
Predicting Changes in and Future Distributions of Plant Habitats of Climate-Sensitive Biological Indicator Species in South Korea



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Setting
2.2. Data Collection and Analysis
2.2.1. Species Data
2.2.2. Environmental Data
2.2.3. MaxEnt Analysis
3. Results
3.1. Model Evaluation and Current Possible Habitats
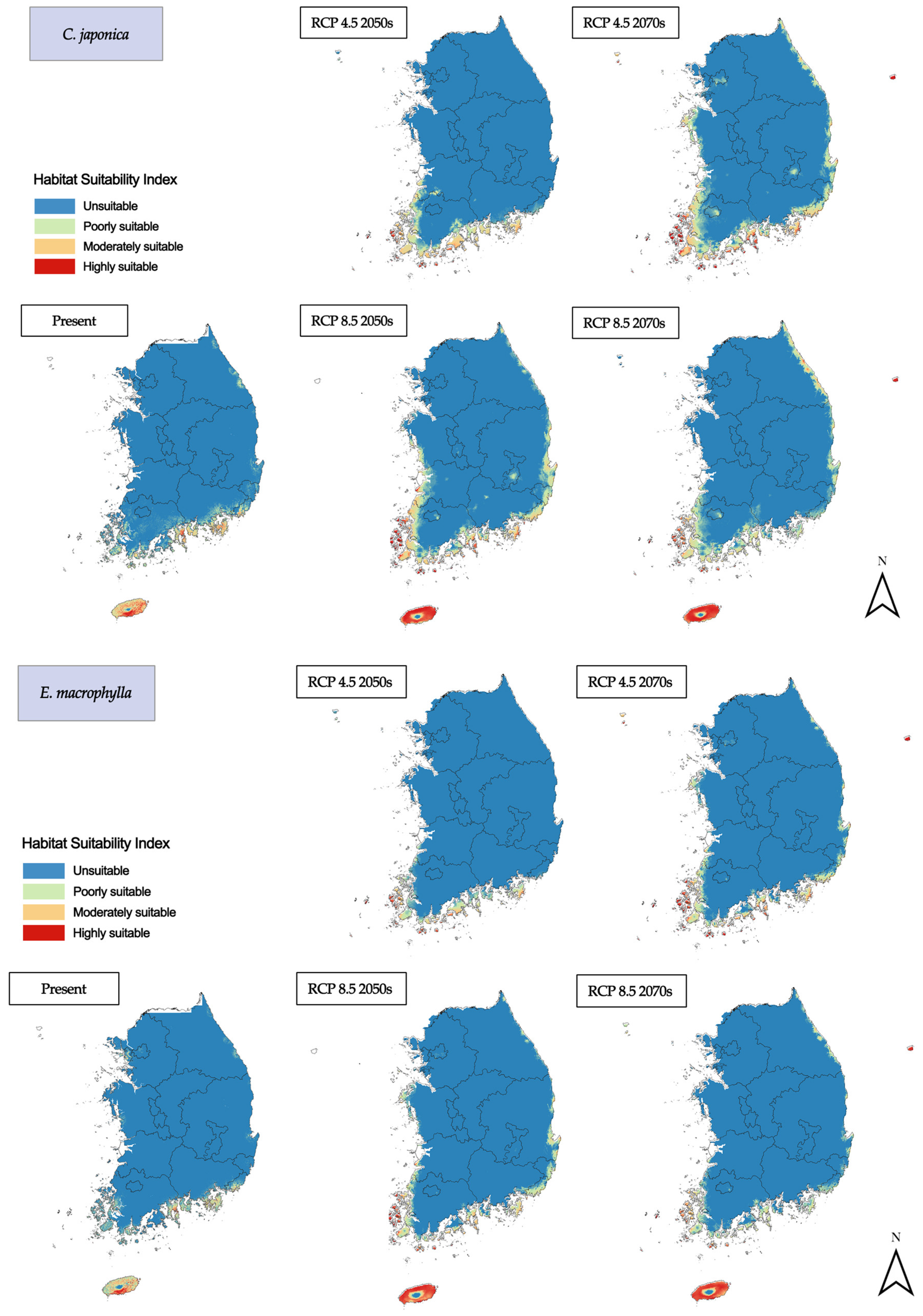
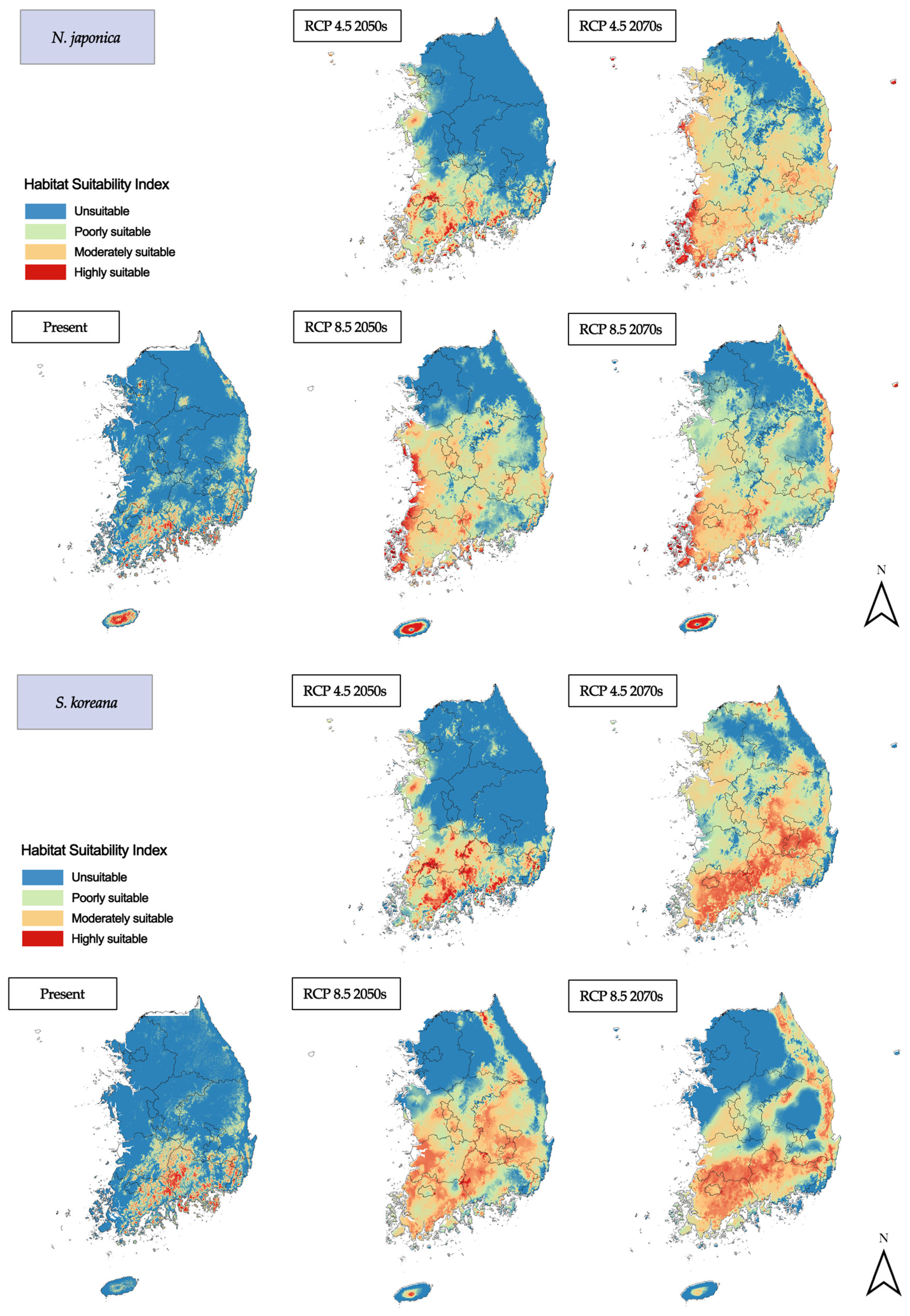
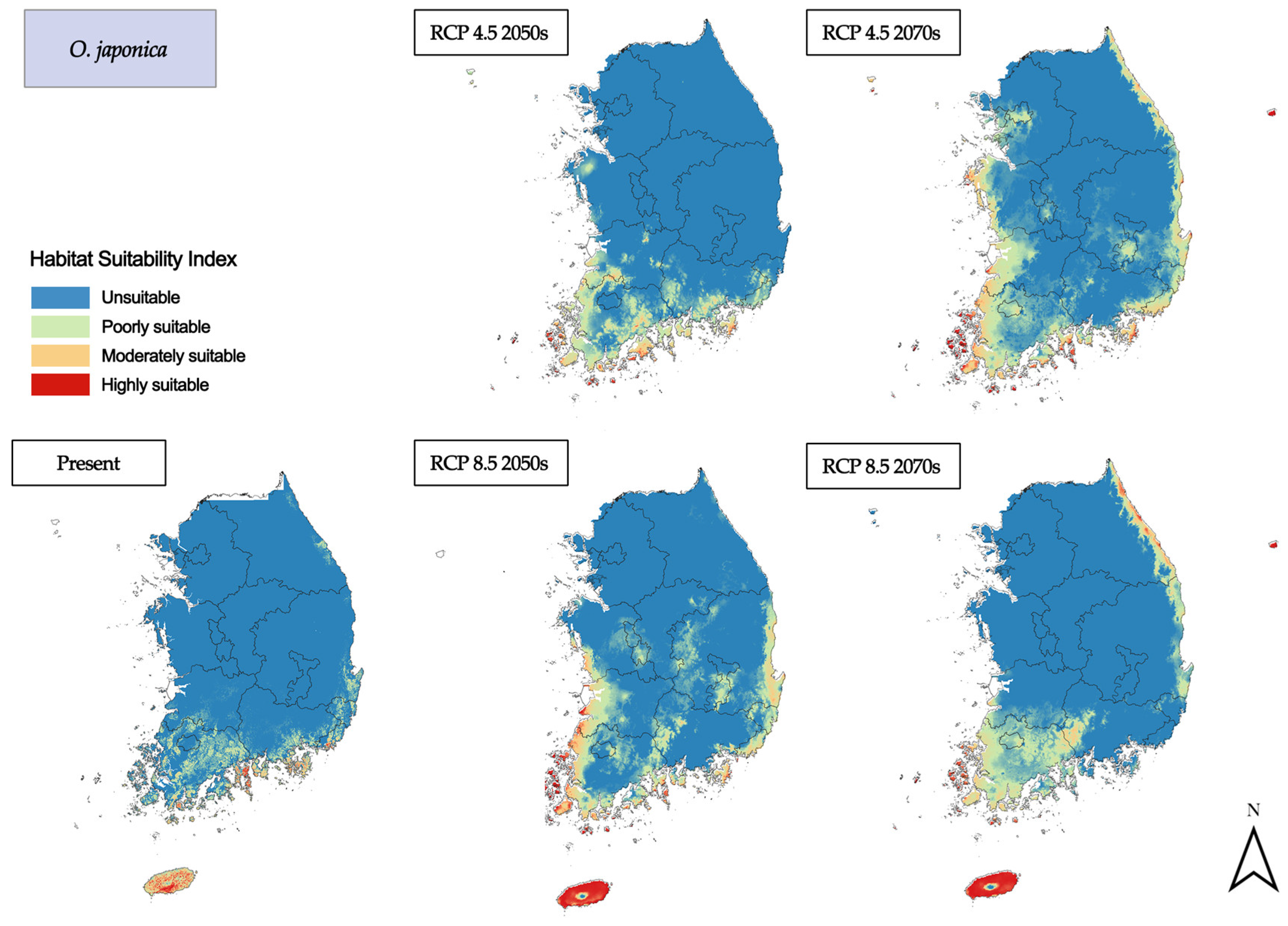
3.2. Habitat Changes under Future Climate Scenarios
3.2.1. Evergreen Coniferous/Broadleaf Trees
3.2.2. Deciduous Broadleaf Trees
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sales, L.P.; Galetti, M.; Pires, M.M. Climate and Land-use Change Will Lead to a Faunal “Savannization” on Tropical Rainforests. Glob. Chang. Biol. 2020, 26, 7036–7044. [Google Scholar] [CrossRef]
- Dornelas, M.; Gotelli, N.J.; Shimadzu, H.; Moyes, F.; Magurran, A.E.; McGill, B.J. A Balance of Winners and Losers in the Anthropocene. Ecol. Lett. 2019, 22, 847–854. [Google Scholar] [CrossRef]
- Urban, M.C. Accelerating Extinction Risk from Climate Change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef] [PubMed]
- Vincent, H.; Bornand, C.N.; Kempel, A.; Fischer, M. Rare Species Perform Worse than Widespread Species under Changed Climate. Biol. Conserv. 2020, 246, 108586. [Google Scholar] [CrossRef]
- Silva, J.L.S.E.; Cruz-Neto, O.; Peres, C.A.; Tabarelli, M.; Lopes, A.V. Climate Change Will Reduce Suitable Caatinga Dry Forest Habitat for Endemic Plants with Disproportionate Impacts on Specialized Reproductive Strategies. PLoS ONE 2019, 14, e0217028. [Google Scholar] [CrossRef] [PubMed]
- Franks, S.J.; Weber, J.J.; Aitken, S.N. Evolutionary and Plastic Responses to Climate Change in Terrestrial Plant Populations. Evol. Appl. 2014, 7, 123–139. [Google Scholar] [CrossRef]
- Vitasse, Y.; Ursenbacher, S.; Klein, G.; Bohnenstengel, T.; Chittaro, Y.; Delestrade, A.; Monnerat, C.; Rebetez, M.; Rixen, C.; Strebel, N.; et al. Phenological and Elevational Shifts of Plants, Animals and Fungi under Climate Change in the European Alps. Biol. Rev. 2021, 96, 1816–1835. [Google Scholar] [CrossRef]
- Araújo, M.B.; Alagador, D.; Cabeza, M.; Nogués-Bravo, D.; Thuiller, W. Climate Change Threatens European Conservation Areas: Climate Change Threatens Conservation Areas. Ecol. Lett. 2011, 14, 484–492. [Google Scholar] [CrossRef]
- Wang, C.; Liu, C.; Wan, J.; Zhang, Z. Climate Change May Threaten Habitat Suitability of Threatened Plant Species within Chinese Nature Reserves. PeerJ 2016, 4, e2091. [Google Scholar] [CrossRef]
- Prober, S.M.; Doerr, V.A.J.; Broadhurst, L.M.; Williams, K.J.; Dickson, F. Shifting the Conservation Paradigm: A Synthesis of Options for Renovating Nature under Climate Change. Ecol. Monogr. 2019, 89, e01333. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A Statistical Explanation of MaxEnt for Ecologists: Statistical Explanation of MaxEnt. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Manish, K.; Telwala, Y.; Nautiyal, D.C.; Pandit, M.K. Modelling the Impacts of Future Climate Change on Plant Communities in the Himalaya: A Case Study from Eastern Himalaya, India. Model. Earth Syst. Environ. 2016, 2, 92. [Google Scholar] [CrossRef]
- Edalat, M.; Dastres, E.; Jahangiri, E.; Moayedi, G.; Zamani, A.; Pourghasemi, H.R.; Tiefenbacher, J.P. Spatial Mapping Zataria multiflora Using Different Machine-Learning Algorithms. CATENA 2022, 212, 106007. [Google Scholar] [CrossRef]
- Pouteau, R.; Meyer, J.-Y.; Taputuarai, R.; Stoll, B. Support Vector Machines to Map Rare and Endangered Native Plants in Pacific Islands Forests. Ecol. Inform. 2012, 9, 37–46. [Google Scholar] [CrossRef]
- Sharifipour, B.; Gholinejad, B.; Shirzadi, A.; Shahabi, H.; Al-Ansari, N.; Farajollahi, A.; Mansorypour, F.; Clague, J.J. Rangeland Species Potential Mapping Using Machine Learning Algorithms. Ecol. Eng. 2023, 189, 106900. [Google Scholar] [CrossRef]
- Sittaro, F.; Hutengs, C.; Vohland, M. Which Factors Determine the Invasion of Plant Species? Machine Learning Based Habitat Modelling Integrating Environmental Factors and Climate Scenarios. Int. J. Appl. Earth Obs. Geoinf. 2023, 116, 103158. [Google Scholar] [CrossRef]
- Mosebo Fernandes, A.C.; Quintero Gonzalez, R.; Lenihan-Clarke, M.A.; Leslie Trotter, E.F.; Jokar Arsanjani, J. Machine Learning for Conservation Planning in a Changing Climate. Sustainability 2020, 12, 7657. [Google Scholar] [CrossRef]
- Shen, T.; Yu, H.; Wang, Y.-Z. Assessing the Impacts of Climate Change and Habitat Suitability on the Distribution and Quality of Medicinal Plant Using Multiple Information Integration: Take Gentiana rigescens as an Example. Ecol. Indic. 2021, 123, 107376. [Google Scholar] [CrossRef]
- Atzeni, L.; Cushman, S.A.; Bai, D.; Wang, J.; Chen, P.; Shi, K.; Riordan, P. Meta-replication, Sampling Bias, and Multi-scale Model Selection: A Case Study on Snow Leopard (Panthera uncia) in Western China. Ecol. Evol. 2020, 10, 7686–7712. [Google Scholar] [CrossRef]
- Fyllas, N.M.; Koufaki, T.; Sazeides, C.I.; Spyroglou, G.; Theodorou, K. Potential Impacts of Climate Change on the Habitat Suitability of the Dominant Tree Species in Greece. Plants 2022, 11, 1616. [Google Scholar] [CrossRef]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt Modeling to Predict the Potential Distribution of the Endemic Plant Rosa arabica Crép. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Mudereri, B.T.; Mukanga, C.; Mupfiga, E.T.; Gwatirisa, C.; Kimathi, E.; Chitata, T. Analysis of Potentially Suitable Habitat within Migration Connections of an Intra-African Migrant-the Blue Swallow (Hirundo Atrocaerulea). Ecol. Inform. 2020, 57, 101082. [Google Scholar] [CrossRef]
- Grimmett, L.; Whitsed, R.; Horta, A. Presence-Only Species Distribution Models Are Sensitive to Sample Prevalence: Evaluating Models Using Spatial Prediction Stability and Accuracy Metrics. Ecol. Model. 2020, 431, 109194. [Google Scholar] [CrossRef]
- Soilhi, Z.; Sayari, N.; Benalouache, N.; Mekki, M. Predicting Current and Future Distributions of Mentha pulegium L. in Tunisia under Climate Change Conditions, Using the MaxEnt Model. Ecol. Inform. 2022, 68, 101533. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of Species Distributions with Maxent: New Extensions and a Comprehensive Evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Kim, J.; Lee, D.K.; Kim, H.G. Suitable Trees for Urban Landscapes in the Republic of Korea under Climate Change. Landsc. Urban Plan. 2020, 204, 103937. [Google Scholar] [CrossRef]
- Bedia, J.; Herrera, S.; Gutiérrez, J.M. Dangers of Using Global Bioclimatic Datasets for Ecological Niche Modeling. Limitations for Future Climate Projections. Glob. Planet. Chang. 2013, 107, 1–12. [Google Scholar] [CrossRef]
- Cramer, W.; Bondeau, A.; Woodward, F.I.; Prentice, I.C.; Betts, R.A.; Brovkin, V.; Cox, P.M.; Fisher, V.; Foley, J.A.; Friend, A.D.; et al. Global Response of Terrestrial Ecosystem Structure and Function to CO2 and Climate Change: Results from Six Dynamic Global Vegetation Models. Glob. Chang. Biol. 2001, 7, 357–373. [Google Scholar] [CrossRef]
- Park, C.-E.; Jeong, S.-J.; Ho, C.-H.; Kim, J. Regional Variations in Potential Plant Habitat Changes in Response to Multiple Global Warming Scenarios. J. Clim. 2015, 28, 2884–2899. [Google Scholar] [CrossRef]
- Sitch, S.; Huntingford, C.; Gedney, N.; Levy, P.E.; Lomas, M.; Piao, S.L.; Betts, R.; Ciais, P.; Cox, P.; Friedlingstein, P.; et al. Evaluation of the Terrestrial Carbon Cycle, Future Plant Geography and Climate-carbon Cycle Feedbacks Using Five Dynamic Global Vegetation Models (DGVMs). Glob. Chang. Biol. 2008, 14, 2015–2039. [Google Scholar] [CrossRef]
- Lucht, W.; Schaphoff, S.; Erbrecht, T.; Heyder, U.; Cramer, W. Terrestrial Vegetation Redistribution and Carbon Balance under Climate Change. Carbon Balance Manag. 2006, 1, 6. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhuang, Q.; Schaphoff, S.; Sitch, S.; Sokolov, A.; Kicklighter, D.; Melillo, J. Uncertainty Analysis of Vegetation Distribution in the Northern High Latitudes during the 21st Century with a Dynamic Vegetation Model. Ecol. Evol. 2012, 2, 593–614. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, P.; Neilson, R.P.; Lenihan, J.M.; Drapek, R.J. Global Patterns in the Vulnerability of Ecosystems to Vegetation Shifts Due to Climate Change: Global Vulnerability to Climate Change. Glob. Ecol. Biogeogr. 2010, 19, 755–768. [Google Scholar] [CrossRef]
- Hojati, M.; Mokarram, M. Determination of a Topographic Wetness Index Using High Resolution Digital Elevation Models. Eur. J. Geogr. 2016, 7, 41–52. [Google Scholar]
- Liu, L.; Guan, L.; Zhao, H.; Huang, Y.; Mou, Q.; Liu, K.; Chen, T.; Wang, X.; Zhang, Y.; Wei, B.; et al. Modeling Habitat Suitability of Houttuynia cordata Thunb (Ceercao) Using MaxEnt under Climate Change in China. Ecol. Inform. 2021, 63, 101324. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the Accuracy of Diagnostic Systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Riordan, E.C.; Rundel, P.W. Land Use Compounds Habitat Losses under Projected Climate Change in a Threatened California Ecosystem. PLoS ONE 2014, 9, e86487. [Google Scholar] [CrossRef]
- Sleeter, B.M.; Sohl, T.L.; Bouchard, M.A.; Reker, R.R.; Soulard, C.E.; Acevedo, W.; Griffith, G.E.; Sleeter, R.R.; Auch, R.F.; Sayler, K.L.; et al. Scenarios of Land Use and Land Cover Change in the Conterminous United States: Utilizing the Special Report on Emission Scenarios at Ecoregional Scales. Glob. Environ. Chang. 2012, 22, 896–914. [Google Scholar] [CrossRef]
- Liu, G.; Mai, J. Habitat Shifts of Jatropha curcas L. in the Asia-Pacific Region under Climate Change Scenarios. Energy 2022, 251, 123885. [Google Scholar] [CrossRef]
- Rahimian Boogar, A.; Salehi, H.; Pourghasemi, H.R.; Blaschke, T. Predicting Habitat Suitability and Conserving Juniperus spp. Habitat Using SVM and Maximum Entropy Machine Learning Techniques. Water 2019, 11, 2049. [Google Scholar] [CrossRef]
- Rahmani, R. Potential Distribution and the Habitat Suitability of the African Mustard (Brassica tournefortii) in Tunisia in the Context of Climate Change. Arab. J. Geosci. 2020, 13, 512. [Google Scholar] [CrossRef]
- Ben Mariem, H. Climate change impacts on the distribution of Stipa tenacissima L. Ecosystems in north african arid zone—A case study in Tunisia. Appl. Ecol. Environ. Res. 2017, 15, 67–82. [Google Scholar] [CrossRef]
- Du, Z.; He, Y.; Wang, H.; Wang, C.; Duan, Y. Potential Geographical Distribution and Habitat Shift of the Genus Ammopiptanthus in China under Current and Future Climate Change Based on the MaxEnt Model. J. Arid Environ. 2021, 184, 104328. [Google Scholar] [CrossRef]
- Xu, N.; Meng, F.; Zhou, G.; Li, Y.; Wang, B.; Lu, H. Assessing the Suitable Cultivation Areas for Scutellaria baicalensis in China Using the Maxent Model and Multiple Linear Regression. Biochem. Syst. Ecol. 2020, 90, 104052. [Google Scholar] [CrossRef]
- Feng, L.; Sun, J.; El-Kassaby, Y.A.; Yang, X.; Tian, X.; Wang, T. Predicting Potential Habitat of a Plant Species with Small Populations under Climate Change: Ostryarehderiana. Forests 2022, 13, 129. [Google Scholar] [CrossRef]
- Feng, L.; Sun, J.; Shi, Y.; Wang, G.; Wang, T. Predicting Suitable Habitats of Camptotheca Acuminata Considering Both Climatic and Soil Variables. Forests 2020, 11, 891. [Google Scholar] [CrossRef]
- Yu, F.; Wang, T.; Groen, T.A.; Skidmore, A.K.; Yang, X.; Ma, K.; Wu, Z. Climate and Land Use Changes Will Degrade the Distribution of Rhododendrons in China. Sci. Total Environ. 2019, 659, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Lim, C.-H.; Chung, H.I.; Kim, Y.; Cho, H.J.; Hwang, J.; Kraxner, F.; Biging, G.S.; Lee, W.-K.; Chon, J.; et al. Forest Management Can Mitigate Negative Impacts of Climate and Land-Use Change on Plant Biodiversity: Insights from the Republic of Korea. J. Environ. Manag. 2021, 288, 112400. [Google Scholar] [CrossRef]
- Corlett, R.T.; Westcott, D.A. Will Plant Movements Keep up with Climate Change? Trends Ecol. Evol. 2013, 28, 482–488. [Google Scholar] [CrossRef]
- Lee, S.; Jung, H.; Choi, J. Projecting the Impact of Climate Change on the Spatial Distribution of Six Subalpine Tree Species in South Korea Using a Multi-Model Ensemble Approach. Forests 2020, 12, 37. [Google Scholar] [CrossRef]
- Wang, G.; Wang, C.; Guo, Z.; Dai, L.; Wu, Y.; Liu, H.; Li, Y.; Chen, H.; Zhang, Y.; Zhao, Y.; et al. Integrating Maxent Model and Landscape Ecology Theory for Studying Spatiotemporal Dynamics of Habitat: Suggestions for Conservation of Endangered Red-Crowned Crane. Ecol. Indic. 2020, 116, 106472. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Species | Family | Life Form |
|---|---|---|---|
| Evergreen coniferous tree | Cephalotaxus harringtonia (Knight ex Forbes) K. Koch | Taxaceae | Tree |
| Evergreen broadleaf tree | Camellia japonical L. | Theaceae | Tree |
| Elaeagnus macrophylla Thunb. | Elaeagnaceae | Shrub | |
| Eurya japonica Thunb. | Pentaphylacaceae | Tree | |
| Machilus thunbergii Siebold & Zucc. | Lauraceae | Tree | |
| Neolitsea sericea (Blume) Koidz. | Lauraceae | Tree | |
| Deciduous broadleaf tree | Neoshirakia japonica (Siebold and Zucc.) Esser | Euphorbiaceae | Tree |
| Orixa japonica Thunb. | Rutaceae | Shrub | |
| Stewartia koreana Nakai ex Rehder | Theaceae | Tree |
| Species | Number of Datasets |
|---|---|
| C. japonica | 129 |
| C. harringtonia | 53 |
| E. macrophylla | 112 |
| E. japonica | 169 |
| M. thunbergii | 114 |
| N. sericea | 106 |
| N. japonica | 126 |
| O. japonica | 92 |
| S. koreana | 64 |
| Category | Variables | Data Type |
|---|---|---|
| Climatic | Annual average temperature | Continuous |
| Maximum temperature | ||
| Minimum temperature | ||
| Average precipitation | ||
| Topographic | Altitude | |
| Topographic wetness index | ||
| Aspect | ||
| Slope | ||
| Soil depth | Categorical |
| Classification | Suitable Habitat Index |
|---|---|
| Unsuitable habitat | p < 0.1 |
| Poorly suitable habitat | 0.1 ≤ p < 0.3 |
| Moderately suitable habitat | 0.3 ≤ p < 0.7 |
| Highly suitable habitat | 0.7 ≤ p < 1.0 |
| Species | AUC Values |
|---|---|
| C. japonica | 0.951 |
| C. harringtonia | 0.801 |
| E. macrophylla | 0.954 |
| E. japonica | 0.944 |
| M. thunbergii | 0.971 |
| N. sericea | 0.983 |
| N. japonica | 0.892 |
| O. japonica | 0.943 |
| S. koreana | 0.873 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, Y.; Shin, E.; Lee, S.-W.; An, K. Predicting Changes in and Future Distributions of Plant Habitats of Climate-Sensitive Biological Indicator Species in South Korea. Sustainability 2024, 16, 1013. https://doi.org/10.3390/su16031013
Shin Y, Shin E, Lee S-W, An K. Predicting Changes in and Future Distributions of Plant Habitats of Climate-Sensitive Biological Indicator Species in South Korea. Sustainability. 2024; 16(3):1013. https://doi.org/10.3390/su16031013
Chicago/Turabian StyleShin, Yeeun, Eunseo Shin, Sang-Woo Lee, and Kyungjin An. 2024. "Predicting Changes in and Future Distributions of Plant Habitats of Climate-Sensitive Biological Indicator Species in South Korea" Sustainability 16, no. 3: 1013. https://doi.org/10.3390/su16031013
APA StyleShin, Y., Shin, E., Lee, S.-W., & An, K. (2024). Predicting Changes in and Future Distributions of Plant Habitats of Climate-Sensitive Biological Indicator Species in South Korea. Sustainability, 16(3), 1013. https://doi.org/10.3390/su16031013