
Optimization of Cellulase and Xylanase Production by Micrococcus Species under Submerged Fermentation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sample Collection
2.2. Isolation of Cellulose and Xylan-Degrading Bacteria
2.3. Growth Media Compositions
2.4. Screening for Cellulase and Xylanase-Producing Bacteria
2.5. Identification of the Bacterial Isolate by 16S rRNA Gene Sequencing
2.6. Cellulase and Xylanase Production
2.7. Cellulase and Xylanase Activity
2.8. Optimization of Culture Conditions for Enzyme Production
2.8.1. Effect of Temperature
2.8.2. Effect of Initial pH of Growth Medium
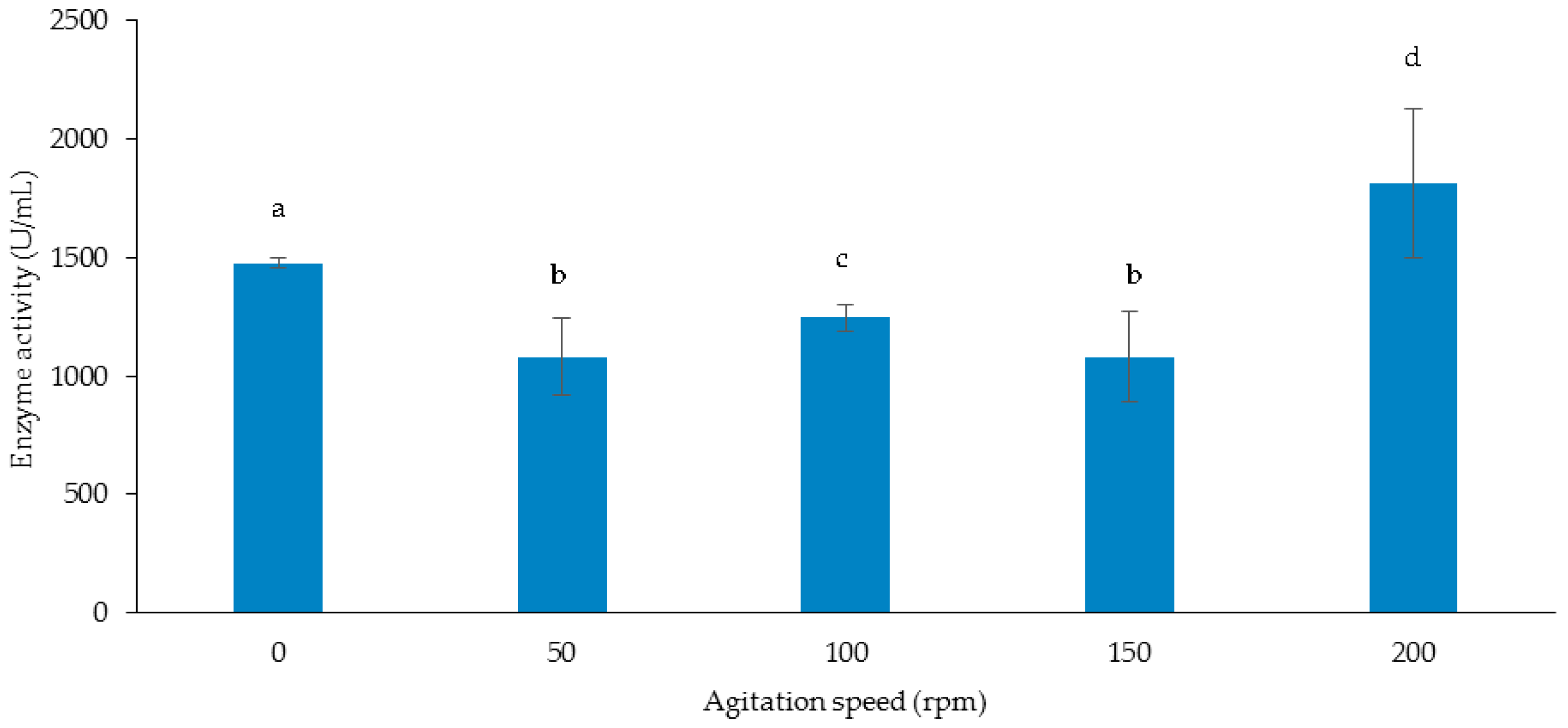
2.8.3. Effect of Agitation Speed
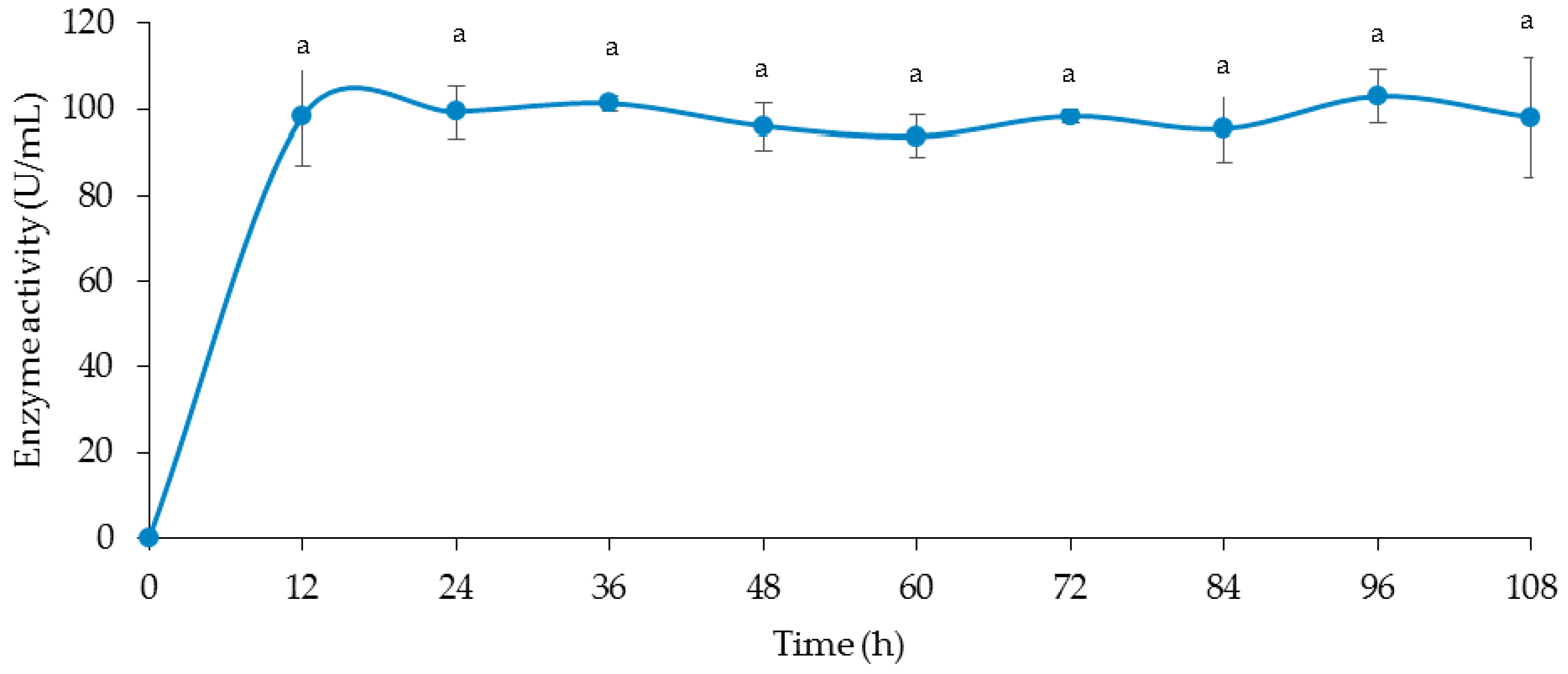
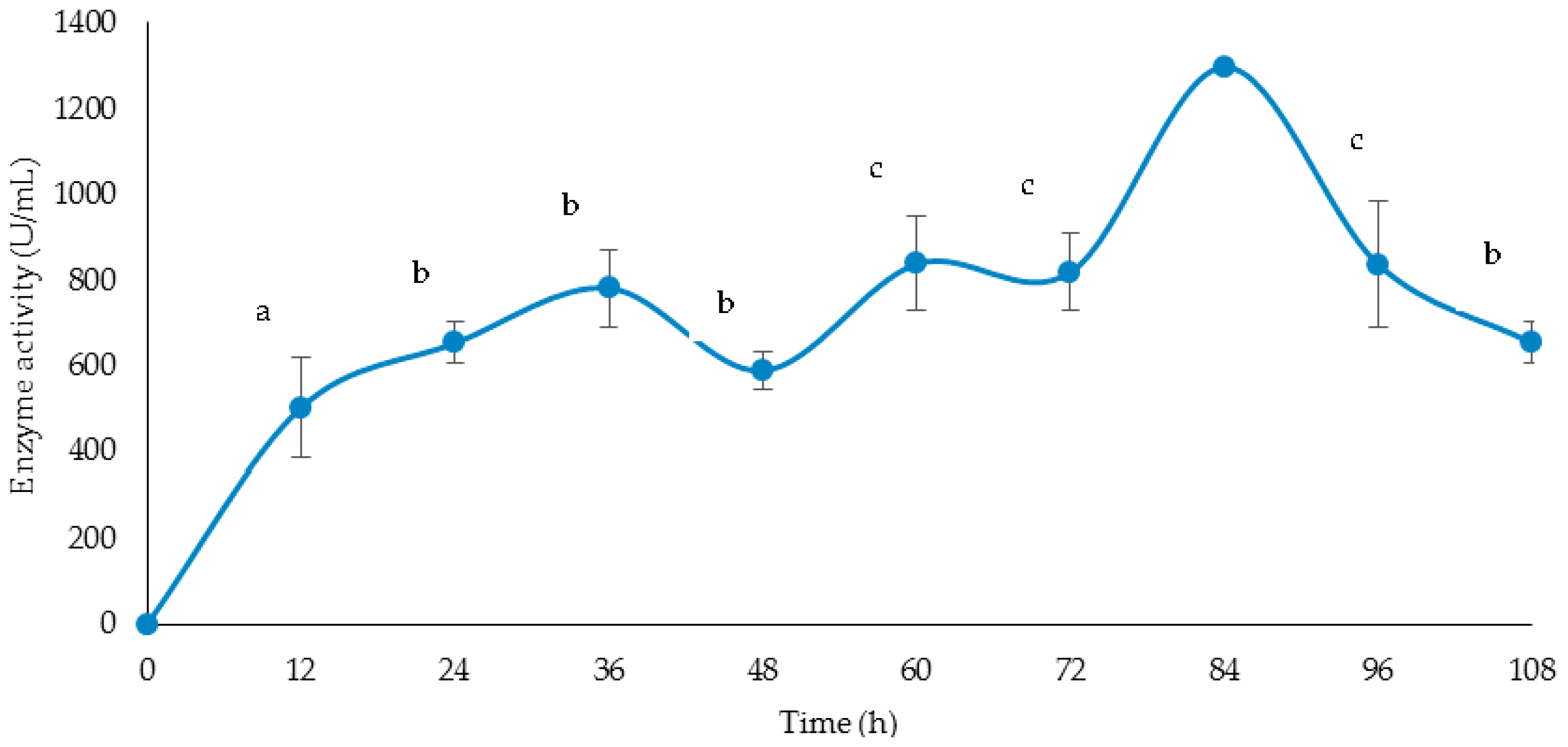
2.9. Time Course for Cellulase and Xylanase Production
2.10. Data Analysis
3. Results
3.1. Isolation and Screening for Cellulase and Xylanase Activity
3.2. Identification of the Bacterial Isolates by 16S rRNA Gene Nucleotide Sequencing
3.3. Optimization of Culture Conditions for Enzymes Production by Micrococcus sp. SAMRC-UFH3
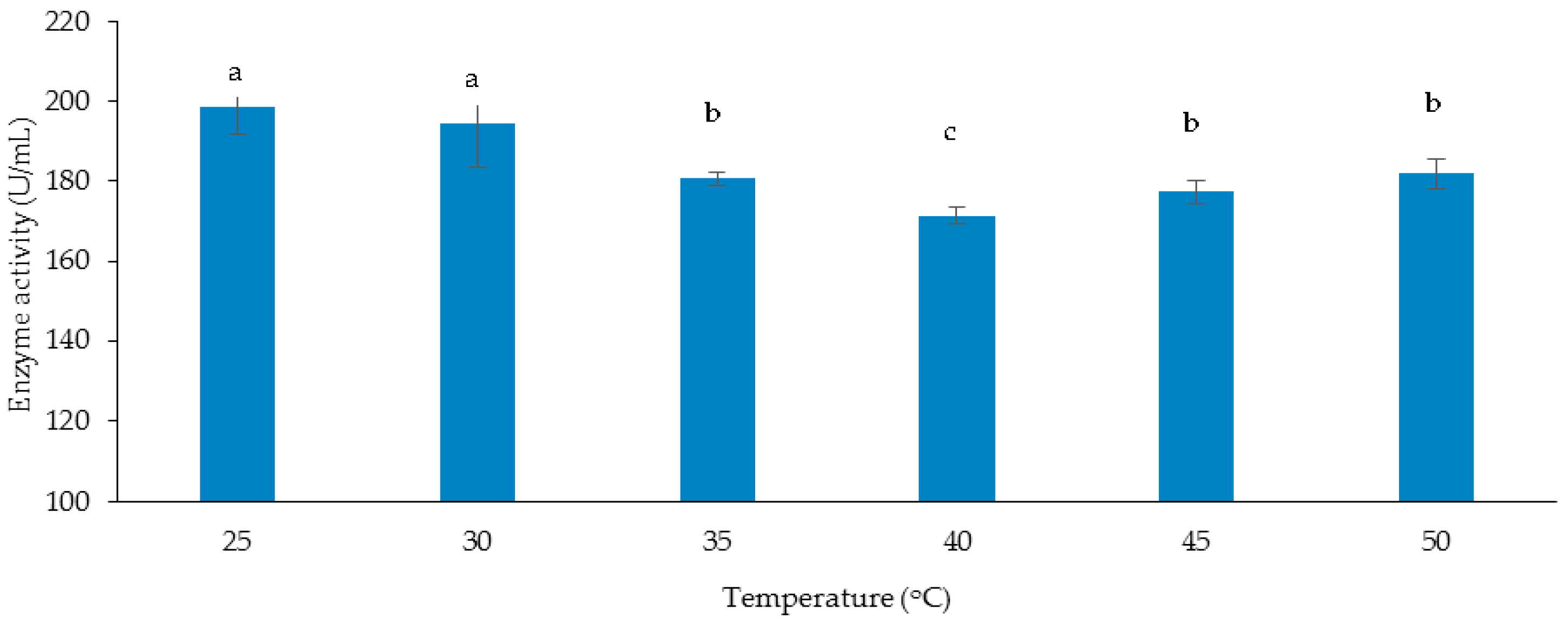
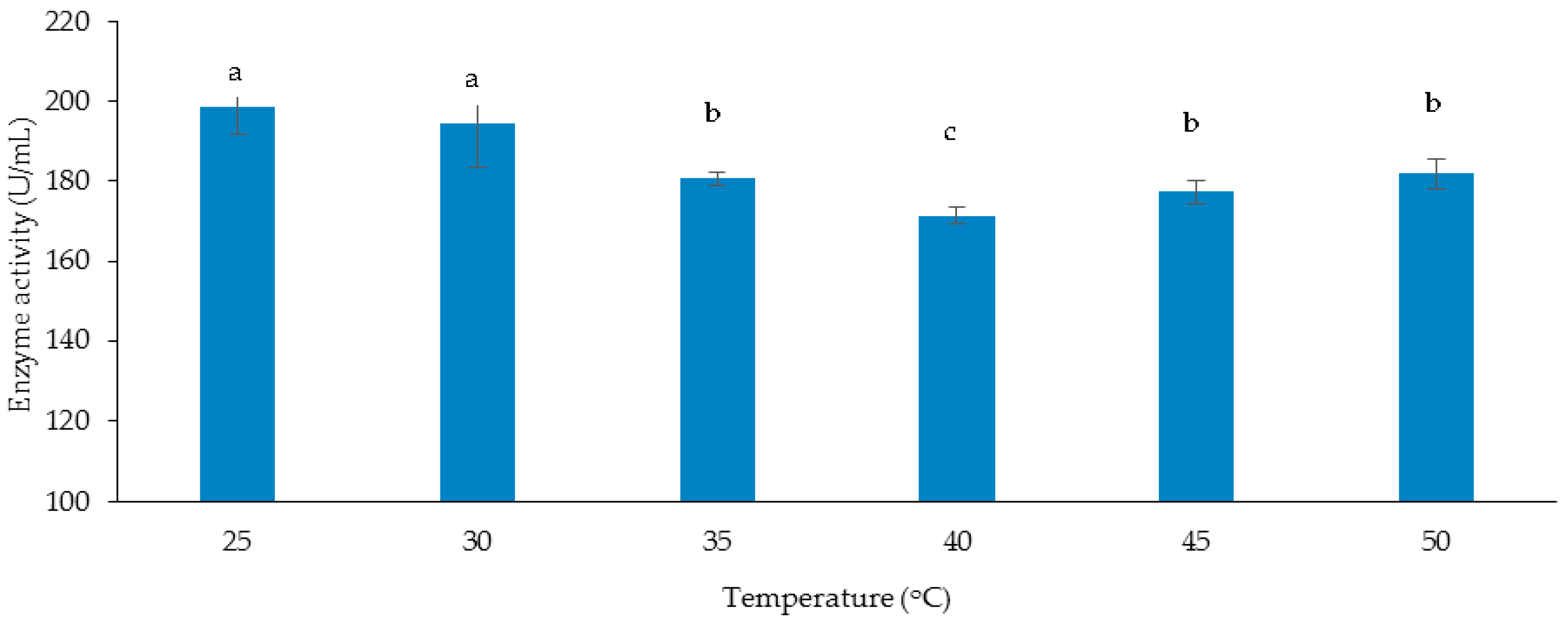
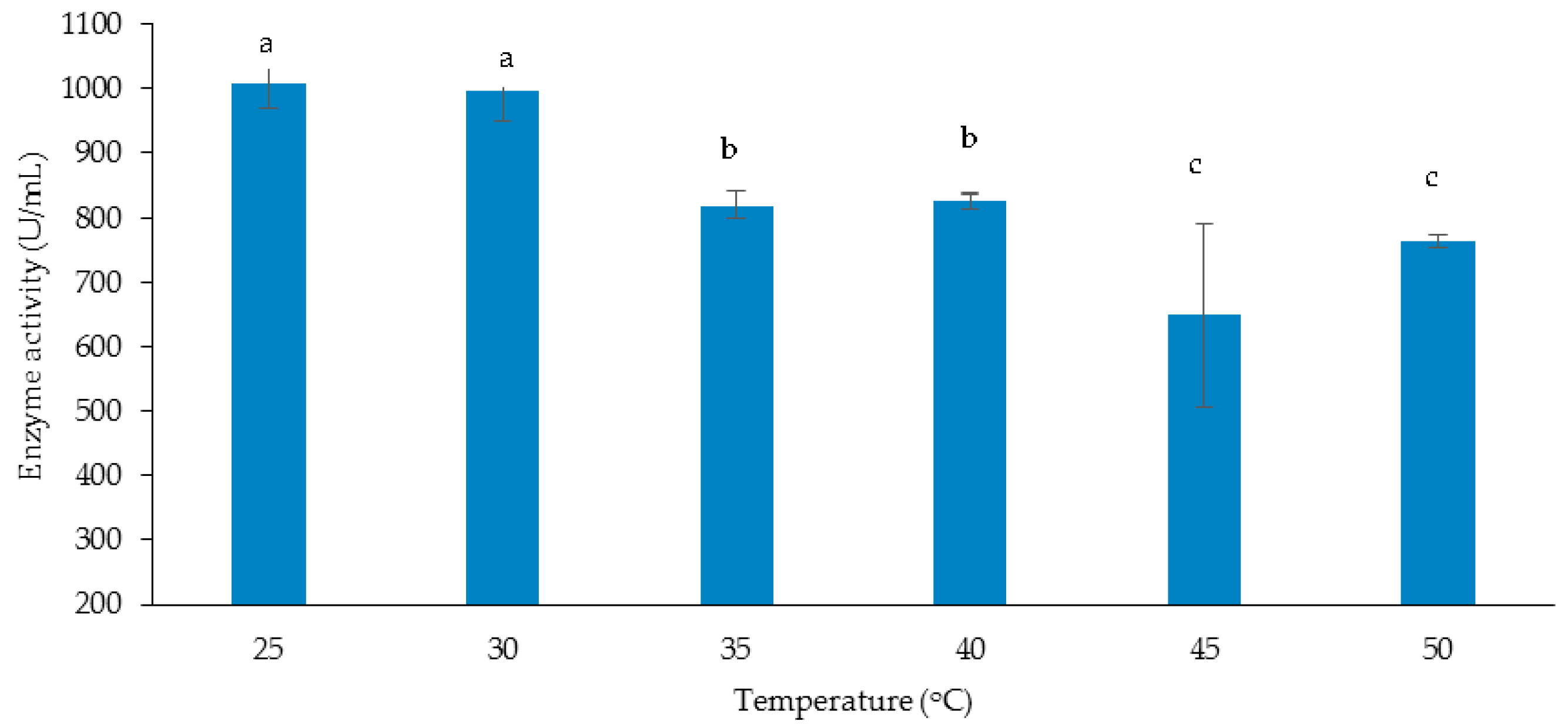
3.3.1. Effect of Temperature on Cellulase and Xylanase Production
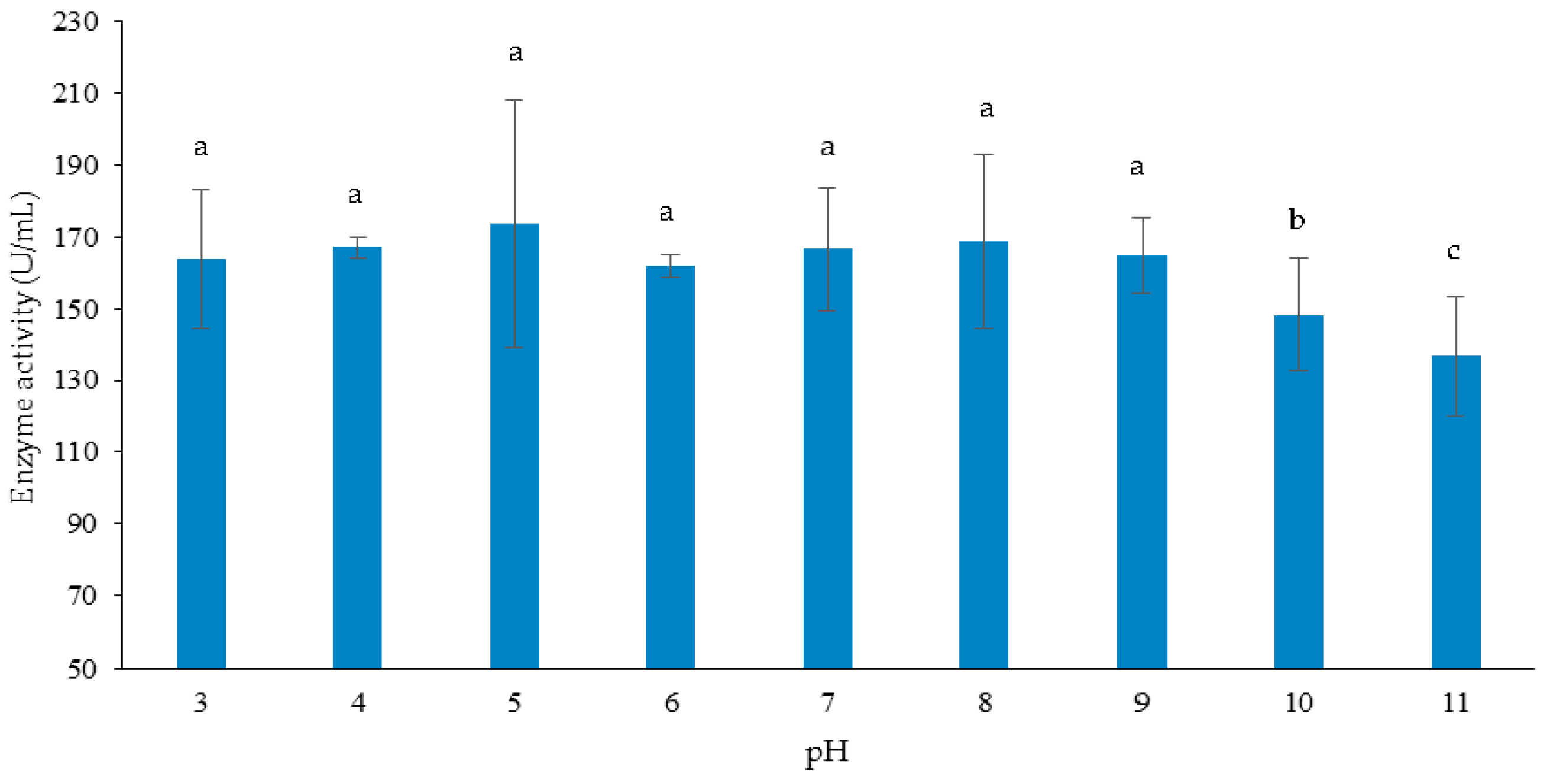
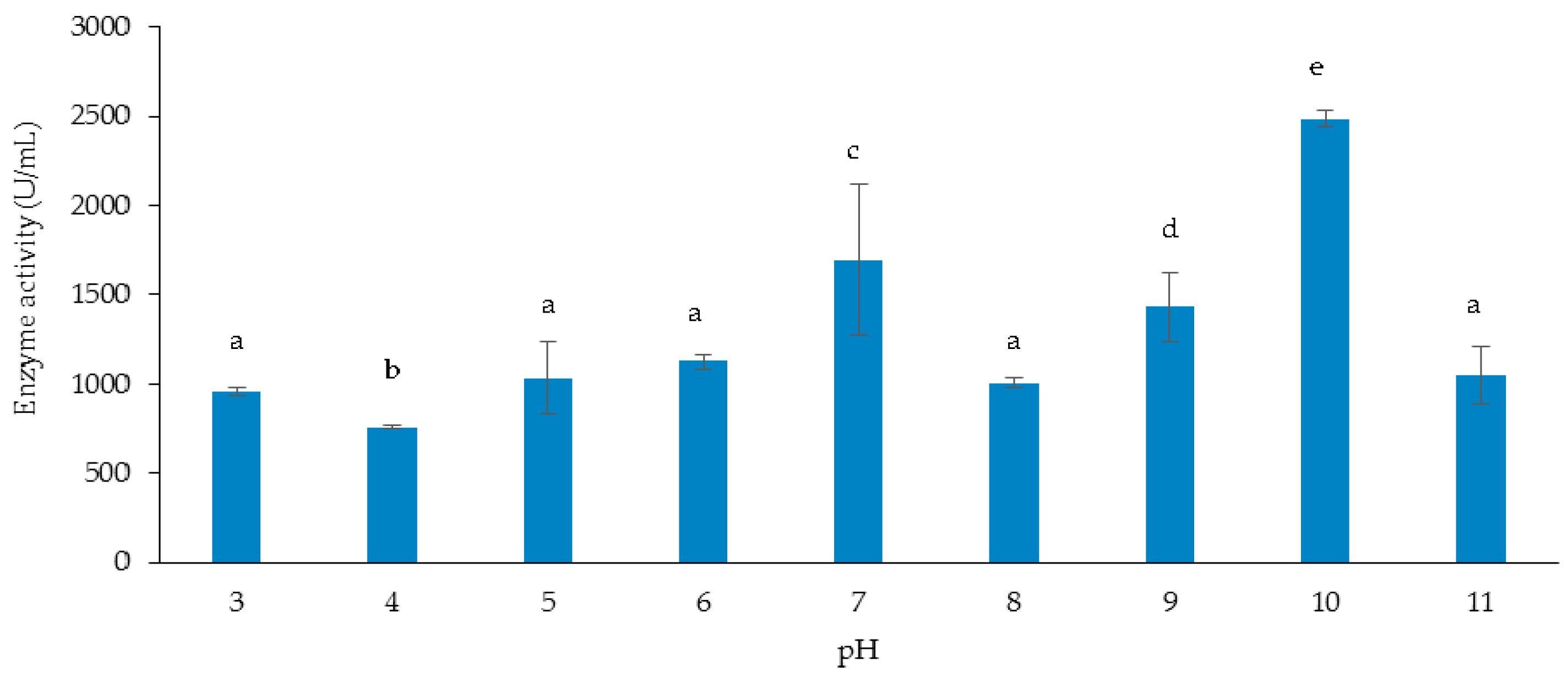
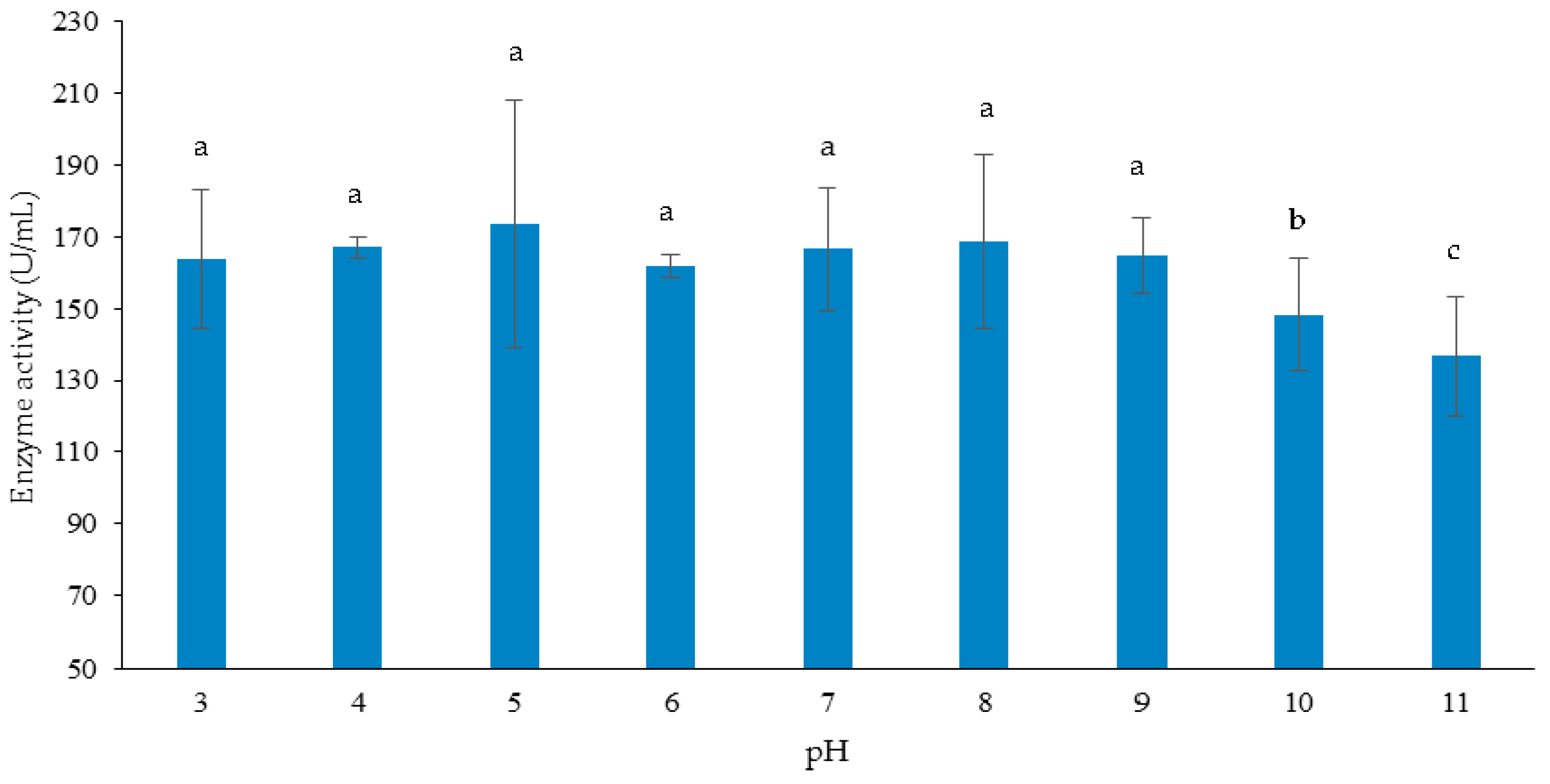
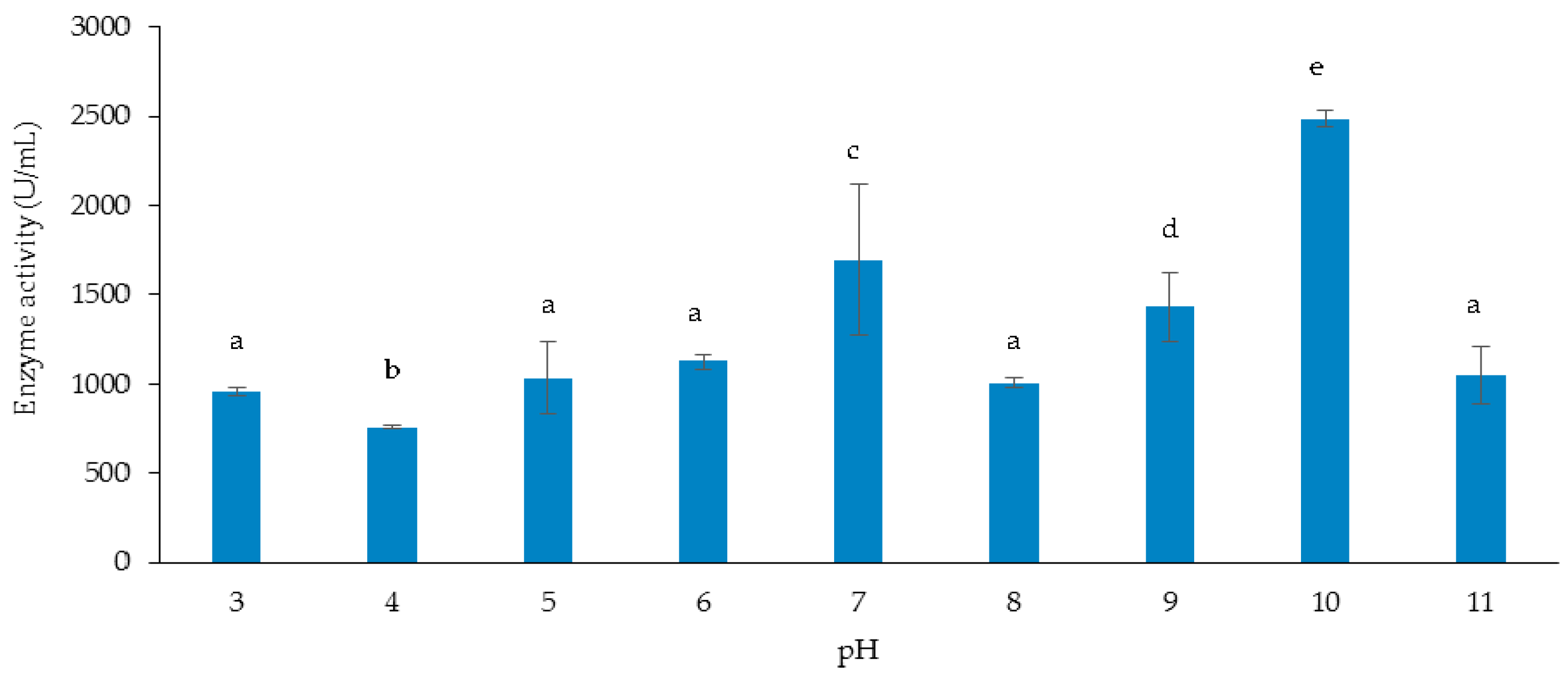
3.3.2. Effect of Initial pH of Growth Medium on Cellulase and Xylanase Production
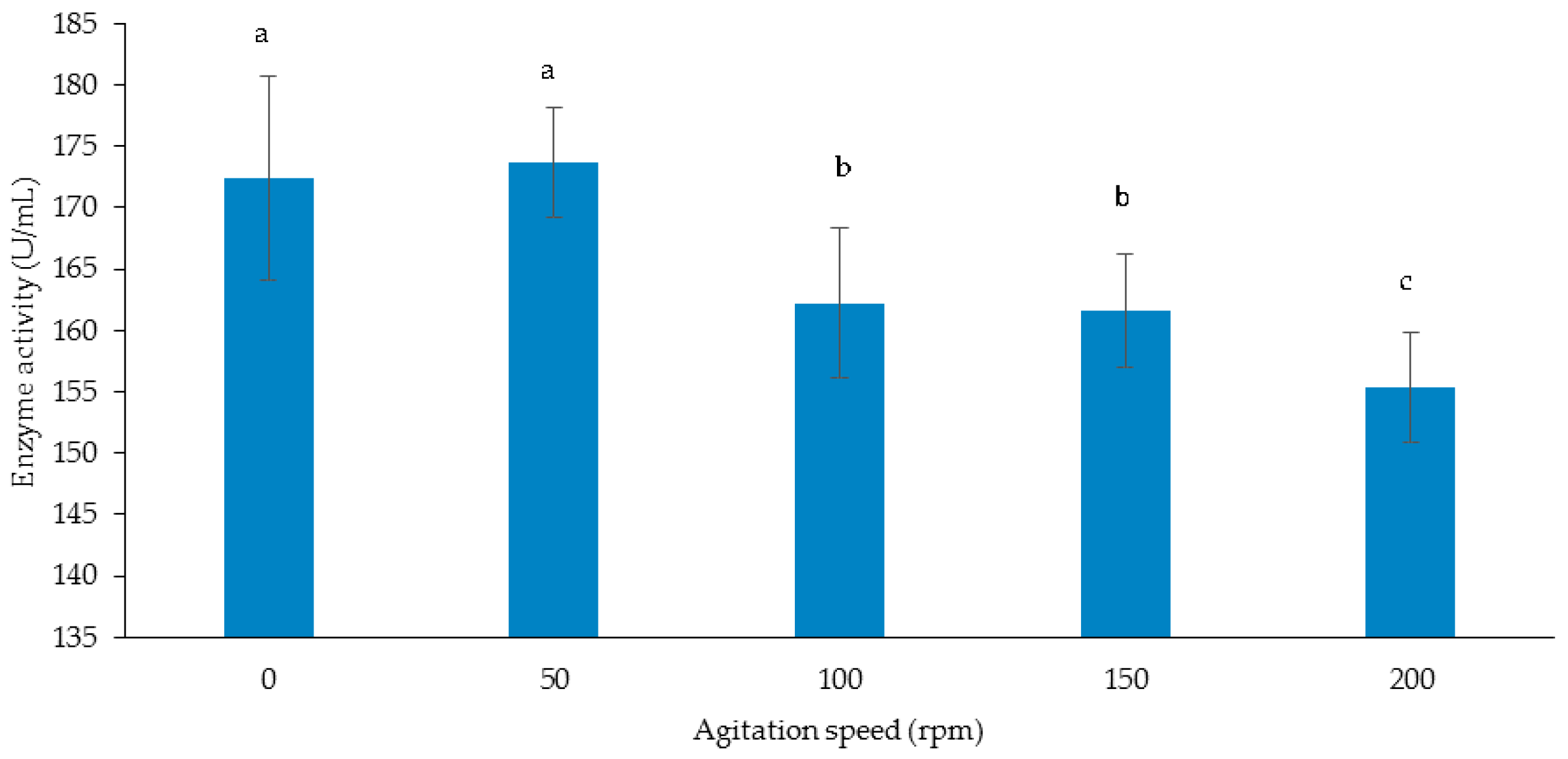
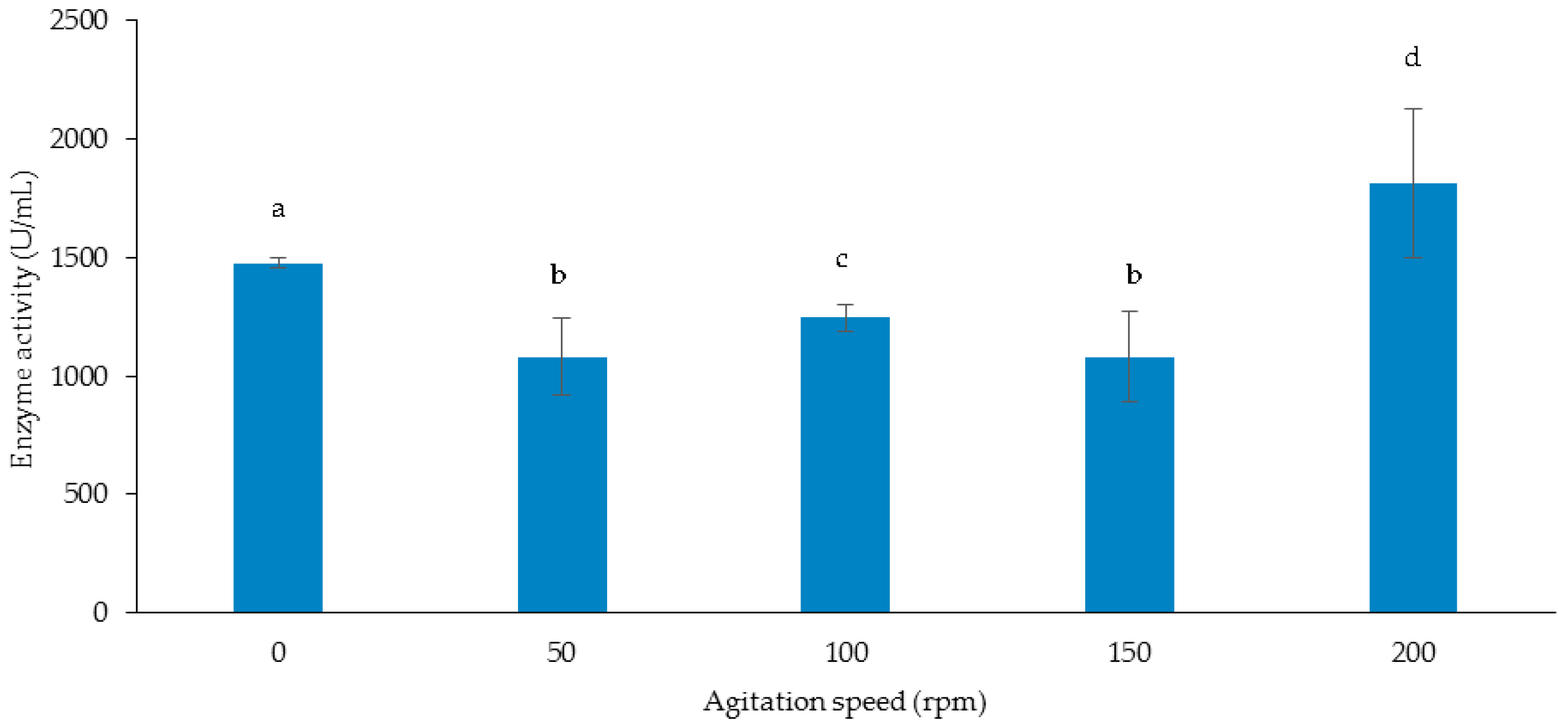
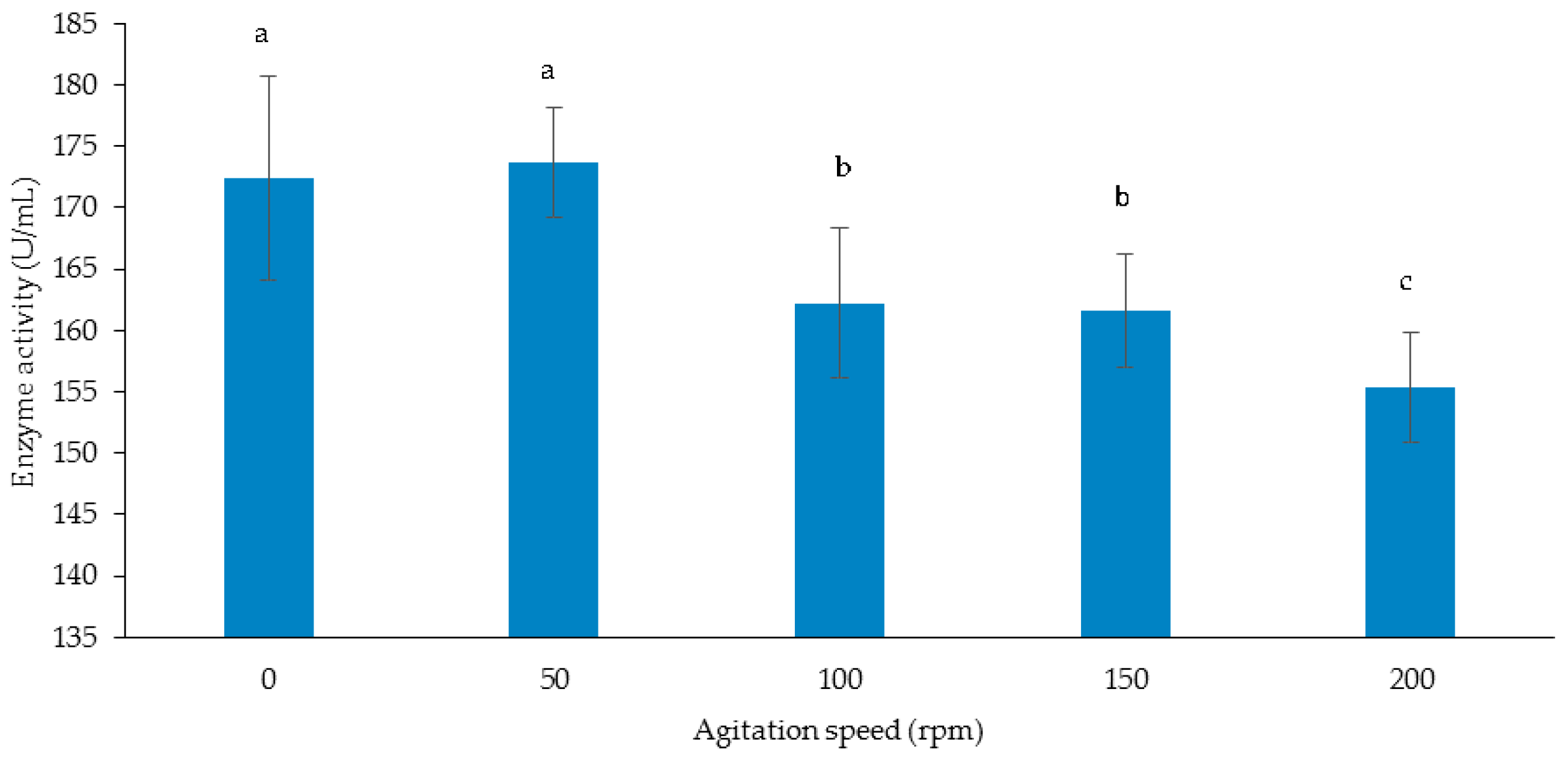
3.3.3. Effect of Agitation Speed on Cellulase and Xylanase Production
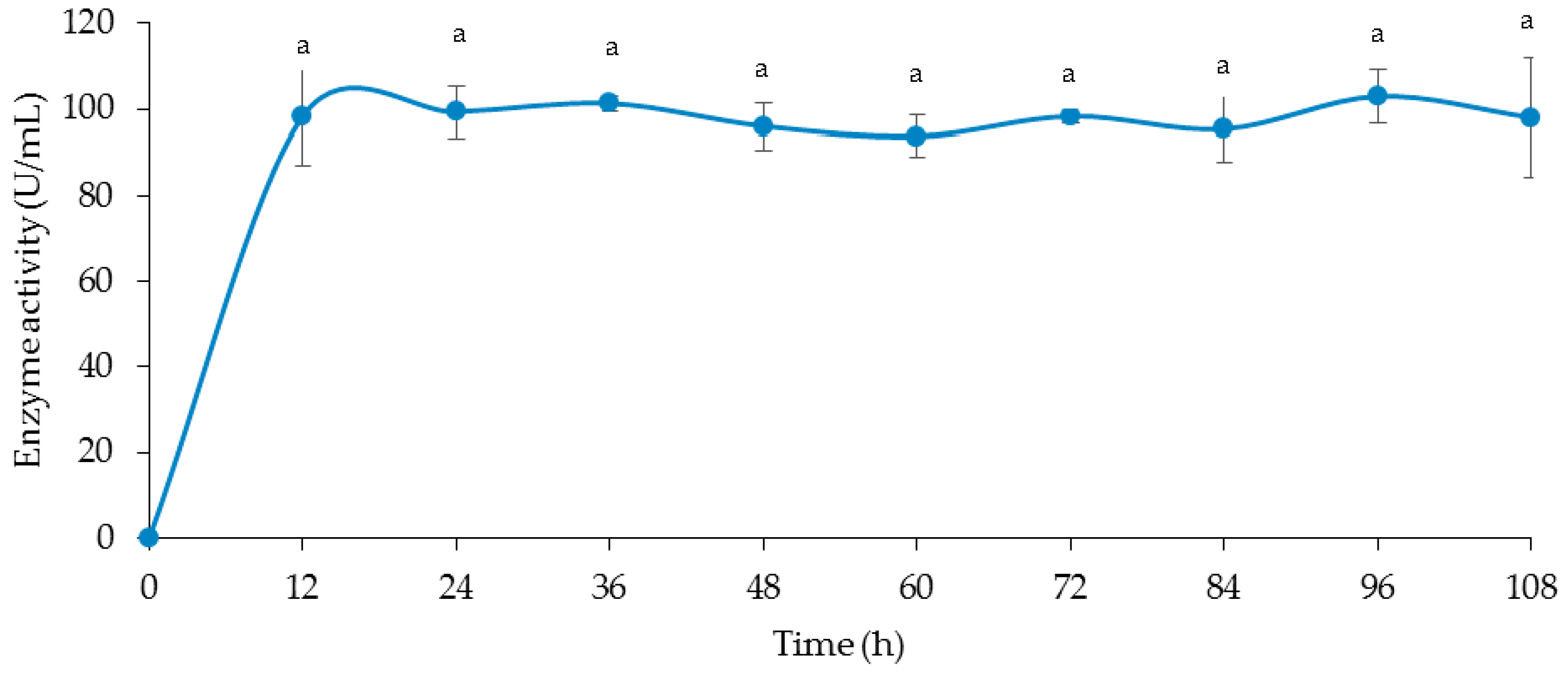
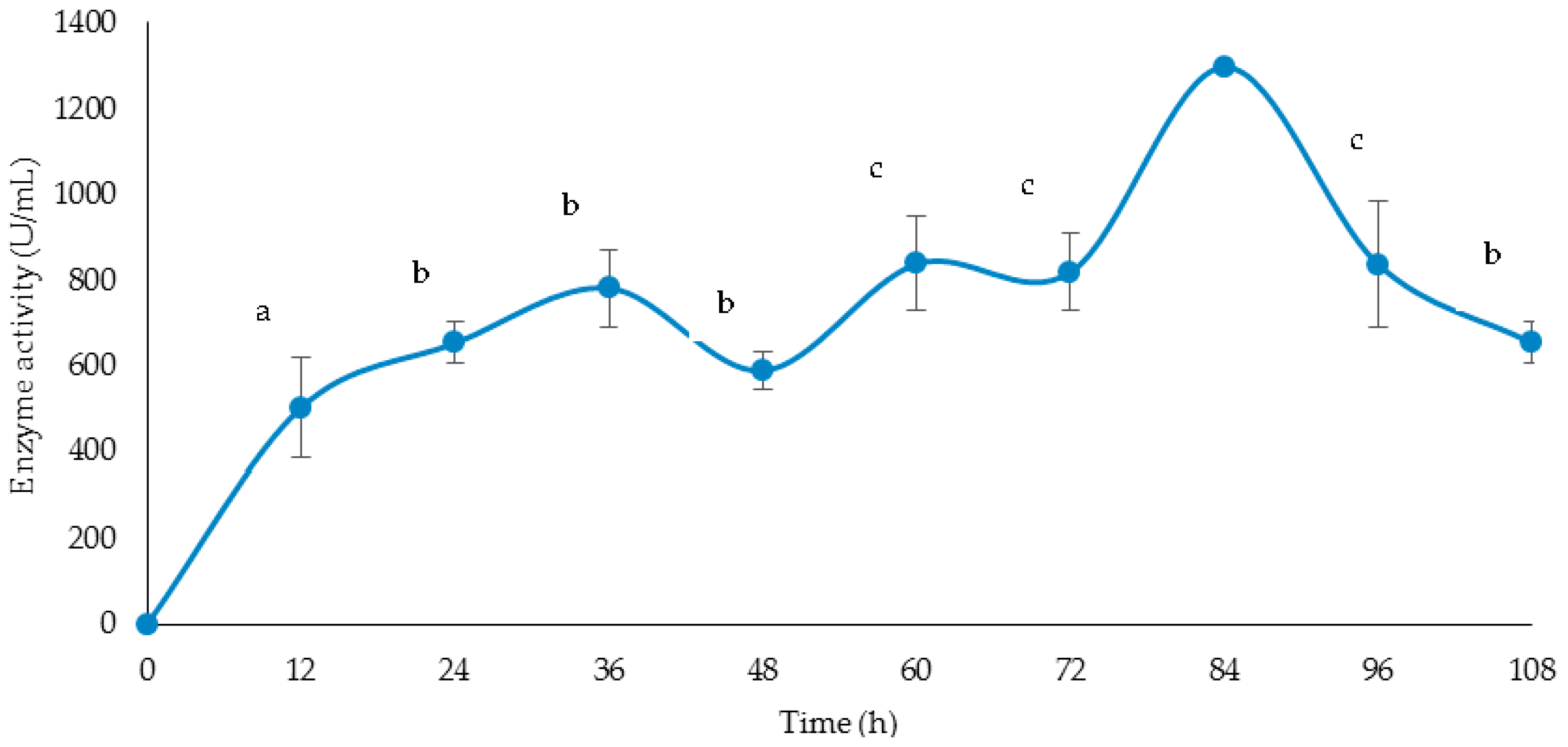
3.4. Time Course of Cellulase and Xylanase Production
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kumar, R.; Singh, S.; Singh, O.V. Bioconversion of lignocellulosic biomass: Biochemical and molecular perspectives. J. Ind. Microbiol. Biotechnol. 2008, 35, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, C. Lignocellulosic residues: Biodegradation and bioconversion by fungi. Biotechnol. Adv. 2009, 27, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.L.; Abotsi, E.; Van Rensburg, E.J.; Howard, S. Lignocellulose biotechnology: Issues of bioconversion and enzyme production. Afr. J. Biotechnol. 2004, 2, 602–619. [Google Scholar] [CrossRef]
- Subramaniyan, S.; Prema, P. Biotechnology of microbial xylanases: Enzymology, molecular biology, and application. Crit. Rev. Biotechnol. 2002, 22, 33–64. [Google Scholar] [CrossRef] [PubMed]
- Techapun, C.; Charoenrat, T.; Watanabe, M.; Sasaki, K.; Poosaran, N. Optimization of thermostable and alkaline-tolerant cellulase-free xylanase production from agricultural waste by thermotolerant Streptomyces sp. Ab106, using the central composite experimental design. Biochem. Eng. J. 2002, 12, 99–105. [Google Scholar] [CrossRef]
- Ghatora, S.K.; Chadha, B.S.; Badhan, A.K.; Saini, H.S.; Bhat, M.K. Identification and characterization of diverse xylanases from thermophilic and thermotolerant fungi. Bioresour. Technol. 2007, 1, 18–33. [Google Scholar]
- Kulkarni, N.; Rao, M. Application of xylanase from alkaliphilic thermophilic Bacillus sp. NCIM 59 in bioleaching of bagasse pulp. J. Biotechnol. 1996, 51, 167–173. [Google Scholar] [CrossRef]
- Yu, H.; Zeng, G.; Huang, H.; Xi, X.; Wang, R.; Huang, D.; Huang, G.; Li, J. Microbial community succession and lignocellulose degradation during agricultural waste composting. Biodegradation 2007, 18, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Doi, R.H. Cellulases of mesophilic microorganisms-Cellulosome and non-cellulosome producers. Incred. Anaerob. Physiol. Genom. Fuels 2008, 1125, 267–279. [Google Scholar]
- Ray, A.K.; Bairagi, A.; Ghosh, K.S.; Sen, S.K. Optimization of fermentation conditions for cellulase production by Bacillus subtilis CY5 and Bacillus circulans TP3 isolated from fish gut. Acta Ichthyol. Piscat. 2007, 1, 47–53. [Google Scholar] [CrossRef]
- Bischoff, K.M.; Rooney, A.P.; Li, X.L.; Liu, S.; Hughes, S.R. Purification and characterization of a family 5 endoglucanase from a moderately thermophilic strain of Bacillus licheniformis. Biotechnol. Lett. 2006, 28, 1761–1765. [Google Scholar] [CrossRef] [PubMed]
- Camassola, M.; De Bittencourt, L.R.; Shenem, N.T.; Andreaus, J.; Dillon, A.J.P. Characterization of the cellulase complex of Penicillium echinulatum. Biocatal. Biotransform. 2004, 22, 391–396. [Google Scholar] [CrossRef]
- Haakana, H.; Miettinen-Oinonen, A.; Joutsjoki, V.; Mäntylä, A.; Suominen, P.; Vehmaanperä, J. Cloning of cellulase genes from Melanocarpus albomyces and their efficient expression in Trichoderma reesei. Enzyme Microb. Technol. 2004, 34, 159–167. [Google Scholar] [CrossRef]
- Silva, R.D.; Lago, E.S.; Merheb, C.W.; Macchione, M.M.; Park, Y.K.; Gomes, E. Production of xylanase and CMCase on solid state fermentation in different residues by Thermoascus aurantiacus miehe. Braz. J. Microbiol. 2005, 36, 235–241. [Google Scholar] [CrossRef]
- Semêdo, L.T.; Gomes, R.C.; Bon, E.P.; Soares, R.M.; Linhares, L.F.; Coelho, R.R. Endocellulase and exocellulase activities of two Streptomyces strains isolated from a forest soil. In Twenty-First Symposium on Biotechnology for Fuels and Chemicals; Humana Press: New York, NY, USA, 2000; pp. 267–276. [Google Scholar]
- Doddamani, H.P.; Ninnekar, H.Z. Biodegradation of carbaryl by a Micrococcus species. Curr. Microbiol. 2001, 43, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Nisha, P.; Das, A.; Saritha, K.V. Production of cellulase from Micrococcus sp. and effect of growth parameters. 2014, 2, 236–240. [Google Scholar]
- Sandrin, T.R.; Maier, R.M. Impact of metals on the biodegradation of organic pollutants. Environ. Health Perspect. 2003, 111, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Young, K.S.; de Abreu, C.N. (Eds.) Internet Addiction: A Handbook and Guide to Evaluation and Treatment; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010.
- Zhuang, L.; Wai, J.S.; Embrey, M.W.; Fisher, T.E.; Egbertson, M.S.; Payne, L.S.; Guare, J.P.; Vacca, J.P.; Hazuda, D.J.; Felock, P.J.; et al. Design and synthesis of 8-hydroxy-[1, 6] naphthyridines as novel inhibitors of HIV-1 integrase in vitro and in infected cells. J. Med. Chem. 2003, 46, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.Z.; Manchur, M.A.; Anwar, M.N. Isolation, purification, characterization of cellulolytic enzymes produced by the isolate Streptomyces omiyaensis. Pak. J. Biol. Sci. 2004, 7, 1647–1653. [Google Scholar]
- Immanuel, G.; Dhanusha, R.; Prema, P.; Palavesam, A. Effect of different growth parameters on endoglucanase enzyme activity by bacteria isolated from coir retting effluents of estuarine environment. Int. J. Environ. Sci. Technol. 2006, 3, 25–34. [Google Scholar] [CrossRef]
- Jeffrey, L.S.H. Isolation, characterization and identification of actinomycetes from agriculture soils at Semongok, Sarawak. Afr. J. Biotechnol. 2008, 7, 3697–3702. [Google Scholar]
- Kasana, R.C.; Salwan, R.; Dhar, H.; Dutt, S.; Gulati, A. A rapid and easy method for the detection of microbial cellulases on agar plates using Gram’s iodine. Curr. Microbiol. 2008, 57, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Pointing, S.B. Qualitative methods for the determination of lignocellulolytic enzyme production by tropical fungi. Funngal Divers. 1999, 2, 17–33. [Google Scholar]
- Teather, R.M.; Wood, P.J. Use of Congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Appl. Environ. Microbiol. 1982, 43, 777–780. [Google Scholar] [PubMed]
- Mangamuri, U.K.; Muvva, V.; Poda, S.; Kamma, S. Isolation, identification and molecular characterization of rare actinomycetes from mangrove ecosystem of Nizampatnam. Malays. J. Microbiol. 2012, 8, 83–91. [Google Scholar]
- Sharma, P.; Bajaj, B.K. Production and partial characterization of alkali-tolerant xylanase from an alkalophilic Streptomyces sp. CD3. J. Sci. Industr. Res. 2005, 64, 688–697. [Google Scholar]
- Bajaj, B.K.; Khajuria, Y.P.; Singh, V.P. Agricultural residues as potential substrates for production of xylanase from alkali-thermo-tolerant bacterial isolate. Biocat. Agric. Biotechnol. 2012, 1, 314–320. [Google Scholar]
- Swe, K.T. Screening Potential Lignin-Degrading Microorganisms and Evaluating Their Optimal Enzyme Producing Growth Conditions; Chalmers University of Technology: Goteborg, Sweden, 2011. [Google Scholar]
- Adhyaru, D.N.; Bhatt, N.S.; Modi, H.A. Enhanced production of cellulase-free, thermo-alkali-solvent-stable xylanase from Bacillus altitudinis DHN8, its characterization and application in sorghum straw saccharification. Biocat. Agric. Biotechnol. 2014, 3, 182–190. [Google Scholar] [CrossRef]
- Nandimath, A.P.; Kharat, K.R.; Gupta, S.G.; Kharat, A.S. Optimization of cellulase production for Bacillus sp. and Pseudomonas sp. soil isolates. Afr. J. Microbiol. Res. 2016, 10, 410–419. [Google Scholar]
- Lugani, Y.; Singla, R.; Sooch, B.S. Optimization of Cellulase Production from Newly Isolated Bacillus sp. Y3. J. Bioproc. Biotech. 2015, 5. [Google Scholar] [CrossRef]
- Vijayakumar, R.; Muthukumar, C.; Thajuddin, N.; Panneerselvam, A.; Saravanamuthu, R. Studies on the diversity of actinomycetes in the Palk Strait region of Bay of Bengal, India. Actinomycetologica 2007, 21, 59–65. [Google Scholar] [CrossRef]
- Insam, H.; Franke-Whittle, I.; Goberna, M. Microbes in aerobic and anaerobic waste treatment. In Microbes at Work; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1–34. [Google Scholar]
- Hamedani, K.; Soudbakhsh, M.N.; Das, A.; Prashanthi, K.; Bhattacharya, S.; Suryan, S. Enzymatic screening, antibacterial potential and molecular characterization of streptomycetes isolated from Wayanad district in Kerala, India. Int. J. Pharm. Biol. Sci. 2012, 2, 201–210. [Google Scholar]
- Das, A.; Bhattacharya, S.; Murali, L. Production of cellulase from a thermophilic Bacillus sp. isolated from cow dung. Am.-Eurasian J. Agric. Environ. Sci. 2010, 8, 685–691. [Google Scholar]
- Gomez, L.D.; Steele-King, C.G.; McQueen-Mason, S.J. Sustainable liquid biofuels from biomass: The writing’s on the walls. New Phytol. 2008, 178, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Madson, P.W.; Tereck, C.D. Lignocellulosic. Feed Stocks for Ethanol Production: The Ultimate Renewable Energy Source. 2004. Available online: http://www.nt.ntnu.no/users/skoge/prost/proceedings/aiche-2004/pdffiles/papers/035a.pdf (accessed on 10 November 2016).
- Bayer, E.A.; Shimon, L.J.; Shoham, Y.; Lamed, R. Cellulosomes—Structure and ultrastructure. J. Struct. Biol. 1998, 124, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Iyer, P.; Singhal, R.S. Production of glutaminase (EC 3.2.1.5) from Zygosaccharomyces rouxii: Statistical optimization using response surface methodology. Bioresour. Technol. 2008, 99, 4300–4307. [Google Scholar] [CrossRef] [PubMed]
- Iyer, P.V.; Singhal, R.S. Screening and selection of marine isolate for l-glutaminase production and media optimization using response surface methodology. Appl. Biochem. Biotechnol. 2009, 159, 233–250. [Google Scholar] [CrossRef] [PubMed]
- Iyer, P.V.; Singhal, R.S. Glutaminase Production using Zygosaccharomyces rouxii NRRL-Y 2547: Effect of aeration, agitation regimes and feeding strategies. Chem. Eng. Technol. 2010, 33, 52–62. [Google Scholar] [CrossRef]
- Lynd, L.R.; Weimer, P.J.; van Zyl, W.H.; Pretorius, I.S. Microbial cellulose utilization: Fundamentals and biotechnology. Microbiol. Mol. Biol. Rev. 2002, 66, 506–577. [Google Scholar] [CrossRef] [PubMed]
- Mg, Z.L.M.; Than, W.M.; Myint, M. Study on the cellulase enzyme producing activity of bacteria isolated from manure waste and degrading soil. Int. J. Tech. Res. Appl. 2015, 3, 165–169. [Google Scholar]
- Onsori, H.; Zamani, M.R.; Motallebi, M.; Zarghami, N. Identification of over producer strain of endo-β-1, 4-glucanase in Aspergillus species: Characterization of crude carboxymethyl cellulase. Afr. J. Biotechnol. 2005, 4, 26–30. [Google Scholar]
- Jansova, E.; Schwarzova, Z.; Chaloupka, J. Sporulation and synthesis of extracellular proteinases in Bacillus subtilis are more temperature-sensitive than growth. Folia Microbiol. 1993, 38, 22–24. [Google Scholar] [CrossRef]
- Bakare, M.K.; Adewale, I.O.; Ajayi, A.; Shonukan, O.O. Purification and characterization of cellulase from the wildtype and two improved mutants of Pseudomonas fluorescens. Afr. J. Biotechnol. 2005, 4, 898–904. [Google Scholar]
- Assareh, R.; Hossein, S.Z.; Noghabi, K.A.; Aminzadeh, S.; Khaniki, G.B. Characterization of the newly isolated Geobacillus sp. T1, the efficient cellulase-producer on untreated barley and wheat straws. Bioresour. Technol. 2012, 120, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Fattah, Y.R.; El-Helow, E.R.; Ghanem, K.M.; Lotfy, W.A. Application of factorial designs for optimization of avicelase production by a thermophilic Geobacillus isolate. Res. J. Microbiol. 2007, 2, 13–23. [Google Scholar]
- Monisha, R.; Uma, M.V.; Murthy, V.K. Partial purification and characterization of Bacillus pumilus xylanase from soil source. Kathmandu Univ. J. Sci. Eng. Technol. 2009, 5, 137–148. [Google Scholar]
- Sepahy, A.A.; Ghazi, S.; Sepahy, M.A. Cost-Effective production and optimization of alkaline xylanase by indigenous Bacillus mojavensis AG137 fermented on agricultural waste. Enzyme Res. 2011. [Google Scholar] [CrossRef]
- Simphiwe, P.; Ademola, B.; Olaniran, O.; Pillay, B. Sawdust and digestive bran as cheap alternate substrates for xylanase production. Afr. J. Microbiol. Res. 2011, 5, 742–752. [Google Scholar] [CrossRef]
- Okaiyeto, K.; Nwodo, U.U.; Okoli, S.A.; Mabinya, L.V.; Okoh, A.I. Implications for public health demands alternatives to inorganic and synthetic flocculants: Bioflocculants as important candidates. Microbiol. Open 2016, 5, 177–211. [Google Scholar] [CrossRef] [PubMed]
- Fatokun, E.N.; Nwodo, U.U.; Okoh, A.I. Classical optimization of cellulase and xylanase production by a marine Streptomyces species. Appl. Sci. 2016, 6. [Google Scholar] [CrossRef]
- Liang, Y.; Feng, Z.; Yesuf, J.; Blackburn, J.W. Optimization of growth medium and enzyme assay conditions for crude cellulases produced by a novel thermophilic and cellulolytic bacterium, Anoxybacillus sp. Appl. Biochem. Biotechnol. 2009. [Google Scholar] [CrossRef] [PubMed]
- Nasr, S.A.; Abozaid, A.A.; Hussein, N.A.; Al-Salemi, F.A. Cellulase production by local bacteria isolated from Taif in Saudi Arabia. J. Agric. Sci. 2011, 19, 163–170. [Google Scholar]
- Padilha, I.Q.M.; Carvalho, L.C.T.; Dias, P.V.S.; Grisi, T.C.S.L.; da Silva, F.L.H.; Santo, S.F.M.; Araujo, D.A.M. Production and characterization of thermophilic carboxymethylcellulase synthesized by Bacillus sp. growing on sugarcane bagasse in submerged fermentation. Braz. J. Chem. Eng. 2015, 32, 35–42. [Google Scholar] [CrossRef]
- Gessesse, A. Purification and properties of two thermostable alkaline xylanases from an alkaliphilic Bacillus sp. Appl. Environ. Microbiol. 1998, 64, 3533–3535. [Google Scholar] [PubMed]
- Rani, D.S.; Nand, K. Production of thermostable cellulase-free xylanase by Clostridium absonum CFR-702. Proc. Biochem. 2000, 36, 355–362. [Google Scholar] [CrossRef]
- Pereira, M.A.; Jacobs, D.R., Jr.; Van Horn, L.; Slattery, M.L.; Kartashov, A.I.; Ludwig, D.S. Dairy consumption, obesity, and the insulin resistance syndrome in young adults: The CARDIA Study. JAMA 2002, 287, 2081–2089. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Zeikus, J. Impact of electrode composition on electricity generation in a single-compartment fuel cell using Shewanella putrefaciens. Appl. Microbiol. Biotechnol. 2002, 59, 58–61. [Google Scholar] [PubMed]
- Bano, S.; Qader, S.A.U.; Aman, A.; Syed, M.N.; Durrani, K. High production of cellulose degrading endo-1, 4-β-D-glucanase using bagasse as a substrate from Bacillus subtilis KIBGE HAS. Carbohydr. Polym. 2013, 91, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Giavasis, I.; Harvey, L.M.; McNeil, B. The effect of agitation and aeration on the synthesis and molecular weight of gellan in batch cultures of Sphingomonas paucimobilis. Enzyme Microb. Technol. 2006, 38, 101–108. [Google Scholar] [CrossRef]
- El-Refai, H.A.; AbdElRahman, H.Y.; Abdulla, H.; Hanna, A.G.; Hashem, A.H.; El-Refai, A.H.; Ahmed, E.M. Studies on the production of actinomycin by Nocardioides luteus, a novel source. Curr. Trends Biotechnol. Pharm. 2011, 5, 1282–1297. [Google Scholar]
- Kokare, C.R.; Kadam, S.S.; Mahadik, K.R.; Chopade, B.A. Studies on bioemulsifier production from marine Streptomyces sp. S1. Indian J. Biotechnol. 2007, 6, 78–84. [Google Scholar]
- Nagendra, H.; Southworth, J.; Tucker, C. Accessibility as a determinant of landscape transformation in western Honduras: Linking pattern and process. Landsc. Ecol. 2003, 18, 141–158. [Google Scholar] [CrossRef]
- Ariffin, H.; Abdullah, N.; Umi Kalsom, M.S.; Shirai, Y.; Hassan, M.A. Production and characterisation of cellulase by Bacillus pumilus EB3. Int. J. Eng. Technol. 2006, 3, 47–53. [Google Scholar]
- Sharma, A.; Adhikari, S.; Satyanarayana, T. Alkali-Thermostable and cellulase-free xylanase production by an extreme thermophile Geobacillus thermoleovorans. World J. Microbiol. Biotechnol. 2007, 23, 483–490. [Google Scholar] [CrossRef]
- Mrudula, S.; Shyam, N. Immobilization of Bacillus megaterium MTCC 2444 by Ca-alginate entrapment method for enhanced alkaline protease production. Braz. Arch. Biol. Technol. 2012, 55, 135–144. [Google Scholar] [CrossRef]
- Gupta, U.; Kar, R. Xylanase production by a thermo-tolerant Bacillus species under solid-state and submerged fermentation. Braz. Arch. Biol. Technol. 2009, 52, 1363–1371. [Google Scholar] [CrossRef]
- Murugan, S.; Arnold, D.; Pongiya, U.D.; Narayanan, P.M. Production of xylanase from Arthrobacter sp. MTCC 6915 using saw dust as substrate under solid state fermentation. Enzyme Res. 2011, 7, 696942. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate Code | Halo Zone Diameter (mm ± SD) on CMC | Halo Zone Diameter (mm ± SD) on Xylan | Halo Zone Diameter of the Two Enzymes (mm ± SD) |
|---|---|---|---|
| PLY1 | 71 ± 7.5 | 43 ± 3.58 | 114 |
| BLY2 | 11.6 ± 2.7 | - | 11.6 |
| BLY10 | 11.4 ± 1.8 | - | 11.4 |
| BLC6 | 22.2 ± 2.4 | - | 22.2 |
| MLY10 | 72.7 ± 6.4 | 40.7 ± 4.7 | 113.4 |
| TLW8 | - | - | 0 |
| TLY1 | 12.5 ± 4.4 | - | 12.5 |
| TLY3 | 32.7 ± 4.7 | 58.7 ± 3.2 | 91.4 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mmango-Kaseke, Z.; Okaiyeto, K.; Nwodo, U.U.; Mabinya, L.V.; Okoh, A.I. Optimization of Cellulase and Xylanase Production by Micrococcus Species under Submerged Fermentation. Sustainability 2016, 8, 1168. https://doi.org/10.3390/su8111168
Mmango-Kaseke Z, Okaiyeto K, Nwodo UU, Mabinya LV, Okoh AI. Optimization of Cellulase and Xylanase Production by Micrococcus Species under Submerged Fermentation. Sustainability. 2016; 8(11):1168. https://doi.org/10.3390/su8111168
Chicago/Turabian StyleMmango-Kaseke, Ziyanda, Kunle Okaiyeto, Uchechukwu U. Nwodo, Leonard V. Mabinya, and Anthony I. Okoh. 2016. "Optimization of Cellulase and Xylanase Production by Micrococcus Species under Submerged Fermentation" Sustainability 8, no. 11: 1168. https://doi.org/10.3390/su8111168
APA StyleMmango-Kaseke, Z., Okaiyeto, K., Nwodo, U. U., Mabinya, L. V., & Okoh, A. I. (2016). Optimization of Cellulase and Xylanase Production by Micrococcus Species under Submerged Fermentation. Sustainability, 8(11), 1168. https://doi.org/10.3390/su8111168