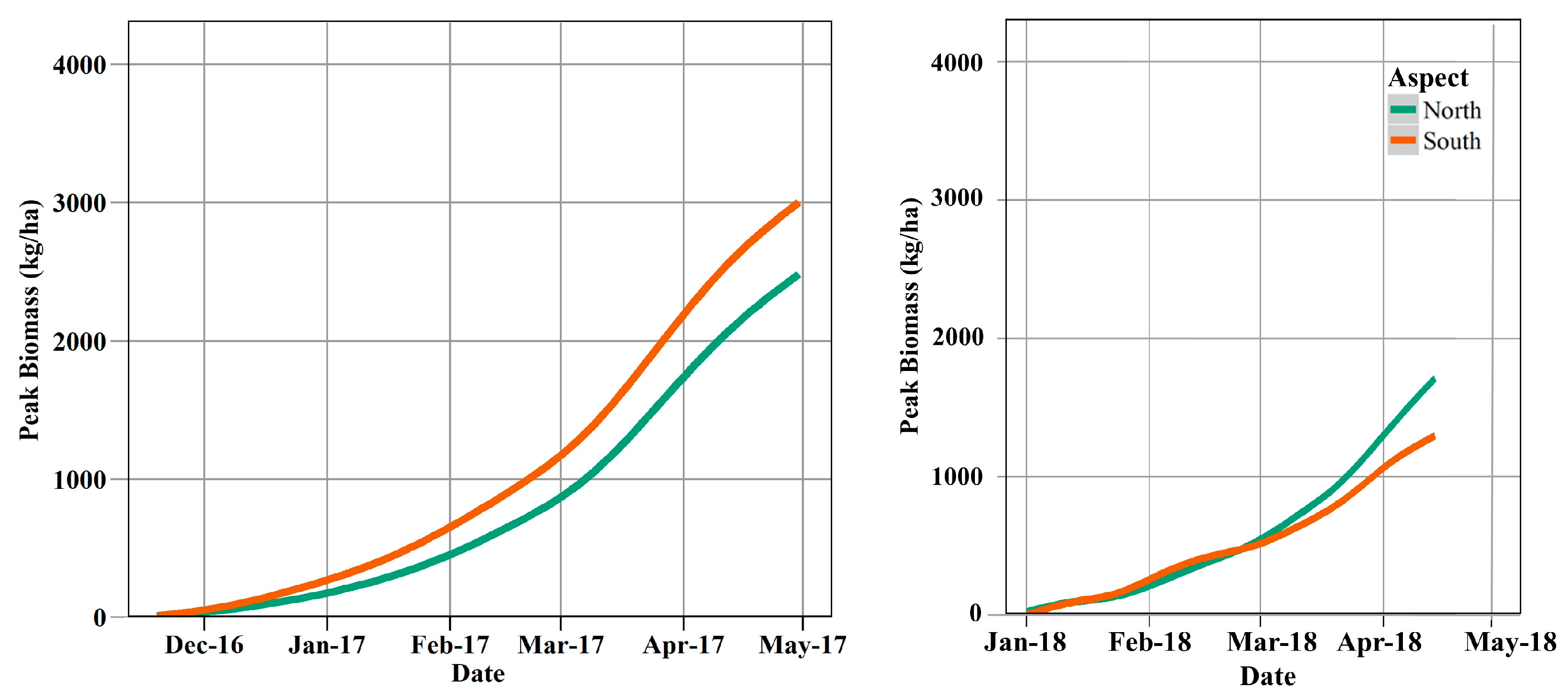
3.5. Forage Production Mapping and Patterns
4. Discussion
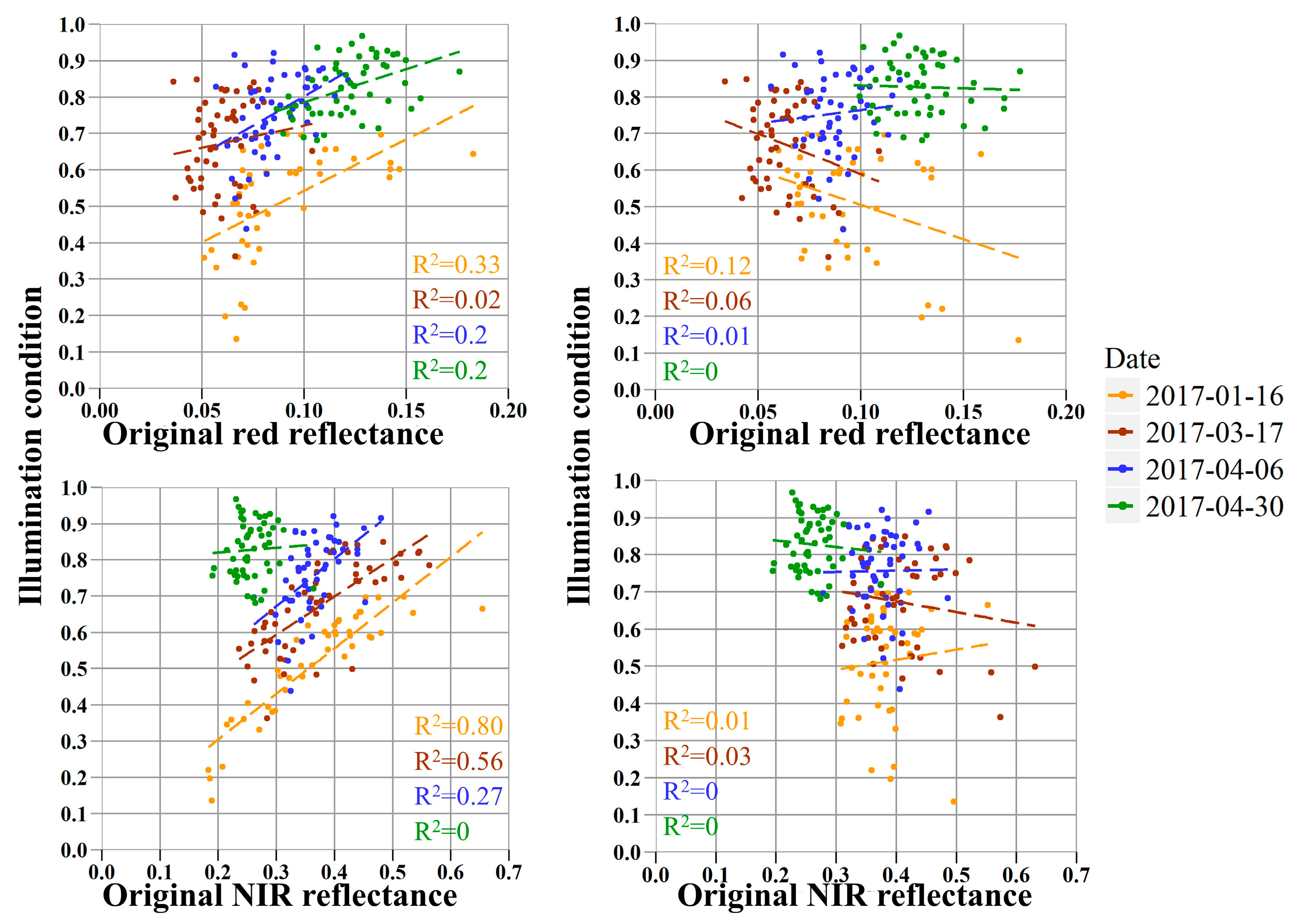
4.1. Variations in the Illumination Effect
4.2. Fusing Satellite and sUAS Data
4.3. LUE Parameterization
4.4. Response of Forage Production and Plant Phenology to Moisture
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Roche, L.M.; Schohr, T.K.; Derner, J.D.; Lubell, M.N.; Cutts, B.B.; Kachergis, E.; Eviner, V.T.; Tate, K.W. Sustaining Working Rangelands: Insights from Rancher Decision Making. Rangel. Ecol. Manag. 2015, 68, 383–389. [Google Scholar] [CrossRef]
- Schwarzenegger, A.; Snow, L.; Walters, D. California’s Forests and Rangelands: 2010 Assessment. Available online: http://frap.fire.ca.gov/data/assessment2010/pdfs/california_forest_assessment_nov22.pdf (accessed on 17 April 2017).
- California Department of Food & Agriculture. 2015 Crop Year Report; California Department of Food & Agriculture: Sacramento, CA, USA, 2015.
- Becchetti, T.; George, M.; McDougald, N.; Dudley, D.M.; Connor, M.; Flavell, D.K.; Vaughn, C.E.; Forero, L.C.; Frost, B.; Oneto, S.R.; et al. Annual Range Forage Production. Univ. Calif. Agric. Nat. Resour. Publ. 2016, 8018, 1–12. [Google Scholar]
- Sloat, L.L.; Gerber, J.S.; Samberg, L.H.; Smith, W.K.; Herrero, M.; Ferreira, L.G.; Godde, C.M.; West, P.C. Increasing Importance of Precipitation Variability on Global Livestock Grazing Lands. Nat. Clim. Chang. 2018, 8, 214–218. [Google Scholar] [CrossRef]
- Larsen, R.; Striby, K.; Horney, M. Fourteen Years of Frage Monitoring on the California Central Coast Shows Tremendous Variation; General Technical Reports PSW-GTR-251; US Department of Agriculture, Forest Service, Pacific Southwest Research Station: Berkeley, CA, USA, 2014; pp. 273–281.
- Intergovernmental Panel on Climate Change [IPCC]. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T., Qin, D., Plattner, G., Tignor, M., Allen, S., Bosching, J., Nauels, A., Xia, Y., Bex, V., Midgley, P., Eds.; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Coppock, D.L.; Fernández-Giménez, M.; Hiernaux, P.; Huber-Sannwald, E.; Schloeder, C.; Valdivia, C.; Arredondo, J.T.; Jacobs, M.; Turin, C.; Turner, M. Rangeland Systems in Developing Nations: Conceptual Advances and Societal Implications. In Rangeland Systems; Briske, D.D., Ed.; Springer: Cham, Switzerland, 2017; pp. 569–641. ISBN 978-3-319-46707-8. [Google Scholar]
- George, M.R.; Williams, W.A.; McDougald, N.K.; Clawson, W.J.; Murphy, A.H. Predicting Peak Standing Crop on Annual Range Using Weather Variables. J. Range Manag. 1989, 42, 508–512. [Google Scholar] [CrossRef]
- Liacos, L.G. Soil Moisture Depletion in the Annual Grass Type. J. Range Manag. 1962, 15, 67–72. [Google Scholar] [CrossRef]
- O’ Geen, A.T. Soil Water Dynamics. Nat. Educ. Knowl. 2012, 3, 12. [Google Scholar]
- Beaudette, D.E.; O’Geen, A.T. Quantifying the Aspect Effect: An Application of Solar Radiation Modeling for Soil Survey. Soil Sci. Soc. Am. J. 2009, 73, 1755. [Google Scholar] [CrossRef]
- Wang, J.Y. A Critique of the Heat Unit Approach to Plant Response Studies. Ecology 1960, 41, 785–790. [Google Scholar] [CrossRef]
- George, M.R.; Raguse, C.A.; Clawson, W.J.; Wilson, C.B.; Willoughby, R.L.; McDougald, N.K.; Duncan, D.A.; Murphy, A.H. Correlation of Degree-Days with Annual Herbage Yields and Livestock Gains. J. Range Manag. 1988, 41. [Google Scholar] [CrossRef]
- Murphy, A.H. Predicted Forage Yield Based on Fall Precipitation in California Annual Grasslands. J. Range Manag. 1970, 23, 363–365. [Google Scholar] [CrossRef]
- Woodmansee, R.G. Critique and Analyses of the Grassland Ecosystem Model ELM. In Grassland Simulation Model; Innis, G.S., Ed.; Springer: New York, NY, USA, 1978; pp. 257–281. ISBN 978-1-4612-9929-5. [Google Scholar]
- Wright, J.; Skiles, J. SPUR Simulation of Production and Utilization of Rangelands: Documentation and User Guide; USDA-Agricultural Research Service: Washington, DC, USA, 1987.
- Pierson, F.B.; Carlson, D.H.; Spaeth, K.E. A Process-based Hydrology Submodel Dynamically Linked to the Plant Component of the Simulation of Production and Utilization on Rangelands SPUR Model. Ecol. Model. 2001, 141, 241–260. [Google Scholar] [CrossRef]
- Stout, W.L.; Vona, L.C.; Skiles, J.W.; Shaffer, J.A.; Jung, G.A.; Reid, R.L. Evaluating SPUR Model for Predicting Animal Gains and Biomass on Eastern Hill Land Pastures. Agric. Syst. 1990, 34, 169–178. [Google Scholar] [CrossRef]
- Corson, M.S.; Skinner, R.H.; Rotz, C.A. Modification of the SPUR Rangeland Model to Simulate Species Composition and Pasture Productivity in Humid Temperate Regions. Agric. Syst. 2006, 87, 169–191. [Google Scholar] [CrossRef]
- Monteith, J.L. Solar Radiation and Productivity in Tropical Ecosystems. J. Appl. Ecol. 1972, 9, 747. [Google Scholar] [CrossRef]
- Potter, C.S.; Randerson, J.T.; Field, C.B.; Matson, P.A.; Vitousek, P.M.; Mooney, H.A.; Klooster, S.A. Terresrial Ecosystem Production: A Process Model Based on Global Satellite and Surface Data. Glob. Biogeochem. Cycles 1993, 7, 811–841. [Google Scholar] [CrossRef]
- Li, S. Monitoring of Net Primary Production in California Rangelands Using Landsat and MODIS Satellite Remote Sensing. Nat. Resour. 2012, 3, 56–65. [Google Scholar] [CrossRef]
- Goetz, S.J.; Prince, D.; Goward, N.; Thawley, M.M.; Small, J.; Johnston, A. Mapping Net Primary Production and Related Biophysical Variables with Remote Sensing: Application to the BOREAS region. J. Geophys. Res. 1999, 104, 27719–27734. [Google Scholar] [CrossRef]
- Grigera, G.; Oesterheld, M.; Pacín, F. Monitoring Forage Production for Farmers’ Decision Making. Agric. Syst. 2007, 94, 637–648. [Google Scholar] [CrossRef]
- Running, S.W.; Thornton, P.E.; Nemani, R.; Glassy, J.M. Global Terrestrial Gross and Net Primary Productivity from the Earth Observing System. In Methods in Ecosystem Science; Springer: New York, NY, USA, 2000; pp. 44–57. [Google Scholar]
- Turner, D.; Gower, S.; Cohen, W.; Gregory, M.; Maiersperger, T. Effects of Spatial Variability in Light Use Efficiency on Satellite-based NPP Monitoring. Remote Sens. Environ. 2002, 80, 397–405. [Google Scholar] [CrossRef]
- Lobell, D.B.; Hicke, J.A.; Asner, G.P.; Field, C.B.; Tucker, C.J.; Los, S.O. Satellite Estimates of Productivity and Light Use Efficiency in United States Agriculture, 1982-98. Glob. Chang. Biol. 2002, 8, 722–735. [Google Scholar] [CrossRef]
- Nestola, E.; Calfapietra, C.; Emmerton, C.A.; Wong, C.Y.S.; Thayer, D.R.; Gamon, J.A. Monitoring Grassland Seasonal Carbon Dynamics, by Integrating MODIS NDVI, Proximal Optical Sampling, and Eddy Covariance Measurements. Remote Sens. 2016, 8, 260. [Google Scholar] [CrossRef]
- Hilker, T.; Coops, N.C.; Wulder, M.A.; Black, T.A.; Guy, R.D. The Use of Remote Sensing in Light Use Efficiency Based Models of Gross Primary Production: A Review of Current Status and Future Requirements. Sci. Total Environ. 2008, 404, 411–423. [Google Scholar] [CrossRef]
- Yuan, W.; Liu, S.; Zhou, G.G.; Zhou, G.G.; Tieszen, L.L.; Baldocchi, D.; Bernhofer, C.; Gholz, H.; Goldstein, A.H.; Goulden, M.L.; et al. Deriving a Light Use Efficiency Model from Eddy Covariance Flux Data for Predicting Daily Gross Primary Production Across Biomes. Agric. For. Meteorol. 2007, 143, 189–207. [Google Scholar] [CrossRef]
- Prince, S.D.; Goward, S.N. Global Primary Production: A Remote Sensing Approach. Source J. Biogeogr. 1995, 22, 815–835. [Google Scholar] [CrossRef]
- Fisher, J.I.; Mustard, J.F.; Vadeboncoeur, M.A. Green Leaf Phenology at Landsat Resolution: Scaling from the Field to the Satellite. Remote Sens. Environ. 2006, 100, 265–279. [Google Scholar] [CrossRef]
- Gärtner, P.; Förster, M.; Kleinschmit, B. The Benefit of Synthetically Generated RapidEye and Landsat 8 data Fusion Time Series for Riparian Forest Disturbance Monitoring. Remote Sens. Environ. 2016, 177, 237–247. [Google Scholar] [CrossRef]
- Hunt, E.R.; Everitt, J.H.; Ritchie, J.C.; Moran, M.S.; Booth, D.T.; Anderson, G.L.; Clark, P.E.; Seyfried, M.S. Applications and Research Using Remote Sensing for Rangeland Management. Photogramm. Eng. Remote Sens. 2003, 69, 675–693. [Google Scholar] [CrossRef]
- Walker, J.J.; De Beurs, K.M.; Wynne, R.H. Dryland Vegetation Phenology across An Elevation Gradient in Arizona, USA, Investigated with Fused MODIS and Landsat Data. Remote Sens. Environ. 2014, 144, 85–97. [Google Scholar] [CrossRef]
- Yu, N.; Li, L.; Schmitz, N.; Tian, L.F.; Greenberg, J.A.; Diers, B.W. Development of methods to improve soybean yield estimation and predict plant maturity with an unmanned aerial vehicle based platform. Remote Sens. Environ. 2016, 187, 91–101. [Google Scholar] [CrossRef]
- Guan, S.; Fukami, K.; Matsunaka, H.; Okami, M.; Tanaka, R.; Nakano, H.; Sakai, T.; Nakano, K.; Ohdan, H.; Takahashi, K.; et al. Assessing Correlation of High-Resolution NDVI with Fertilizer Application Level and Yield of Rice and Wheat Crops using Small UAVs. Remote Sens. 2019, 11, 112. [Google Scholar] [CrossRef]
- Yue, J.; Yang, G.; Li, C.; Li, Z.; Wang, Y.; Feng, H.; Xu, B.; Yue, J.; Yang, G.; Li, C.; et al. Estimation of Winter Wheat Above-Ground Biomass Using Unmanned Aerial Vehicle-Based Snapshot Hyperspectral Sensor and Crop Height Improved Models. Remote Sens. 2017, 9, 708. [Google Scholar] [CrossRef]
- Näsi, R.; Viljanen, N.; Kaivosoja, J.; Alhonoja, K.; Hakala, T.; Markelin, L.; Honkavaara, E.; Näsi, R.; Viljanen, N.; Kaivosoja, J.; et al. Estimating Biomass and Nitrogen Amount of Barley and Grass Using UAV and Aircraft Based Spectral and Photogrammetric 3D Features. Remote Sens. 2018, 10, 1082. [Google Scholar] [CrossRef]
- Brocks, S.; Bareth, G.; Brocks, S.; Bareth, G. Estimating Barley Biomass with Crop Surface Models from Oblique RGB Imagery. Remote Sens. 2018, 10, 268. [Google Scholar] [CrossRef]
- Sarron, J.; Malézieux, É.; Sané, C.; Faye, É.; Sarron, J.; Malézieux, É.; Sané, C.A.B.; Faye, É. Mango Yield Mapping at the Orchard Scale Based on Tree Structure and Land Cover Assessed by UAV. Remote Sens. 2018, 10, 1900. [Google Scholar] [CrossRef]
- Planet. Planet Imagery Product Specification: PlanetScope and RapidEye; Planet Labs Inc.: San Francisco, CA, USA, 2018. [Google Scholar]
- Bartolome, J.; Frost, W.; Mcdougald, N. Guidelines for Residual Dry Matter on Coastal and Foothill Rangelands in California. Agric. Nat. Resour. Publ. 2002, 8092, 1–7. [Google Scholar]
- Ball, G.H.; Hall, D.J. ISODATA, A Novel Method of Data Analysis and Pattern Classification; Stanford Research Institute: Menlo Park, CA, USA, 1965. [Google Scholar]
- Pix4D Support Radiometric Corrections. Available online: https://support.pix4d.com/hc/en-us/articles/202559509-Radiometric-corrections#%23label2 (accessed on 26 September 2018).
- Teillet, P.M.; Guindon, B.; Goodenough, D.G. On the Slope-Aspect Correction of Multispectral Scanner Data. Can. J. Remote Sens. 1982, 8, 84–106. [Google Scholar] [CrossRef]
- Meyer, P.; Itten, K.I.; Kellenberger, T.; Sandmeier, S.; Sandmeier, R. Radiometric Corrections of Topographically Induced Effects on Landsat TM Data in an Alpine Environment. ISPRS J. Photogramm. Remote Sens. 1993, 48, 17–28. [Google Scholar] [CrossRef]
- Soenen, S.A.; Peddle, D.R.; Coburn, C.A. SCS+C: A Modified Sun-canopy-sensor Topographic Correction in Forested Terrain. IEEE Trans. Geosci. Remote Sens. 2005, 43, 2148–2159. [Google Scholar] [CrossRef]
- Tan, B.; Masek, J.G.; Wolfe, R.; Gao, F.; Huang, C.; Vermote, E.F.; Sexton, J.O.; Ederer, G. Improved Forest Change Detection with Terrain Illumination Corrected Landsat Images. Remote Sens. Environ. 2013, 136, 469–483. [Google Scholar] [CrossRef]
- Gao, F.; Masek, J.; Schwaller, M.; Hall, F. On the Blending of The landsat and MODIS Surface Reflectance: Predicting Daily Landsat Surface Reflectance. IEEE Trans. Geosci. Remote Sens. 2006, 44, 2207–2218. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Epstein, H.E.; Lauenroth, W.K.; Burke, I.C. ANPP Estimates from NDVI for the Central Grassland Region of the United States ANPP. Ecology 1997, 78, 953–958. [Google Scholar] [CrossRef]
- Anderson, G.; Hanson, J.; Haas, R. Evaluating Landsat Thematic Mapper Derived Vegetation Indices for Estimating Above-Ground Biomass on Semiarid Rangelands. Remote Sens. Environ. 1993, 45, 165–175. [Google Scholar] [CrossRef]
- Moulin, S.; Kergoat, L.; Viovy, N.; Dedieu, G. Global-scale Assessment of Vegetation Phenology using NOAA/AVHRR Satellite Measurements. J. Clim. 1997, 10, 1154–1170. [Google Scholar] [CrossRef]
- Tucker, C.J.; Sellers, P.J. Satellite Remote Sensing of Primary Production. Int. J. Remote Sens. 1986, 7, 1395–1416. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring Vegetation Phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Sellers, P.J.; Los, S.O.; Tucker, C.J.; Justice, C.O.; Dazlich, D.A.; Collatz, G.J.; Randall, D.A. A Revised Land Surface Parameterization (SiB2) Ffor Atmospheric GCMs. Part II: The Generation of Global Fields of Terrestrial Biophysical Parameters from Datellite Data. J. Clim. 1996, 9, 706–737. [Google Scholar] [CrossRef]
- Hart, Q.J.; Brugnach, M.; Temesgen, B.; Rueda, C.; Ustin, S.L.; Frame, K. Daily Reference Evapotranspiration for California using Satellite Imagery and Weather Station Measurement Interpolation. Civ. Eng. Environ. Syst. 2009, 26, 19–33. [Google Scholar] [CrossRef]
- Akitsu, T.; Kume, A.; Hirose, Y.; Ijima, O.; Nasahara, K.N. On the Stability of Radiometric Ratios of Photosynthetically Active Radiation to Global Solar Radiation in Tsukuba, Japan. Agric. For. Meteorol. 2015, 209, 59–68. [Google Scholar] [CrossRef]
- Blackburn, W.J.; Proctor, J.T.A. Estimating Photosynthetically Active Radiation from Measured Solar Irradlance. Sol. Energy 1983, 3183, 233–234. [Google Scholar] [CrossRef]
- Li, R.; Zhao, L.; Ding, Y.; Wang, S.; Ji, G.; Xiao, Y.; Liu, G.; Sun, L. Monthly Ratios of PAR to Global Solar Radiation Measured at Northern Tibetan Plateau, China. Sol. Energy 2010, 84, 964–973. [Google Scholar] [CrossRef]
- Papaioannou, G.; Papanikolaou, N.; Retalis, D. Relationships of Photosynthetically Active Radiation and Shortwave Irradiance. Theor. Appl. Climatol. 1993, 48, 23–27. [Google Scholar] [CrossRef]
- Tsubo, M.; Walker, S. Relationships between Photosynthetically Active Radiation and Clearness Index at Bloemfontein, South Africa. Theor. Appl. Climatol. 2005, 80, 17–25. [Google Scholar] [CrossRef]
- Frouin, R.; Pinker, R.T. Estimating Photosynthetically Active Radiation (PAR) at the Earth’s Surface from Satellite Observations. Remote Sens. Environ. 1995, 51, 98–107. [Google Scholar] [CrossRef]
- ArcGIS for Desktop Area Solar Radiation. Available online: http://desktop.arcgis.com/en/arcmap/10.3/tools/spatial-analyst-toolbox/area-solar-radiation.htm (accessed on 11 March 2019).
- Klute, A.; Cassel, D.K.; Nielsen, D.R. Field Capacity and Available Water Capacity. In Methods of Soil Analysis: Part 1—Physical and Mineralogical Methods; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1986; pp. 901–926. [Google Scholar]
- Gibson, W.; Daly, C.; Kittel, T.; Nychka, D.; Johns, C.; Rosenbloom, N.; McNab, A.; Taylor, G. Development of A 103-year high-resolution Climate Data Set for the Conterminous United States. In AMS Conference on Applied Climatology; American Meteorological Society: Portland, OR, USA, 2002; pp. 181–183. [Google Scholar]
- Robbins, H.; Monro, S. A Stochastic Approximation Method. Annu. Math. Stat. 1951, 22, 400–407. [Google Scholar] [CrossRef]
- Bottou, L. Stochastic gradient descent tricks. In Neural Networks: Tricks of the Trade; Springer: Berlin/Heidelberg, Germany, 2012; pp. 421–436. ISBN 9783642352881. [Google Scholar]
- Trishchenko, A.P.; Cihlar, J.; Li, Z. Effects of Spectral Response Function on Surface Reflectance and NDVI Measured with Moderate Resolution Satellite Sensors. Remote Sens. Environ. 2002, 81, 1–18. [Google Scholar] [CrossRef]
- Roy, D.P.; Kovalskyy, V.; Zhang, H.K.; Vermote, E.F.; Yan, L.; Kumar, S.S.; Egorov, A. Characterization of Landsat-7 to Landsat-8 Reflective Wavelength and Normalized Difference Vegetation Index Continuity. Remote Sens. Environ. 2016, 185, 57–70. [Google Scholar] [CrossRef]
- Li, A.; Bian, J.; Lei, G.; Huang, C. Estimating the Maximal Light Use Efficiency for Different Vegetation Through the CASA Model Combined with Time-series Remote Sensing Data and Ground Measurements. Remote Sens. 2012, 4, 3857–3876. [Google Scholar] [CrossRef]
- Paloscia, S.; Pettinato, S.; Santi, E.; Notarnicola, C.; Pasolli, L.; Reppucci, A. Soil Moisture Mapping using Sentinel-1 Images: Algorithm and Rreliminary Validation. Remote Sens. Environ. 2013, 134, 234–248. [Google Scholar] [CrossRef]
- Wagner, W.; Sabel, D.; Doubkova, M.; Bartsch, A.; Pathe, C. The Potential of Sentinel-1 for Monitoring Soil Moisture With a High Spatial Resolution At Global Scale. Earth Obs. Water Cycle Sci. 2009, 2009, 18–20. [Google Scholar]
- Balenzano, A.; Mattia, F.; Satalino, G.; Pauwels, V.; Snoeij, P. SMOSAR Algorithm for Soil Moisture Retrival using Sentinel-1 Data. In Proceedings of the2012 IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Munich, Germany, 22–27 July 2012; pp. 1200–1203. [Google Scholar]
- Chaplin-Kramer, R.; George, M.R. Effects of Climate Change on Range Forage Production in the San Francisco Bay Area. PLoS ONE 2013, 8, 1–11. [Google Scholar] [CrossRef]












© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Dahlgren, R.A.; Larsen, R.E.; Devine, S.M.; Roche, L.M.; O’ Geen, A.T.; Wong, A.J.Y.; Covello, S.; Jin, Y. Estimating Rangeland Forage Production Using Remote Sensing Data from a Small Unmanned Aerial System (sUAS) and PlanetScope Satellite. Remote Sens. 2019, 11, 595. https://doi.org/10.3390/rs11050595
Liu H, Dahlgren RA, Larsen RE, Devine SM, Roche LM, O’ Geen AT, Wong AJY, Covello S, Jin Y. Estimating Rangeland Forage Production Using Remote Sensing Data from a Small Unmanned Aerial System (sUAS) and PlanetScope Satellite. Remote Sensing. 2019; 11(5):595. https://doi.org/10.3390/rs11050595
Chicago/Turabian StyleLiu, Han, Randy A. Dahlgren, Royce E. Larsen, Scott M. Devine, Leslie M. Roche, Anthony T. O’ Geen, Andy J.Y. Wong, Sarah Covello, and Yufang Jin. 2019. "Estimating Rangeland Forage Production Using Remote Sensing Data from a Small Unmanned Aerial System (sUAS) and PlanetScope Satellite" Remote Sensing 11, no. 5: 595. https://doi.org/10.3390/rs11050595
APA StyleLiu, H., Dahlgren, R. A., Larsen, R. E., Devine, S. M., Roche, L. M., O’ Geen, A. T., Wong, A. J. Y., Covello, S., & Jin, Y. (2019). Estimating Rangeland Forage Production Using Remote Sensing Data from a Small Unmanned Aerial System (sUAS) and PlanetScope Satellite. Remote Sensing, 11(5), 595. https://doi.org/10.3390/rs11050595