
Influence of Organic Farming on the Potato Transcriptome


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Farming Site
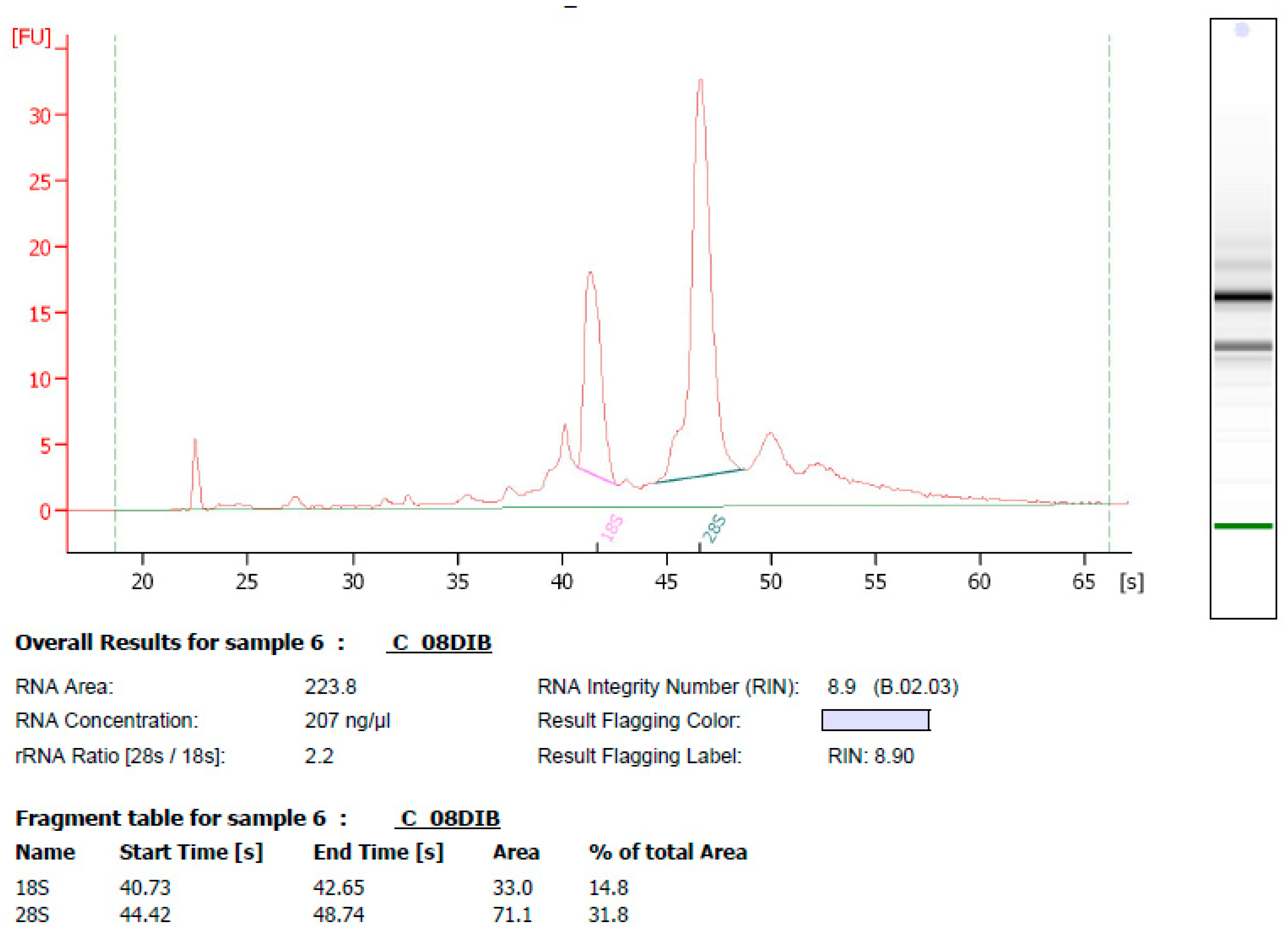
2.2. Total RNA Isolation from Potato Tubers and Fluorescent Labelling of cRNA
2.3. Microarray Data Mining
2.4. Quantitative Real-Time PCR Analysis
3. Results
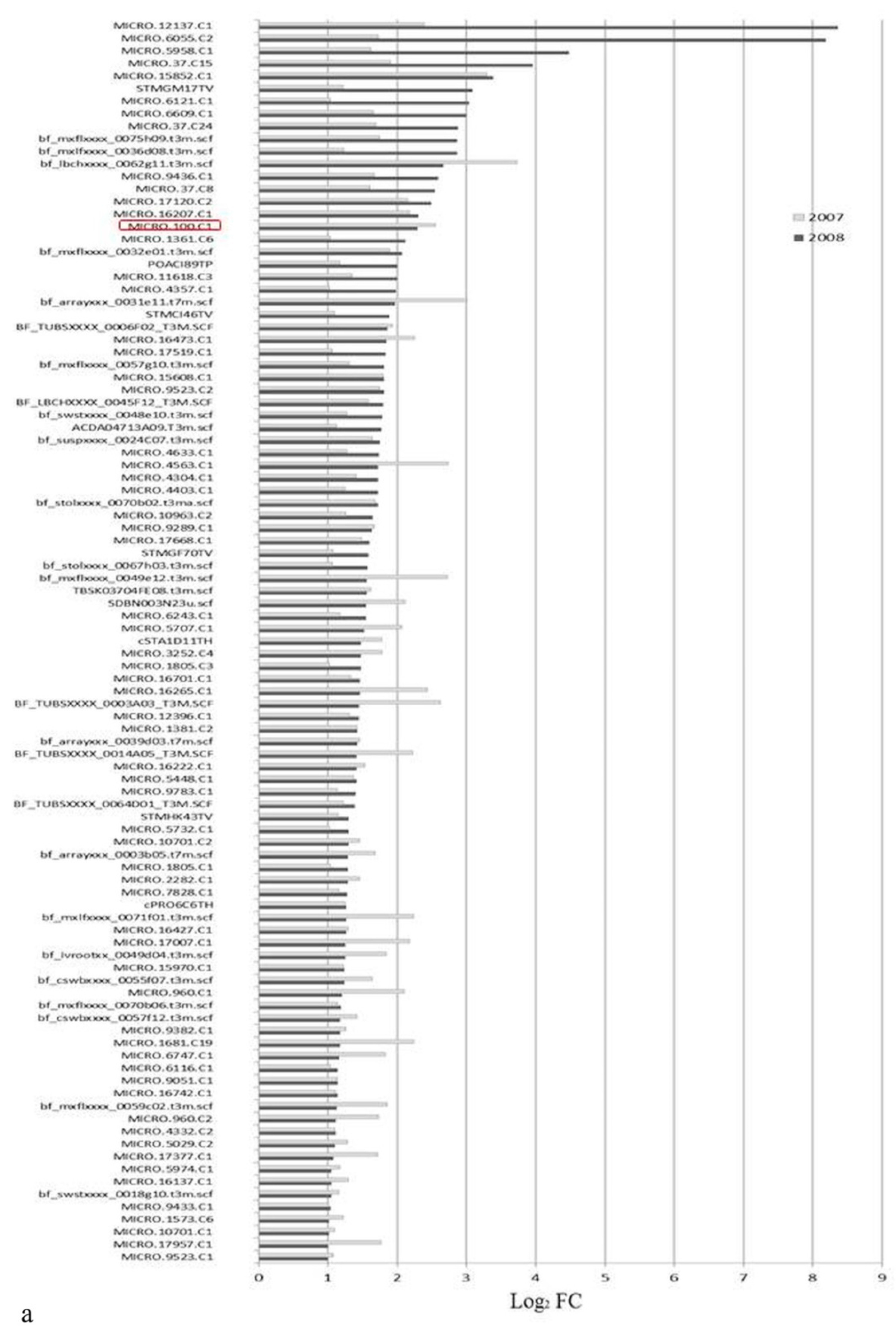
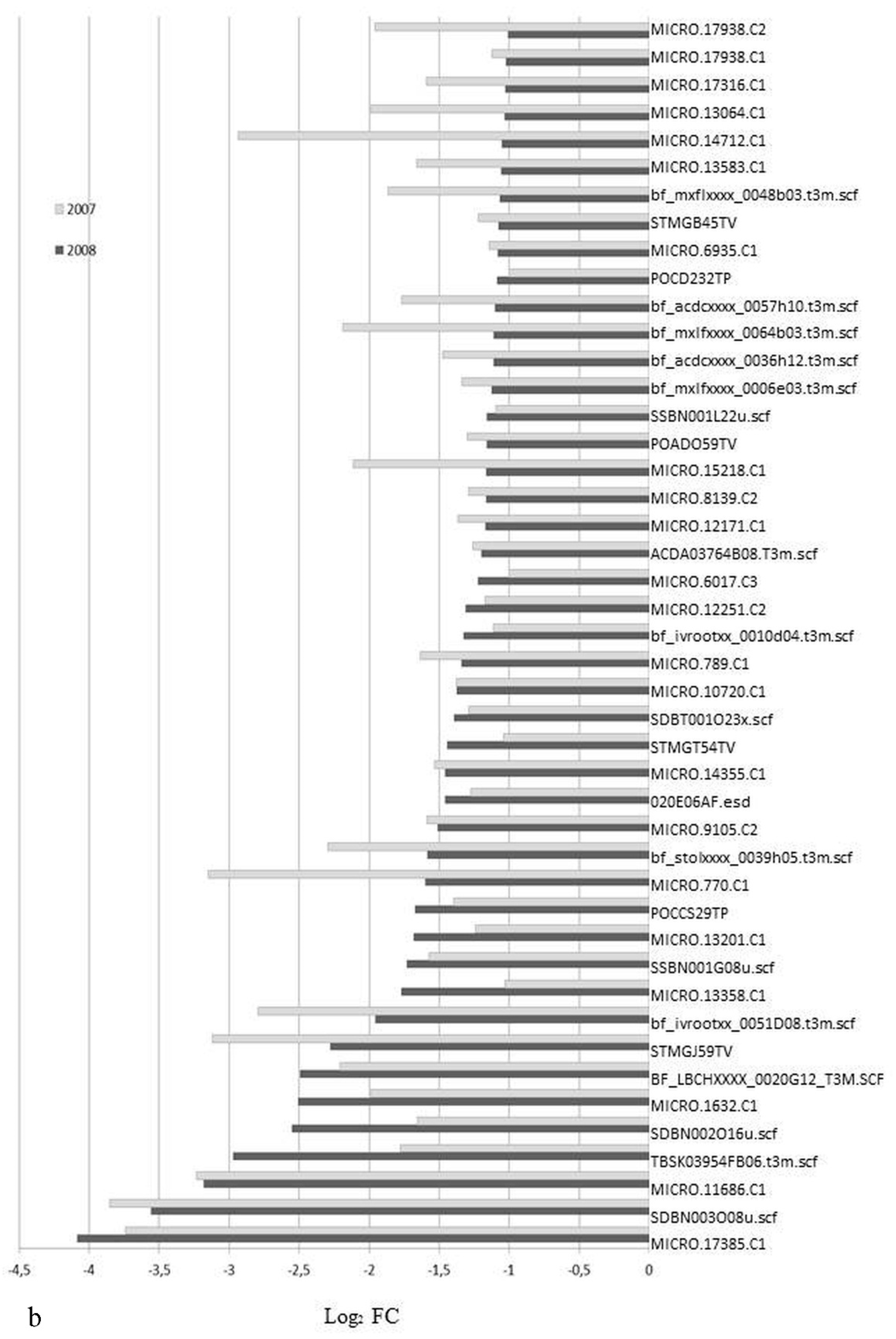
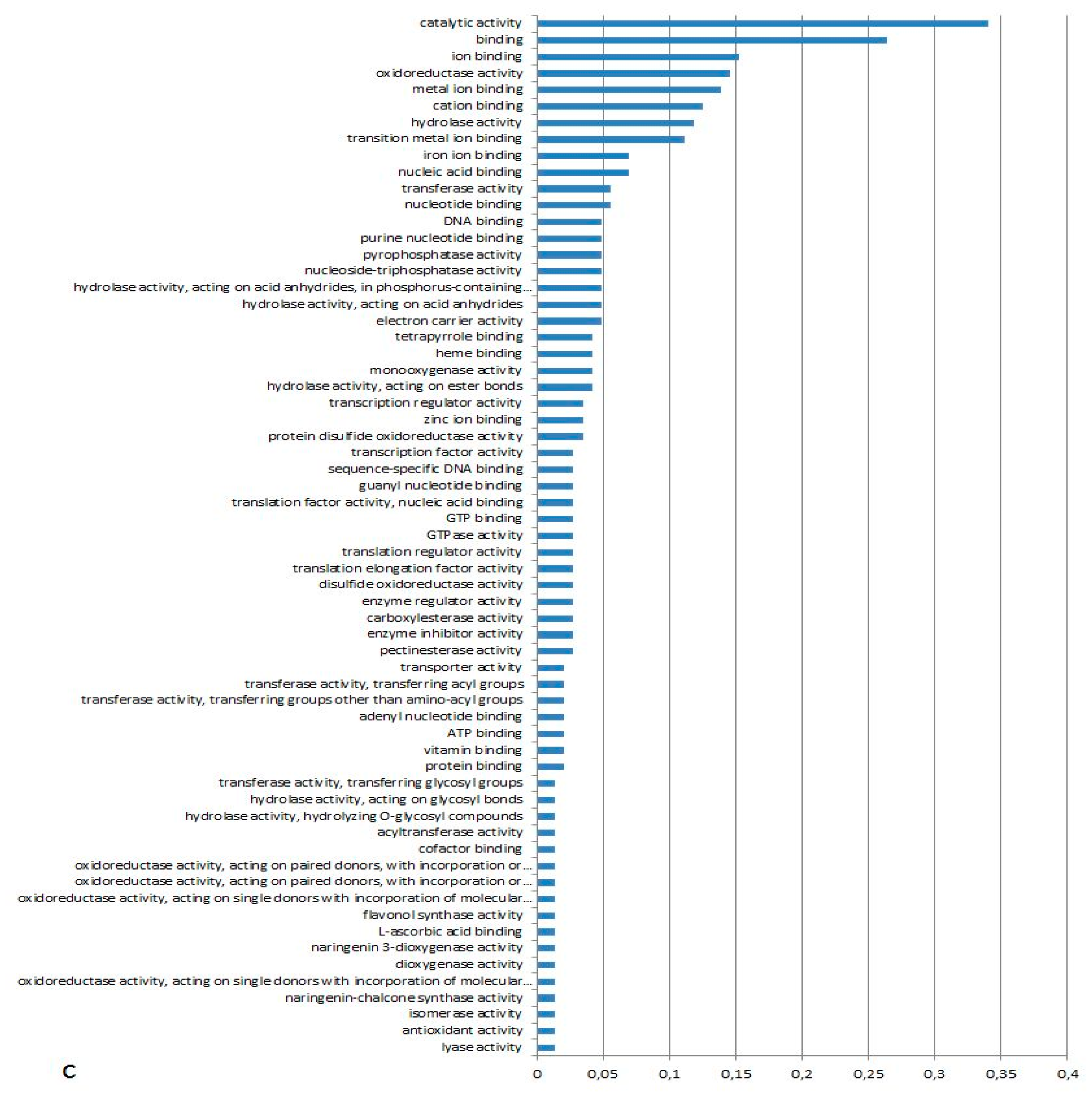
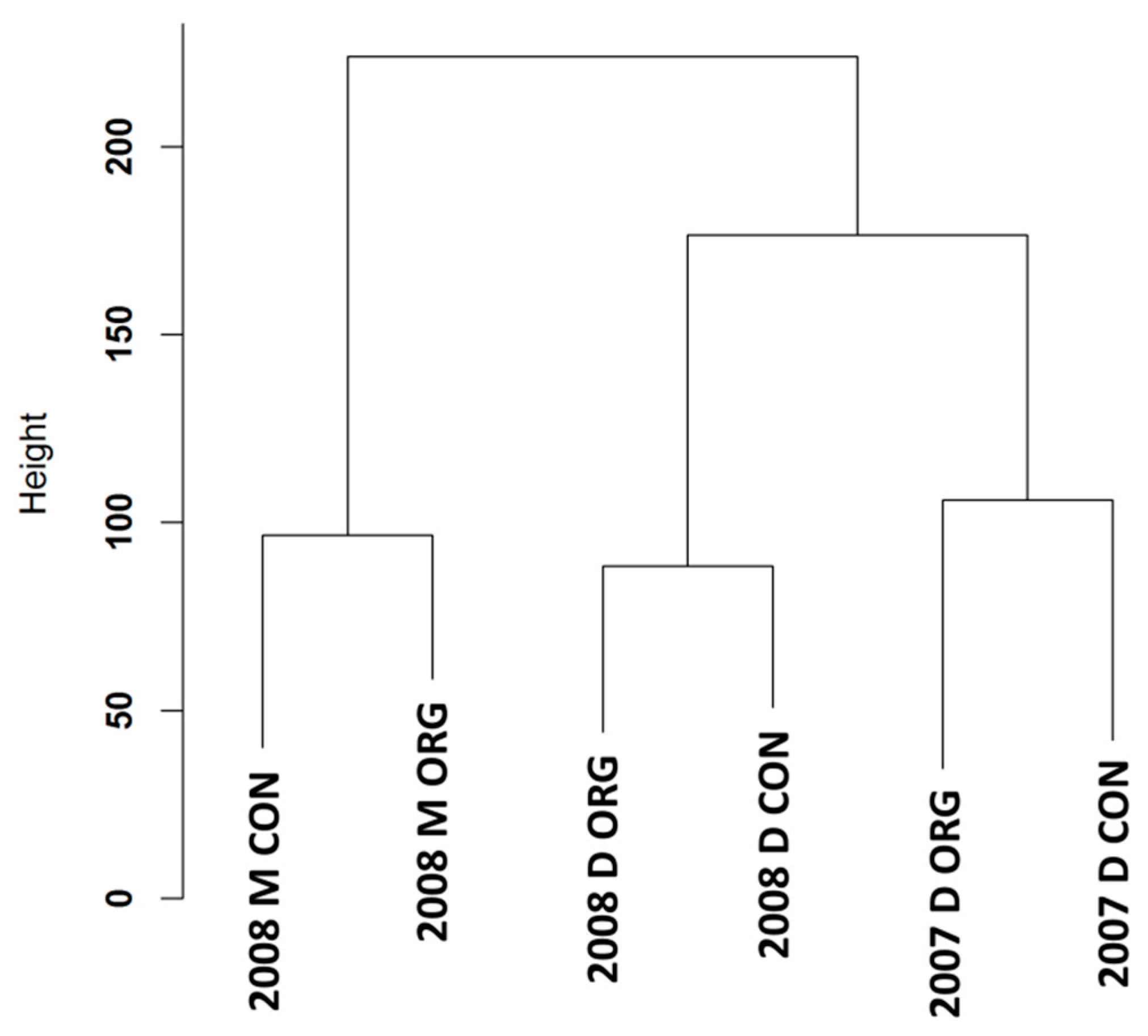
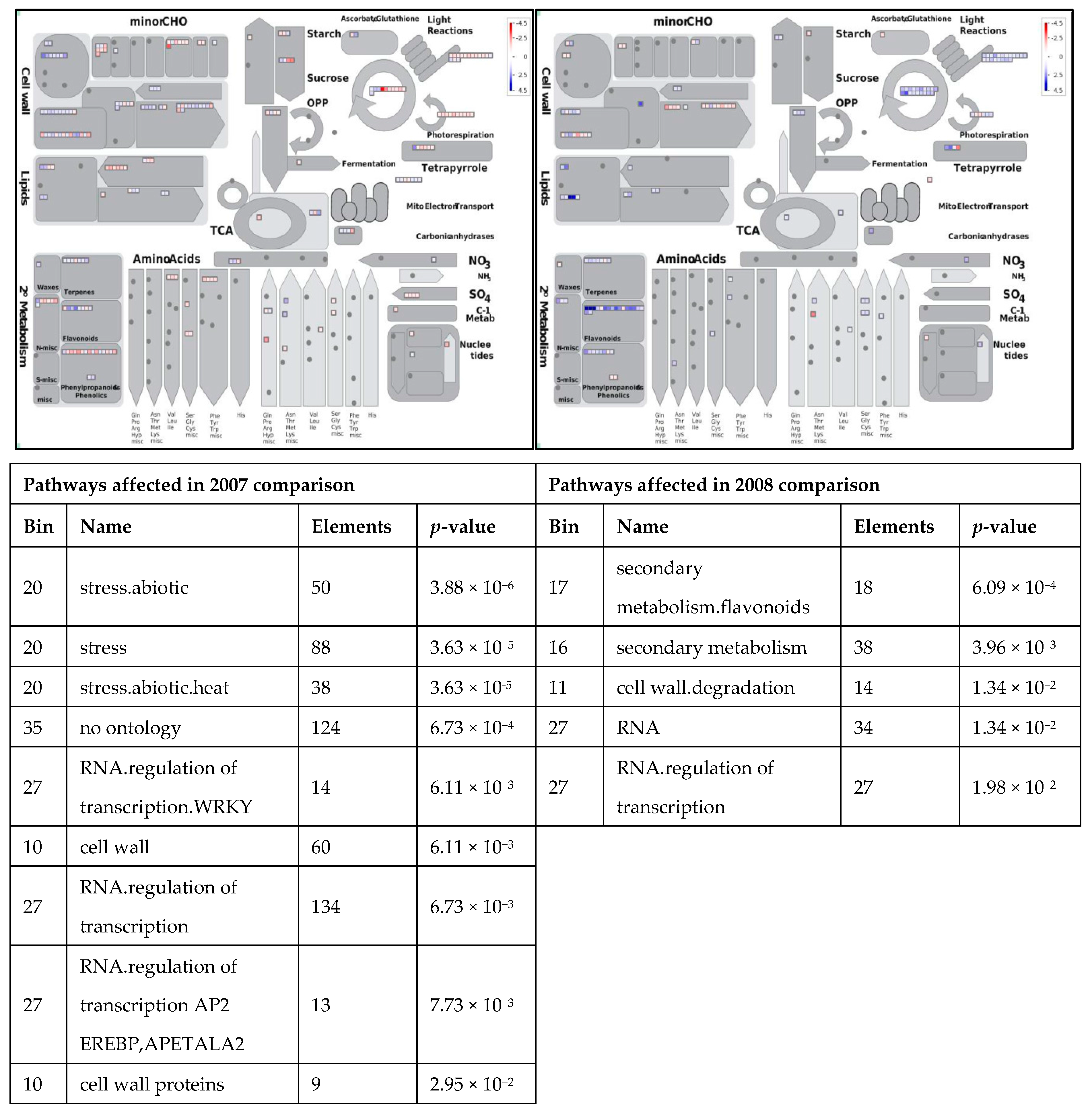
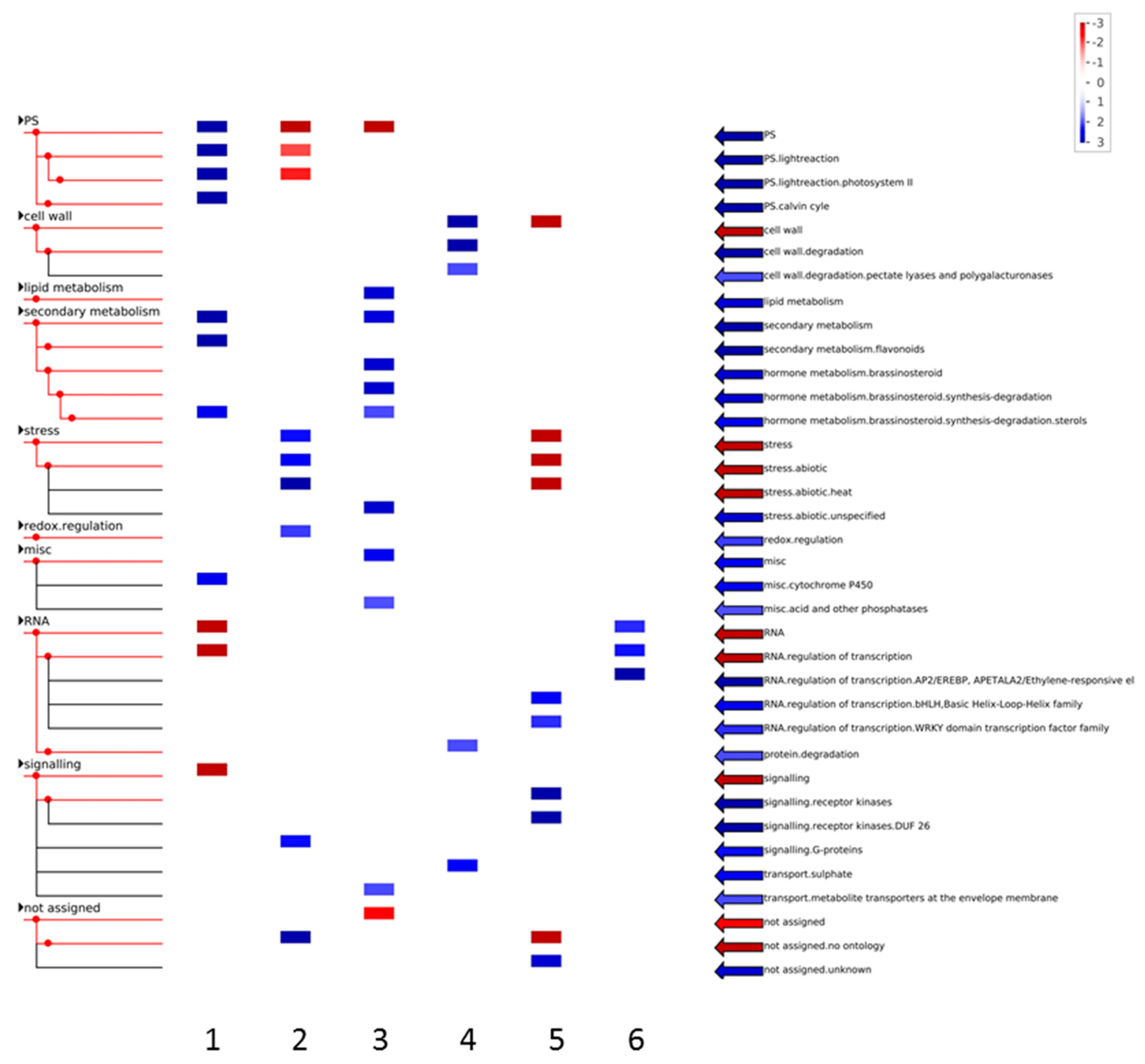
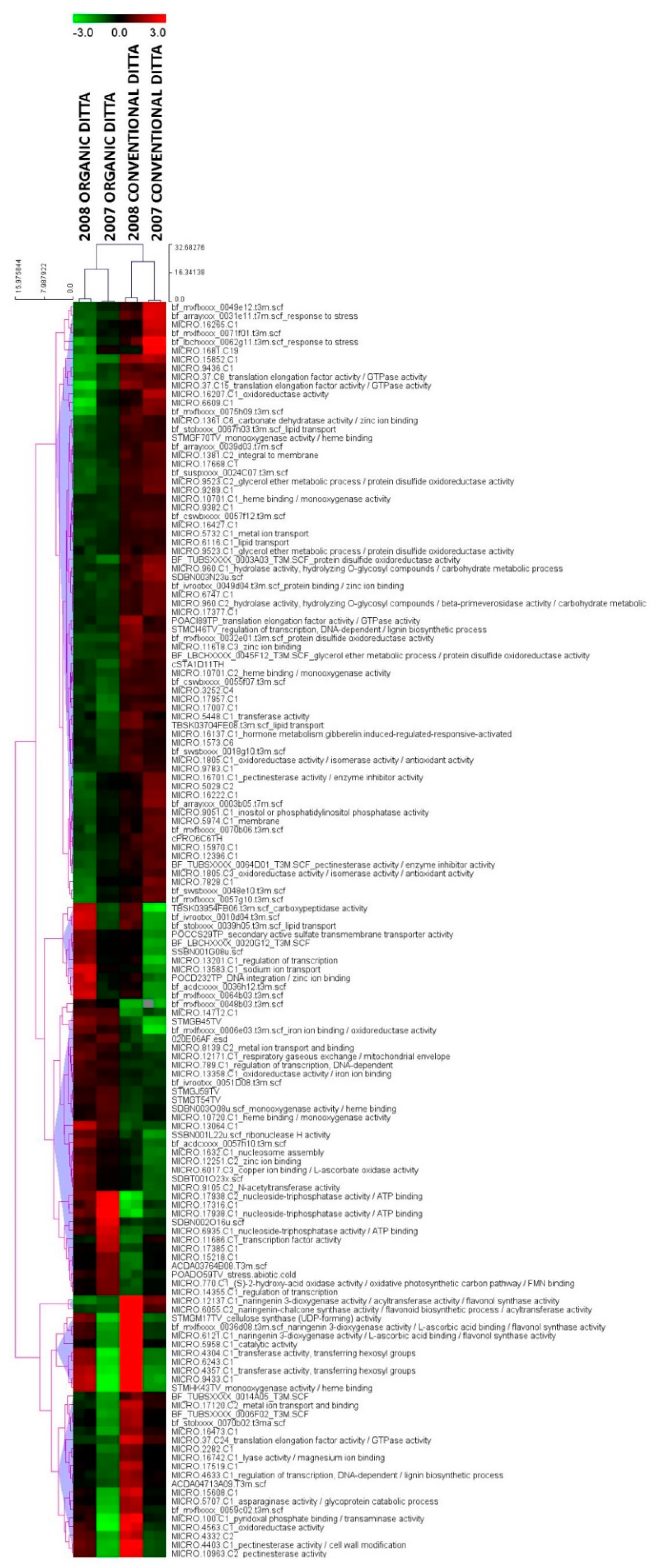
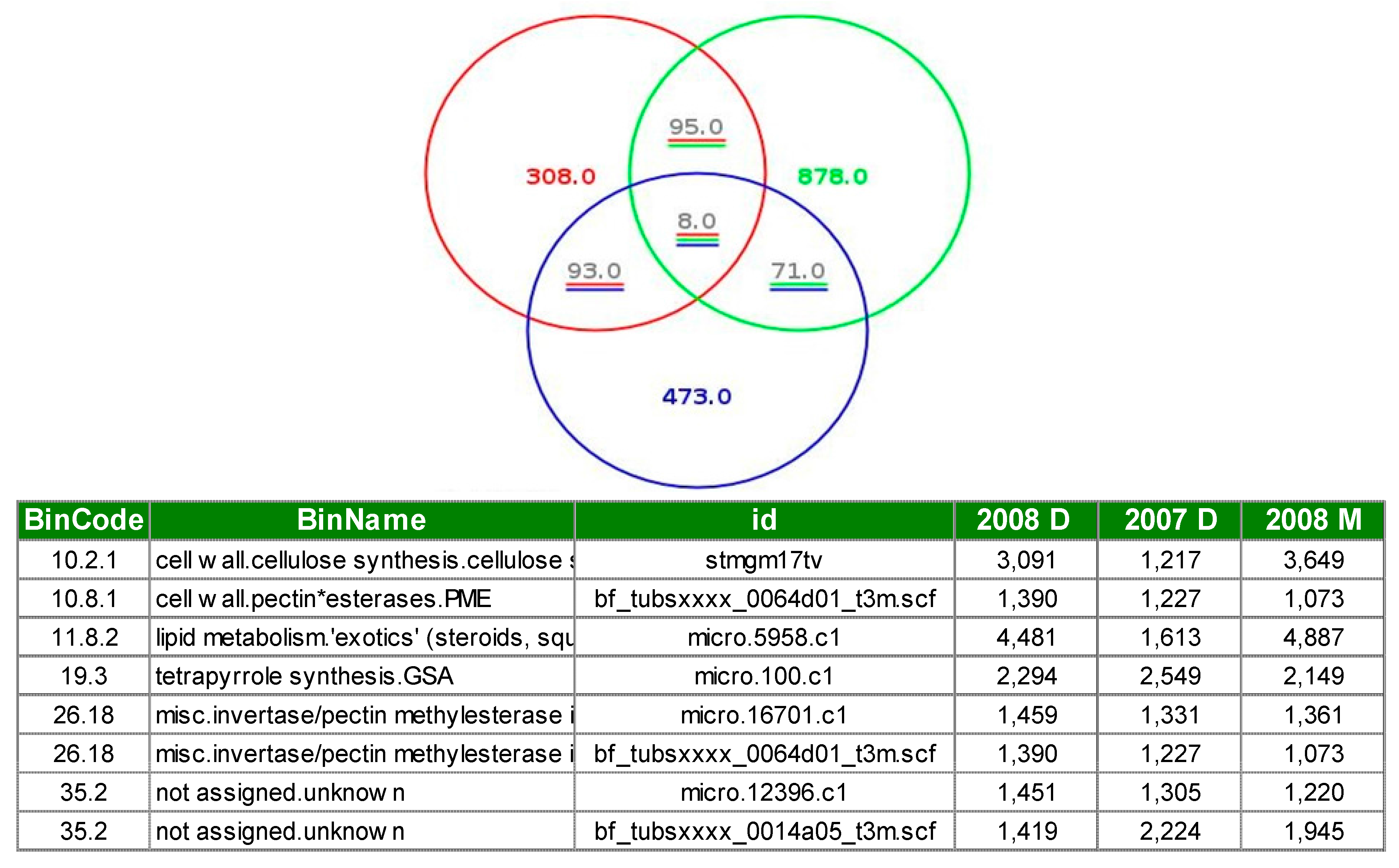
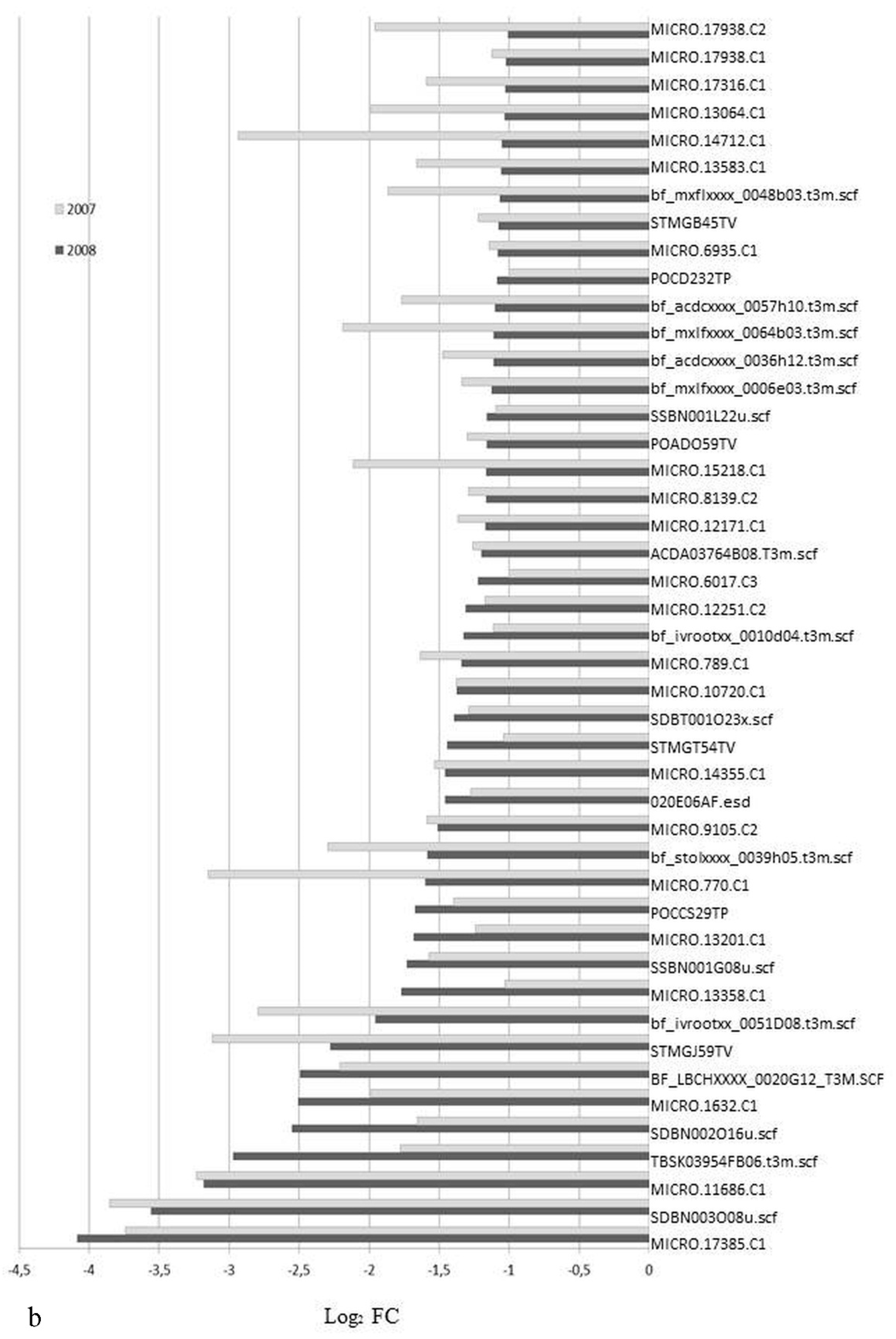
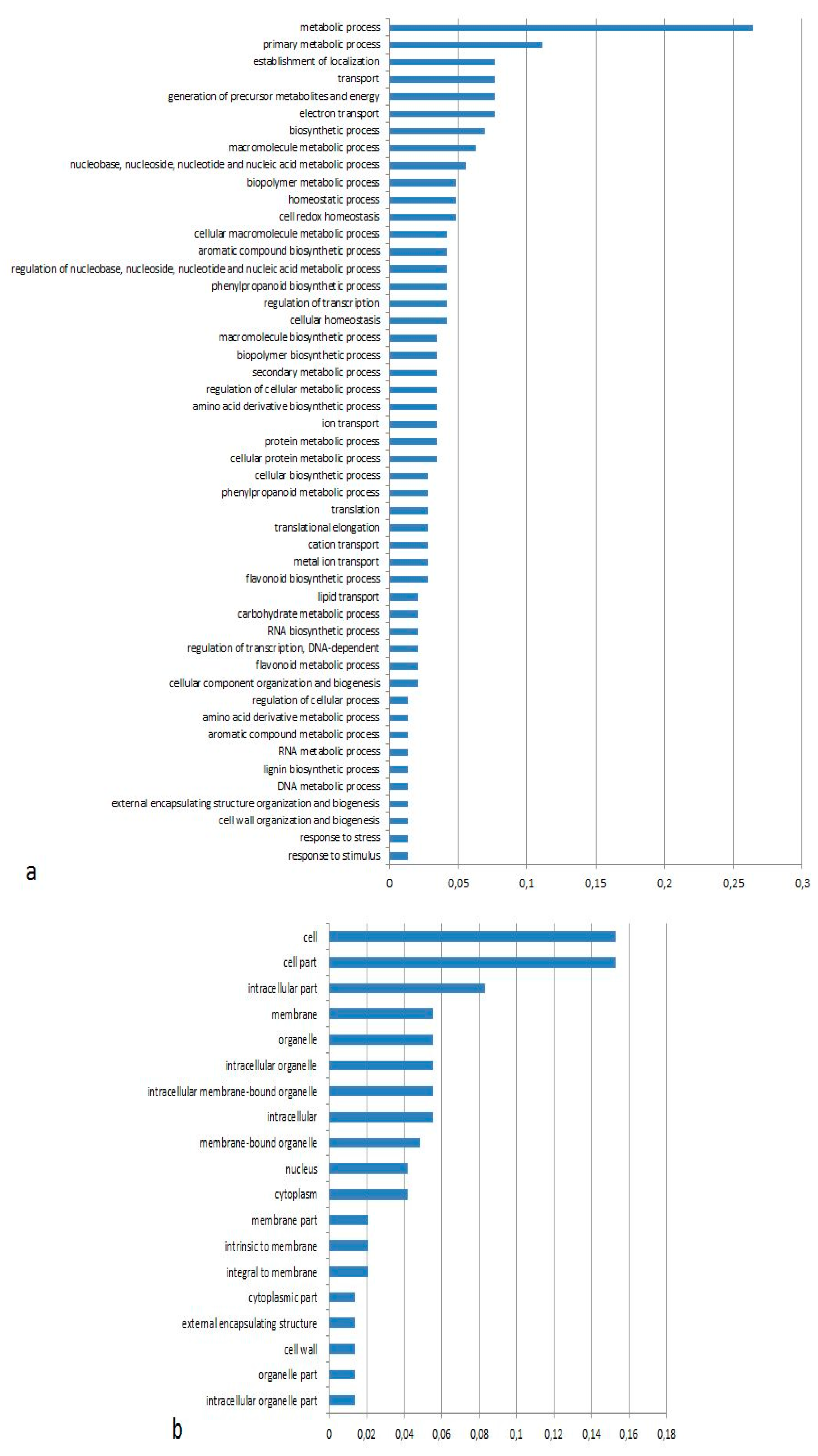
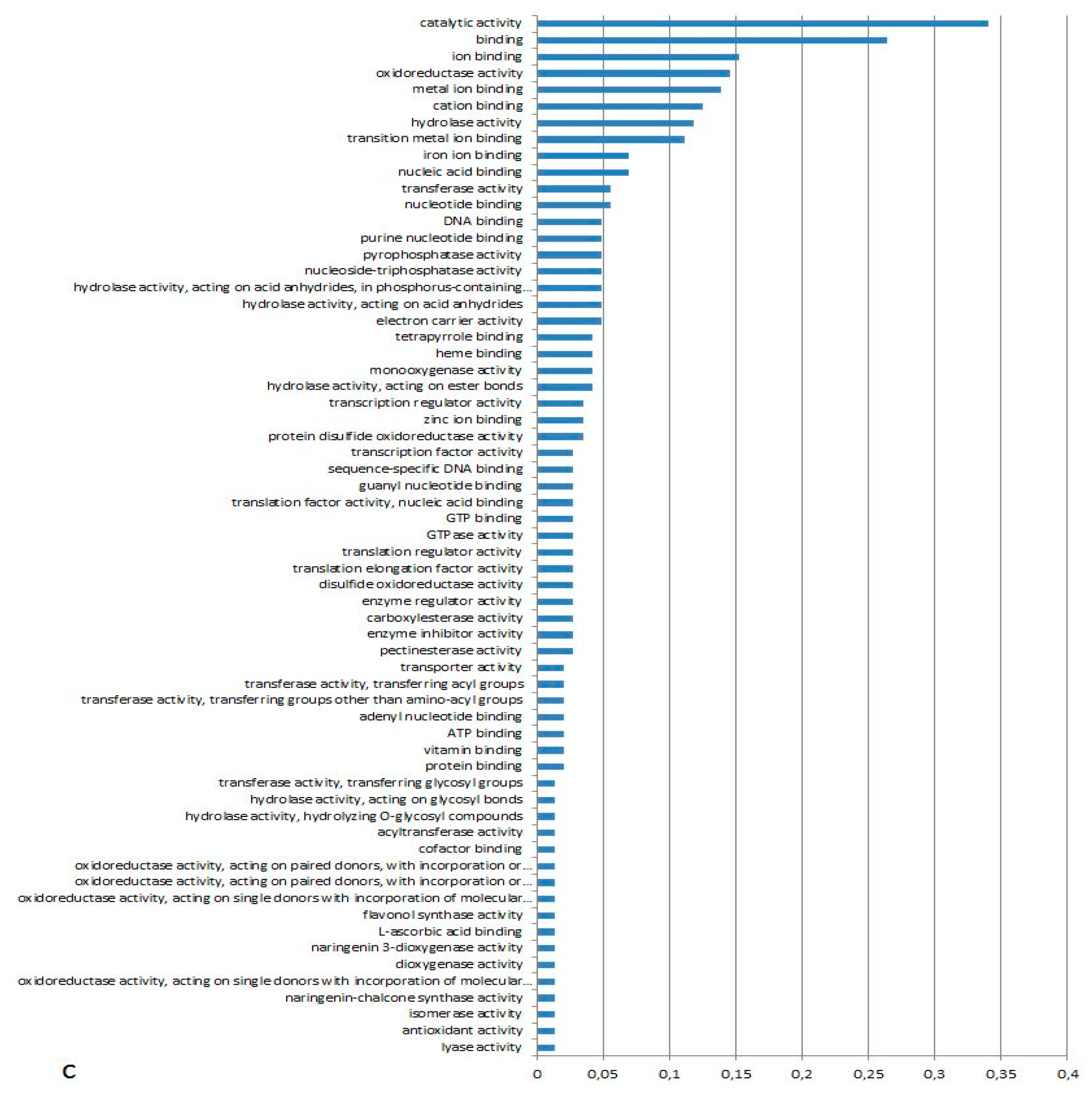
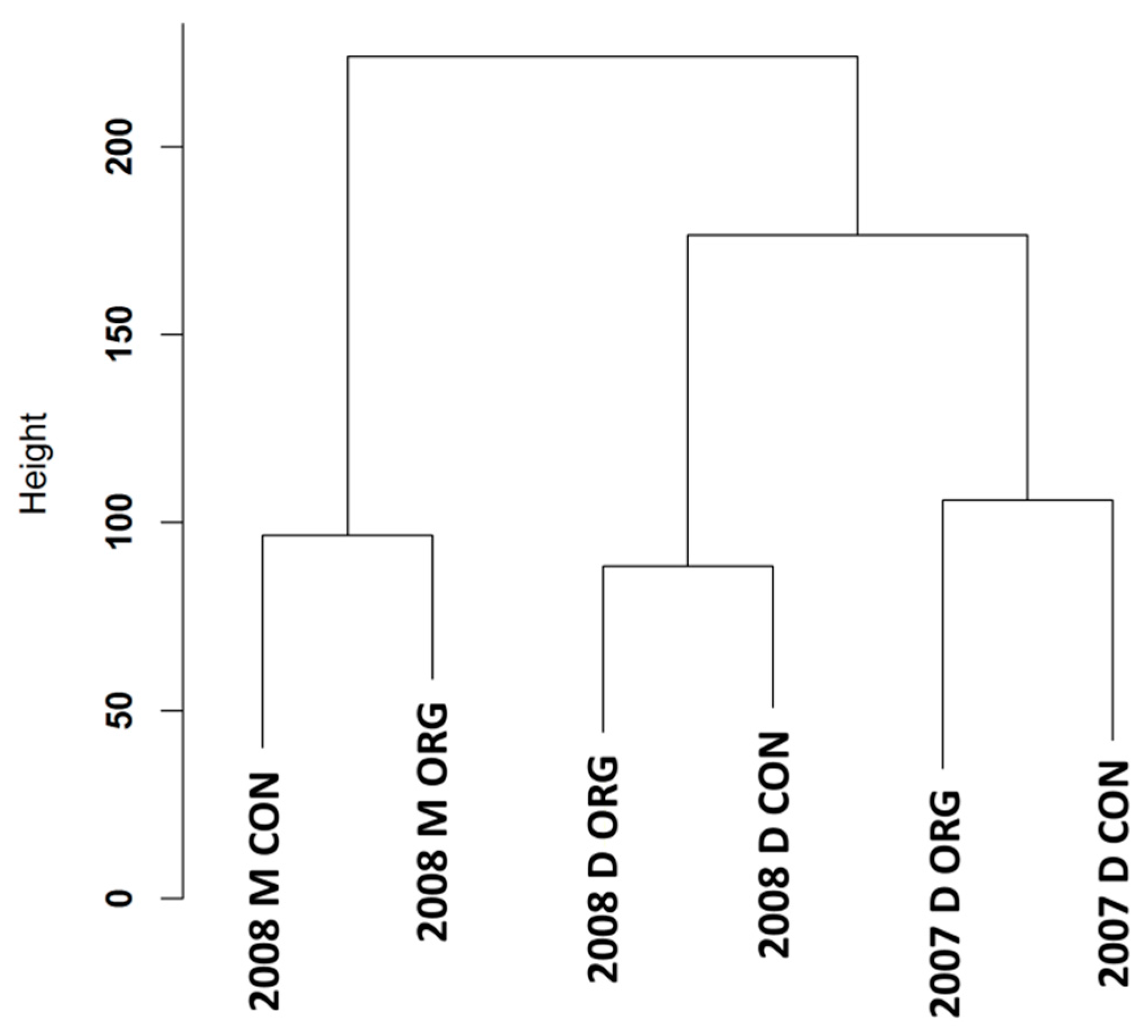
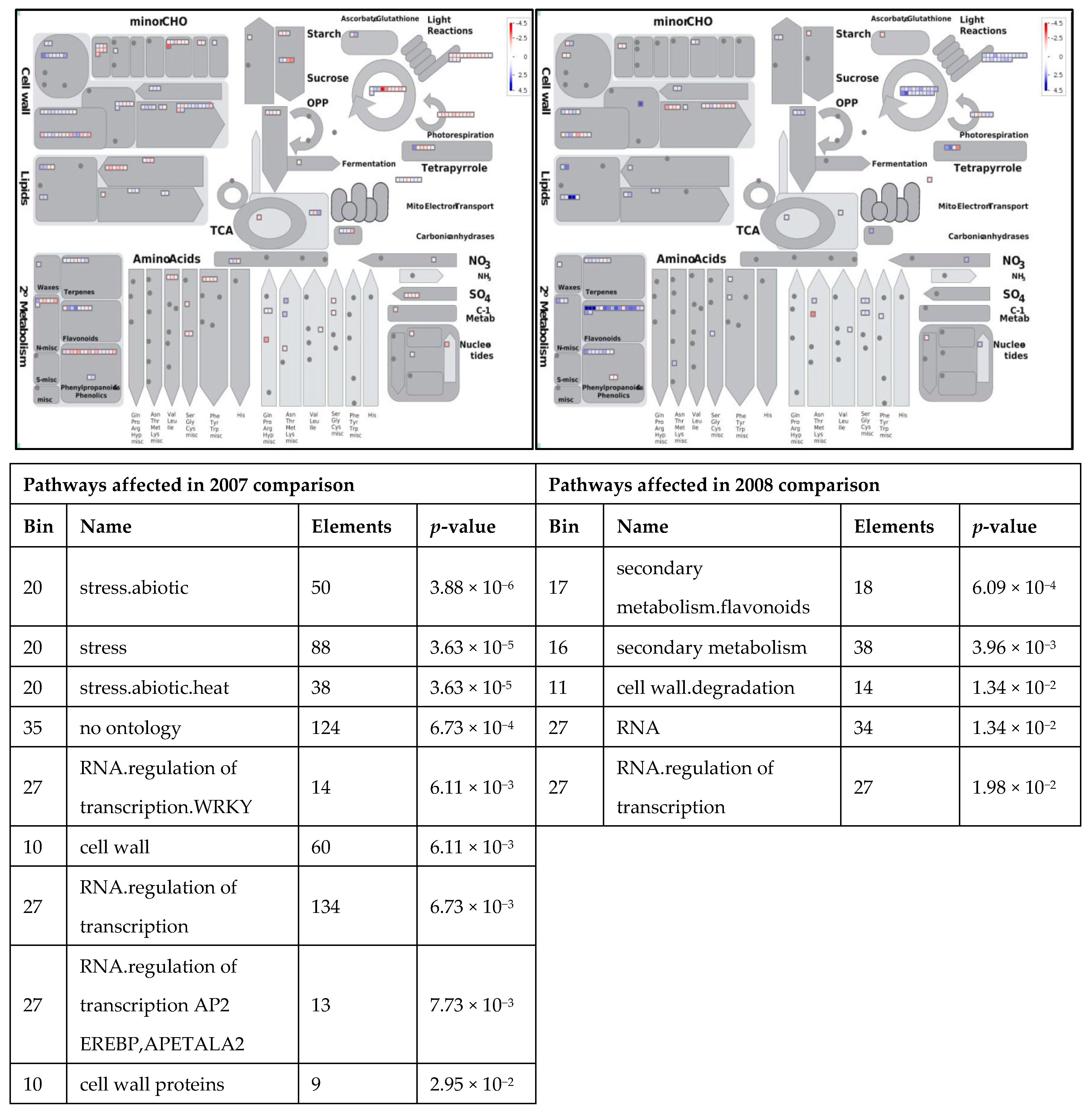
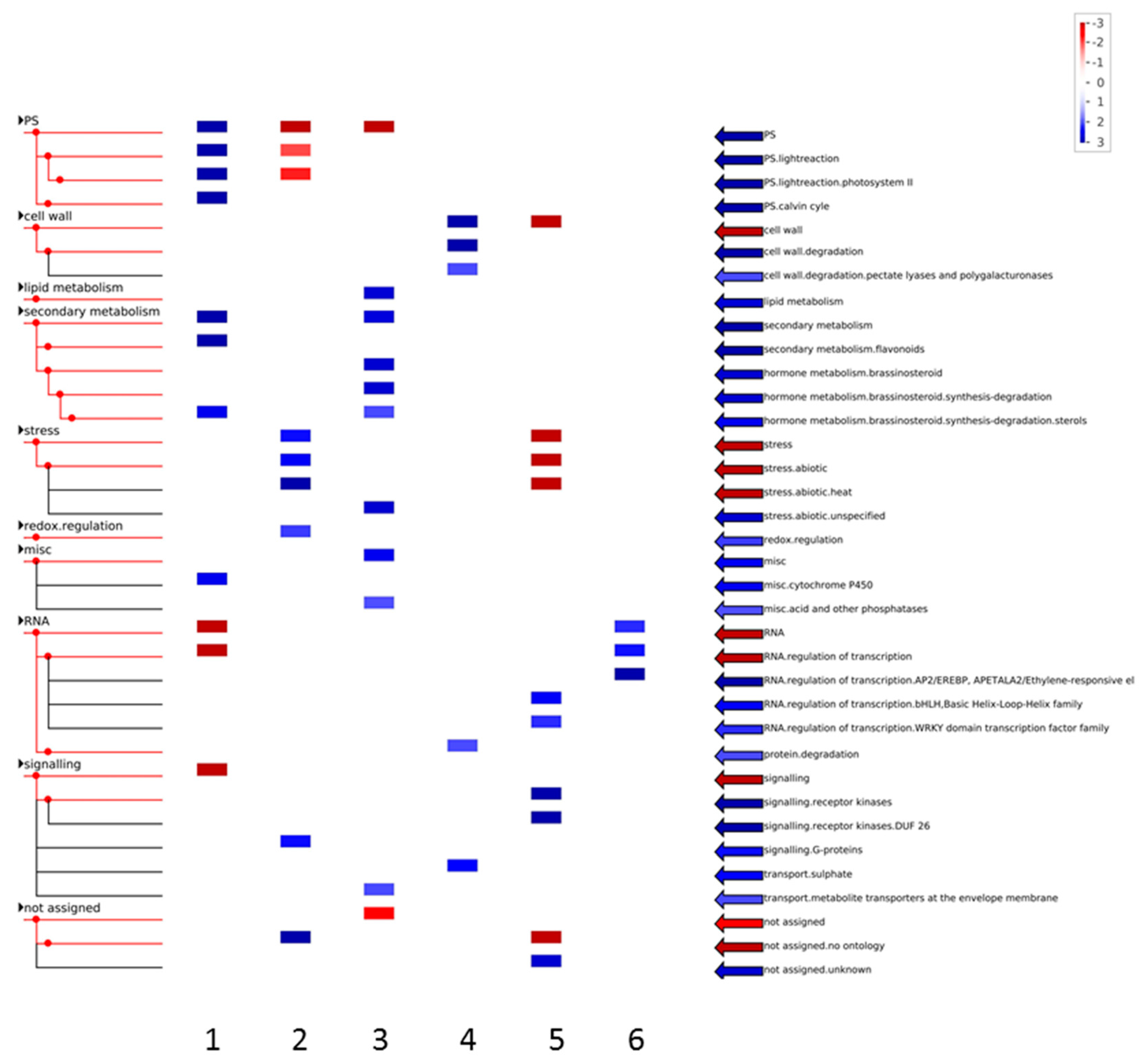
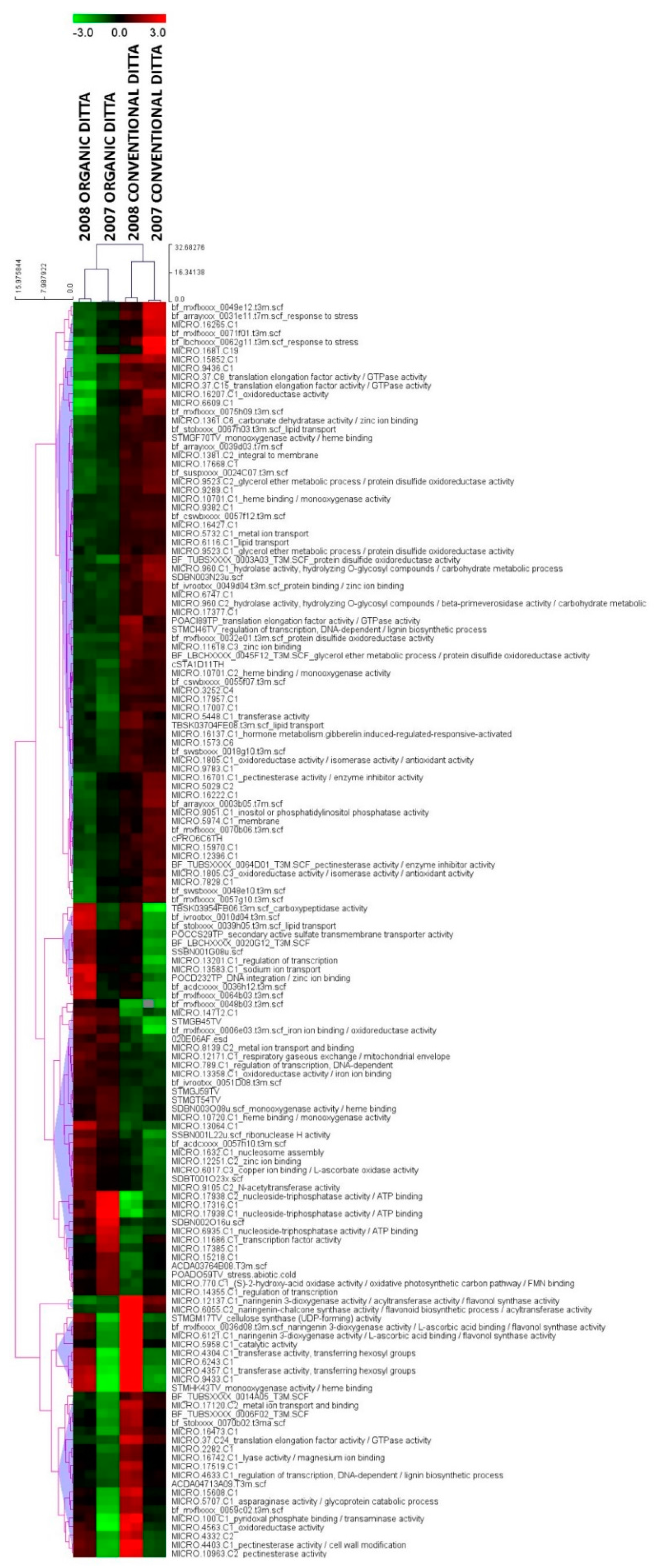
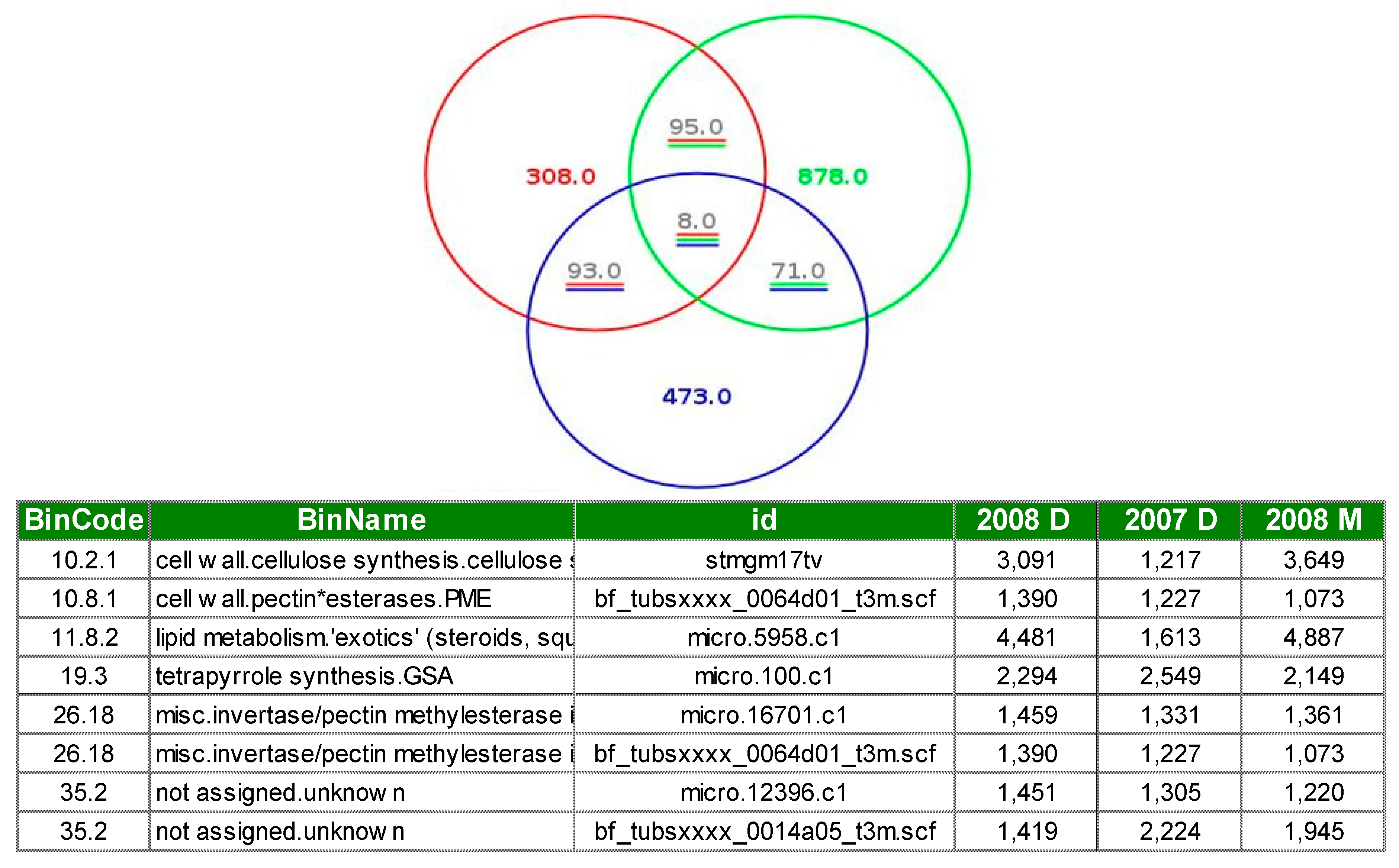
3.1. Expression Profiling and Functional Classification of Differentially Expressed Features Modulated in cv. Ditta over Two Years
3.2. Expression Profiling and Functional Classification of Differentially Expressed Features Modulated in Clone Ditta and Clone MN1404 O5
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTAB | PVP40 | EDTA | TrisHCL | NaCl |
|---|---|---|---|---|
| 20 g L−1 | 20 g L−1 | 0.025 mol L−1 | 0.1 mol L−1 | 2 mol L−1 |
| Probes | GenBank | PGSC ID | Sequenze Primer (5′–3′) | Size (pb) | Cluster | ||
|---|---|---|---|---|---|---|---|
| FW | REV | Expected Ampicon | Tanscript | ||||
| bf_mxlfxxxx_0036d08.t3m | gb|CAA63092 | >PGSC0003DMT400036565 | TCCACATTGGTGACCAAGTT | ATGAGGGCTCCAAGAAAACA | 120 | 1460 | E |
| MICRO.6055.C2 | gb|U47738.1 | >PGSC0003DMT400049165 | GTTTGCTCAGAGATCACTGCA | ATCACCAAAAAGGGCTTGTC | 87 | 1442 | D |
| MICRO 5707.C1 | gb|AAM23265 | >PGSC0003DMT400020647 | CACCGTGAAAAACCCAATCT | GCCTCTTTGGCTAGTTTCAGC | 174 | 1214 | F |
| MICRO.100.C1 | XM_006351918 | >PGSC0003DMT400062385 | CCACTAGAATGGGGTGTTGG | TGAGACTTGAGTTCTTCCACTCT | 194 | 1234 | F |
| MICRO .6017.C3 | gb|AAY47050 | >PGSC0003DMT400071763 | GGCAGCTTGACTGCTCTTTC | TTACCACAAATGGCTCCACA | 100 | 1118 | C |
| MICRO.17316.C1 | XM_006343852 | >PGSC0003DMT400031731 | CGATTCAGGGATGAACAGGA | GAAAACAAAACGCGTCACAA | 138 | 3572 | C |
| MICRO.17938.C1 | XM_006343852 | >PGSC0003DMT400031731 | ACCGAGTCTTGCTGACCTGT | AAGAAGTACAAGCGGTTGGC | 237 | 3572 | C |
| MICRO.17938.C2 | XM_006343852 | >PGSC0003DMT400031731 | TTCATGACTCATTGGGATGG | CCGTCATCAACTCTCTCCTTG | 197 | 3572 | C |
| MICRO.13064.C2 | XR368500 | >PGSC0003DMT400039114 | CGATCAACTCAACCCCAAG | GTCCCGGGAACATCAACTC | 150 | 487 | C |
| Elongation factor alpha 1 | AJ536671.1 | ------- | ATTGGAAATGGATATGCTCC | TCCTTACCTGAACGCCTGTCA | 102 | 1753 | - |
References
- FAO Statistical Pocketbook 2015. Available online: http://www.fao.org/3/a-i4691e.pdf (accessed on 4 April 2017).
- Organic Food Sales Growth in the United States from 2000 to 2015. Available online: https://www.statista.com/statistics/196962/organic-food-sales-growth-in-the-us-since-2000/ (accessed on 4 April 2017).
- Organic in Europe. Prospects and Developments 2016. IFOAM EU Group. Available online: https://shop.fibl.org/fileadmin/documents/shop/1708-organic-europe-2016.pdf (accessed on 8 May 2017).
- Pacifico, D.; Paris, R. Effect of Organic Potato Farming on Human and Environmental Health and Benefits from New Plant Breeding Techniques. Is It Only a Matter of Public Acceptance? Sustainability 2016, 8, 1054. [Google Scholar] [CrossRef]
- Lu, C.; Hawkesford, M.J.; Barraclough, P.B.; Poulton, P.R.; Wilson, I.D.; Barker, G.L.; Edwards, K.J. Markedly different gene expression in wheat grown with organic or inorganic fertilizer. Proc. Biol. Sci. 2005, 272, 1901–1908. [Google Scholar] [CrossRef] [PubMed]
- Maggio, A.; Carillo, P.; Bulmetti, G.S.; Fuggi, A.; Barbieri, G.; De Pascale, S. Potato yield and metabolic profiling under conventional and organic farming. Eur. J. Agron. 2008, 28, 343–350. [Google Scholar] [CrossRef]
- Hoefkens, C.; Verbeke, W.; Aertsens, J.; Mondelaers, K.; Camp, J.V. The nutritional and toxicological value of organic vegetables. Br. Food J. 2009, 111, 1062–1077. [Google Scholar] [CrossRef]
- Lombardo, S.; Lo Monaco, A.; Pandino, G.; Parisi, B.; Mauromicale, G. The phenology, yield and tuber composition of ‘early’ crop potatoes: A comparison between organic and conventional cultivation systems. Renew. Agric. Food Syst. 2013, 28, 50–58. [Google Scholar] [CrossRef]
- Lehesranta, S.J.; Koistinen, K.M.; Massat, N.; Davies, H.V.; Shepherd, L.V.T.; McNicol, J.W.; Cakmak, I.; Cooper, J.; Lück, L.; Kärenlampi, S.O. Effects of agricultural production systems and their components on protein profiles of potato tubers. Proteomics 2007, 7, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, D.; Casciani, L.; Ritota, M.; Mandolino, G.; Onofri, C.; Moschella, A.; Parisi, B.; Cafiero, C.; Valentini, M. NMR-based metabolomics for organic farming traceability of early potatoes. J. Agric. Food Chem. 2013, 61, 11201–11211. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, J.P.; Cankar, K.; Hendriksen, P.J.; Beenen, H.G.; Zhu, M.; Scheffer, S.; Shepherd, L.V.; Stewart, D.; Davies, H.V.; Leifert, C. The identification and interpretation of differences in the transcriptomes of organically and conventionally grown potato tubers. J. Agric. Food Chem. 2012, 60, 2090–2101. [Google Scholar] [CrossRef] [PubMed]
- Siddappa, S.; Tiwari, J.K.; Sindhu, R.; Sharma, S.; Bhardwaj, V.; Chakrabarti, S.K.; Singh, B.P. Phytophthora infestans associated global gene expression profile in a late blight resistant Indian potato cv. Aust. J. Crop Sci. 2014, 8, 215–222. [Google Scholar]
- Kaminski, K.P.; Petersen, A.H.; Sønderkær, M.; Pedersen, L.H.; Pedersen, H.; Feder, C.; Nielsen, K.L. Transcriptome Analysis Suggests That Starch Synthesis May Proceed via Multiple Metabolic Routes in High Yielding Potato Cultivars. PLoS ONE 2012, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kloosterman, B.; De Koeyer, D.; Griffiths, R.; Flinn, B.; Steuernagel, B.; Scholz, U.; Sonnewald, S.; Sonnewald, U.; Bryan, G.J.; Prat, S. Genes driving potato tuber initiation and growth: Identification based on transcriptional changes using the POCI array. Funct. Integr. Genom. 2008, 8, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Stushnoff, C.; Ducreux, L.J.; Hancock, R.D.; Hedley, P.E.; Holm, D.G.; McDougall, G.J.; McNicol, J.W.; Morris, J.; Morris, W.L.; Sungurtas, J.A. Flavonoid profiling and transcriptome analysis reveals new gene-metabolite correlations in tubers of Solanum tuberosum L. J. Exp. Bot. 2010, 61, 1225–1238. [Google Scholar] [CrossRef] [PubMed]
- Onofri, C.; Pacifico, D.; Ostano, P.; Mello-Grand, M.; Ghimenti, C.; Parisi, B.; Mandolino, G. Strumenti di genomica funzionale per la tipizzazione dell’origine di coltivazione della patata. Minerva Biotecnol. 2012, 24, 11–23. [Google Scholar]
- Ducreux, J.M.L.; Morris, W.L.; Prosser, I.M.; Morris, J.A.; Beale, M.H.; Wright, F.; Shepherd, T.; Bryan, G.J.; Hedley, P.E.; Taylor, M.A. Expression profiling of potato germplasm differentiated in quality traits leads to the identification of candidate flavour and texture genes. J. Exp. Bot. 2008, 59, 4219–4231. [Google Scholar] [CrossRef] [PubMed]
- Perkins, J.R.; Martins, A.A.; Calvo, M.; Grist, J.; Rust, W.; Schmid, R.; Hildebrandt, T.; Kohl, M.; Orengo, C.; McMahon, S.B.; et al. A comparison of RNA-seq and exon arrays for whole genome transcription profiling of the L5 spinal nerve transection model of neuropathic pain in the rat. Mol. Pain 2014, 10, 2–26. [Google Scholar] [CrossRef] [PubMed]
- Bioconductor. Available online: http://www.bioconductor.org (accessed on 4 April 2017).
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S. Bioconductor: Open software development for computational biology and bioinformatics. Genome Biol. 2004, 5, 80. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.I.; Bhagabati, N.K.; Braisted, J.C.; Liang, W.; Sharov, V.; Howe, E.A.; Li, J.; Thiagarajan, M.; White, J.A.; Quackenbush, J. TM4 microarray software suite. Methods Enzymol. 2006, 411, 134–139. [Google Scholar] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar]
- POCI 015425 Annotation v1. Available online: http://pgrc-35.ipk-gatersleben.de (accessed on 8 May 2017).
- Spud DB. Potato Genomics Resources. Available online: http://solanaceae.plantbiology.msu.edu/integrated_searches.shtml (accessed on 8 May 2017).
- Radonic, A.; Thulke, S.; Nackay, I.M.; Landt, O.; Siegert, W.; Nitsche, A. Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2004, 313, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Magill, R.; Frederiksen, R.; Magill, C. Chalcone synthase and phenylalanine ammonia-lyase mRNA levels following exposure of sorghum seedlings to three fungal pathogens. Phys. Mol. Plant Pathol. 1996, 49, 187–199. [Google Scholar] [CrossRef]
- Helliwell, C.A.; Poole, A.; Peacock, W.J.; Dennis, E.S. Arabidopsis ent-Kaurene Oxidase Catalyzes Three Steps of Gibberellin Biosynthesis. Plant Physiol. 1999, 119, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.; Leifert, C.; Sanderson, R.; Seal, C.J. Agroecosystem management and nutritional quality of plant food: The case of organic fruits and vegetables. Crit. Rev. Plant Sci. 2011, 30, 177–197. [Google Scholar] [CrossRef]
- Merkl, R.; Hrádková, I.; Filip, V.; Šmidrkal, J. Antimicrobial and antioxidant properties of phenolic acids alkyl esters. Czech J. Food Sci. 2010, 28, 275–279. [Google Scholar]
- Skrabule, I.; Muceniece, R.; Kirhnere, I. Evaluation of Vitamins and Glycoalkaloids in Potato Genotypes Grown Under Organic and Conventional Farming Systems. Potato Res. 2013, 56, 259–276. [Google Scholar] [CrossRef]
- Hendriksen, H.V.; Kornbrust, B.A.; Oestergaard, P.R.; Stringer, M.A. Evaluating the Potential for Enzymatic Acrylamide Mitigation in a Range of Food Products Using an Asparaginase from Aspergillus oryzae. J. Agric. Food Chem. 2009, 57, 4168–4176. [Google Scholar] [CrossRef] [PubMed]
- Weisshaar, R.; Gutsche, B. Formation of acrylamide in heated potato products—Model experiments pointing to asparagine as precursor. Deutsch. Lebensm. Rundsch. 2002, 98, 397–400. [Google Scholar]
- Laing, W.A.; Wright, M.A.; Cooney, J.; Bulley, S.M. From the cover: The missing step of the l-galactose pathway of ascorbate biosynthesis in plants, an l-galactose guanyltransferase, increases leaf ascorbate content. Proc. Natl. Acad. Sci. USA 2007, 104, 9534–9539. [Google Scholar] [CrossRef] [PubMed]
- Lorence, A.; Chevone, B.I.; Mendes, P.; Nessler, C.L. Myo-inositol oxygenase offers a possible entry point into plant ascorbate biosynthesis. Plant Physiol. 2004, 134, 1200–1205. [Google Scholar] [CrossRef] [PubMed]
- Lester, E.; Saftner, R.A. Organically versus Conventionally Grown Produce: Common Production Inputs, Nutritional Quality, and Nitrogen Delivery between the Two Systems. J. Agric. Food Chem. 2011, 59, 10401–10406. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pacifico, D.; Onofri, C.; Parisi, B.; Ostano, P.; Mandolino, G. Influence of Organic Farming on the Potato Transcriptome. Sustainability 2017, 9, 779. https://doi.org/10.3390/su9050779
Pacifico D, Onofri C, Parisi B, Ostano P, Mandolino G. Influence of Organic Farming on the Potato Transcriptome. Sustainability. 2017; 9(5):779. https://doi.org/10.3390/su9050779
Chicago/Turabian StylePacifico, Daniela, Chiara Onofri, Bruno Parisi, Paola Ostano, and Giuseppe Mandolino. 2017. "Influence of Organic Farming on the Potato Transcriptome" Sustainability 9, no. 5: 779. https://doi.org/10.3390/su9050779
APA StylePacifico, D., Onofri, C., Parisi, B., Ostano, P., & Mandolino, G. (2017). Influence of Organic Farming on the Potato Transcriptome. Sustainability, 9(5), 779. https://doi.org/10.3390/su9050779