1. Introduction
In the context of global climate change, low-temperature stress poses a severe challenge to agricultural production. Low temperatures limit plant growth and development and directly affect crop yield and quality [
1]. Over the course of evolution, plants have developed complex physiological and molecular mechanisms to adapt to low-temperature environments. Among them, the endogenous plant hormones jasmonic acid (JA) and methyl jasmonate (MeJA) regulate plant responses to cold stress. Under low-temperature stress, electrolyte leakage, enzyme inactivation, and ice crystal formation in plant cells cause different degrees of damage to plants [
2]. In particular, during freezing (<0 °C), intracellular water molecules are transformed into extracellular ice crystals, which disrupts the integrity of the cell membrane and leads to the denaturation of membrane proteins. This injury is often fatal [
3]. Plants respond to these environmental stresses through finely regulated stress responses, activating the antioxidant system [
4]. The antioxidant system consists of the activation of the ascorbate system 4. The antioxidant system includes metabolites such as ascorbic acid (AsA), reduced glutathione (GSH), and antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), peroxidase (POD), ascorbate peroxidase (APX), and glutathione reductase (GR) [
5,
6,
7,
8]. Together, these antioxidant components constitute the plant’s defense against the damaging effects of ROS. As an essential endogenous plant hormone, JA has a broad spectrum of physiological effects [
9]. It regulates plant growth and development and plays a vital role in plant response to biotic and abiotic stresses, especially low-temperature stresses. Studies have shown that JA enhances cold stress responses in plants by activating downstream signaling pathways. For example, the exogenous application of MeJA could increase the chlorophyll fluorescence parameter (Fv/Fm) and decrease the relative electrical conductivity (REC) of the tomato, thereby improving its cold tolerance [
10]. In addition, MeJA also promotes the scavenging of reactive oxygen species (ROS) under cold stress conditions, which effectively improves the tolerance to cold stress in tea trees [
11].
However, although the potential role of JAs in plant cold tolerance has been widely recognized, its specific mechanism of action and the effects of its application under different conditions still lack systematic understanding. The present review aimed to fill this knowledge gap by investigating the effects of exogenous JAs on plant cold stress acclimation through systematic evaluations and meta-analyses to address the following scientific questions: (1) How do JAs systematically affect cold stress response in plants? (2) What are the potentials and limitations of exogenous JA applications in practical agricultural production?
3. Results
3.1. Systematic Evaluation
A total of 247 research papers involving JAs’ involvement in plants in cold stress were collected between 2018 and 2022. The analysis (
Figure 2A) shows that although the annual publication volume fluctuated, the overall trend was stable. In particular, the number of published papers peaked in 2020, with a total of 60 papers. Upon further analysis of the literature according to the country of the first author (
Figure 2B), China’s research contribution in this field is the most significant, with a total of 177 papers, accounting for 72% of the total, followed by India (10 papers); Spain (9 papers); Turkey (7 papers); the United States and Iran, both with 5 papers; Germany, Russia, and South Korea, all with 3 papers; Germany, Hungary, Egypt, Canada, Malaysia, Thailand, and the UK, all with 2 articles; and other countries not listed contributed 1 document.
After analyzing the research content of the 247 papers, this review grouped them into four main research directions: low-temperature response, fruit storage, structure and function, and breeding and cultivation. The literature on low-temperature responses mainly focused on the physiological and molecular responses of plants sprayed with JA under low-temperature stress, as well as the effects of low-temperature treatments on JA content in plants. Fruit storage studies in the literature focus on the improvement of fruit quality by substance treatments, including the enhancement of fruit freshness and the extension of storage time. The structure–function literature focuses on cis-acting elements in gene promoter structures in response to low temperature and JAs. In contrast, the breeding and cultivation literature focuses on the selection of cold-resistant varieties and agronomic measures to improve cold resistance in plants.
Of all 247 articles analyzed, 99 (40%) included both molecular-level and physiological-level tests. In addition, 82 articles (33% of the total) were purely molecular tests, while 66 articles (27%) were strictly physiological tests. Upon further analysis of the source of sampling of research material in the literature, we found that most studies (243 articles or 98%) used indoor testing methods, in contrast to only 4 articles (2%) that used outdoor testing as a research method.
In this review, we conducted a comprehensive statistics analysis, and the organization of the indicators measured in the literature was included in the analysis. As shown in
Figure 2C, the physiological indicators that have received extensive attention for the role of JAs in plant cold tolerance include, in order of importance, JA, MDA, SOD, Chl, CAT, electrolyte permeability, ABA, and total TPC and ABA.
The plant material used in the literature and included in the analysis was evaluated, and this material covered a wide range of classifications, including angiosperms and gymnosperms. Among angiosperms, dicots were used most frequently. Specifically, for single species, the most used material in the studies was tomato, with 19 articles, as shown in
Figure 2D. Among monocots, wheat was the most popular material, with 12 articles in the relevant research literature.
Based on the main content of the research material in 247 publications, this review found that the main spray substances involved in studies analyzing the effect of JA application on plant cold resistance were mainly JA and MeJA. Specifically, 33 publications reported experiments with exogenous spraying of JA, while 37 explored the effect of exogenous spraying of MeJA.
The gene studies involved in the included literature were systematically organized and analyzed in detail according to the threshold classification (
Supplemental Data S2). Currently, studies on the involvement of JAs in plant chilling resistance focus on the following gene categories: genes encoding enzymes, genes encoding proteins, pathway genes, transcription factors, and functional genes. In particular, transcription factor gene families play an essential role in jasmonic acid-mediated chilling resistance responses, involving the following families: ERF, EIN, WRKY, MADS, MYB, BHLH, TIFY, bHLH, NAC, bZIP, DREB, WOX, BBX, ICE, NF-Y, SlERF, GRF, MYC, and CBF, for a total of 19 gene families. In terms of antioxidant enzyme synthesis genes, this analysis covered vital genes such as SOD, CAT, APX, PRX, and LOX.
3.2. Analysis of Combined Effects
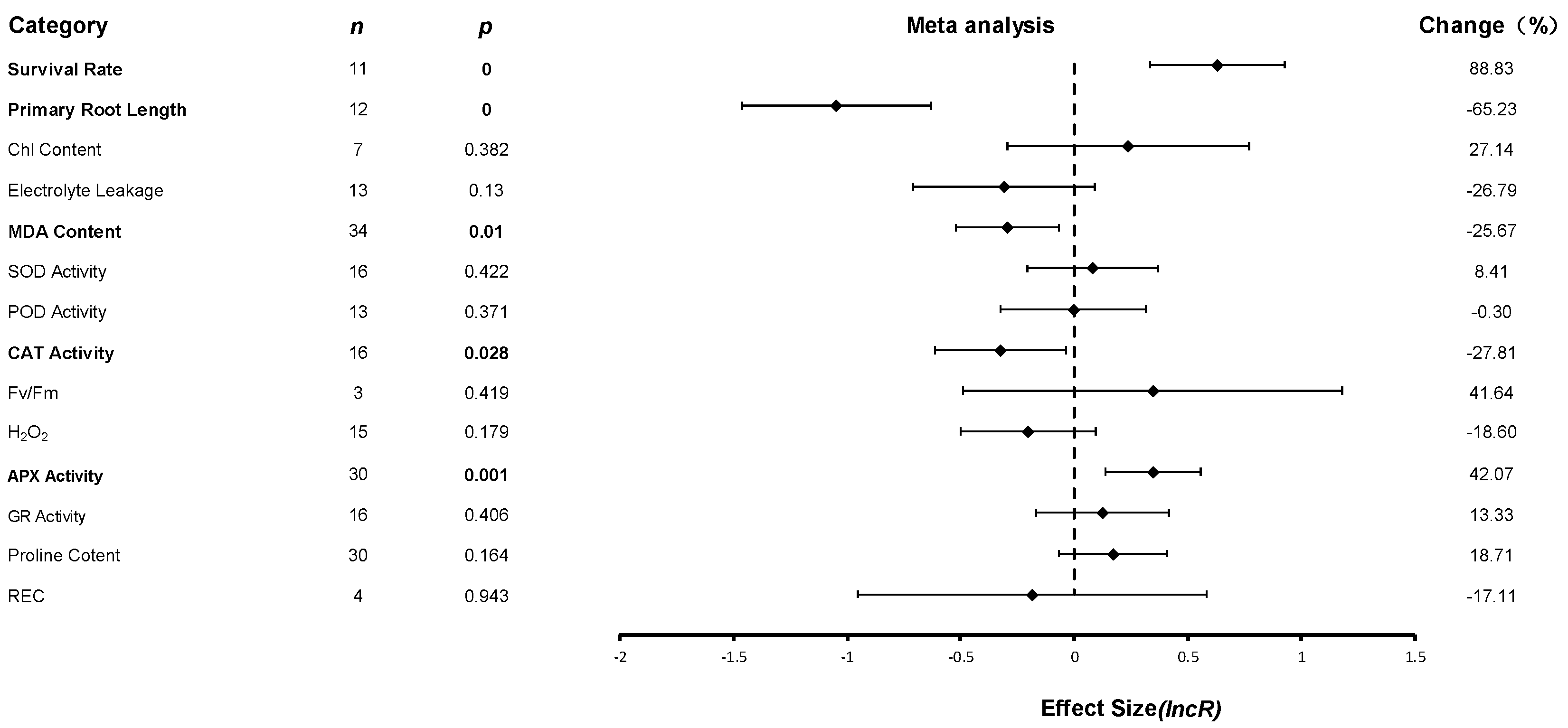
Data for all pooled effect values were uniformly transformed to the corresponding natural logarithms and presented in forest plots. Effect values for 14 indicators were aggregated, and 220 studies from 16 papers were included in the meta-analysis. The JA-induced changes in the various indicators of plant cold tolerance and their combined effect values are presented in a forest plot, as shown in
Figure 3.
The dotted line in the forest plot (value of 0) indicates that JA treatment did not affect plant physiological indices under low-temperature conditions. If the value is less than 0 (i.e., to the left of the vertical line), it means that the physiological index was reduced after JA treatment relative to the control; if the value is greater than 0 (i.e., to the right of the vertical line), it means that the physiological index was increased after JA treatment. The changes in APX activity were highly significant (p ≤ 0.01). The changes in the surviving rate, the length of the primary root, and the content of MDA were substantial (p ≤ 0.05), which indicated that JAs improved plant cold resistance under low temperatures by changing these indexes to enhance the freezing tolerance of plants at low temperatures. JA treatment had the most significant effect on survival rates at low temperatures, which were increased by 88%. The promotion effect on APX activity was 42%. JA treatment significantly inhibited primary root length (65%), MDA content (25%), and CAT activity (27%).
3.3. Publication Bias
Positive findings are more likely to be published and, therefore, more likely to cause publication bias [
18], which leads to incomplete inclusion of studies. To test for publication bias, several statistical methods have been used that are based on exploring the relationship between study effect size and precision, and it is generally recognized that high-quality studies with large sample sizes or low variance tend to show smaller effect sizes. In contrast, low-quality studies with small sample sizes or high variance may show larger effect sizes [
19].
Nearly half of the composite effects studied in this paper had asymmetric funnel plots, suggesting potential publication bias (
Table 1). Using Kendall’s tau test, it was observed that 13 out of 14 composite effects had tau values higher than 0.2 or
p-values > 0.05, indicating that these composite effects were subject to slight bias; however, for the composite effect of MDA content,
p ≤ 0.05, which may indicate the presence of publication bias. Egger’s regression test of the two-tailed significance test criterion further revealed the possible bias for the four composite effects. The results of the analyses by the trim and fill method indicated that the adjusted values of the seven composite effects were closer to 0, which further supported the idea that the composite effects might be affected by publication bias. Despite these indications, as shown in
Table 1, the combined assessment of the four statistical methods did not find that the aggregated effect sizes of the tested indicators were affected by bias. Therefore, we decided to include all indicators in the meta-analysis.
3.4. Heterogeneity Analysis
As shown in
Table 2, statistically significant heterogeneity was not found in any of the 14 composite effect sizes. In the meta-analysis, heterogeneity in effect sizes usually refers to the variation in accurate effect sizes across studies; however, in experimental biological studies, heterogeneity may be affected by a variety of factors, such as insufficient sample sizes or significant differences in experimental conditions, which may lead to the observed variation in effect sizes. It is worth noting that even if phetero > 0.100 or I
2 has a value of 0, this does not necessarily mean no true heterogeneity between studies [
14]. Similarly, a significant variation in accurate effect sizes may result in phetero < 0.100. Therefore, in assessing heterogeneity, a random effects model was chosen as appropriate for subgroup analyses of the combined effect values of the different variables.
3.5. Subgroup Analyses
To investigate the heterogeneity between the true effect values, five physiological indicators were analyzed that showed significant changes in combined effect values under exogenous JA treatment: survival rate, primary root length, MDA content, CAT activity, and APX activity. Subgroup analyses of these indicators were performed to reveal possible effect size variation between studies.
3.5.1. Effect of JA Treatment on Plant Biomass
Exogenous spraying of JAs significantly promoted plant survival. The impact of exogenously sprayed JAs on the combined effect value of plant survival under low-temperature stress conditions was determined by analyzing five variables: stress time, plant type, stress level, JA concentration, and gene treatment (
Figure 4). At low temperatures, plant survival was increased by 83% when plants were exogenously sprayed with JAs at concentrations ranging from 10 to 50 μM; exogenous spraying of JAs had a more significant positive effect on survival when plants were cold-stressed for 2–10 days; exogenous spraying of JAs increased survival by 73% when plants were subjected to freezing as a low-temperature stressor, and by 118% when plants were subjected to cold stress. When plants were annuals, low-temperature spraying of JAs did not significantly affect survival; when plants were overexpressing a gene, low-temperature spraying of JAs increased survival by 83%.
Exogenous spraying of JAs significantly inhibited the primary root length of plants. The effect of exogenously sprayed JAs on the combined effect value of plant primary root growth under low-temperature stress conditions was determined by analyzing four variables: time of stress, type of JAs, concentration of JAs, and gene treatment (
Figure 5). At low temperatures, exogenous sprays of MeJA and JA both inhibited primary root growth, where spraying MeJA inhibited primary root growth by 72% while spraying JA inhibited primary root growth by 58%. Primary root growth was significantly inhibited by exogenous sprays of 50–100 μM, or less than 10 μM of JAs, whereas the effect of exogenous JAs on primary root length was not significant in the range of 10–50 μM; furthermore, the length of time the plants were subjected to cold stress and the changes in genes within the plant had a similar effect on the composite effect value when exogenous sprays of JAs were applied at low temperatures and both significantly inhibited primary root length.
3.5.2. Effect of JA Treatment on Plant Ability to Respond to Adversity
Exogenously sprayed JAs significantly promoted APX activity in plants. The effects of exogenously sprayed JAs on the combined effect values of APX activity of the plants under low-temperature stress conditions were determined by analyzing five variables, namely, culture medium, stress time, plant type, degree of stress, and gene treatment (
Figure 6). At low temperatures, exogenously sprayed JAs promoted APX activity when plants were in a liquid medium; exogenously sprayed JAs promoted APX activity by 106% and 98% when plants were under cold stress for 11–20 days and greater than 20 days, respectively; exogenously sprayed JAs boosted APX activity by 44% when plants suffered from cold stress, whereas freezing damage did not significantly affect APX activity; and exogenously sprayed JAs enhanced APX activity by 44% when plants were annual, while freeze damage did not considerably influence APX activity. When plants were annual, exogenous spraying of JAs promoted APX activity under low-temperature conditions; when a gene was repressed in plants, exogenous spraying of JAs did not have a significant effect on APX activity under low-temperature conditions.
Exogenously sprayed JAs significantly inhibited the CAT activity of the plants. The effect of exogenously sprayed JAs on the combined effect value of CAT activity of the plants under low-temperature stress conditions was determined by analyzing four variables: culture medium, stress time, degree of stress, and gene treatment (
Figure 7). At low temperatures, exogenously sprayed JAs inhibited CAT activity by 36% when plants were in a liquid medium, whereas CAT activity was not significantly affected when plants were in a soil medium; exogenously sprayed JAs inhibited CAT activity when plants were subjected to low-temperature stress with cold damage, whereas neither the time of stress nor the gene type variables had a significant effect on the combined effect value of CAT activity.
Exogenously sprayed JAs significantly inhibited the MDA content of the plants. The effect of exogenously sprayed JAs on the combined effect value of MDA content of plants under low-temperature stress conditions was determined by analyzing six variables: culture medium, stress time, JA type, degree of stress, JA concentration, and gene treatments, as shown in
Figure 8. Exogenously sprayed JAs at low temperatures, when plants were in the solid medium, compared with plants in the soil and liquid mediums, reduced MDA content by 73%; exogenous spraying of JA reduced MDA content by 77% when plants were under cold stress compared to spraying MeJA; exogenous spraying of JAs did not have a significant effect on MDA content when plants were under cold stress for more than 20 days compared to less than 2 days; exogenous spraying of JAs did not have a significant effect on MDA content; and any JA concentration did not have a substantial effect on MDA content.
4. Discussion
For systematic evaluation, this review collected and analyzed 247 research papers published between 2018 and 2022 involving the role of JAs in plant cold stress. The studies focused on four primary categories, namely low-temperature response, fruit storage, structure–function, and breeding cultivation, covering multiple research directions from the molecular to the physiological level. The physiological indicators of JAs in plant cold resistance are, in order of interest: JAs, malondialdehyde (MDA), superoxide dismutase (SOD), chlorophyll (Chl), catalase (CAT), electrolyte osmosis (EO) and chlorophyll (Chl), electrolyte permeability, abscisic acid (ABA), and total phenol content (TPC).
In addition, the 247 studies were analyzed comprehensively, with only 4 field experiments, and the rest were indoor studies, reflecting that most of the studies remained in the laboratory and could not be widely applied to field production. Future studies should also focus on the potential application of JAs in actual agricultural production, and field experiments should be carried out to validate the effectiveness and feasibility of JA application to meet the exact needs of agricultural production.
In addition, the studies covered 83 plant species, including food and model crops such as tomato and Arabidopsis, but fewer cash crops. There were far more dicotyledonous plant studies (211) than monocotyledonous plants (36), suggesting the need for increased attention to monocotyledonous plants, an essential source of food, in the future.
Exogenous spraying of JAs significantly inhibited the primary root length and increased the survival rate of plants. The survival rate is the most direct judgment of plant health status; a meta-analysis showed that spraying JAs increased the survival rate of plants significantly, and exogenous spraying of JAs was able to resist cold stress and make plants less burdened effectively. The ability of root formation is controlled by various endogenous and environmental factors, among which growth factors play the most crucial role. It has been shown that JAs induce growth hormone biosynthesis, and it was revealed that JAs play a regulatory role in the attenuation of growth hormone transport in roots. The local distribution of growth hormone in root meristematic tissues is essential for lateral root formation [
20]. Therefore, low concentrations of JA and MeJA inhibited primary root growth and promoted lateral root formation in Arabidopsis thaliana [
21]. This inhibitory effect may be related to the regulatory role of JAs under adversity, i.e., under cold stress, JAs may promote the development of lateral roots or root systems by inhibiting the growth of primary roots, thus improving the plant’s adaptation to cold stress.
In the present review, exogenous JA treatment significantly enhanced plant survival under low-temperature conditions, a result consistent with previous studies [
22,
23]. This suggests that JAs enhance plants’ cold stress response by activating downstream signaling pathways. In addition, the promotion of APX activity by JA treatment suggests that JAs may enhance cold stress tolerance in plants by enhancing their antioxidant capacity [
24,
25]. This was also confirmed by subgroup analyses, especially in liquid media, where exogenous spraying of JAs significantly promoted APX activity [
26,
27].
Furthermore, the role of JAs in regulating oxidative stress in plants should be considered. JA treatment significantly reduced MDA content, suggesting its role in regulating oxidative stress in plants [
21,
28]. The reduction in MDA content, the end product of membrane lipid peroxidation, suggests that JAs may protect the integrity of plant cell membranes by reducing the accumulation of reactive oxygen species (ROS) [
11,
29]. This was also reflected in the effect of exogenously sprayed JAs on CAT activity. Despite the inhibition of CAT activity, the reduction in MDA content suggests that JAs may regulate oxidative stress in plants through other antioxidant pathways [
27].
JA treatment significantly promoted ascorbate peroxidase (APX) activity while inhibiting malondialdehyde (MDA) content and catalase (CAT) activity. MDA content is an important lipid peroxidation marker for assessing cold injury. It increases when membrane permeability is elevated due to the increase in peroxidation of cellular membranes under cold stress [
30]. This is consistent with the results of the meta-analysis that exogenous spraying of JAs can inhibit the MDA content, thus effectively alleviating the injury of plants subjected to cold stress Studies have shown that overexpression of the APX gene in the cytoplasm removes excessive reactive oxygen species in Arabidopsis thaliana [
31]. This also suggests that with an increase in APX activity, the reactive oxygen species that can be efficiently removed can withstand the cold stress. Notably, spraying JAs reduced CAT activity by 27%. Research indicates that, under low-temperature conditions, the catalase (CAT) activity in the jasmonic acid-sprayed treatment group is significantly lower than that in the non-sprayed control group when a recovery period is incorporated into the experimental design [
11]. In contrast, when no recovery time is added, the CAT activity of the JA-sprayed treatment group will be significantly higher than that of the unsprayed treatment group, which suggests that the response of CAT activity is rapid, as well as that changes in the activity of CAT with the increase in time will show different trends. The article showed that CAT activity, with the decrease in temperature and the prolongation of low-temperature stress time, showed a trend of first increasing and then decreasing, indicating that a large number of reactive oxygen radicals were formed in the body of the plant after low-temperature stress; to eliminate such free radicals to protect the enzyme, activity rose sharply. The CAT activity showed a decline at the late stage of stress, which may be because the leaves slowly adapted to the low temperature, and the status was eased [
32,
33]. In summary, the CAT activity increased significantly in a short period after spraying JAs. Then, it showed a decreasing trend in the later period with the prolongation of time, which was consistent with the meta results.
The mechanism of action of JAs may be more complex, involving the regulation of multiple signaling pathways and genes. The current study focuses on the effects of JAs on several critical physiological and biochemical indicators. In conclusion, although JAs improve cold tolerance in plants, the meta-analysis reveals that its mechanism of action and scope of application still need to be explored in depth. It is recommended that the study of JAs be extended to a variety of plant species, in particular, to combine indoor experiments with field experiments to meet the needs of practical agricultural production. In the future, the mechanism of JAs against cold stress should be explored in depth, which will help to lay the foundation for future work related to plant stress tolerance breeding and pave the way for new techniques.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}