
Germination of Bouteloua dactyloides and Cynodon dactylon in a Multi-Polluted Soil


Abstract
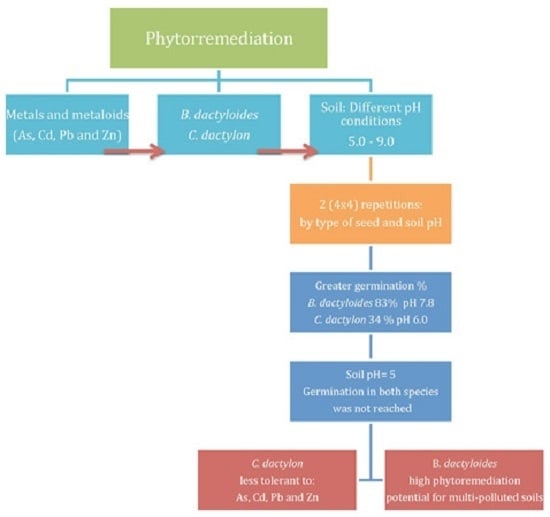
:
1. Introduction
2. Materials and Methods
2.1. Soil Collection
2.2. Soil pH
2.3. Total Metals
2.4. Soluble Metals
2.5. Seed Germination
3. Results
3.1. Total Metals
3.2. Soluble Metals
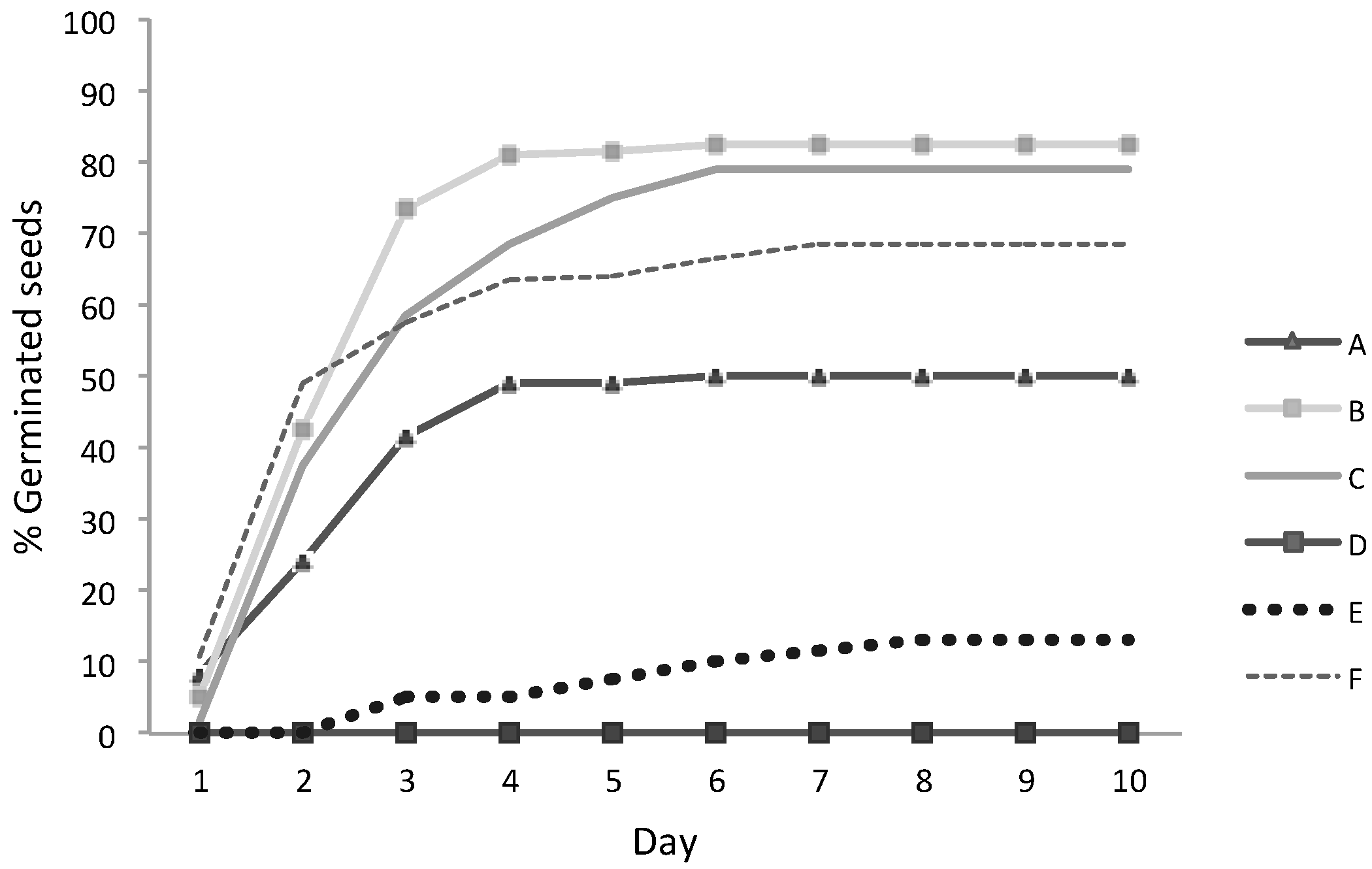
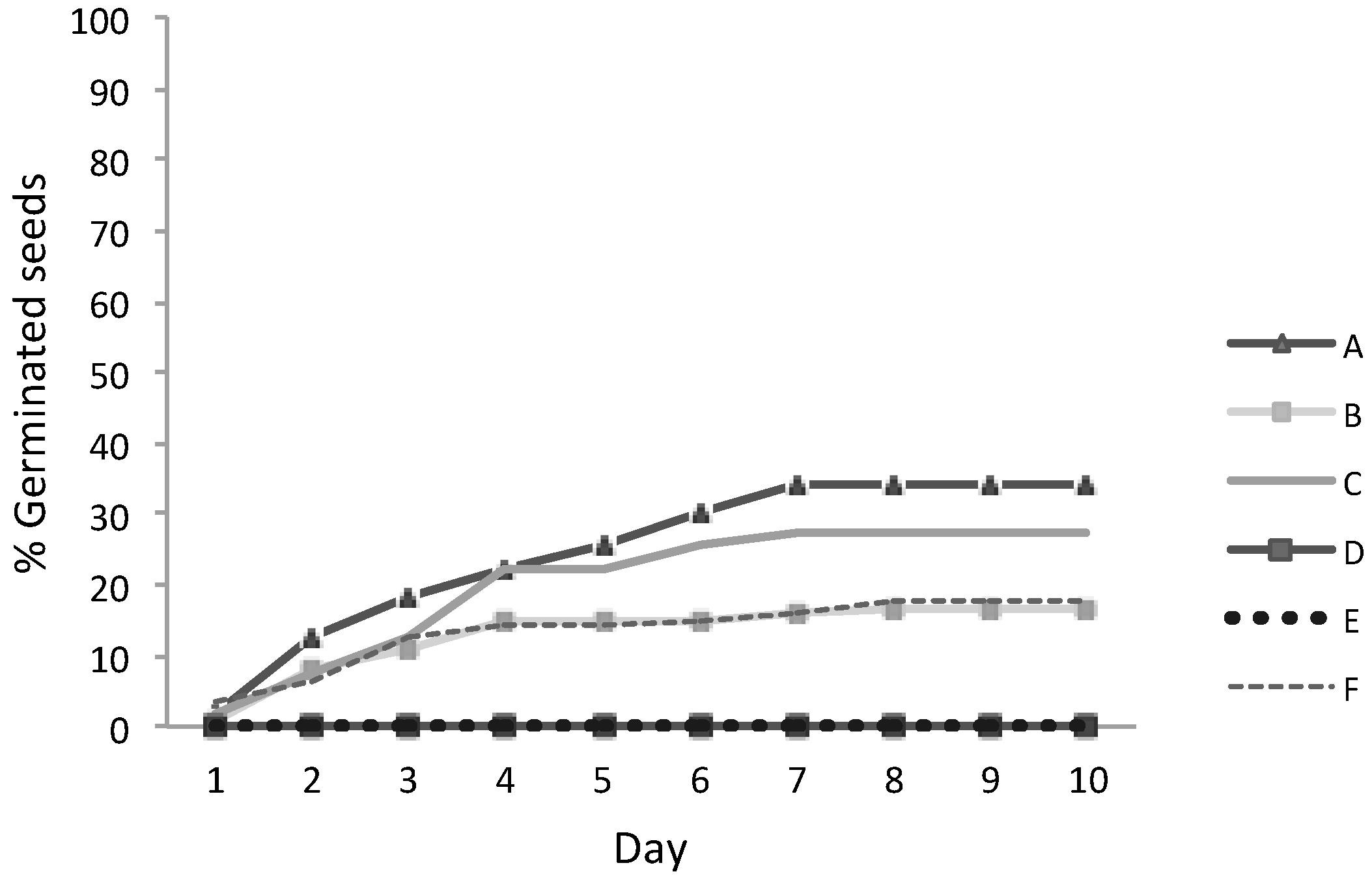
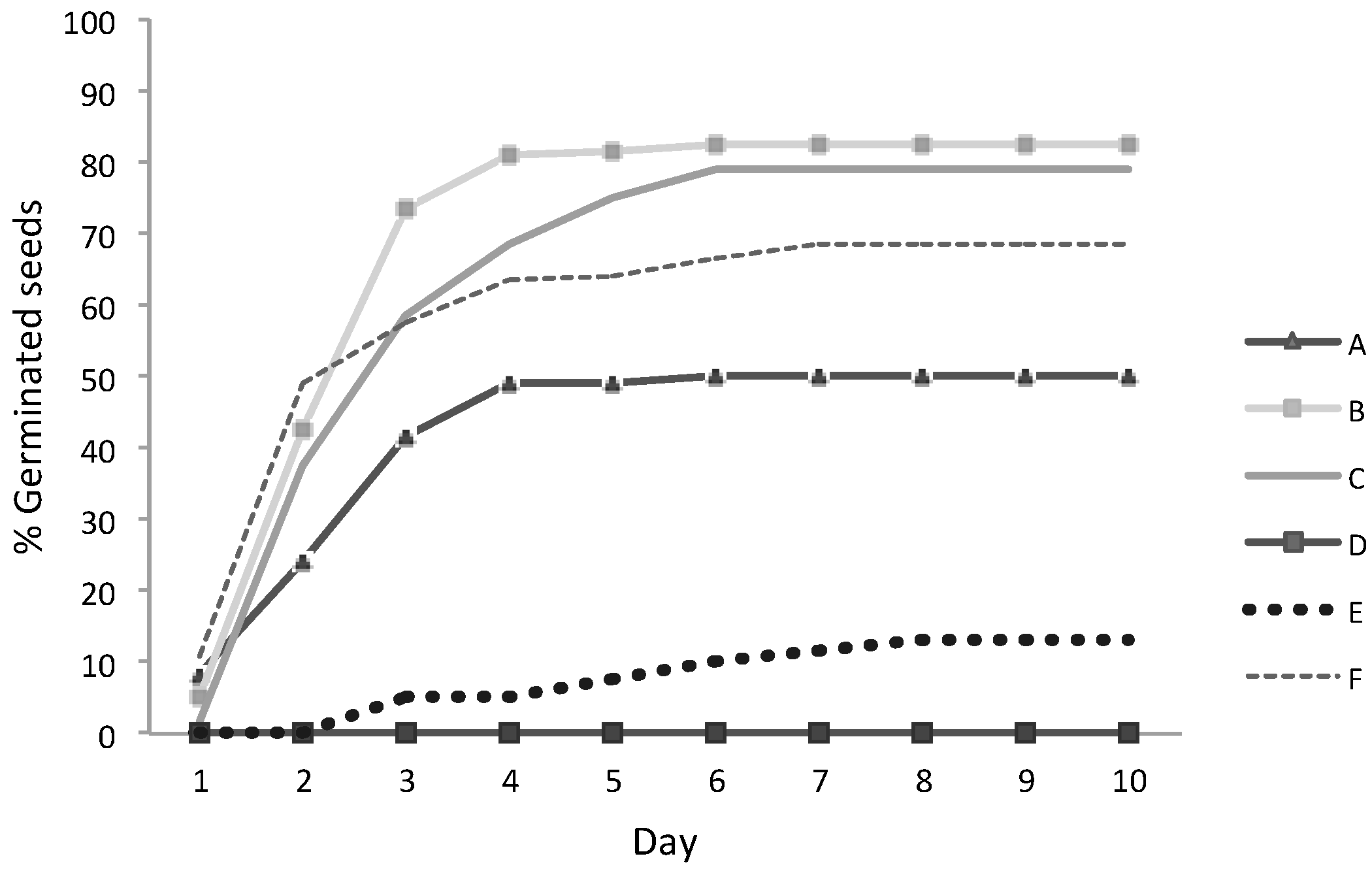
3.3. Seeds Germination
3.4. Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- United States Environmental Protection Agency. Residential Lead Hazard Standards-TSCA Section 403: Office of Pollution Prevention and Toxics; United States Environmental Protection Agency: Washington, DC, USA, 2001.
- Kumar, R.; Kumar Mishra, R.; Mishra, V.; Qidwai, A.; Pandey, A.; Kant Shukla, S.; Pandey, M.; Pathak, A.; Dikshit, A. Detoxification and tolerance of heavy metals in plants. In Plant Metal Interaction; Elsevier: Amsterdam, The Netherlands, 2016; Chapter 13. [Google Scholar]
- Marichali, A.; Dallali, S.; Ouerghemmi, S.; Sebei, H.; Hosni, K. Germination, morpho-physiological and biochemical responses ofcoriander (Coriandrum sativum L.) to zinc excess. Ind. Crop. Prod. 2014, 55, 248–257. [Google Scholar] [CrossRef]
- Chul Kong, I. Joint effects of heavy metal binary mixtures on seed germination, root and shoot growth, bacterial bioluminescence, and gene mutation. J. Environ. Sci. 2013, 25, 889–894. [Google Scholar] [CrossRef]
- Gutiérrez, M.E.; Moreno, M. Los Residuos en la Minería Mexicana; Instituto Nacional de Ecología (INE): D.F., Mexico, 1997. (In Spanish) [Google Scholar]
- Secretaría de Medio Ambiente y Recursos Naturales. Información Proporcionada por la Dirección General de Gestión Integral de Materiales y Actividades Riesgosas; Secretaría de Medio Ambiente y Recursos Naturales: D.F., Mexico, 2004. (In Spanish) [Google Scholar]
- Gulson, B.L.; Masón, K.J.; Korsch, M.J.; Howarth, D. Non-orebody sources are significant contributors to blood lead of some children with low to moderate lead exposure in a mayor mining community. Sci. Total Environ. 1996, 181, 223–230. [Google Scholar] [CrossRef]
- Kranner, I.; Colville, L. Metals and seeds: Biochemical and molecular implications and their significance for seed germination. Environ. Exp. Bot. 2011, 72, 93–105. [Google Scholar] [CrossRef]
- Salt, D.E.; Smith, R.D.; Raskin, I. Phytoremediation. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 643–668. [Google Scholar] [CrossRef] [PubMed]
- Márquez-García, B.; Márquez, C.; Sanjosé, I.; Nieva, F.J.J.; Rodríguez-Rubio, P.; Muñoz-Rodríguez, A.F. The effects of heavy metals on germination and seedling characteristics in two halophyte species in Mediterranean marshes. Mar. Pollut. Bull. 2013, 70, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Čudić, V.; Stojiljković, D.; Jovović, A. Phytoremediation potential of wild plants growing on soil contaminated with heavy metals. Arh. Hig. Rada. Toksikol. 2016, 67, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Grandlic, C.J.; Palmer, M.W.; Maier, R.M. Optimization of plant growth-promoting bacteria-assisted phytostabilization of mine tailings. Soil. Biol. Biochem. 2009, 41, 1734–1740. [Google Scholar] [CrossRef] [PubMed]
- Solís-Dominguez, F.A.; White, S.A.; Borrillo Hutter, T.; Amistadi, M.K.; Root, R.A.; Chorover, J.; Maier, R.M. Response of Key Soil Parameters during Compost-Assisted Phytostabilization in Extremely Acidic Tailings: Effect of Plant Species. Environ. Sci. Technol. 2012, 46, 1019–1027. [Google Scholar] [CrossRef] [PubMed]
- Abou-Shanab, R.; Ghanem, N.; Ghanem, K.; Al-Kolaibe, A. Phytoremediation Potential of Crop and Wild Plants for Multi-metal Contaminated Soils. Res. J. Agric. Biol. Sci. 2007, 3, 370–376. [Google Scholar]
- Sekabira, K.; Oryem–Origa, H.; Mutumba, G.; Kakudidi, E.; Basamba, T.A. Heavy metal phytoremediation by Commelina benghalensis (L.) and Cynodon dactylon (L.) growing in Urban stream sediments. Int. J. Plant Physiol. Biochem. 2011, 3, 133–142. [Google Scholar]
- Brede, D. Turfgrass Maintenance Reduction Handbook: Sports, Lawns, and Golf; Sleeping Bear Press: Chelsea, MI, USA, 2000; p. 386. [Google Scholar]
- Secretaría de Medio Ambiente y Recursos Naturales. NOM-147-SEMARNAT/SSA1-2004, Que Establece Criterios para Determinar las Concentraciones de Remediación de Suelos Contaminados por Arsénico, Bario, Berilio, Cadmio, Cromo Hexavalente, Mercurio, Níquel, Plata, Plomo, Selenio, Talio y/o vanadio; Secretaría de Medio Ambiente y Recursos Naturales: D.F., Mexico, 2004. (In Spanish) [Google Scholar]
- United States Environmental Protection Agency. Method 3051 Microwave Assisted Acid Digestion of Sediments, Sludges, Soils and Oils; United States Environmental Protection Agency: Washington, DC, USA, 2007. Available online: https://www.epa.gov/sites/production/files/2015-12/documents/3051a.pdf (accessed on 21 November 2015).
- Procuraduría Federal de Protección al Ambiente. Disposición y Procedimientos Para la Caracterización y Restauración de Suelos Contaminados; Secretaría de Medio Ambiente y Recursos Naturales, Procuraduría Federal de Protección al Ambiente, Subprocuraduría de Auditoría Ambiental, Dirección General de Emergencias Ambientales: D.F., Mexico, 2000; p. 225. (In Spanish) [Google Scholar]
- United States Environmental Protection Agency. Method 3015A Microwave Assisted Acid Digestion of Aqueous Samples and Extracts; United States Environmental Protection Agency: Washington, DC, USA, 2007. Available online: https://www.epa.gov/sites/production/files/2015-12/documents/3015a.pdf (accessed on 21 November 2015).
- Traina, S.; Laparche, V. Contaminant bioavailability in soils, sediments and aquatic environments. In Proceedings of the National Academy of Sciences Colloquium “Geology, Mineralogy, and Human Welfare”, Irvine, CA, USA, 8–9 November 1999. Proc. Nat. Acad. Sci. USA 1999, 96, 3365–3371. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.M. Toxic Metals in Soil-Plant Systems; John Wiley & Sons: New York, NY, USA, 1994; pp. 63–152. [Google Scholar]
- Wang, D.; Zhang, S.H.; Wang, P.F.; Hou, J.; Zhang, W.J.; Li, W.; Lin, Z.P. The effect of excess Zn on mineral nutrition and antioxidative response in rapeseed seedlings. Chemosphere 2009, 75, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Silveira, M.L.A.; Alleonl, L.R.F.; Guilherme, L.R.G. Biosolids and heavy metals in soils. Sci. Agric. 2003, 60, 793–806. [Google Scholar] [CrossRef]
- Bae, J.; Benoit, D.L.; Watson, A.K. Effect of heavy metals on seed germination and seedling growth of common ragweed and roadside ground cover legumes. Environ. Pollut. 2016, 213, 112–118. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Soil Sample | Soil pH | Irrigation Water pH |
|---|---|---|
| A | 6.0 | 5.4 |
| B | 7.8 | 7.9 |
| C | 7.0 | 7.9 |
| D | 5.0 | 4.6 |
| E | 9.0 | 9.0 |
| F | 8.0 | 7.9 |
| Concentration (mg·kg−1) | |||||
|---|---|---|---|---|---|
| Soil Sample | pH | As | Cd | Pb | Zn |
| A | 6.0 | 2153.30 | 82.58 | 6340.38 | 8082.77 |
| B | 7.8 | 2447.15 | 98.72 | 6227.56 | 11,441.56 |
| C | 7.0 | 1171.16 | 200.52 | 9172.16 | 9506.29 |
| D | 5.0 | 2956.18 | 126.89 | 5455.38 | 14,502.99 |
| E | 9.0 | 2549.33 | 142.91 | 5516.44 | 15,309.95 |
| F | 8.0 | 27.61 | 4.60 | 401.56 | 229.49 |
| Concentration (mg·L−1) | |||||
|---|---|---|---|---|---|
| Soil Sample | pH | As | Cd | Pb | Zn |
| A | 6.0 | 0.09 | 0.04 | 0.03 | 1.28 |
| B | 7.8 | 0.10 | 0.05 | 0.06 | 1.62 |
| C | 7.0 | 0.08 | 0.10 | 0.05 | 0.17 |
| D | 5.0 | 0.07 | 0.36 | 0.02 | 29.51 |
| E | 9.0 | 0.10 | 0.007 | 0.10 | 0.51 |
| F | 8.0 | 0.13 | 0.004 | 0.04 | 0.05 |
| Group | Mean | N | Treatment | |
|---|---|---|---|---|
| a | 24.75 | 4 | BDB | |
| a | ||||
| b | a | 23.75 | 4 | BDC |
| b | ||||
| b | 20.50 | 4 | BDF | |
| c | 15.00 | 4 | BDA | |
| d | 10.25 | 4 | CDA | |
| d | ||||
| e | d | 8.25 | 4 | CDC |
| e | ||||
| e | f | 5.25 | 4 | CDF |
| e | f | |||
| e | f | 5.00 | 4 | CDB |
| f | ||||
| f | 4.00 | 4 | BDE | |
| g | 0.00 | 4 | CDD | |
| g | ||||
| g | 0.00 | 4 | CDE | |
| g | ||||
| g | 0.00 | 4 | BDD | |
| Soil Sample | pH | % Germination | Germinated Seeds Mean | Variation Coefficient (Vc) |
|---|---|---|---|---|
| A | 6.0 | 50.0 | 15.0 ± 5.9 | 39.6 |
| B | 7.8 | 83 | 24.8 ± 3.4 | 13.8 |
| C | 7.0 | 79 | 23.8 ± 2.5 | 10.5 |
| D | 5.0 | 0 | 0 | - |
| E | 9.0 | 13 | 4.0 ± 3.6 | 89.0 |
| F | 8.0 | 68 | 20.5 ± 2.6 | 12.9 |
| Soil Sample | pH | % Germination | Germinated Seeds Mean | Variation Coefficient (Vc) |
|---|---|---|---|---|
| A | 6.0 | 34 | 10.3 ± 3.0 | 29.1 |
| B | 7.8 | 17 | 5.0 ± 1.8 | 36.5 |
| C | 7.0 | 28 | 8.3 ± 1.0 | 11.6 |
| D | 5.0 | 0 | 0 | - |
| E | 9.0 | 0 | 0 | - |
| F | 8.0 | 18 | 5.3 ± 2.2 | 42.2 |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado-Caballero, M.D.R.; Alarcón-Herrera, M.T.; Valles-Aragón, M.C.; Melgoza-Castillo, A.; Ojeda-Barrios, D.L.; Leyva-Chávez, A. Germination of Bouteloua dactyloides and Cynodon dactylon in a Multi-Polluted Soil. Sustainability 2017, 9, 81. https://doi.org/10.3390/su9010081
Delgado-Caballero MDR, Alarcón-Herrera MT, Valles-Aragón MC, Melgoza-Castillo A, Ojeda-Barrios DL, Leyva-Chávez A. Germination of Bouteloua dactyloides and Cynodon dactylon in a Multi-Polluted Soil. Sustainability. 2017; 9(1):81. https://doi.org/10.3390/su9010081
Chicago/Turabian StyleDelgado-Caballero, María Del Rosario, María Teresa Alarcón-Herrera, María Cecilia Valles-Aragón, Alicia Melgoza-Castillo, Dámaris Lepoldina Ojeda-Barrios, and Arwell Leyva-Chávez. 2017. "Germination of Bouteloua dactyloides and Cynodon dactylon in a Multi-Polluted Soil" Sustainability 9, no. 1: 81. https://doi.org/10.3390/su9010081
APA StyleDelgado-Caballero, M. D. R., Alarcón-Herrera, M. T., Valles-Aragón, M. C., Melgoza-Castillo, A., Ojeda-Barrios, D. L., & Leyva-Chávez, A. (2017). Germination of Bouteloua dactyloides and Cynodon dactylon in a Multi-Polluted Soil. Sustainability, 9(1), 81. https://doi.org/10.3390/su9010081