
A Model for Spatially Explicit Landscape Configuration and Ecosystem Service Performance, ESMAX: Model Description and Explanation


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Extending Ecological Field Theory
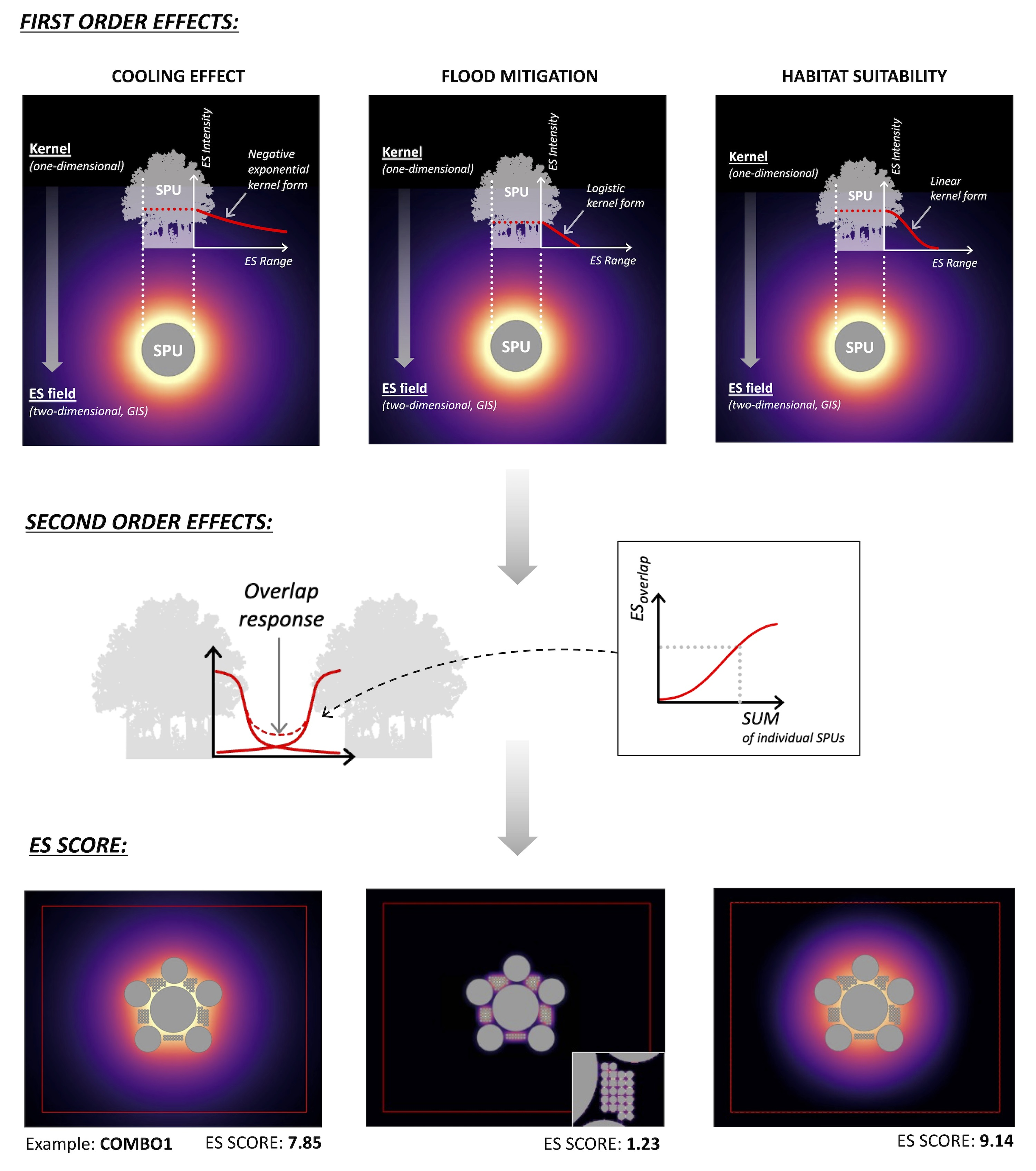
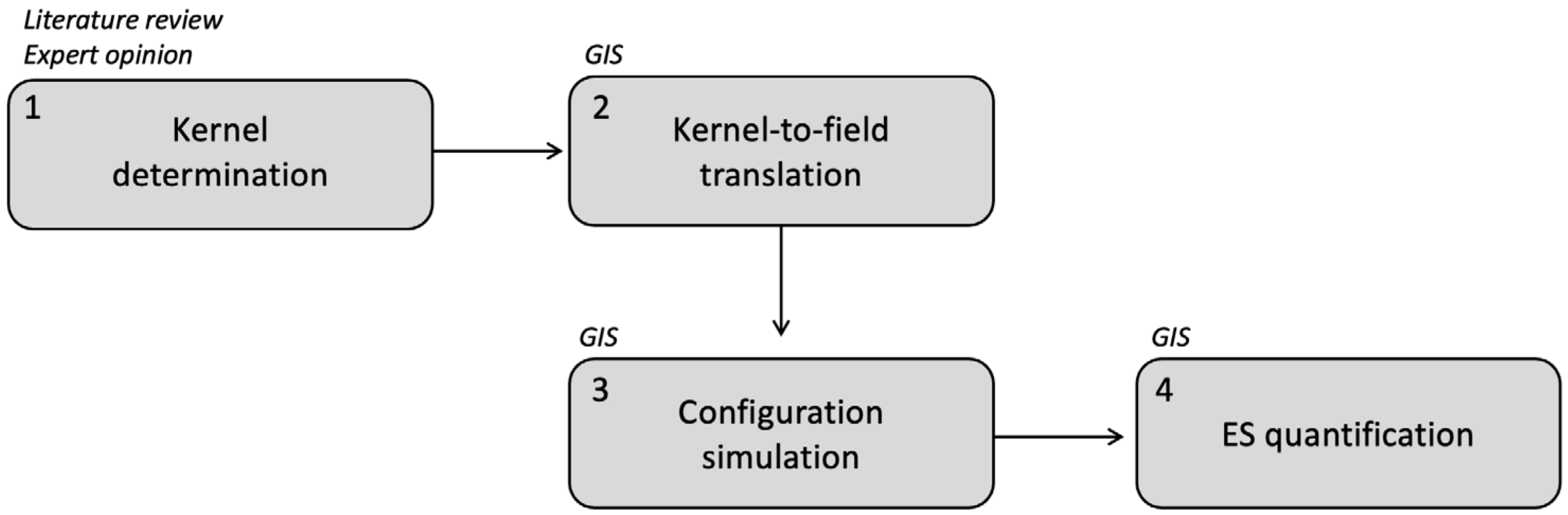
2.2. Model Structure
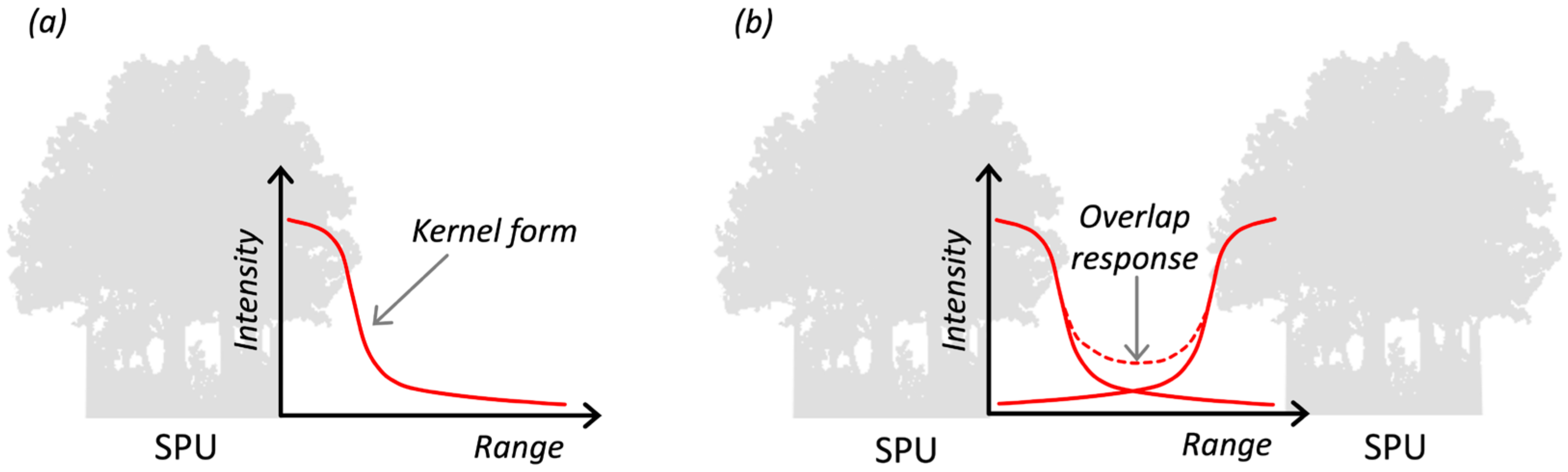
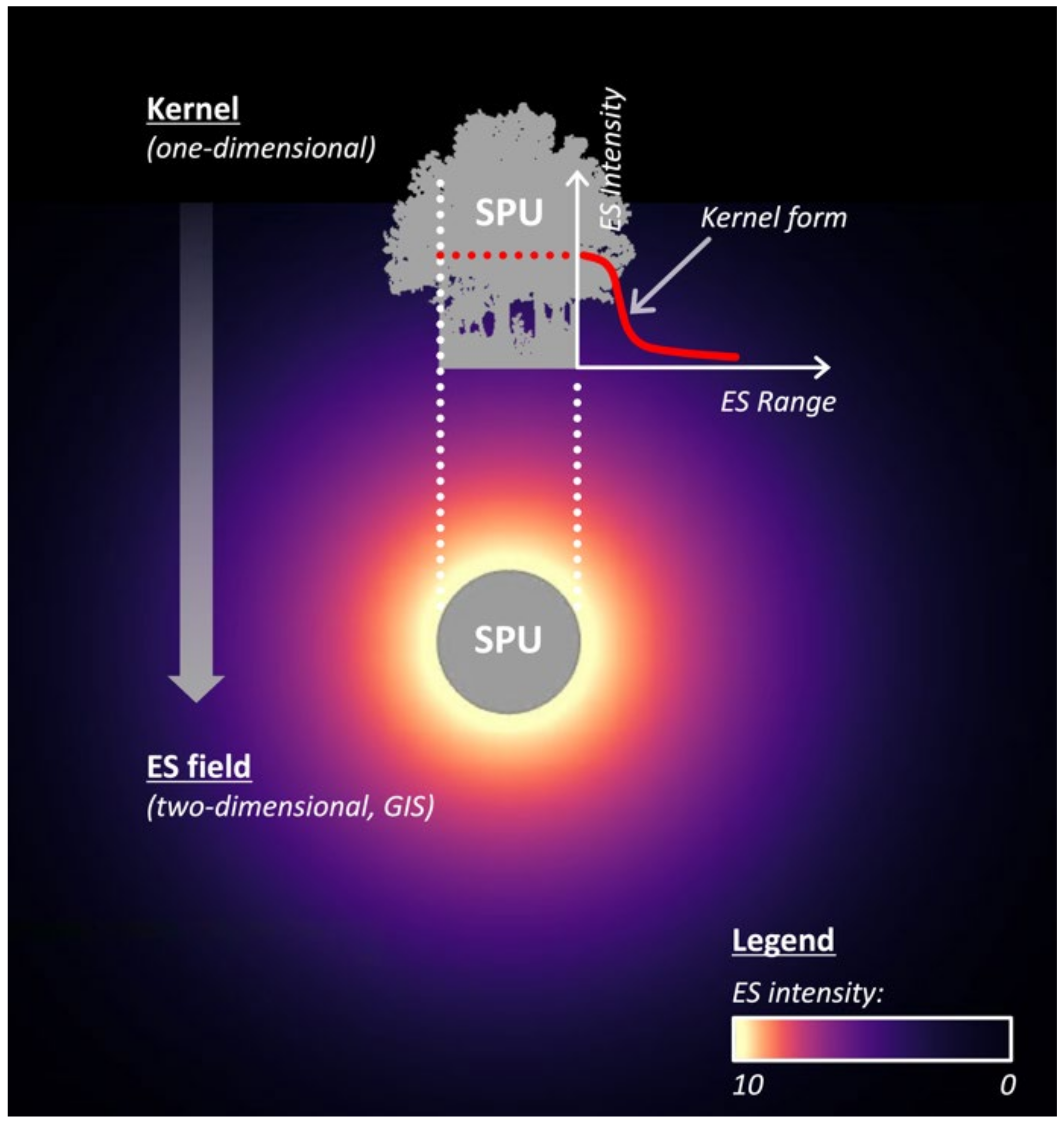
2.2.1. Step 1—Kernel Determination
Intensity
Range
Kernel Form
Overlap Response
2.2.2. Step 2—Kernel-to-ES Field Translation
2.2.3. Step 3—Configuration Simulation
- As the SPUs are circular shapes, the respective ES fields of the SPUs are isotropic radiating circles, meaning that ES distance-decay is constant in all possible directions from the centre of the SPU.
- A neutral background landscape is assumed, contributing no ESs and homogeneous in terms of land use, soil type, etc., without landscape features (such as watercourses).
- Each SPU is assumed to comprise species suitable for testing the ESs included in our conceptual model. Future research could add composition as a further explanatory variable.
- Seasonality is not considered, and we assume the trees to be at peak maturity. The context of this initial development of the ESMAX is mainly spatial—we acknowledge there is a temporal dimension to ES supply, but we consider it implicitly by using a hypothetical time period and resolution that permits the assumption that rates are constant.
- SPUs are limited to three sizes: 6 ha, 2 ha, and 0.02 ha (designated large—L, medium—M, and extra small—XS, respectively). These areas represent thresholds for the specific ES being assessed. For example, Meurk et al. (2006) establish a functional hierarchy of bird nesting/feeding patch sizes, which we adapt for use in this research [45]. These thresholds are detailed in Section 2.3.3 below.
- Setting the total SPU area to 18 ha allows straightforward, whole-number distribution of homogeneously sized SPUs, i.e., 3 × 6 ha, 9 × 2 ha, and 900 × 0.02 ha. ‘COMBO’ configurations are also generated, which combined the three SPU sizes, while still maintaining a total SPU area of 18 ha (Figure 4).

2.2.4. Step 4—Field Quantification
2.3. Application to Specific ESs
2.3.1. Selection of ESs
2.3.2. Cooling Effect
- For the purposes of this model, the correlation of cooling performance increasing with SPU size noted in the literature is reflected in the kernel range (dcool) increasing proportionally to the SPU diameter [38].
- The range is calculated with reference to the literature on micrometeorological phenomena characteristic of forest edge contexts [52,53]. The displaced, cool sub-canopy air will travel the same distance (dcool) as the diameter of the source SPU. This assumption is supported by urban cooling research, which notes a cooling influence range of one park width away from the cooling source park [50,54].
- The kernel form is determined using the negative exponential distance-decay used in the InVEST (Integrated Valuation of Ecosystem Services and Trade-offs) urban cooling model [10].
- Overlap intensity of cooling effect is poorly understood. Our model ESMAX assumes that the overlapping of cooling fields will follow the same principle as the nonlinear amplification of temperature between two heat sources [55].
2.3.3. Habitat Suitability
- SPUs of 6 ha approximate a minimum core habitat area of 2.5 ha (once a perimeter buffer to the SPU edge of around 50 m is established), suitable for sanctuaries in human-modified landscapes [45].
- SPUs of 2 ha provide habitat for most plants, lizards, insectivorous birds, and invertebrates and provide resource-rich ‘steppingstones’ for nectivorous birds [45].
- SPUs of 200 m2 (0.02 ha) provide finer-grained steppingstones and feeding locations [45].
- Intensity, in this case, is a measure of feeding activity from a nesting base and was given an arbitrary maximum value. It is recognised that there is a minimum SPU area required for the establishment of nesting and establishment of a home range [45]. The insectivore will nest even in the smallest 0.02 ha SPUs (which translates to a 16 m diameter tree clump), as long as the distance to a neighbouring clump is no greater than 150 m [72]. Therefore, for the insectivore, intensity is set to zero in the smallest 0.02 ha SPUs if these were located more than 150 m away from other SPUs. For the nectivore, the minimum SPU area for nesting is set at 2 ha [79], and the intensity is set to zero in all 0.02 ha SPUs.
- Range (for feeding) is set at 100 m for the insectivore and 500 m for the nectivore.
- Kernel form is based on the triweight kernel used by Laca (2021), considered a reasonable representation of population distribution from a nesting site [23]. Its single parameter, λ, is the reciprocal of the range.
- ES overlap intensity is affected by territorial characteristics. The insectivore is noted to be territorial and will aggressively defend its territories from other members of the same species [80]. However, conspecific attraction of external individuals for breeding is demonstrated even in highly territorial species [81], so the overlapping fields are considered additive. The nectivore exhibits some territorial behaviour during the breeding season, but generally, feeding ranges may overlap [76,77,79]. The nonlinear logistic overlap response for both species reflects a negative density-dependent relationship, where population growth is curtailed by crowding, predators, and competition [82,83,84].
2.3.4. Nitrogen Retention
- Maximum nitrate interception, the measure of ES intensity in this case, is given a constant arbitrary value.
- Range and kernel form are based on physical root spatial distribution measurements carried out in field research into the mechanical influence of trees on soil erosion [39,99,100]. We consider implicitly that there is some extension of ES range beyond the rhizosphere because nitrates move with water and by diffusion to volumes of lower concentration.
- We selected alder (Alnus viridis) as the focal species for this model, as it exhibited the greatest root biomass at the end of the field research referred to. This species exhibits a high degree of root interweaving [100], and we assume therefore nitrate interception capacity to be additive in areas of overlap. The rhizosphere is a vastly complex environment. For the purposes of this model, we base the additive nonlinear logistic overlap response on the following upper and lower asymptote assumptions: nutrient saturation establishes the upper asymptote [101,102,103] (i.e., maximum nitrate retention occurs closer to the root trunk), and root branching density decreasing further away from the trunk, lowering nitrate retention capacity, establishes the lower asymptote [104,105,106].
3. Results
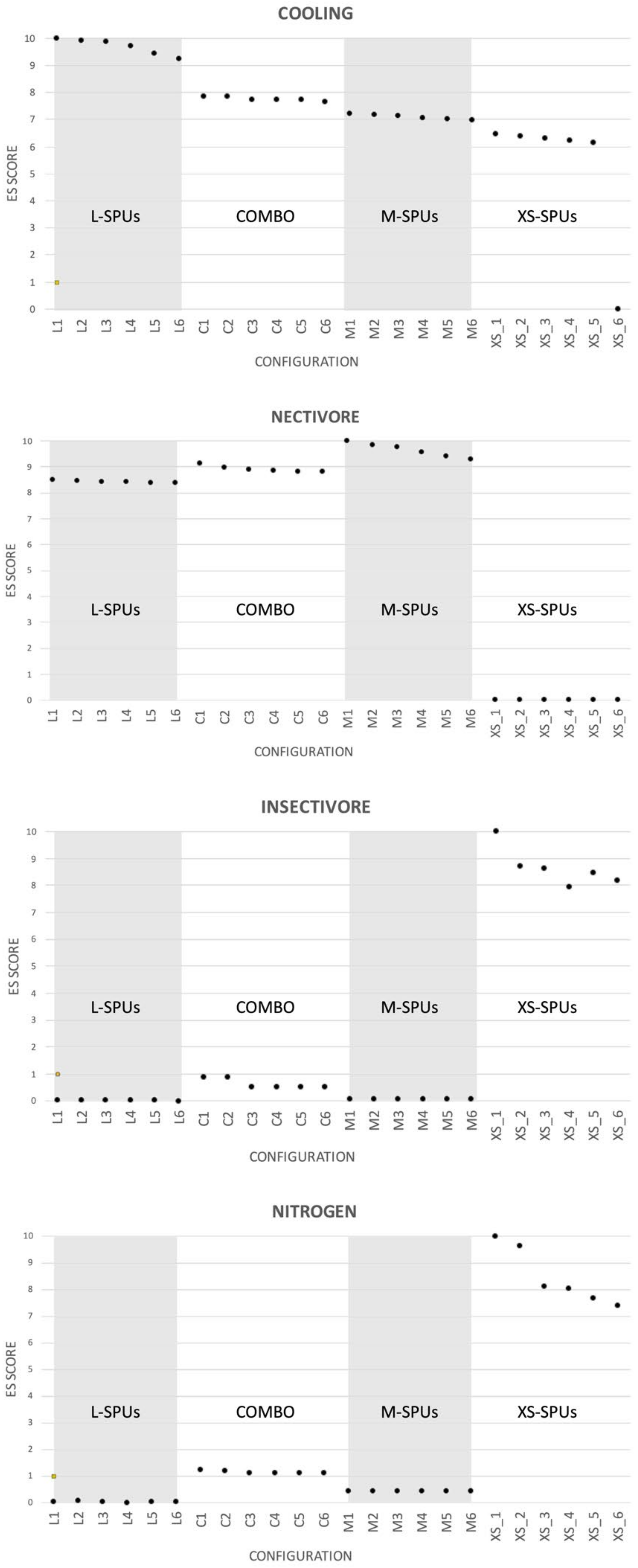
3.1. Result 1: Configuration (Size)
3.2. Result 2: Configuration (Aggregation)
4. Discussion
4.1. ES Field Theory: Addressing the Biophysical Gap
4.2. Spatial Configuration and Implications for ES-Based Design
5. Conclusions
- The protocol for development of ES kernels, based on the Ecological Field Theory approach, is a simple iterative process and provides a ready platform for understanding the spatial implications of ecological functions.
- ESMAX is potentially applicable to a wide range of other flow-based regulating ESs, including air filtration, noise abatement, bioremediation, soil erosion control, flood protection, and pollination.
- The translation of kernels, specific to each ES, to a two-dimensional ES field is a novel use of basic GIS tools, which require minimal modification for this purpose.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bennett, E.M. Changing the agriculture and environment conversation. Nat. Ecol. Evol. 2017, 1, 18. [Google Scholar] [CrossRef] [PubMed]
- Wu, J. Landscape sustainability science (II): Core questions and key approaches. Landsc. Ecol. 2021, 36, 2453–2485. [Google Scholar] [CrossRef]
- Costanza, R. Valuing natural capital and ecosystem services toward the goals of efficiency, fairness, and sustainability. Ecosyst. Serv. 2020, 43, 101096. [Google Scholar] [CrossRef]
- Starfield, A.M.; Smith, K.A.; Bleloch, A.L. How to Model It: Problem Solving for the Computer Age; Burgess International Group: Edina, MN, USA, 1994. [Google Scholar]
- Groot, J.C.J.; Rossing, W.A.H. Model-aided learning for adaptive management of natural resources: An evolutionary design perspective. Methods Ecol. Evol. 2011, 2, 643–650. [Google Scholar] [CrossRef]
- Starfield, A.M. Building Models for Conservation and Wildlife Management; Macmillan: New York, NY, USA, 1986. [Google Scholar]
- Caswell, H. Theory and models in ecology: A different perspective. Ecol. Model. 1988, 43, 33–44. [Google Scholar] [CrossRef]
- Hamann, M.; Johnson, J.A.; Chaigneau, T.; Chaplin-Kramer, R.; Mandle, L.; Rieb, J.T. Ecosystem service modelling. In The Routledge Handbook of Research Methods for Social-Ecological Systems; Biggs, R.O., Ed.; Routledge: London, UK, 2021; pp. 426–439. [Google Scholar]
- Sharps, K.; Masante, D.; Thomas, A.; Jackson, B.; Redhead, J.; May, L.; Prosser, H.; Cosby, B.; Emmett, B.; Jones, L. Comparing strengths and weaknesses of three ecosystem services modelling tools in a diverse UK river catchment. Sci. Total. Environ. 2017, 584–585, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Sharp, R.; Douglass, J.; Wolny, S. InVEST 3.10.2. User’s Guide; Stanford University: Stanford, CA, USA, 2020. [Google Scholar]
- Trodahl, M.I.; Jackson, B.M.; Deslippe, J.R.; Metherell, A.K. Investigating trade-offs between water quality and agricultural productivity using the Land Utilisation and Capability Indicator (LUCI)–A New Zealand application. Ecosyst. Serv. 2017, 26, 388–399. [Google Scholar] [CrossRef]
- Bagstad, K.J.; Johnson, G.W.; Voigt, B.; Villa, F. Spatial dynamics of ecosystem service flows: A comprehensive approach to quantifying actual services. Ecosyst. Serv. 2013, 4, 117–125. [Google Scholar] [CrossRef]
- Verhagen, W.; Van Teeffelen, A.J.A.; Baggio Compagnucci, A.; Poggio, L.; Gimona, A.; Verburg, P.H. Effects of landscape configuration on mapping ecosystem service capacity: A review of evidence and a case study in Scotland. Landsc. Ecol. 2016, 31, 1457–1479. [Google Scholar] [CrossRef]
- Rieb, J.T.; Bennett, E.M. Landscape structure as a mediator of ecosystem service interactions. Landsc. Ecol. 2020, 35, 2863–2880. [Google Scholar] [CrossRef]
- Lavorel, S.; Bayer, A.; Bondeau, A.; Lautenbach, S.; Ruiz-Frau, A.; Schulp, N.; Seppelt, R.; Verburg, P.; Teeffelen, A.v.; Vannier, C.; et al. Pathways to bridge the biophysical realism gap in ecosystem services mapping approaches. Ecol. Indic. 2017, 74, 241–260. [Google Scholar] [CrossRef]
- Seppelt, R.; Dormann, C.F.; Eppink, F.V.; Lautenbach, S.; Schmidt, S. A quantitative review of ecosystem service studies: Approaches, shortcomings and the road ahead. J. Appl. Ecol. 2011, 48, 630–636. [Google Scholar] [CrossRef]
- Pickett, S.T. Ecological Understanding the Nature of Theory and the Theory of Nature; Elsevier/Academic Press: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Sutherland, I.J.; Villamagna, A.M.; Dallaire, C.O.; Bennett, E.M.; Chin, A.T.M.; Yeung, A.C.Y.; Lamothe, K.A.; Tomscha, S.A.; Cormier, R. Undervalued and under pressure: A plea for greater attention toward regulating ecosystem services. Ecol. Indic. 2018, 94, 23–32. [Google Scholar] [CrossRef]
- Perrotti, D.; Iuorio, O. Green Infrastructure in the Space of Flows: An Urban Metabolism Approach to Bridge Environmental Performance and User’s Wellbeing. In Planning Cities with Nature; Springer: Berlin/Heidelberg, Germany, 2019; pp. 265–277. [Google Scholar] [CrossRef]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Dominati, E.J.; Mackay, A.D.; Bouma, J.; Green, S. An Ecosystems Approach to Quantify Soil Performance for Multiple Outcomes: The Future of Land Evaluation? Soil Sci. Soc. Am. J. 2016, 80, 438–449. [Google Scholar] [CrossRef]
- Mitchell, M.G.E.; Bennett, E.M.; Gonzalez, A. Strong and nonlinear effects of fragmentation on ecosystem service provision at multiple scales. Environ. Res. Lett. 2015, 10, 94014. [Google Scholar] [CrossRef]
- Laca, E.A. Multi-Scape Interventions to Match Spatial Scales of Demand and Supply of Ecosystem Services. Front. Sust. Food Syst. 2021, 4, 607276. [Google Scholar] [CrossRef]
- Cadenasso, M.L.; Pickett, S.T.A.; Weathers, K.C.; Jones, C.G. A Framework for a Theory of Ecological Boundaries. BioScience 2003, 53, 750–758. [Google Scholar] [CrossRef]
- Naveh, Z. Biocybernetic and thermodynamic perspectives of landscape functions and land use patterns. Landsc. Ecol. 1987, 1, 75–83. [Google Scholar] [CrossRef]
- Kremen, C. Managing ecosystem services: What do we need to know about their ecology? Ecol. Lett. 2005, 8, 468–479. [Google Scholar] [CrossRef]
- Luck, G.W.; Harrington, R.; Harrison, P.A.; Kremen, C.; Berry, P.M.; Bugter, R.; Dawson, T.R.; Bello, F.d.; Díaz, S.; Feld, C.K.; et al. Quantifying the Contribution of Organisms to the Provision of Ecosystem Services. BioScience 2009, 59, 223–235. [Google Scholar] [CrossRef]
- Andersson, E.; McPhearson, T.; Kremer, P.; Gomez-Baggethun, E.; Haase, D.; Tuvendal, M.; Wurster, D. Scale and context dependence of ecosystem service providing units. Ecosyst. Serv. 2015, 12, 157–164. [Google Scholar] [CrossRef]
- Walker, J.; Sharpe, P.J.H.; Penridge, L.K.; Wu, H. Ecological Field Theory: The Concept and Field Tests. Vegetatio 1989, 83, 81–95. [Google Scholar] [CrossRef]
- Li, B.-L.; Wu, H.-i.; Zou, G. Self-thinning rule: A causal interpretation from ecological field theory. Ecol. Model. 2000, 132, 167–173. [Google Scholar] [CrossRef]
- Wu, H.-I.; Sharpe, P.J.H.; Walker, J.; Penridge, L.K. Ecological field theory: A spatial analysis of resource interference among plants. Ecol. Model. 1985, 29, 215–243. [Google Scholar] [CrossRef]
- Unwin, D.J. Effects, First and Second-Order. In Encyclopedia of Geographic Information Science; Kemp, K., Ed.; SAGE Publications, Incorporated: Thousand Oaks, CA, USA, 2007; p. 123. [Google Scholar]
- Bailey, T.C.; Gatrell, A.C. Interactive Spatial Data Analysis; Longman Scientific & Technical Harlow Essex, England: Harlow, UK, 1995. [Google Scholar]
- Meron, E. Nonlinear Physics of Ecosystems; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2015. [Google Scholar]
- Wu, J.; Loucks, O.L. From Balance of Nature to Hierarchical Patch Dynamics: A Paradigm Shift in Ecology. Q. Rev. Biol. 1995, 70, 439–466. [Google Scholar] [CrossRef]
- Capra, F. The Systems View of Life:A Unifying Vision; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Schiere, J.B.; Gregorini, P. Complexity, Crash and Collapse of Chaos: Clues for Designing Sustainable Systems, with Focus on Grassland-Based Systems. Sustainability 2023, 15, 4356. [Google Scholar] [CrossRef]
- Zardo, L.; Geneletti, D.; Pérez-Soba, M.; Van Eupen, M. Estimating the Cooling Capacity of Green Infrastructures to Support Urban Planning. Ecosyst. Serv. 2017, 26, 225–235. [Google Scholar] [CrossRef]
- Phillips, C.J.; Ekanayake, J.C.; Marden, M. Root site occupancy modelling of young New Zealand native plants: Implications for soil reinforcement. Plant Soil 2011, 346, 201–214. [Google Scholar] [CrossRef]
- Lindborg, R.; Gordon, L.J.; Malinga, R.; Bengtsson, J.; Peterson, G.; Bommarco, R.; Deutsch, L.; Gren, Å.; Rundlöf, M.; Smith, H.G. How spatial scale shapes the generation and management of multiple ecosystem services. Ecosphere 2017, 8, e01741. [Google Scholar] [CrossRef]
- Eigenbrod, F. Redefining Landscape Structure for Ecosystem Services. Curr. Landsc. Ecol. Rep. 2016, 1, 80–86. [Google Scholar] [CrossRef]
- Wiens, J.A. Spatial Scaling in Ecology. Funct. Ecol. 1989, 3, 385–397. [Google Scholar] [CrossRef]
- Mastrangelo, M.E.; Weyland, F.; Villarino, S.H.; Barral, M.P.; Nahuelhual, L.; Laterra, P. Concepts and Methods for Landscape Multifunctionality and a Unifying Framework based on Ecosystem Services. Landsc. Ecol. 2014, 29, 345–358. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.A.; Ene, E. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps; University of Massachusetts: Amherst, MA, USA, 2012. [Google Scholar]
- Meurk, C.; Hall, G.; Parkes, J. Options for enhancing forest biodiversity across New Zealand’s managed landscapes based on ecosystem modelling and spatial design. N. Z. J. Ecol. 2006, 30, 131–146. [Google Scholar]
- Hamada, S.; Tanaka, T.; Ohta, T. Impacts of land use and topography on the cooling effect of green areas on surrounding urban areas. Urban For. Urban Green. 2013, 12, 426–434. [Google Scholar] [CrossRef]
- Vidrih, B.; Medved, S. Multiparametric model of urban park cooling island. Urban For. Urban Green. 2013, 12, 220–229. [Google Scholar] [CrossRef]
- Cochrane, M.A.; Laurance, W.F. Synergisms among Fire, Land Use, and Climate Change in the Amazon. Ambio 2008, 37, 522–527. [Google Scholar] [CrossRef]
- Laurance, W.F. Theory meets reality: How habitat fragmentation research has transcended island biogeographic theory. Biol. Conserv. 2008, 141, 1731–1744. [Google Scholar] [CrossRef]
- Chang, C.-R.; Li, M.-H.; Chang, S.-D. A preliminary study on the local cool-island intensity of Taipei city parks. Landsc. Urban Plan. 2007, 80, 386–395. [Google Scholar] [CrossRef]
- Cao, X.; Onishi, A.; Chen, J.; Imura, H. Quantifying the cool island intensity of urban parks using ASTER and IKONOS data. Landsc. Urban Plan. 2010, 96, 224–231. [Google Scholar] [CrossRef]
- Eder, F.; Serafimovich, A.; Foken, T. Coherent Structures at a Forest Edge: Properties, Coupling and Impact of Secondary Circulations. Bound.-Layer Meteorol. 2013, 148, 285–308. [Google Scholar] [CrossRef]
- Huang, J.; Cassiani, M.; Albertson, J.D. Coherent Turbulent Structures Across a Vegetation Discontinuity. Bound.-Layer Meteorol. 2011, 140, 1–22. [Google Scholar] [CrossRef]
- Jauregui, E. Influence of a large urban park on temperature and convective precipitation in a tropical city. Energy Build. 1990, 15, 457–463. [Google Scholar] [CrossRef]
- Yang, C.; Li, A.; Gao, X.; Ren, T. Interaction of the thermal plumes generated from two heat sources of equal strength in a naturally ventilated space. J. Wind Eng. Ind. Aerodyn. 2020, 198, 104085. [Google Scholar] [CrossRef]
- Martini, A.; Biondi, D.; Batista, A.C. Distance and Intensity of Microclimatic Influence Provided by Urban Forest Typologies. Floresta Ambiente 2018, 25. [Google Scholar] [CrossRef]
- Lin, B.-S.; Lin, C.-T. Preliminary study of the influence of the spatial arrangement of urban parks on local temperature reduction. Urban For. Urban Green. 2016, 20, 348–357. [Google Scholar] [CrossRef]
- Honjo, T.; Takakura, T. Simulation of thermal effects of urban green areas on their surrounding areas. Energy Build. 1990, 15, 443–446. [Google Scholar] [CrossRef]
- Isbell, F.; Gonzalez, A.; Loreau, M.; Cowles, J.; Díaz, S.; Hector, A.; MacE, G.M.; Wardle, D.A.; O’Connor, M.I.; Duffy, J.E.; et al. Linking the influence and dependence of people on biodiversity across scales. Nature 2017, 546, 65–72. [Google Scholar] [CrossRef]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of Biodiversity on Ecosystem Functioning: A Consensus of Current Knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Manning, P.; Van Der Plas, F.; Soliveres, S.; Allan, E.; Maestre, F.T.; Mace, G.; Whittingham, M.J.; Fischer, M. Redefining ecosystem multifunctionality. Nat. Ecol. Evol. 2018, 2, 427–436. [Google Scholar] [CrossRef]
- Haddaway, N.R.; Brown, C.; Eales, J.; Eggers, S.; Josefsson, J.; Kronvang, B.; Randall, N.P.; Uusi-Kämppä, J. The multifunctional roles of vegetated strips around and within agricultural fields. Environ. Evid. 2018, 7, 14. [Google Scholar] [CrossRef]
- Butler, S.J.; Vickery, J.A.; Norris, K. Farmland Biodiversity and the Footprint of Agriculture. Science 2007, 315, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.; Meurk, C. Protecting and Restoring Our Natural Heritage—A Practical Guide; Department of Conservation: Wellington, New Zealand, 2001. [Google Scholar]
- Park, G.N. New Zealand as Ecosystems: The Ecosystem Concept as a Tool for Environmental Management and Conservation; Department of Conservation: Wellington, New Zealand, 2000. [Google Scholar]
- MacArthur, R.H. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Palmer, M.W. The Coexistence of Species in Fractal Landscapes. Am. Nat. 1992, 139, 375–397. [Google Scholar] [CrossRef]
- Meurk, C.D.; Sullivan, J.; McWilliam, W. Vegetation History and Dynamics in New Zealand: Future Scenarios and Improved Trajectories Towards Restoring Natural Patterns; Springer International Publishing: Cham, Switzerland, 2016; pp. 517–528. [Google Scholar] [CrossRef]
- Higgins, P.J.; Peter, J.M.; Cowling, S.J. Handbook of Australian, New Zealand & Antarctic Birds; Oxford University Press: Melbourne, Australia, 1990. [Google Scholar]
- Fitter, J. A Field Guide to the Birds of New Zealand; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Coleman, G. Nest Site Selection of the New Zealand Fantail (Rhipidura fugilinosa) on South Island Production Land; University of Otago: Dunedin, New Zealand, 2008. [Google Scholar]
- Powlesland, R.G. New Zealand Fantail|Pīwakawaka. Available online: www.nzbirdsonline.org.nz (accessed on 12 December 2022).
- Howe, R.W. Local dynamics of bird assemblages in small forest habitat islands in Australia and North America. Ecology 1984, 65, 1585–1601. [Google Scholar] [CrossRef]
- Berry, L. Edge effects on the distribution and abundance of birds in a southern Victorian forest. Wildl. Res. 2001, 28, 239–245. [Google Scholar] [CrossRef]
- Spurr, E.B.; Borkin, K.M.; Rod, S. Use of radio telemetry to determine home range and movements of the bellbird (Anthornis melanura)—A feasibility study. Notornis 2010, 57, 63–70. [Google Scholar]
- Anderson, S.H.; Craig, J.L. Breeding biology of bellbirds (Anthornis melanura) on Tiritiri Matangi Island. Notornis 2003, 50, 75–82. [Google Scholar]
- Spurr, E.B.; Crossland, A.C.; Sagar, P.M. Increased abundance of the bellbird (Anthornis melanura) in Christchurch, New Zealand. Notornis 2014, 61, 67–74. [Google Scholar]
- Dent, J.M. Information Use during Foraging by New Zealand Bellbirds (Anthornis melanura): A Thesis Submitted in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy at Lincoln University. Ph.D. Thesis, Lincoln University, Lincoln, New Zealand, 2019. [Google Scholar]
- Del Hoyo, J.; Elliott, A.; Christie, D.A. Handbook of the Birds of the World; Lynx Edicions: Barcelona, Spain, 2006; Volume 11. [Google Scholar]
- Valente, J.J.; LeGrande-Rolls, C.L.; Rivers, J.W.; Tucker, A.M.; Fischer, R.A.; Betts, M.G. Conspecific attraction for conservation and management of terrestrial breeding birds: Current knowledge and future research directions. Ornithol. Appl. 2021, 123, duab007. [Google Scholar] [CrossRef]
- Schowalter, T.D. Population Dynamics. In Insect Ecology, 2nd ed.; Schowalter, T.D., Ed.; Academic Press: Burlington, NJ, USA, 2006; pp. 153–177. [Google Scholar] [CrossRef]
- Aubier, T.G. Positive density dependence acting on mortality can help maintain species-rich communities. eLife 2020, 9, e57788. [Google Scholar] [CrossRef] [PubMed]
- Matthysen, E. Density-dependent dispersal in birds and mammals. Ecography 2005, 28, 403–416. [Google Scholar] [CrossRef]
- Peterjohn, W.T.; Correll, D.L. Nutrient dynamics in an agricultural watershed: Observations on the role of a riparian forest. Ecology 1984, 65, 1466–1475. [Google Scholar] [CrossRef]
- Vought, L.B.M.; Dahl, J.; Lauge Pedersen, C.; Lacoursiere, J.O. Nutrient retention in riparian ecotones. Ambio 1994, 23, 342–348. [Google Scholar]
- Mayer, P.M.; Reynolds, S.K.; McCutchen, M.D.; Canfield, T.J. Meta-Analysis of Nitrogen Removal in Riparian Buffers. J. Environ. Qual. 2007, 36, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Marshall, C.J.; Beck, M.R.; Garrett, K.; Barrell, G.K.; Al-Marashdeh, O.; Gregorini, P. Grazing dairy cows with low milk urea nitrogen breeding values excrete less urinary urea nitrogen. Sci. Total Environ. 2020, 739, 139994. [Google Scholar] [CrossRef] [PubMed]
- Beukes, P.C.; Scarsbrook, M.R.; Gregorini, P.; Romera, A.J.; Clark, D.A.; Catto, W. The relationship between milk production and farm-gate nitrogen surplus for the Waikato region, New Zealand. J. Environ. Manag. 2012, 93, 44–51. [Google Scholar] [CrossRef]
- Di, H.J.; Cameron, K.C. Nitrate leaching in temperate agroecosystems: Sources, factors and mitigating strategies. Nutr. Cycl. Agroecosyst. 2002, 64, 237–256. [Google Scholar] [CrossRef]
- Parkyn, S. Review of Riparian Buffer Zone Effectiveness; Ministry of Agriculture and Forestry: Wellington, New Zealand, 2004. [Google Scholar]
- Clode, P.L.; Kilburn, M.R.; Jones, D.L.; Stockdale, E.A.; Cliff, J.B.; Herrmann, A.M.; Murphy, D.V. In Situ Mapping of Nutrient Uptake in the Rhizosphere Using Nanoscale Secondary Ion Mass Spectrometry. Plant Physiol 2009, 151, 1751–1757. [Google Scholar] [CrossRef]
- Finzi, A.C.; Abramoff, R.Z.; Spiller, K.S.; Brzostek, E.R.; Darby, B.A.; Kramer, M.A.; Phillips, R.P. Rhizosphere processes are quantitatively important components of terrestrial carbon and nutrient cycles. Glob. Change Biol. 2015, 21, 2082–2094. [Google Scholar] [CrossRef]
- Wu, L.; McGechan, M.B.; McRoberts, N.; Baddeley, J.A.; Watson, C.A. SPACSYS: Integration of a 3D root architecture component to carbon, nitrogen and water cycling—Model description. Ecol. Model. 2007, 200, 343–359. [Google Scholar] [CrossRef]
- Tobin, B.; Čermák, J.; Chiatante, D.; Danjon, F.; Di Iorio, A.; Dupuy, L.; Eshel, A.; Jourdan, C.; Kalliokoski, T.; Laiho, R.; et al. Towards developmental modelling of tree root systems. Plant Biosyst. 2007, 141, 481–501. [Google Scholar] [CrossRef]
- Pressland, A.J. Productivity and Management of Mulga in South-Western Queensland in Relation to Tree Structure and Density. Aust. J. Bot. 1975, 23, 965–976. [Google Scholar] [CrossRef]
- Saint Cast, C.; Meredieu, C.; Défossez, P.; Pagès, L.; Danjon, F. Modelling root system development for anchorage of forest trees up to the mature stage, including acclimation to soil constraints: The case of Pinus pinaster. Plant Soil 2019, 439, 405–430. [Google Scholar] [CrossRef]
- Marden, M.; Lambie, S.; Phillips, C. Biomass and root attributes of eight of New Zealand’s most common indigenous evergreen conifer and broadleaved forest species during the first 5 years of establishment. N. Z. J. For. Sci. 2018, 48, 9. [Google Scholar] [CrossRef]
- Spiekermann, R.I.; McColl, S.; Fuller, I.; Dymond, J.; Burkitt, L.; Smith, H.G. Quantifying the influence of individual trees on slope stability at landscape scale. J. Environ. Manag. 2021, 286, 112194. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.J.; Marden, M.; Lambie, S.M. Observations of “coarse” root development in young trees of nine exotic species from a New Zealand plot trial. N. Z. J. For. Sci. 2015, 45, 1. [Google Scholar] [CrossRef]
- Gress, S.E.; Nichols, T.D.; Northcraft, C.C.; Peterjohn, W.T. Nutrient limitation in soils exhibiting differing nitrogen availabilities: What lies beyond nitrogen saturation? Ecology 2007, 88, 119–130. [Google Scholar] [CrossRef]
- Blake, G.R.; Steinhardt, G.C.; Pombal, X.P.; Muñoz, J.C.N.; Cortizas, A.M.; Arnold, R.W.; Schaetzl, R.J.; Stagnitti, F.; Parlange, J.Y.; Steenhuis, T.S.; et al. Plant Nutrients. In Encyclopedia of Soil Science; Chesworth, W., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 2008; pp. 560–571. [Google Scholar] [CrossRef]
- Ågren, G.I.; Bosatta, E. Nitrogen saturation of terrestrial ecosystems. Environ. Pollut. 1988, 54, 185–197. [Google Scholar] [CrossRef]
- Valenzuela-Estrada, L.R.; Vera-Caraballo, V.; Ruth, L.E.; Eissenstat, D.M. Root anatomy, morphology, and longevity among root orders in Vaccinium corymbosum (Ericaceae). Am. J. Bot. 2008, 95, 1506–1514. [Google Scholar] [CrossRef]
- Atkinson, D. The growth, activity and distribution of the fruit tree root system. Plant Soil 1983, 71, 23–35. [Google Scholar] [CrossRef]
- Landsberg, J.J.; Fowkes, N.D. Water Movement Through Plant Roots. Ann. Bot. 1978, 42, 493–508. [Google Scholar] [CrossRef]
- Burns, I.G. Influence of the spatial distribution of nitrate on the uptake of N by plants: And review and model for rooting depth. J. Soil Sci. 1980, 31, 155–173. [Google Scholar] [CrossRef]
- Dunbabin, V.; Rengel, Z.; Diggle, A.J. Simulating Form and Function of Root Systems: Efficiency of Nitrate Uptake Is Dependent on Root System Architecture and the Spatial and Temporal Variability of Nitrate Supply. Funct. Ecol. 2004, 18, 204–211. [Google Scholar] [CrossRef]
- Kuusemets, V.; Mander, Ü.; Lõhmus, K.; Ivask, M. Nitrogen and phosphorus variation in shallow groundwater and assimilation in plants in complex riparian buffer zones. Water. Sci. Technol. 2001, 44, 615–622. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Zak, D.R.; Loya, W.M.; Karberg, N.J.; King, J.S.; Burton, A.J. Chapter 7—The Contribution of Root—Rhizosphere Interactions to Biogeochemical Cycles in a Changing World; Elsevier Inc.: Amsterdam, The Netherlands, 2007; pp. 155–178. [Google Scholar] [CrossRef]
- Yan, X.-L.; Jia, L.; Dai, T. Fine root morphology and growth in response to nitrogen addition through drip fertigation in a Populus × euramericana “Guariento” plantation over multiple years. Ann. For. Sci. 2019, 76, 13. [Google Scholar] [CrossRef]
- Burkhard, B.; Maes, J. Mapping Ecosystem Services; Pensoft Publishers: Sofia, Bulgaria, 2017. [Google Scholar]
- Eigenbrod, F.; Armsworth, P.R.; Anderson, B.J.; Heinemeyer, A.; Gillings, S.; Roy, D.B.; Thomas, C.D.; Gaston, K.J. The impact of proxy-based methods on mapping the distribution of ecosystem services. J. Appl. Ecol. 2010, 47, 377–385. [Google Scholar] [CrossRef]
- Rykiel, E.J. Testing ecological models: The meaning of validation. Ecol. Model. 1996, 90, 229–244. [Google Scholar] [CrossRef]
- Haycock, N.E.; Pinay, G. Groundwater Nitrate Dynamics in Grass and Poplar Vegetated Riparian Buffer Strips during the Winter. J. Environ. Qual. 1993, 22, 273–278. [Google Scholar] [CrossRef]
- Lamy, T.; Liss, K.N.; Gonzalez, A.; Bennett, E.M. Landscape structure affects the provision of multiple ecosystem services. Environ. Res. Lett. 2016, 11, 124017. [Google Scholar] [CrossRef]
- Spake, R.; Lasseur, R.; Crouzat, E.; Bullock, J.M.; Lavorel, S.; Parks, K.E.; Schaafsma, M.; Bennett, E.M.; Maes, J.; Mulligan, M.; et al. Unpacking ecosystem service bundles: Towards predictive mapping of synergies and trade-offs between ecosystem services. Glob. Environ. Change 2017, 47, 37–50. [Google Scholar] [CrossRef]
- Jones, K.B.; Zurlini, G.; Kienast, F.; Petrosillo, I.; Edwards, T.; Wade, T.G.; Li, B.-l.; Zaccarelli, N. Informing landscape planning and design for sustaining ecosystem services from existing spatial patterns and knowledge. Landsc. Ecol. 2012, 28, 1175–1192. [Google Scholar] [CrossRef]
- Kupfer, J.A. Landscape ecology and biogeography: Rethinking landscape metrics in a post-FRAGSTATS landscape. Prog. Phys. Geogr. 2012, 36, 400–420. [Google Scholar] [CrossRef]
- Koch, E.W.; Barbier, E.B.; Silliman, B.R.; Reed, D.J.; Perillo, G.M.; Hacker, S.D.; Granek, E.F.; Primavera, J.H.; Muthiga, N.; Polasky, S.; et al. Non-linearity in ecosystem services: Temporal and spatial variability in coastal protection. Front. Ecol. Environ. 2009, 7, 29–37. [Google Scholar] [CrossRef]
- Burkhard, B.; Fath, B.D.; Müller, F. Adapting the adaptive cycle: Hypotheses on the development of ecosystem properties and services. Ecol. Model. 2011, 222, 2878–2890. [Google Scholar] [CrossRef]
- Troll, C. The geographic landscape and its investigation. In Foundation Papers in Landscape Ecology; Wiens, J.A., Moss, M.R., Turner, M.G., Mladenoff, D.J., Eds.; Columbia University Press: New York, NY, USA, 2007; pp. 71–101. [Google Scholar]
- Arts, B.; Buizer, M.; Horlings, L.; Ingram, V.; van Oosten, C.; Opdam, P. Landscape Approaches: A State-of-the-Art Review. Annu. Rev. Environ. Resour. 2017, 42, 439–463. [Google Scholar] [CrossRef]
- Turner, M.G. Landscape Ecology: The Effect of Pattern on Process. Annu. Rev. Ecol. Syst. 1989, 20, 171–197. [Google Scholar] [CrossRef]
- Cohen, J.; Stewart, I. The Collapse of Chaos: Discovering Simplicity in a Complex World; Penguin: London, UK, 2000. [Google Scholar]
- Larson-Praplan, S.; George, M.R.; Buckhouse, J.C.; Laca, E.A. Spatial and temporal domains of scale of grazing cattle. Anim. Prod. Sci. 2015, 55, 284–297. [Google Scholar] [CrossRef]
- Ryszkowski, L.; French, N.R.; Kędziora, A. Dynamics of an Agricultural Landscape; Zaklad Badan Srodowiska Rolniczego i Lesnego PAN: Poznan, Poland, 1996. [Google Scholar]
- Cortinovis, C.; Geneletti, D. A Performance-based Planning Approach Integrating Supply and Demand of Urban Ecosystem Services. Landsc. Urban Plan. 2020, 201, 103842. [Google Scholar] [CrossRef]
- Biggs, R.; Schlüter, M.; Biggs, D.; Bohensky, E.L.; BurnSilver, S.; Cundill, G.; Dakos, V.; Daw, T.M.; Evans, L.S.; Kotschy, K.; et al. Toward Principles for Enhancing the Resilience of Ecosystem Services. Annu. Rev. Environ. Resour. 2012, 37, 421–448. [Google Scholar] [CrossRef]
- Deilami, K.; Kamruzzaman, M.; Liu, Y. Urban heat island effect: A systematic review of spatio-temporal factors, data, methods, and mitigation measures. ITC J. 2018, 67, 30–42. [Google Scholar] [CrossRef]
- Bateman, I.J.; Anderson, K.; Argles, A.; Belcher, C.; Betts, R.A.; Binner, A.; Brazier, R.E.; Cho, F.H.T.; Collins, R.M.; Day, B.H.; et al. A review of planting principles to identify the right place for the right tree for ‘net zero plus’ woodlands: Applying a place-based natural capital framework for sustainable, efficient and equitable (SEE) decisions. People Nat. 2023, 5, 271–301. [Google Scholar] [CrossRef]
- Di Sacco, A.; Hardwick, K.A.; Blakesley, D.; Brancalion, P.H.S.; Breman, E.; Cecilio Rebola, L.; Chomba, S.; Dixon, K.; Elliott, S.; Ruyonga, G.; et al. Ten golden rules for reforestation to optimize carbon sequestration, biodiversity recovery and livelihood benefits. Glob. Change Biol. 2021, 27, 1328–1348. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.G.; Gardner, R.H. Landscape Ecology in Theory and Practice: Pattern and Process; Springer: New York, NY, USA, 2015. [Google Scholar]
- Wu, J.; Jones, K.; Li, H.; Loucks, O. Scaling and Uncertainty Analysis in Ecology: Methods and Applications; Springer: Dordrecht, The Netherlands, 2006. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morris, R.; Davis, S.; Grelet, G.-A.; Doscher, C.; Gregorini, P. A Model for Spatially Explicit Landscape Configuration and Ecosystem Service Performance, ESMAX: Model Description and Explanation. Sustainability 2024, 16, 876. https://doi.org/10.3390/su16020876
Morris R, Davis S, Grelet G-A, Doscher C, Gregorini P. A Model for Spatially Explicit Landscape Configuration and Ecosystem Service Performance, ESMAX: Model Description and Explanation. Sustainability. 2024; 16(2):876. https://doi.org/10.3390/su16020876
Chicago/Turabian StyleMorris, Richard, Shannon Davis, Gwen-Aëlle Grelet, Crile Doscher, and Pablo Gregorini. 2024. "A Model for Spatially Explicit Landscape Configuration and Ecosystem Service Performance, ESMAX: Model Description and Explanation" Sustainability 16, no. 2: 876. https://doi.org/10.3390/su16020876
APA StyleMorris, R., Davis, S., Grelet, G.-A., Doscher, C., & Gregorini, P. (2024). A Model for Spatially Explicit Landscape Configuration and Ecosystem Service Performance, ESMAX: Model Description and Explanation. Sustainability, 16(2), 876. https://doi.org/10.3390/su16020876