



(Micro-)Plastics in Saturated and Unsaturated Groundwater Bodies: First Evidence of Presence in Groundwater Fauna and Habitats
 ,
,  ,
,  , , , , , , ,
, , , , , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sample Collection
2.3. Microplastic Extraction
2.4. Contamination Control
2.5. Polymer Analysis
2.6. Fluorescence Microscopy
3. Results
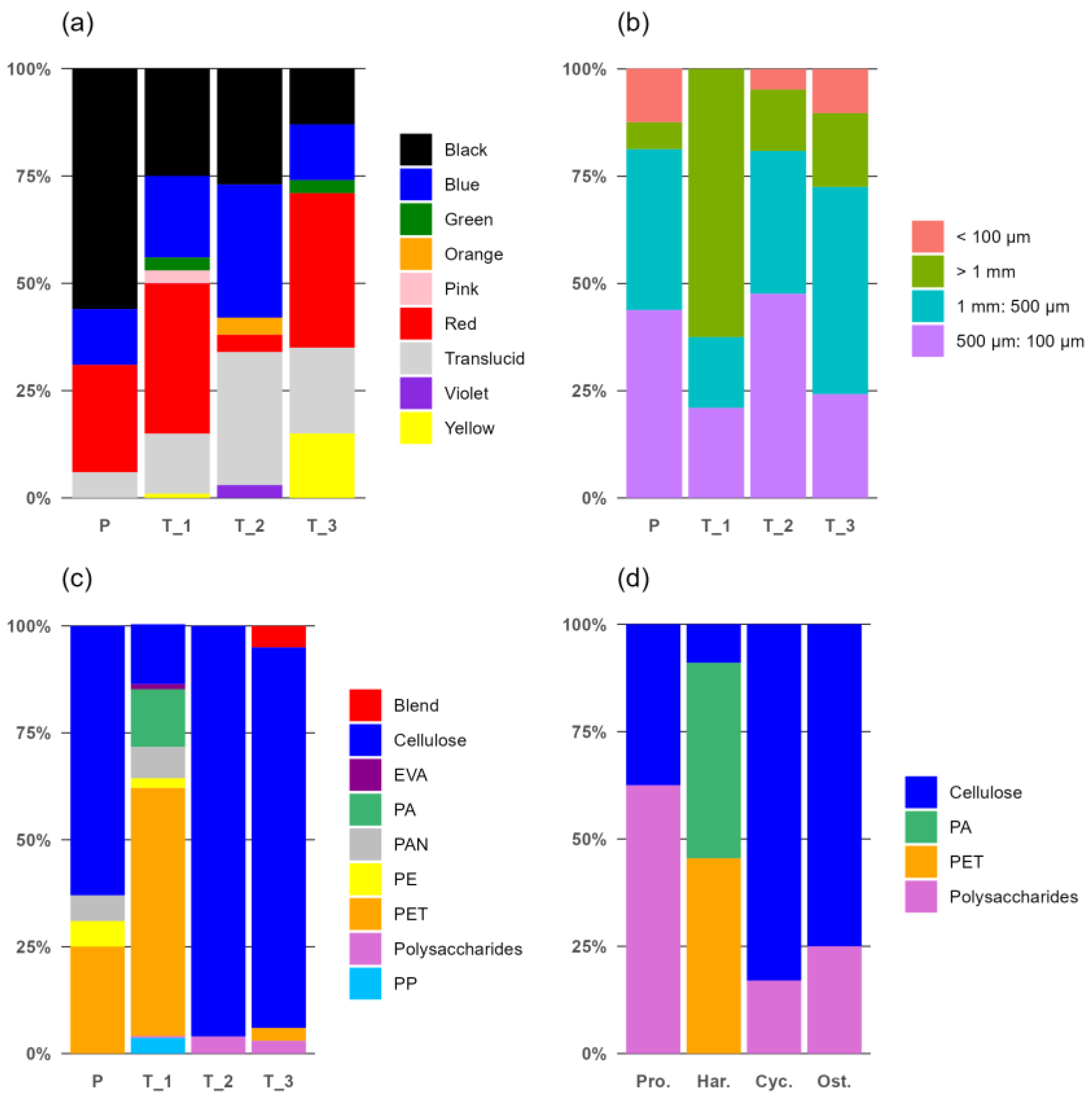
3.1. Groundwater Samples
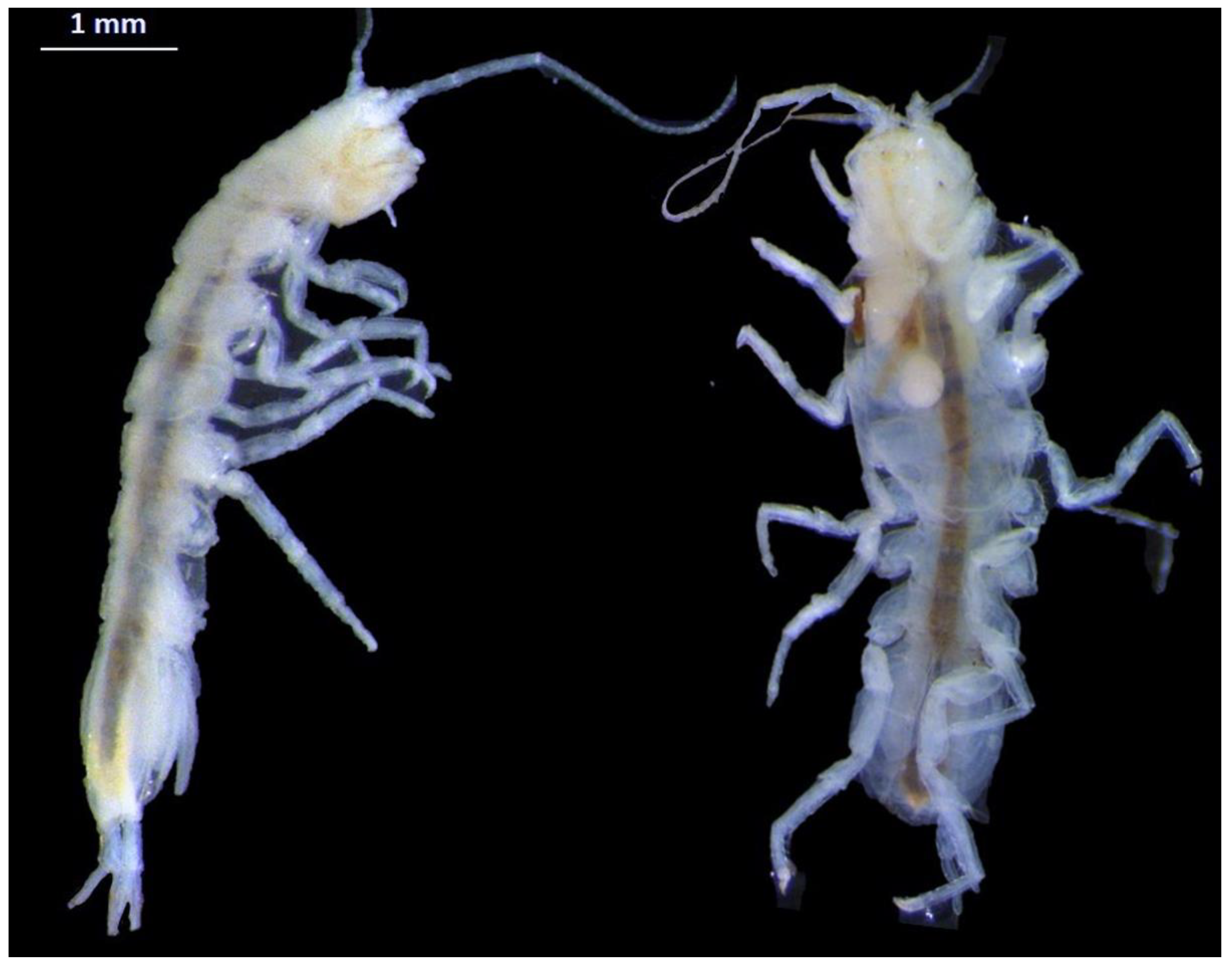
3.2. Fauna Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- PlasticsEurope; EPRO. Plastics—The Facts 2021 an Analysis of European Plastics Production, Demand and Waste Data; PlasticsEurope: Brussels, Belgium; EPRO: Wemmel, Belgium, 2021. [Google Scholar]
- Taylor, M.L.; Gwinnett, C.; Robinson, L.F.; Woodall, L.C. Plastic microfibre ingestion by deep-sea organisms. Sci. Rep. 2016, 6, 33997. [Google Scholar] [CrossRef]
- ISO/TR 21960:2020(en); Plastics—Environmental Aspects—State of Knowledge and Methodologies. 2020. Available online: https://www.iso.org/obp/ui/en/#iso:std:iso:tr:21960:ed-1:v1:en (accessed on 9 October 2023).
- Guerranti, C.; Martellini, T.; Perra, G.; Scopetani, C.; Cincinelli, A. Microplastics in cosmetics: Environmental issues and needs for global bans. Environ. Toxicol. Pharmacol. 2019, 68, 75–79. [Google Scholar] [CrossRef]
- UNEP. Plastic in Cosmetics: Are We Polluting the Environment through Our Personal Care? UNEP: Nairobi, Kenya, 2015. [Google Scholar]
- Cole, M.; Lindeque, P.; Halsband, C.; Galloway, T.S. Microplastics as contaminants in the marine environment: A review. Mar. Pollut. Bull. 2011, 62, 2588–2597. [Google Scholar] [CrossRef] [PubMed]
- Crawford, C.B.; Quinn, B. Microplastic Pollutants, 1st ed.; Elsevier Science: Amsterdam, The Netherlands, 2016; pp. 1–315. [Google Scholar]
- Zhang, K.; Hamidian, A.H.; Tubić, A.; Zhang, Y.; Fang, J.K.; Wu, C.; Lam, P.K. Understanding plastic degradation and microplastic formation in the environment: A review. Environ. Pollut. 2021, 274, 116554. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.; Allen, D.; Phoenix, V.R.; Le Roux, G.; Jiménez, P.D.; Simonneau, A.; Binet, S.; Galop, D. Atmospheric transport and deposition of microplastics in a remote mountain catchment. Nat. Geosci. 2019, 12, 339–344. [Google Scholar] [CrossRef]
- Sarma, H.; Hazarika, R.P.; Kumar, V.; Roy, A.; Pandit, S.; Prasad, R. Microplastics in marine and aquatic habitats: Sources, impact, and sustainable remediation approaches. Environ. Sustain. 2022, 5, 39–49. [Google Scholar] [CrossRef]
- Scopetani, C.; Chelazzi, D.; Cincinelli, A.; Martellini, T.; Leiniö, V.; Pellinen, J. Hazardous contaminants in plastics contained in compost and agricultural soil. Chemosphere 2022, 293, 133645. [Google Scholar] [CrossRef]
- Yang, H.; Chen, G.; Wang, J. Microplastics in the marine environment: Sources, fates, impacts and microbial degradation. Toxics 2021, 9, 41. [Google Scholar] [CrossRef]
- Cincinelli, A.; Scopetani, C.; Chelazzi, D.; Martellini, T.; Pogojeva, M.; Slobodnik, J. Microplastics in the Black Sea sediments. Sci. Total. Environ. 2020, 760, 143898. [Google Scholar] [CrossRef] [PubMed]
- Giarrizzo, T.; Andrade, M.C.; Schmid, K.; O Winemiller, K.; Ferreira, M.; Pegado, T.; Chelazzi, D.; Cincinelli, A.; Fearnside, P.M. Amazonia: The new frontier for plastic pollution. Front. Ecol. Environ. 2019, 17, 309–310. [Google Scholar] [CrossRef]
- Van Cauwenberghe, L.; Vanreusel, A.; Mees, J.; Janssen, C.R. Microplastic pollution in deep-sea sediments. Environ. Pollut. 2013, 182, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Mintenig, S.M.; Löder, M.G.J.; Primpke, S.; Gerdts, G. Low numbers of microplastics detected in drinking water from ground water sources. Sci. Total. Environ. 2019, 648, 631–635. [Google Scholar] [CrossRef] [PubMed]
- Panno, S.V.; Kelly, W.R.; Scott, J.; Zheng, W.; McNeish, R.E.; Holm, N.; Hoellein, T.J.; Baranski, E.L. Microplastic Contamination in Karst Groundwater Systems. Groundwater 2019, 57, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Samandra, S.; Johnston, J.M.; Jaeger, J.E.; Symons, B.; Xie, S.; Currell, M.; Ellis, A.V.; Clarke, B.O. Microplastic contamination of an unconfined groundwater aquifer in Victoria, Australia. Sci. Total. Environ. 2022, 802, 149727. [Google Scholar] [CrossRef] [PubMed]
- Re, V. Shedding light on the invisible: Addressing the potential for groundwater contamination by plastic microfibers. Hydrogeol. J. 2019, 27, 2719–2727. [Google Scholar] [CrossRef]
- Belkhiri, A.H.; Carre, F.; Quiot, F. State of knowledge and future research needs on microplastics in groundwater. J. Water Health 2022, 20, 1479–1496. [Google Scholar] [CrossRef]
- Severini, E.; Ducci, L.; Sutti, A.; Robottom, S.; Sutti, S.; Celico, F. River–Groundwater Interaction and Recharge Effects on Microplastics Contamination of Groundwater in Confined Alluvial Aquifers. Water 2022, 14, 1913. [Google Scholar] [CrossRef]
- Jiang, S.; Wang, J.; Wu, F.; Xu, S.; Liu, J.; Chen, J. Extensive abundances and characteristics of microplastic pollution in the karst hyporheic zones of urban rivers. Sci. Total. Environ. 2023, 857, 159616. [Google Scholar] [CrossRef]
- Bergami, E.; Manno, C.; Cappello, S.; Vannuccini, M.; Corsi, I. Nanoplastics affect moulting and faecal pellet sinking in Antarctic krill (Euphausia superba) juveniles. Environ. Int. 2020, 143, 105999. [Google Scholar] [CrossRef]
- Expósito, N.; Rovira, J.; Sierra, J.; Gimenez, G.; Domingo, J.L.; Schuhmacher, M. Levels of microplastics and their characteristics in molluscs from North-West Mediterranean Sea: Human intake. Mar. Pollut. Bull. 2002, 181, 113843. [Google Scholar] [CrossRef]
- Hirai, H.; Takada, H.; Ogata, Y.; Yamashita, R.; Mizukawa, K.; Saha, M.; Kwan, C.; Moore, C.; Gray, H.; Laursen, D.; et al. Organic micropollutants in marine plastics debris from the open ocean and remote and urban beaches. Mar. Pollut. Bull. 2011, 62, 1683–1692. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Lin, X.; Wu, M.; Lu, G.; Hao, Y.; Mo, C.; Li, Q.; Wu, J.; Wu, J.; Hu, B.X. Combined Effects of Polyamide Microplastics and Hydrochemical Factors on the Transport of Bisphenol A in Groundwater. Separations 2023, 10, 123. [Google Scholar] [CrossRef]
- Perestrelo, R.; Silva, C.L.; Algarra, M.; Câmara, J.S. Evaluation of the Occurrence of Phthalates in Plastic Materials Used in Food Packaging. Appl. Sci. 2021, 11, 2130. [Google Scholar] [CrossRef]
- Selvam, S.; Jesuraja, K.; Venkatramanan, S.; Roy, P.D.; Kumari, V.J. Hazardous microplastic characteristics and its role as a vector of heavy metal in groundwater and surface water of coastal south India. J. Hazard. Mater. 2021, 402, 123786. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, K.; Zhang, H. Adsorption of antibiotics on microplastics. Environ. Pollut. 2018, 237, 460–467. [Google Scholar] [CrossRef]
- Anbumani, S.; Kakkar, P. Ecotoxicological effects of microplastics on biota: A review. Environ. Sci. Pollut. Res. 2018, 25, 14373–14396. [Google Scholar] [CrossRef]
- Hahladakis, J.N.; Velis, C.A.; Weber, R.; Iacovidou, E.; Purnell, P. An overview of chemical additives present in plastics: Migration, release, fate and environmental impact during their use, disposal and recycling. J. Hazard. Mater. 2018, 344, 179–199. [Google Scholar] [CrossRef]
- Malard, F.; Griebler, C.; Rétaux, S. Groundwater Ecology and Evolution, 2nd ed.; Academic Press: Cambridge, MA, USA, 2023; pp. 1–610. [Google Scholar] [CrossRef]
- Rétaux, S.; Jeffery, W.R. Voices from the underground: Animal models for the study of trait evolution during groundwater colonization and adaptation. In Groundwater Ecology and Evolution, 2nd ed.; Malard, F., Griebler, C., Rétaux, S., Eds.; Academic Press: Cambridge, MA, USA, 2023; Chapter 12; pp. 285–304. [Google Scholar] [CrossRef]
- Cooper, S.; Fišer, C.; Zakšek, V.; Delić, T.; Borko, Š.; Faille, A.; Humphreys, W. Phylogenies reveal speciation dynamics: Case studies from groundwater. In Groundwater Ecology and Evolution, 2nd ed.; Malard, F., Griebler, C., Rétaux, S., Eds.; Academic Press: Cambridge, MA, USA, 2023; Chapter 7; pp. 165–183. [Google Scholar] [CrossRef]
- Zagmajster, M.; Ferreira, R.L.; Humphreys, W.F.; Niemiller, M.L.; Malard, F. Patterns and determinants of richness and composition of the groundwater fauna. In Groundwater Ecology and Evolution, 2nd ed.; Malard, F., Griebler, C., Rétaux, S., Eds.; Academic Press: Cambridge, MA, USA, 2023; Chapter 6; pp. 141–164. [Google Scholar] [CrossRef]
- Mermillod-Blondin, F.; Hose, G.C.; Simon, K.S.; Korbel, K.; Avramov, M.; Vorste, R. Vander. Role of invertebrates in groundwater ecosystem processes and services. In Groundwater Ecology and Evolution, 2nd ed.; Malard, F., Griebler, C., Rétaux, S., Eds.; Academic Press: Cambridge, MA, USA, 2023; Chapter 11; pp. 263–281. [Google Scholar] [CrossRef]
- Deng, L.; Krauss, S.; Feichtmayer, J.; Hofmann, R.; Arndt, H.; Griebler, C. Grazing of heterotrophic flagellates on viruses is driven by feeding behaviour. Environ. Microbiol. Rep. 2014, 6, 325–330. [Google Scholar] [CrossRef]
- de Sá, L.C.; Luís, L.G.; Guilhermino, L. Effects of microplastics on juveniles of the common goby (Pomatoschistus microps): Confusion with prey, reduction of the predatory performance and efficiency, and possible influence of developmental conditions. Environ. Pollut. 2015, 196, 359–362. [Google Scholar] [CrossRef]
- Jeong, C.-B.; Won, E.-J.; Kang, H.-M.; Lee, M.-C.; Hwang, D.-S.; Hwang, U.-K.; Zhou, B.; Souissi, S.; Lee, S.-J.; Lee, J.-S. Microplastic Size-Dependent Toxicity, Oxidative Stress Induction, and p-JNK and p-p38 Activation in the Monogonont Rotifer (Brachionus koreanus). Environ. Sci. Technol. 2016, 50, 8849–8857. [Google Scholar] [CrossRef]
- Banzato, C.; Dallagiovanna, G.; Maino, M.; Peano, G.; Vigna, B. Correlation between the geological setting and groundwater flow: The Bossea karst underground laboratory. Epitome 2011, 4, 14. [Google Scholar]
- Peano, G.; Vigna, B.; Villavecchia, E. L’evento alluvionale nell’ottobre 1996 nella Grotta di Bossea. Bossea 2005, 5–8, 407–422. [Google Scholar]
- Forti, P.; Gamberi, M.L. Le pisoliti della Buca del Vasaio di Montrone e l’ipotesi del minimo e massimo diametro possibile. Sotterranea 1982, 59, 18–23. [Google Scholar]
- Regione Toscana. Delibera n. 939 del 26/10/2009. Individuazione e Caratterizzazione dei Corpi Idrici Della Toscana—Attuazione delle Disposizioni di Cui ALL’ART.2 del DM 131/08 (Acque Superficiali) e Degli Art. 1,3 e all. 1 del D.Lgs. 30/09 (Acque Sotterranee). 2009. Available online: https://www.arpat.toscana.it/documentazione/normativa/normativa-regionale-toscana/2009/delibera-giunta-regionale-toscana-n.-939-del-26-10-2009 (accessed on 5 January 2024).
- Bencini, A.; Ercolanelli, R.; Sbaragli, A.; Verrucchi, C. Groundwaters of Florence (Italy): Trace element distribution and vul-nerability of the aquifers. Environl. Geol. 1993, 22, 193–200. [Google Scholar] [CrossRef]
- Koelmans, A.A.; Nor, N.H.M.; Hermsen, E.; Kooi, M.; Mintenig, S.M.; De France, J. Microplastics in freshwaters and drinking water: Critical review and assessment of data quality. Water Res. 2019, 155, 410–422. [Google Scholar] [CrossRef]
- Dris, R.; Imhof, H.; Sanchez, W.; Gasperi, J.; Galgani, F.; Tassin, B.; Laforsch, C. Beyond the ocean: Contamination of freshwater ecosystems with (micro-)plastic particles. Environ. Chem. 2015, 12, 539–550. [Google Scholar] [CrossRef]
- Santini, S.; De Beni, E.; Martellini, T.; Sarti, C.; Randazzo, D.; Ciraolo, R.; Scopetani, C.; Cincinelli, A. Occurrence of Natural and Synthetic Micro-Fibers in the Mediterranean Sea: A Review. Toxics 2022, 10, 391. [Google Scholar] [CrossRef]
- Castaño-Sánchez, A.; Malard, F.; Kalčíková, G.; Reboleira, A.S.P.S. Novel Protocol for Acute In Situ Ecotoxicity Test Using Native Crustaceans Applied to Groundwater Ecosystems. Water 2021, 13, 1132. [Google Scholar] [CrossRef]
- Malard, F.; Bernard, C.; Lyon, U.; Claude, M.-J.D.-O.; Lyon, B.U.; Stoch, F. Sampling Manual for the Assessment of Regional Groundwater Biodiversity. 2002. Available online: https://www.researchgate.net/publication/267567541 (accessed on 12 January 2024).
- Korbel, K.; Chariton, A.; Stephenson, S.; Greenfield, P.; Hose, G.C. Wells provide a distorted view of life in the aquifer: Implications for sampling, monitoring and assessment of groundwater ecosystems. Sci. Rep. 2017, 7, 40702. [Google Scholar] [CrossRef]
- Reiss, J.; Schmid-Araya, J.M. Existing in plenty: Abundance, biomass and diversity of ciliates and meiofauna in small streams. Freshw. Biol. 2008, 53, 652–668. [Google Scholar] [CrossRef]
- Cummins, K.W.; Wilzbach, M.; Kolouch, B.; Merritt, R. Estimating Macroinvertebrate Biomass for Stream Ecosystem Assessments. Int. J. Environ. Res. Public Health 2022, 19, 3240. [Google Scholar] [CrossRef]
- Culver, D.C.; Pipan, T.; Fišer, Ž. Ecological and evolutionary jargon in subterranean biology. In Groundwater Ecology and Evolution, 2nd ed.; Malard, F., Griebler, C., Rétaux, S., Eds.; Academic Press: Cambridge, MA, USA, 2023; Chapter 4; pp. 89–100. [Google Scholar] [CrossRef]
- Venarsky, M.; Niemiller, M.L.; Fišer, C.; Saclier, N.; Moldovan, O.T. Life histories in groundwater organisms. In Groundwater Ecology and Evolution, 2nd ed.; Malard, F., Griebler, C., Rétaux, S., Eds.; Academic Press: Cambridge, MA, USA, 2023; Chapter 19; pp. 439–456. [Google Scholar] [CrossRef]
- An, X.; Li, W.; Lan, J.; Adnan, M. Preliminary Study on the Distribution, Source, and Ecological Risk of Typical Microplastics in Karst Groundwater in Guizhou Province, China. Int. J. Environ. Res. Public Health 2022, 19, 14751. [Google Scholar] [CrossRef]
- Cutroneo, L.; Reboa, A.; Geneselli, I.; Capello, M. Considerations on salts used for density separation in the extraction of microplastics from sediments. Mar. Pollut. Bull. 2021, 166, 112216. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Cabigliera, S.B.; Martellini, T.; Laurati, M.; Chelazzi, D.; Galassi, D.M.P.; Cincinelli, A. Ingestion of microplastics and textile cellulose particles by some meiofaunal taxa of an urban stream. Chemosphere 2023, 310, 136830. [Google Scholar] [CrossRef]
- ISO 6330:2021; Textiles—Domestic Washing and Drying Procedures for Textile Testing. 2021. Available online: https://www.iso.org/standard/75934.html (accessed on 20 February 2024).
- Alfonso, M.B.; Takashima, K.; Yamaguchi, S.; Tanaka, M.; Isobe, A. Microplastics on plankton samples: Multiple digestion techniques assessment based on weight, size, and FTIR spectroscopy analyses. Mar. Pollut. Bull. 2021, 173, 113027. [Google Scholar] [CrossRef]
- Prata, J.C.; Sequeira, I.F.; Monteiro, S.S.; Silva, A.L.P.; da Costa, J.P.; Dias-Pereira, P.; Fernandes, A.J.S.; da Costa, F.M.; Duarte, A.C.; Rocha-Santos, T. Preparation of biological samples for microplastic identification by Nile Red. Sci. Total. Environ. 2021, 783, 147065. [Google Scholar] [CrossRef]
- Scopetani, C.; Esterhuizen-Londt, M.; Chelazzi, D.; Cincinelli, A.; Setälä, H.; Pflugmacher, S. Self-contamination from clothing in microplastics research. Ecotoxicol. Environ. Saf. 2020, 189, 110036. [Google Scholar] [CrossRef] [PubMed]
- Brander, S.M.; Renick, V.C.; Foley, M.M.; Steele, C.; Woo, M.; Lusher, A.; Carr, S.; Helm, P.; Box, C.; Cherniak, S.; et al. Sampling and Quality Assurance and Quality Control: A Guide for Scientists Investigating the Occurrence of Microplastics Across Matrices. Appl. Spectrosc. 2020, 74, 1099–1125. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Lusher, A.L.; Bråte, I.L.N.; Munno, K.; Hurley, R.R.; Welden, N.A. Is It or Isn’t It: The Importance of Visual Classification in Microplastic Characterization. Appl. Spectrosc. 2020, 74, 1139–1153. [Google Scholar] [CrossRef] [PubMed]
- UNESCO. Groundwater: Making the invisible visible. The United Nations World Water Development Report 2022. 2022. Available online: https://www.unwater.org/publications/un-world-water-development-report-2022 (accessed on 18 December 2023).
- Esfandiari, A.; Abbasi, S.; Peely, A.B.; Mowla, D.; Ghanbarian, M.A.; Oleszczuk, P.; Turner, A. Distribution and transport of microplastics in groundwater (Shiraz aquifer, southwest Iran). Water Res. 2022, 220, 118622. [Google Scholar] [CrossRef]
- Lapworth, D.J.; Baran, N.; Stuart, M.E.; Ward, R.S. Emerging organic contaminants in groundwater: A review of sources, fate and occurrence. Environ. Pollut. 2012, 163, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Baraza, T.; Hasenmueller, E.A. Floods enhance the abundance and diversity of anthropogenic microparticles (including microplastics and treated cellulose) transported through karst systems. Water Res. 2023, 242, 120204. [Google Scholar] [CrossRef]
- Balestra, V.; Vigna, B.; De Costanzo, S.; Bellopede, R. Preliminary investigations of microplastic pollution in karst systems, from surface watercourses to cave waters. J. Contam. Hydrol. 2023, 252, 104117. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Gui, X.; Xu, X.; Zhao, L.; Qiu, H.; Cao, X. Microplastics in the soil-groundwater environment: Aging, migration, and co-transport of contaminants—A critical review. J. Hazard. Mater. 2021, 419, 126455. [Google Scholar] [CrossRef]
- Viaroli, S.; Lancia, M.; Re, V. Microplastics contamination of groundwater: Current evidence and future perspectives. A review. Sci. Total. Environ. 2022, 824, 153851. [Google Scholar] [CrossRef]
- Browne, M.A.; Crump, P.; Niven, S.J.; Teuten, E.; Tonkin, A.; Galloway, T.; Thompson, R. Accumulation of microplastic on shorelines worldwide: Sources and sinks. Environ. Sci. Technol. 2011, 45, 9175–9179. [Google Scholar] [CrossRef]
- De Falco, F.; Cocca, M.; Avella, M.; Thompson, R.C. Microfiber Release to Water, Via Laundering, and to Air, via Everyday Use: A Comparison between Polyester Clothing with Differing Textile Parameters. Environ. Sci. Technol. 2020, 54, 3288–3296. [Google Scholar] [CrossRef]
- Schuyler, Q.; Hardesty, B.D.; Wilcox, C.; Townsend, K. To Eat or not to eat? debris selectivity by marine turtles. PLoS ONE 2012, 7, e40884. [Google Scholar] [CrossRef]
- Scherer, C.; Brennholt, N.; Reifferscheid, G.; Wagner, M. Feeding type and development drive the ingestion of microplastics by freshwater invertebrates. Sci. Rep. 2017, 7, 17006. [Google Scholar] [CrossRef]
- Wright, S.L.; Thompson, R.C.; Galloway, T.S. The physical impacts of microplastics on marine organisms: A review. Environ. Pollut. 2013, 178, 483–492. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Reboleira, A.S.P.S.; Galassi, D.M.P.; Hervant, F.; Avramov, M.; Iepure, S.G.; Mammola, S. Physiological tolerance and ecotoxicological constraints of groundwater fauna. In Groundwater Ecology and Evolution, 2nd ed.; Malard, F., Griebler, C., Rétaux, S., Eds.; Academic Press: Cambridge, MA, USA, 2023; Chapter 20; pp. 457–479. [Google Scholar] [CrossRef]
- Pan, C.-G.; Mintenig, S.M.; Redondo-Hasselerharm, P.E.; Neijenhuis, P.H.M.W.; Yu, K.-F.; Wang, Y.-H.; Koelmans, A.A. Automated μFTIR Imaging Demonstrates Taxon-Specific and Selective Uptake of Microplastic by Freshwater Invertebrates. Environ. Sci. Technol. 2021, 55, 9916–9925. [Google Scholar] [CrossRef] [PubMed]
- Balestra, V.; Bellopede, R. Microplastic pollution in show cave sediments: First evidence and detection technique. Environ. Pollut. 2022, 292, 118261. [Google Scholar] [CrossRef]
- Liu, S.; Huang, Y.; Luo, D.; Wang, X.; Wang, Z.; Ji, X.; Chen, Z.; Dahlgren, R.A.; Zhang, M.; Shang, X. Integrated effects of polymer type, size and shape on the sinking dynamics of biofouled microplastics. Water Res. 2022, 220, 118656. [Google Scholar] [CrossRef]
- Haegerbaeumer, A.; Mueller, M.-T.; Fueser, H.; Traunspurger, W. Impacts of micro- and nano-sized plastic particles on benthic invertebrates: A literature review and gap analysis. Front. Environ. Sci. 2019, 7, 425457. [Google Scholar] [CrossRef]
- Vroom, R.J.; Koelmans, A.A.; Besseling, E.; Halsband, C. Aging of microplastics promotes their ingestion by marine zooplankton. Environ. Pollut. 2017, 231, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, D.; Bréchon, A.; Thompson, R. Ingestion and fragmentation of plastic carrier bags by the amphipod Orchestia gammarellus: Effects of plastic type and fouling load. Mar. Pollut. Bull. 2018, 127, 154–159. [Google Scholar] [CrossRef]
- Powell, M.D.; Berry, A. Ingestion and regurgitation of living and inert materials by the estuarine copepod Eurytemora affinis (Poppe) and the influence of salinity. Estuar. Coast. Shelf Sci. 1990, 31, 763–773. [Google Scholar] [CrossRef]
- Xu, B.; Liu, S.; Zhou, J.L.; Zheng, C.; Weifeng, J.; Chen, B.; Zhang, T.; Qiu, W. PFAS and their substitutes in groundwater: Occurrence, transformation and remediation. J. Hazard. Mater. 2021, 412, 125159. [Google Scholar] [CrossRef]
- Zhang, S.; Wu, H.; Hou, J. Progress on the Effects of Microplastics on Aquatic Crustaceans: A Review. Int. J. Mol. Sci. 2023, 24, 5523. [Google Scholar] [CrossRef]
- Batel, A.; Linti, F.; Scherer, M.; Erdinger, L.; Braunbeck, T. Transfer of benzo[a]pyrene from microplastics to Artemia nauplii and further to zebrafish via a trophic food web experiment: CYP1A induction and visual tracking of persistent organic pollutants. Environ. Toxicol. Chem. 2016, 35, 1656–1666. [Google Scholar] [CrossRef]
- Beckie, R.D. Groundwater. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar] [CrossRef]
- Saccò, M.; Mammola, S.; Altermatt, F.; Alther, R.; Bolpagni, R.; Brancelj, A.; Brankovits, D.; Fišer, C.; Gerovasileiou, V.; Griebler, C.; et al. Groundwater is a hidden global keystone ecosystem. Glob. Chang. Biol. 2024, 30, e17066. [Google Scholar] [CrossRef]
- Garside, P.; Wyeth, P. Identification of Cellulosic Fibres by FTIR Spectroscopy: Thread and Single Fibre Analysis. Stud. Conserv. 2003, 48, 269–275. [Google Scholar] [CrossRef]
- Jung, M.R.; Horgen, F.D.; Orski, S.V.; Rodriguez, C.V.; Beers, K.L.; Balazs, G.H.; Jones, T.T.; Work, T.M.; Brignac, K.C.; Royer, S.J.; et al. Validation of ATR FT-IR to identify polymers of plastic marine debris, including those ingested by marine organisms. Mar. Pollut. Bull. 2018, 127, 704–716. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, Y.; Dang, Y.; Jiao, Q.; Li, H.; Wu, Q.; Zhao, Y. Synthesis of a novel titanium complex catalyst and its catalytic performance for olefin polymerization. Russ. J. Appl. Chem. 2015, 88, 1723–1727. [Google Scholar] [CrossRef]
- Faria, S.; Petkowicz, C.L.D.O.; de Morais, S.A.L.; Terrones, M.G.H.; de Resende, M.M.; de França, F.P.; Cardoso, V.L. Characterization of xanthan gum produced from sugar cane broth. Carbohydr. Polym. 2011, 86, 469–476. [Google Scholar] [CrossRef]
- Hong, T.; Yin, J.-Y.; Nie, S.-P.; Xie, M.-Y. Applications of infrared spectroscopy in polysaccharide structural analysis: Progress, challenge and perspective. Food Chem. 2012, 12, 100168. [Google Scholar] [CrossRef] [PubMed]
- Mizher, R.M.; Adawiya J, H.; Naser, J.Z.; Wee, T.T.; Zaki B, A.R.M.; Bin, K.A. Synthesis and characterization of grafted Acrylonitrile on Polystyrene modified with carbon nanotubes using Gamma-irradiation. Res. J. Chem. Sci. 2012, 2, 790–795. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Abundance | Fibers | Fragments |
|---|---|---|---|
| % | % | ||
| P | 64 | 88 | 12 |
| T_1 | 911 | 90 | 10 |
| T_2 | 18 | 94 | 6 |
| T_3 | 28 | 93 | 7 |
| Taxonomic Pool | Site | µ_L (mm) | SD_L (mm) | µ_W (mm) | SD_W (mm) | µ_DW (µg) | SD_DW (µg) | TOT DW (µg) | Items/ind. | Items/µg (TOT DW) |
|---|---|---|---|---|---|---|---|---|---|---|
| Proasellus franciscoloi (n = 1) | P | 7.095 | 1.656 | 1,032 | 191 | 0.2 | ||||
| Harpacticoida (n = 15) | T_1 | 0.468 | 0.120 | 0.095 | 0.013 | 0.274 | 0.125 | 4 | 35 | 130.5 |
| Cyclopoida (n = 6) | T_2 | 0.916 | 0.138 | 0.214 | 0.038 | 2.358 | 1.023 | 14 | 112 | 47.6 |
| Ostracoda (n = 19) | T_3 | 0.562 | 0.070 | 0.334 | 0.046 | 3.160 | 1.251 | 62 | 17 | 5.3 |
| Taxonomic Pool | Pellet | Fiber | Fragment | ||||||
|---|---|---|---|---|---|---|---|---|---|
| % | µ | SD | % | µ | SD | % | µ | SD | |
| Proasellus franciscoloi (n = 1) | 93 | 1 | 1 | 1 | 16 | 3 | 6 | 32 | 37 |
| Harpacticoida (n = 15) | 85 | 1 | 1 | 4 | 16 | 9 | 11 | 25 | 34 |
| Cyclopoida (n = 6) | 75 | 1 | 1 | 4 | 24 | 19 | 21 | 22 | 34 |
| Ostracoda (n = 19) | 87 | 2 | 2 | 3 | 21 | 10 | 10 | 23 | 14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sforzi, L.; Tabilio Di Camillo, A.; Di Lorenzo, T.; Galassi, D.M.P.; Balestra, V.; Piccini, L.; Cabigliera, S.B.; Ciattini, S.; Laurati, M.; Chelazzi, D.; et al. (Micro-)Plastics in Saturated and Unsaturated Groundwater Bodies: First Evidence of Presence in Groundwater Fauna and Habitats. Sustainability 2024, 16, 2532. https://doi.org/10.3390/su16062532
Sforzi L, Tabilio Di Camillo A, Di Lorenzo T, Galassi DMP, Balestra V, Piccini L, Cabigliera SB, Ciattini S, Laurati M, Chelazzi D, et al. (Micro-)Plastics in Saturated and Unsaturated Groundwater Bodies: First Evidence of Presence in Groundwater Fauna and Habitats. Sustainability. 2024; 16(6):2532. https://doi.org/10.3390/su16062532
Chicago/Turabian StyleSforzi, Laura, Agostina Tabilio Di Camillo, Tiziana Di Lorenzo, Diana Maria Paola Galassi, Valentina Balestra, Leonardo Piccini, Serena Benedetta Cabigliera, Samuele Ciattini, Marco Laurati, David Chelazzi, and et al. 2024. "(Micro-)Plastics in Saturated and Unsaturated Groundwater Bodies: First Evidence of Presence in Groundwater Fauna and Habitats" Sustainability 16, no. 6: 2532. https://doi.org/10.3390/su16062532
APA StyleSforzi, L., Tabilio Di Camillo, A., Di Lorenzo, T., Galassi, D. M. P., Balestra, V., Piccini, L., Cabigliera, S. B., Ciattini, S., Laurati, M., Chelazzi, D., Martellini, T., & Cincinelli, A. (2024). (Micro-)Plastics in Saturated and Unsaturated Groundwater Bodies: First Evidence of Presence in Groundwater Fauna and Habitats. Sustainability, 16(6), 2532. https://doi.org/10.3390/su16062532