Abstract
The growth of seedlings depends on many factors: the availability of light, water, and minerals, as well as the type and physical properties of the substrate, including its density. The effect of different levels of compaction of the peat-perlite substrate on the growth of European beech (Fagus sylvatica L.) seedlings in container nursery has not been investigated so far. In presented research, nine variants of peat-perlite substrate compaction (in three replications), with the actual bulk density ranging between 0.196 and 0.317 g·cm−3, were prepared in Hiko V265 nursery trays. European beech seeds were sown in the trays, and a seedling was grown in the production field. After the cultivation period, selected parameters were measured. The results reveal that substrate in a wide range of compaction (dry bulk density: 0.078–0.127 g·cm−3) had an impact on the height of seedlings, root dry matter, and the mean length of coarse roots and fine roots. The best shoot-to-root ratio and sturdiness quotient were observed in the seedlings growing on the substrate with the lowest compaction. Based on the results, compaction at the level of 0.196 g·cm−3 is recommended for the cultivation of European beech in Hiko V265 nursery trays.
1. Introduction
European beech is an Atlantic climate species found throughout central and west Europe [1]. In Poland, coniferous monocultures (pine, spruce) have been intensively restructured due to the declining health and quality of trees. Between 2005 and 2018, the area contribution of deciduous species in Poland increased from 35% to 37.3% (an increase of 0.5%—from 5.5% to 6.0% in the case of beech) [2]. Regeneration in this region is mainly carried out using seedlings bred in forest nurseries (86% of cultivation area) [3]. The use of nursery trays for the production of seedlings allows regulating the breeding conditions, such as irrigation and fertilization, and the substrate parameters [4]. Seedlings bred in trays have a good quality, contributing to their adaptation after planting on the culture. Nursery substrate compaction is a significant factor influencing the growth and quality of seedlings [5,6,7,8,9].
The major component used for tray seedling production is a substrate consisting of peat and a small addition of perlite. However, the influence of a wide range different levels of substrate compaction on the growth of seedlings in container nurseries has not been investigated so far. Seedling growth is influenced by the physical properties of the applied substrate, including its compaction level, which is typically expressed as bulk density. Substrate compaction determines the air–water ratio, including the availability of water and nutrients [10,11,12], and microbial activity [13,14]. Several authors have studied the effect of soil compaction on the growth and development of plants. Kozlowski [15] characterized the impact of soil compaction on tree growth. Lipiec et al. [16] analyzed the changes in the growth and function of roots based on soil compaction. Similarly, Passioura [17] investigated how soil compaction can influence root growth. In another study, Hamza and Anderson [18] described the nature and causes of excessive soil compaction on plants and possible solutions to reduce soil compaction. In 2002, the European Commission recognized soil compaction as one of the leading environmental hazards [19], and various authors have confirmed this [20,21,22,23,24,25,26,27]. Their results indicate that soil’s excessive and inadequate bulk density can affect plant development depending on the species and substrate type. Studies on soil compaction have been conducted on Nicotiana L. [28], several grass species [29], nonforest trees, such as Malus Mill. [30] and Ailanthus altissima Mill. Swingle [31], and forest tree species of low significance for Polish forest management, such as Pinus contorta Dougl. ex Loud [32], Pseudotsuga menziesii (Mirb.) Franco [33], Pinus nigra Arn. [5], Pinus ponderosa var. ponderosa Dougl. ex Laws, Pinus taeda L., Pinus echinata Mill. [34], Quercus rubra L. [35], Quercus coccinea Muencch [36], and Quercus pyrenaica Willd. [37]. In studies from Poland, the effect of soil compaction on seedlings growth has been analyzed for Fagus sylvatica L. under laboratory conditions [7], for Quercus petraea (Matt.) Liebl. on soil collected from a nursery bed [38], and for Pinus sylvestris L. in trays containing bark-based substrate [39]. Kormanek et al. [40] demonstrated the effect of soil compaction on the growth of F. sylvatica.
The applicability of seedling for planting on a culture can be assessed using different standards based on the values of single traits. As a rule, it is assumed that higher seedlings are better for forest culture compared with smaller seedlings and ground cover plants [41]. The adaptive potential of seedlings is characterized by synthetic indicators [9,42,43,44,45]—namely, seedling sturdiness quotient (SQ) and stem-to-root ratio (S/R). SQ is a measure of seedling stability in the culture (resistance to wind and drought), with smaller values of SQ corresponding to more sturdy seedlings. According to Roller [46], the value of SQ depends on the species and type of nursery tray used for breeding. The S/R value determines the balance between the area of transpiration and the area of water absorption. The SQ of European beech seedlings was calculated based on the limit values of height and diameter at the root collar recommended by the Polish standard PN-R-67025 [47]. In ground nurseries, for first-class seedlings, the value of SQ is 6.3, and for second-class seedlings, it is 6.7. In the absence of recommended values of SQ for container nurseries, for the purpose of this experiment we set SQ to be at most 6.5. For seedlings growing in soil, the S/R value should not exceed 3:1, while for seedlings growing in trays, it should be 2:1 [42].
The research on the impact of compaction most often concerns agriculture; there is scarcely any research on the peat substrate compaction (especially in such a wide range of compactions) in container nurseries. The density level used in container nurseries is chosen based on the intuition and experience of forest nursery employees. The same level of compaction is most often used for each species grown at one nursery. For specific species, there is insufficient research on whether very high or very low densities affect the growth of the species and what is the optimal compaction of the perlite–peat substrate.
In the present study, the following hypothesis was tested for European beech (Fagus sylvatica L.) seedlings grown in Hiko V265 containers: there is an optimum peat-perlite compaction at which the grown seedlings will have the desirable growth parameters (allometric) for good adaptation on the crop—i.e., the value of the S/R should equal to or be below 2 (S/R ≤ 2:1), and the SQ should be below 6.5 (SQ ≤ 6.5).
2. Materials and Methods
2.1. Levels of Compaction
The experiment was performed on the substrate obtained from a nursery farm in Nędza, Rudy Raciborskie Forest District (GPS N 50.167964, E 18.3138334,17). The substrate contained a high amount of peat (93%) and a low amount of perlite (7%), with dolomite added (3 kg m−3 substrate) to achieve a pH of 5.5. The proportion of perlite to peat is selected individually by substrate producers for each batch of the substrate in accordance with the guidelines [4]. The peat used in the substrate was characterized by a maximum decomposition level of 15% and organic matter content of >85%. Its granulometric composition was as follows: 10.1–20 mm, 2.5%; 4.1–10 mm, 12.5%; 2.1–4.0 mm, 12.5%; <2.0 mm, 72.5%. The peat had an air capacity of 15–25% (v/v), water capacity (at 10 cm H2O) of 70–80% (v/v), total porosity of 85–95% (v/v), humidity of approximately 65%, pH at H2O of 3.0–4.5, and salinity of up to 0.12 mS∙cm−1.
Polyethylene Hiko V265 trays, measuring 352 × 216 × 150 mm (L × W × H) and consisting of 28 cells (each with 265 cm3 volume), were used for breeding. The cells in the tray taper down and are equipped with vertical guides for the root system. A total of 9 variants with different levels of peat-perlite substrate compaction were prepared. The minimum bulk density (V1 variant) was achieved by filling by hand a single cell with un-compacted, loose substrate of 60% humidity, which was subsequently poured out and weighed. The procedure was repeated 3 times, and after averaging for the V1 variant, a substrate with a weight of 52.0 ± 0.2 g was obtained. The substrate in the variant with maximum compaction (V9) was compacted using a wooden stamp until the cell in the tray was completely filled; next, the substrate was poured out of the cell and weighed. The procedure was repeated 3 times, and the average weight of the three samples was calculated. A variant with maximum compaction (V9) with a weight of 84.0 ± 0.3 g was obtained. For variants V2–V8, the substrate mass was calculated by equally dividing the mass of extreme compaction variants ((84.0 g − 52.0 g): 8 = 4) and an increase in weight in subsequent variants by 4 g (Table 1). Next, for each variant, the substrate was filled in 3 trays (replications) by pouring the calculated substrate mass to a cell, weighed on the analytical scale with an accuracy of ±0.1 g. Each cell of the tray was filled by hand.

Table 1.
Wet substrate weight used to fill cells in individual experimental variants and its corresponding actual and dry bulk density and porosity.
2.2. Plant Growth
After tray preparation (filling with substrate), beech seeds were manually sown (April 2015). A total of 756 seeds were sown in compaction variants, with 84 seeds per variant (28 cells in tray × 3 replications). After sowing, the trays were placed in a vegetation hall for 4 weeks and then transported to an outdoor production field. The trays (27 pcs.) were marked with the variant name and replication number and randomly distributed on a stand (pallet) placed among other seedlings of the species in the middle section of the production field. The seedlings were allowed to grow for a period of 5 months following the tray nursery procedure [4]. In the process of growing seedlings, foliar fertilization was carried out along with irrigation (5 times in the tent, 10 times in the production field (total: 0.270 dm3∙m−2)—Floralesad and once Florasin K, at the end of the vegetation season). Floralesad fertilizer consisted of (in g∙dm−3) N 103.1, N-NO2 0.0214, N-NO3− 16.369, N-NH4+ 2.602, N-NH2 84.107, P 17.231, K 47.423, Mg 3.567, Ca 0.737, Na 0.28, S 1.24, B 1.107, Cu 0.123, Fe 1.04, Mn 0.281, Mo 0.048, and Zn 0.231, and Florasin K 500 consisted of (in g∙dm−3) K 420.75, Na 2.2, and Ca 0.003. Throughout the vegetative season, the conductivity of the aqueous fertilizer solution was maintained at approximately 600 µS∙cm−1. After the fertilizer had been poured into the fertilizer distribution on irrigation ramp, the conductivity test was made by hand tester on a sample. The seedlings were subjected to protective treatments (plant protection products were used) and care (manual weeding). During the rearing period, the precipitation was only at 78 mm. A HAB-T1 BCC sprinkler ramp was used to supplement the water deficit irrigation. Irrigation was carried out for 103 days, providing water up to 904 mm.
2.3. Laboratory Analysis
At the end of the production cycle, the number of bred seedlings was determined, and using electronic calipers, the root collar diameter (RCD) (±0.1 mm) and seedling height (±1 mm) were determined. SQ was calculated as the proportion of SH to RCD [9,48] as well as S/R—i.e., the ratio of dry stem weight to roots [42]. Leaves from each seedling were scanned using an Epson V800 Photo scanner, at 800 DPI resolution, and their surface was analyzed using WinFolia software (Regent Instruments Inc.). In each replication per variant, 3 seedlings closest to the mean were selected, and their root system was analyzed in detail. Selected seedlings were removed from cells, while the root ball was cut into 3 equal portions (upper (a), 0–5 cm cell depth; middle (b), 5–10 cm; lower (c), 10–15 cm). Subsequently, roots were manually separated from the substrate under running water. The roots collected from each part were scanned using the Epson V800 Photo scanner, at 800 DPI resolution. Using WinRhizo software 2015a (Regent Instruments Inc., Québec, QC, Candada), the root length was determined for the following 3 ranges of diameter: <0.5 mm (very fine), 0.5–2.0 mm (fine), and >2.0 mm (coarse) [49,50]. All parts of seedlings (assimilation apparatus, stem, roots) were dried at a temperature of 105 °C (48 h) and weighed with an accuracy of ±0.1 mg. For each variant, the contribution of seedlings that met both qualitative criteria (SQ ≤ 6.5 and S/R ≤ 2:1), as well as that of seedlings that met one of the criteria or that did not meet any of the criteria was calculated.
2.4. Statistical Analysis
The effect of compaction on the parameters of the bred seedlings was studied using single-variant analysis of variance and post hoc Tukey multiple comparison test (p = 0.05). Calculations were carried out in Statistica 13.3 software [51]. Using nonlinear regression, the variability of mean values of the tested seedling parameters in the analyzed experimental variants was visualized.
3. Results
Of the 756 beech seeds sown, 404 seedlings were bred, indicating a success rate of 53%. The remaining seeds did not germinate (22%) or withered during growth (25%). The differences in emergence success between individual variants were not statistically significant (F = 0.977; p = 0.452).
3.1. Growth
The effect of compaction variant was statistically significant for some of the tested traits of the bred nursery material (Table 2 and Table 3). The mean height of seedlings from the V6 and V9 variants, which were tallest, differed significantly from that of seedlings from the V1 variant (with the lowest height) by 7.1 cm. SH showed an increase with the increase in substrate compaction (Table 2, Figure 1). The differences in root collar diameter (RCD) and leaf surface (LA) were not statistically significant (Table 2).
Table 2.
Mean growth parameters (±SD) depending on compaction variant; SH—seedling height, RCD—root collar diameter, LA—leaf area; letters a, b, indicate significant differences between the means (Tukey test) at p = 0.05.
Table 3.
Mean values (±SD) of dry weight depending on compaction variant and mean ratio (±SD) of dry weight of aboveground parts to underground parts (shoot-to-root ratio); letters a, b, and c indicate significant differences between means (Tukey test) at p = 0.05.
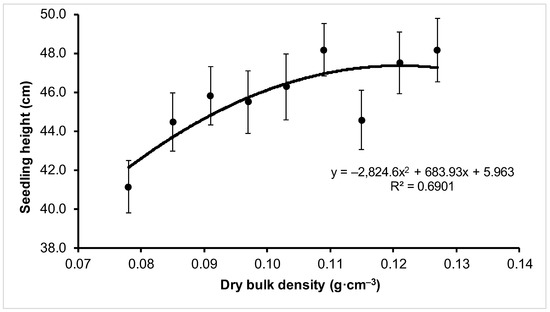
Figure 1.
Change in the mean seedling height (±SE) depending on the bulk density of peat-perlite substrate. Values of dry bulk density (in g·cm−3) for each variant: V1 = 0.078, V2 = 0.085, V3 = 0.091, V4 = 0.097, V5 = 0.103, V6 = 0.109, V7 = 0.115, V8 = 0.121, V9 = 0.127.
3.2. Dry Matter
No statistically significant differences were noted between variants with respect to the dry weight of stems and leaves. The root dry weight of the V1 variant was significantly higher than the remaining variants and was almost 2-fold higher compared with that of the V2 variant, which had the lowest dry weight among all variants. The highest dry weight of part (a) of root was obtained for the variants with extreme compaction, i.e., V1 and V9, and the lowest value for the V2 variant. The highest dry weight of parts (b) and (c) was found in the least compacted V1 variant, and lowest was found in the most compacted V9 variant. In contrast with stem and leaf dry weight, the values of root dry weight differed statistically significantly between the variants. Except for the V1 variant, the dry weight of the examined seedling parts (stems, leaves, and roots) showed an upward trend with the increase in substrate compaction (Figure 2 and Figure 3). However, high variability in the results was not observed for the medium values of compaction. The highest value of the determination coefficient (76%) was obtained for the leaf dry weight, a lower value for stem dry weight (54%), and the lowest for root dry weight (43%) (Figure 2). For roots, the value of R2 reached approximately 50% for parts (a) and (b) (Figure 3). The most pronounced influence of compaction was noted for part (c) of root, with as much as 80% of dry weight variability explained by the change in substrate compaction.
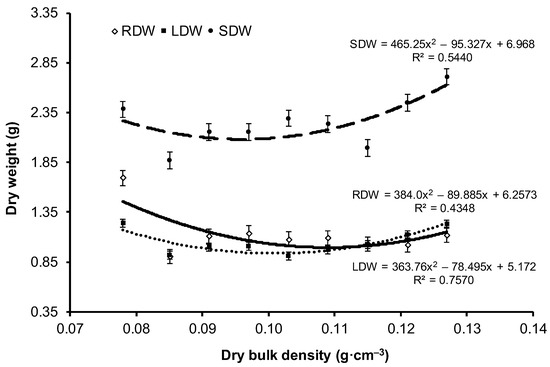
Figure 2.
Change in the mean value (±SE) of root dry weight (RDW), stem dry weight (SDW), and leaf dry weight (LDW) of European beech seedlings depending on the bulk density of peat-perlite substrate. Values of dry bulk density (in g·cm−3) for each variant: V1 = 0.078, V2 = 0.085, V3 = 0.091, V4 = 0.097, V5 = 0.103, V6 = 0.109, V7 = 0.115, V8 = 0.121, V9 = 0.127.
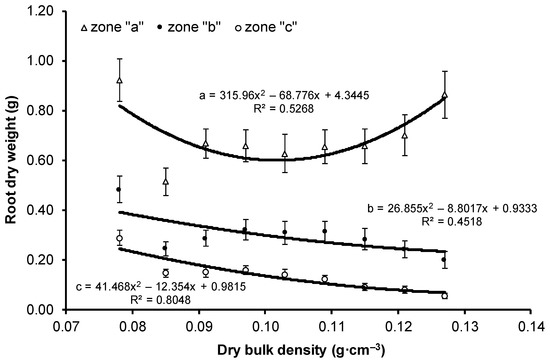
Figure 3.
Change in the mean value (±SE) of root dry weight of European beech seedlings in root ball zone (parts (a), (b), (c)) depending on the bulk density of peat-perlite substrate. Values of dry bulk density (in g·cm−3) for each variant: V1 = 0.078, V2 = 0.085, V3 = 0.091, V4 = 0.097, V5 = 0.103, V6 = 0.109, V7 = 0.115, V8 = 0.121, V9 = 0.127.
3.3. Quality Indices
Considering the ratio of SH to RCD, the best SQ was attained by seedlings from the V1 variant (7.82), whereas the least favorable result was observed for the V6 variant (10.35). The V1 variant showed significant differences from the remaining results (Figure 4).
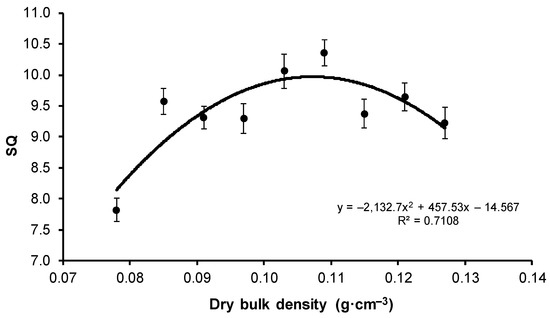
Figure 4.
Mean value of sturdiness quotient (SQ) (±SE) depending on the bulk density of peat-perlite substrate. Values of dry bulk density (in g·cm−3) for each variant: V1 = 0.078, V2 = 0.085, V3 = 0.091, V4 = 0.097, V5 = 0.103, V6 = 0.109, V7 = 0.115, V8 = 0.121, V9 = 0.127.
The lowest stem dry weight (without assimilation apparatus) to root ratio, i.e., S/R, was attained by the V1 variant (1.47), and highest by the V9 variant (2.80), which are respectively variants with the lowest and highest compaction of peat-perlite substrate (Table 4). The value of S/R increased with the increase in substrate compaction (Figure 5), and about 65% variability of the S/R ratio was explained by the change in substrate compaction.
Table 4.
Mean values of quality indicators (±SD) depending on compaction variant; SQ—sturdiness quotient, S/R—stem-to-root ratio; letters a, b, c, d, and e indicate significant differences between means (Tukey test) at p = 0.05.
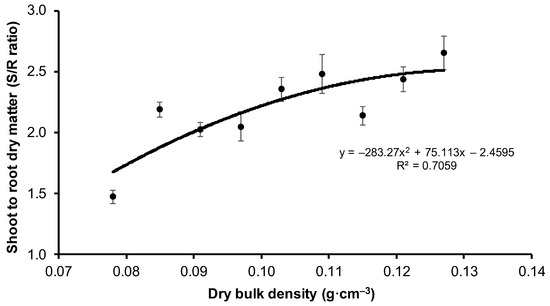
Figure 5.
S/R value (±SE) depending on the bulk density of peat-perlite substrate. Values of dry bulk density (in g·cm−3) for each variant: V1 = 0.078, V2 = 0.085, V3 = 0.091, V4 = 0.097, V5 = 0.103, V6 = 0.109, V7 = 0.115, V8 = 0.121, V9 = 0.127.
The percentage of seedlings with admissible values of SQ and S/R (i.e., not exceeding the maximum defined for this experiment) decreased with increasing substrate compaction. When the dry bulk density was above 0.091 g·cm−3 (V3) (Figure 6), the percentage of seedlings with admissible values dropped below 50%.
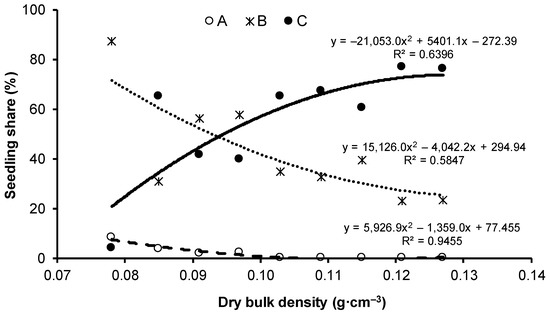
Figure 6.
Change in the share of seedlings meeting the requirement for sturdiness quotient (SQ, ≤6.5) and shoot-to-root ratio (S/R, ≤2:1); A—both met, B—only one met, C—neither met. Values of dry bulk density (in g·cm−3) for each variant: V1 = 0.078, V2 = 0.085, V3 = 0.091, V4 = 0.097, V5 = 0.103, V6 = 0.109, V7 = 0.115, V8 = 0.121, V9 = 0.127.
In total, the longest roots were obtained in seedlings from the V1 variant, and the root length of this variant differed by 1236 cm from V2 variant seedlings characterized by the shortest roots (Table 5). The root length of seedlings from the V1 variant was 39% higher than the mean and differed statistically significantly from the seedlings of other variants, whereas the root length of seedlings from the V2 variant was 42% lower than the mean root length of seedlings from all variants. Except for the V1 variant as a deviating observation, the remaining results of total seedling root length were set into a unimodal function with a single maximum observed for the V5 variant (Figure 7). The change in substrate compaction explained about 20% of root length variability.
Table 5.
Mean length (cm) of roots from individual cell levels grouped according to root diameter (mm) with mean total root length (cm) from all levels per variant ±SD. No letter indicates the lack of statistically significant differences. Letters a, and b indicate significant differences between means (Tukey test) at p = 0.05.
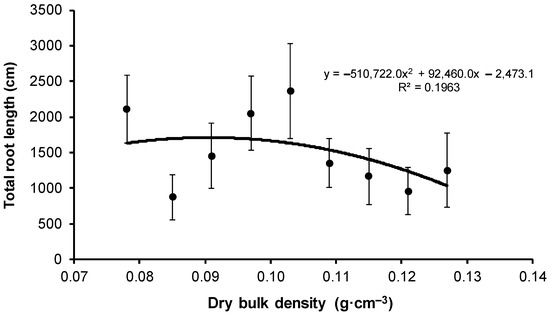
Figure 7.
Change in the mean total root length depending on the bulk density of peat-perlite substrate. Values of dry bulk density (in g·cm−3) for each variant: V1 = 0.078, V2 = 0.085, V3 = 0.091, V4 = 0.097, V5 = 0.103, V6 = 0.109, V7 = 0.115, V8 = 0.121, V9 = 0.127.
Coarse roots with the greatest diameter (>2 mm) were found to be most numerous in level (a) of the root ball, without statistically significant differences between the studied variants (Table 5). In level (b), the highest number of roots with this diameter was found for the V1 variant with the lowest compaction. The value differed significantly from the number of roots growing in the V9 variant with the highest compaction. Coarse roots were least common in level (c), and the highest number of these roots was found in the V1 variant. When the substrate compaction was higher than the mean (from V6 to V9 variants), roots with this diameter did not occur at this level. No statistically significant differences in the number of fine roots (2–0.5) in level (a) were found between variants. In levels (b) and (c), the highest number of fine roots was found for the V1 variant, and the value differed significantly from the V2, V3, and V7–V9 variants, which had the lowest number of these roots. No statistically significant differences in the number of very fine roots (≤0.5 mm) could be found at each root ball level.
4. Discussion
Research on different plant species has revealed that the growth of plants increases with the bulk density of the substrate. Alameda et al. [28] observed increased growth of Nicotiana L. seedlings in a substrate consisting of sand, silt, and gravel at a bulk density of 1.4 g·cm−3. Studies have also demonstrated that plant growth is inhibited above a certain compaction threshold. For instance, some authors reported growth inhibition of Q. rubra L. at a compaction level above 1.75 g·cm−3 [35], P. contorta Dougl. ex Loud at a compaction level above 1.7 g·cm−3 [33], and P. menziesii (Mirb.) Franco at a bulk density of 0.72 g·cm−3 [24]. The present study showed that increased substrate compaction did not have a limiting effect on the height of the aboveground part and RCD of European beech seedlings. The highest level of substrate compaction applied in the study did not cause inhibition of seedling growth. For European beach, a higher bulk density per cell would be achieved by applying a different substrate compaction procedure in the tray cells. Increased compaction would lead to the deformation of tray cells and an increase in substrate volume, which would incur higher production costs. In turn, this could not be compensated for by the enhanced quality of the produced seedlings. The study showed high SQ values (7.8–10.4) for seedlings from the tested variants, which indicates a high increase in the height of seedings, with a lower increase in thickness at the root collar. SH increased with substrate compaction and stabilized at a dry bulk density of approximately 0.11 g·cm−3. A similar course of changes was not observed for diameter at root collar. The response of beech to compaction differed from that of other species, for which a reduction in SH has been observed. These species include Quercus alba L. [52], Q. rubra L. [35], Q. petraea (Matt.) Liebl. [38], and P. contorta Dougl. ex Loud [33]. No statistically significant differences between variants were found in the leaf area of beech seedlings. Similar results were obtained in the study of Maupin and Struve [35], who reported that the leaf area of Q. rubra L. seedlings remained at the same level, independent of the compaction level of substrate.
The present study revealed that increasing substrate compaction did not have an inhibiting effect on the dry weight of beech seedlings. A contrasting result was obtained in the study of Pan and Bassuk at compaction 1.67 g/cm3 and 1.88 g/cm3 for mason sand and 1.30 g/cm3 and 1.64 g/cm3 sandy loam [31], in which the total dry weight of A. altissima Mill. Swingle seedlings was approximately 50% lower in variants with high compaction. Compared with the tested soils types, peat is more difficult to compact. The highest dry weight of leaves and shoots was observed for seedlings from two extreme compaction variants. In a different study on beech [40], the highest dry weight of leaves and stems was observed in seedlings bred in medium soil compaction variants. This discrepancy in findings may be due to differences in the type of compacted nursery substrate—i.e., the peat-perlite substrate in our research and mineral nursery soil without the organic layer in the cited study. The dry weight of the assimilation apparatus of beech observed in the present study comports with other studies on Q. petraea (Matt.) Liebl. [38]. The highest dry weight of the root system was achieved for the lowest compaction variant (0.08 g·cm−3). In the lower and middle zones (parts (c) and (b), respectively), the dry mass of roots was lower, while the lowest root mass was found in the upper zone (part (a)) of seedlings grown in the medium compaction variants. Numerous studies have indicated that increased substrate compaction results in the shortening of the seedlings’ root systems [8,9,26,28,39]. However, a correlation has been observed between root dry weight and RCD for Picea abies (L.) Karst. [53], F. sylvatica L. [54], Abies alba Mill. [55], and Quercus robur L. [38]. The results of the present study also confirm this relationship because the obtained Pearson’s r coefficient was positive and statistically significant (r = 0.668; p < 0.05).
Grossnickle [56] demonstrated that the resistance of seedlings to post-transplant stress and to drought was greater when the ratio of the weight of the aboveground part to the weight of the underground system, i.e., S/R, was smaller [42]. In the present study, the ratio of dry weight of shoots to the dry weight of the root system changed with the increase in substrate compaction, and the most favorable S/R was obtained in the variants of lower substrate compaction. The remaining high values of S/R (exceeding the permissible limit of ≤2) and SQ could be achieved by providing optimal conditions for the growth of seedlings throughout the production period—i.e., successive and intensive foliar fertilization and lack of moisture stress.
The mean total length of the beech root system determined in the present study was over 3.5 times smaller than the value obtained by Wrzesiński [54] for the same species. This remarkable difference, despite the use of the same trays, could be attributed to the use of mycorrhiza in the cited study. Unlike in the study by Wrzesiński [54], the aboveground part with a relatively small root system dominated in the present research. Increased SH at the cost of root system size is unfavorable in the aspect of production quality. A more extensive root system is typical of a more durable seedling, which will be characterized by a greater growth potential and thus greater survival after planting on the culture [23,42,57,58], which is in turn associated with a lower number of refillings after the loss of seedlings [54]. The highest total root length was observed in seedlings from uncompacted and medium compaction (0.10–0.11 g·cm−3) variants. A reduction in root growth in response to increased soil compaction has been confirmed by Kormanek et al. [38] for Q. petraea Liebl. and by Tworkoski et al. [52] for Q. alba L. and P. nigra Arn. seedlings growing on dusty loam with a bulk density of 1.4 g·cm−3 and on clay sand with a density of 1.6 g·cm−3 [59]. Similarly, reduced root system growth was observed in the study on Eucalyptus nitens (H.Deane & Maiden) Maiden seedlings in response to high soil density, where the total length of main and lateral roots decreased by 71% and 31%, respectively, at elevated compaction level [60]. The formation of a shorter and thicker root due to increased soil compaction was also found in a study on Araucaria angustifolia (Bertol.) Kunze [61].
At the beginning of the experiment, the substrate used was sterile and contained no elements. Due to the small volume of the substrate in each cell tray, no soil fertilizers were used because it could quickly be leached of the substrate. However, the elements appeared in the substrate during cultivation from the foliar fertilizer. Differences in the physical properties (e.g., bulk density) of the substrate may have an impact on the amount of the elements retained in the substrate.
5. Conclusions
This study showed that European beech seedlings growing for a period of 5 months on a peat-perlite substrate with added perlite in BCC Hiko trays (capacity 265 cm3) exhibited low sensitivity to substrate compaction changes. Although the differences observed between the variants in the measured parameters were small, statistically significant differences were found in the height of seedlings, dry weight of roots, and average length of thick and fine roots, as well as SQ and S/R values. The height of seedlings and dry weight of their individual parts showed a slight upward trend with increasing substrate compaction (dry bulk density in the range of 0.078–0.127 g·cm−3). SQ and S/R were improved in seedlings growing on a substrate with lower compaction. Increased substrate compaction (dry bulk density > 0.103 g·cm−3, actual bulk density 0.257 g·cm−3) limited the total root length, particularly in the lower zone of the substrate—i.e., at a cell depth from 5 to 15 cm. The highest values of SH, RCD, leaf surface area, and dry weight of individual parts of seedlings were achieved with the lowest density (dry bulk density 0.078 g·cm−3). The obtained results of the study indicate that a low level of peat-perlite substrate compaction is required for breeding European beech seedlings in nursery trays. The obtained results of the study indicate that a low level of peat-perlite substrate compaction is recommended for breeding European beech seedlings in nursery trays in Hiko V265. Since the highest level of substrate compaction applied in the study did not cause inhibition of seedling growth, additional research using another kind and size of nursery tray is needed to verify whether the substrate compaction at bulk densities above 0.317 g·cm−3 limits the growth responses of European beech seedlings.
Author Contributions
Conceptualization, K.P., S.M. and M.K.; methodology, K.P., S.M. and M.K.; software, K.P. and J.B.; validation, S.M. and M.K.; formal analysis, K.P.; investigation, K.P., S.M. and M.K.; resources, S.M. and M.K.; data curation, S.M. and M.K.; writing—original draft preparation, K.P.; writing—review and editing, K.P., S.M. and M.K.; visualization, K.P. and J.B.; supervision, S.M. and M.K.; project administration, S.M. and M.K.; funding acquisition, S.M. All authors have read and agreed to the published version of the manuscript.
Funding
The research was Funded by the STATE FORESTS NATIONAL FOREST HOLDING. Grant number ER-2717-4/14.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Acknowledgments
We would like to thank the staff of the Rudy Raciborskie Forest District and the management and employees of the Nędza Nursery Farm for allowing the research and assisting us in the performance of this research work, as well as Rafał Czuchta, for providing help in conducting experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jaworski, A. Hodowla Lasu. T. 3. Charakterystyka Hodowlana Drzew i Krzewów Leśnych; Powszechne Wydawnictwo Rolnicze i Leśne: Warszawa, Poland, 2019; ISBN 978-83-09-01117-0. [Google Scholar]
- Tomaś, Ł.; Jagodziński, A.M. Przebudowa Drzewostanów. ACADEMIA-Mag. Pol. Akad. Nauk. 2019, 3–4, 94–97. [Google Scholar]
- Zajączkowski, G.; Jabłoński, M.; Jabłoński, T.; Szmidla, H.; Kowalska, A.; Małachowska, J.; Piwnicki, J. Raport o Stanie Lasów w Polsce 2020 (Report on the State of Forests in Poland 2020); General Directorate of State Forests: Warszawa, Poland, 2021; pp. 1–162.
- Szabla, K.; Pabian, R. Szkółkarstwo Kontenerowe: Nowe Technologie i Techniki w Szkółkarstwie Leśnym; Centrum Informacji Lasów Państwowych: Warszawa, Poland, 2009; ISBN 978-83-89744-80-7.
- Zahreddine, H.G.; Struve, D.K.; Quigley, M. Growing Pinus Nigra Seedlings in SpinoutTM-Treated Containers Reduces Root Malformation and Increases Growth After Transplanting. J. Environ. Hortic. 2004, 22, 176–182. [Google Scholar] [CrossRef]
- Alameda, D.; Villar, R. Moderate Soil Compaction: Implications on Growth and Architecture in Seedlings of 17 Woody Plant Species. Soil Tillage Res. 2009, 103, 325–331. [Google Scholar] [CrossRef]
- Kormanek, M. Determination of the Impact of Soil Compaction on Germination and Seedling Growth Parameters of Common Beech in the Laboratory Conditions. Acta Sci. Pol. Silvarum. Colendarum. Ratio Et Ind. Lignaria 2013, 12, 14–27. [Google Scholar]
- Jourgholami, M.; Khoramizadeh, A.; Zenner, E. Effects of Soil Compaction on Seedling Morphology, Growth, and Architecture of Chestnut-Leaved Oak (Quercus castaneifolia). iForest 2017, 10, 145–153. [Google Scholar] [CrossRef]
- Banach, J.; Kormanek, M.; Jaźwiński, J. Quality of Scots Pine, European Beech and Pedunculate Oak Grown from Sowing on Soil with Different Compaction Levels. Leśne Pr. Badaw. 2020, 81, 167–174. [Google Scholar] [CrossRef]
- Onweremadu, E.U.; Eshett, E.T.; Ofoh, M.C.; Nwufo, M.I.; Obiefuna, J.C. Seedling Performance as Affected by Bulk Density and Soil Moisture on a Typic Tropaquept. J. Plant Sci. 2007, 3, 43–51. [Google Scholar] [CrossRef]
- Rykhlik, A.E.; Bezuglova, O.S. Method of Intra-Soil Pulse Continuous-Discrete Moistening (Model Experiment). Biogeosystem Tech. 2017, 4, 39–65. [Google Scholar] [CrossRef]
- Gudkov, S.V.; Shafeev, G.A.; Glinushkin, A.P.; Shkirin, A.V.; Barmina, E.V.; Rakov, I.I.; Simakin, A.V.; Kislov, A.V.; Astashev, M.E.; Vodeneev, V.A.; et al. Production and Use of Selenium Nanoparticles as Fertilizers. ACS Omega 2020, 5, 17767–17774. [Google Scholar] [CrossRef]
- Marshall, V.G. Impacts of forest harvesting on biological processes in northern forest soils. For. Ecol. Manag. 2000, 133, 43–60. [Google Scholar] [CrossRef]
- Gajda, A.; Przewłoka, B. Soil Biological Activity as Affected by Tillage Intensity. Int. Agrophysics 2012, 26, 15–23. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Soil Compaction and Growth of Woody Plants. Scand. J. For. Res. 1999, 14, 596–619. [Google Scholar] [CrossRef]
- Lipiec, J.; Medvedev, V.V.; Birkas, M.; Dumitru, E.; Lyndina, T.E.; Rousseva, S.; Fulajtár, E. Effect of Soil Compaction on Root Growth and Crop Yield in Central and Eastern Europe. Int. Agrophys. 2003, 17, 61–69. [Google Scholar]
- Passioura, J.B. Soil Conditions and Plant Growth’: Soil Conditions and Plant Growth. Plant Cell Environ. 2002, 25, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Hamza, M.A.; Anderson, W.K. Soil Compaction in Cropping Systems. Soil Tillage Res. 2005, 82, 121–145. [Google Scholar] [CrossRef]
- Towards a Thematic Strategy for Soil Protection; Commission of the European Communities: Brussels, Belgium, 2002; pp. 1–35.
- Arvidsson, J. Influence of Soil Texture and Organic Matter Content on Bulk Density, Air Content, Compression Index and Crop Yield in Field and Laboratory Compression Experiments. Soil Tillage Res. 1998, 49, 159–170. [Google Scholar] [CrossRef]
- Brais, S. Persistence of Soil Compaction and Effects on Seedling Growth in Northwestern Quebec. Soil Sci. Soc. Am. J. 2001, 65, 1263–1271. [Google Scholar] [CrossRef]
- Fleming, R.; Powers, R.; Foster, N.; Kranabetter, J.; Scott, A.; Jr, F.; Berch, S.; Chapman, W.; Kabzems, R.; Ludovici, K.; et al. Effects of Organic Matter Removal, Soil Compaction, and Vegetation Control on 5-Year Seedling Performance: A Regional Comparison of Long-Term Soil Productivity Sites. Can. J. For. Res. 2006, 36, 529–550. [Google Scholar] [CrossRef]
- Picchio, R.; Tavankar, F.; Nikooy, M.; Pignatti, G.; Venanzi, R.; Lo Monaco, A. Morphology, Growth and Architecture Response of Beech (Fagus orientalis Lipsky) and Maple Tree (Acer velutinum Boiss.) Seedlings to Soil Compaction Stress Caused by Mechanized Logging Operations. Forests 2019, 10, 771. [Google Scholar] [CrossRef]
- Zhao, Y.; Krzic, M.; Bulmer, C.E.; Schmidt, M.G.; Simard, S.W. Relative Bulk Density as a Measure of Compaction and Its Influence on Tree Height. Can. J. For. Res. 2010, 40, 1724–1735. [Google Scholar] [CrossRef]
- Kormanek, M.; Banach, J. Influence of unit pressure exerted on soil on quality of renewal of chosen species of trees. Acta Agroph. 2012, 19, 51–63. [Google Scholar]
- Lipiec, J.; Hajnos, M.; Świeboda, R. Estimating Effects of Compaction on Pore Size Distribution of Soil Aggregates by Mercury Porosimeter. Geoderma 2012, 179–180, 20–27. [Google Scholar] [CrossRef]
- Kalinitchenko, V.P.; Glinushkin, A.P.; Swidsinski, A.V.; Minkina, T.M.; Andreev, A.G.; Mandzhieva, S.S.; Sushkova, S.N.; Makarenkov, D.A.; Ilyina, L.P.; Chernenko, V.V.; et al. Thermodynamic Mathematical Model of the Kastanozem Complex and New Principles of Sustainable Semiarid Protective Silviculture Management. Environ. Res. 2021, 194, 110605. [Google Scholar] [CrossRef] [PubMed]
- Alameda, D.; Anten, N.P.R.; Villar, R. Soil Compaction Effects on Growth and Root Traits of Tobacco Depend on Light, Water Regime and Mechanical Stress. Soil Tillage Res. 2012, 120, 121–129. [Google Scholar] [CrossRef]
- Bartholomew, P.W.; Williams, R.D. Effects of Soil Bulk Density and Strength on Seedling Growth of Annual Ryegrass and Tall Fescue in Controlled Environment: Soil Bulk Density Effects on Grass Seedling Growth. Grass Forage Sci. 2010, 65, 348–357. [Google Scholar] [CrossRef]
- Ferree, D.C.; Streeter, J.G.; Yuncong, Y. Response of Container-Grown Apple Trees to Soil Compaction. HortScience 2004, 39, 40–48. [Google Scholar] [CrossRef]
- Pan, E.; Bassuk, N. Effects of Soil Type and Compaction on the Growth of Ailanthus Altissima Seedlings. J. Environ. Hortic. 1985, 3, 158–162. [Google Scholar] [CrossRef]
- Blouin, V.M.; Schmidt, M.G.; Bulmer, C.E.; Krzic, M. Effects of Compaction and Water Content on Lodgepole Pine Seedling Growth. For. Ecol. Manag. 2008, 255, 2444–2452. [Google Scholar] [CrossRef]
- Conlin, T.S.S.; van den Driessche, R. Soil Compaction Studies; FRDA Report; Canadian Forest Service and B.C. Ministry of Forests: Ottawa, ON, Canada, 1996; pp. 1–14.
- Siegel-Issem, C.M.; Burger, J.A.; Powers, R.F.; Ponder, F.; Patterson, S.C. Seedling Root Growth as a Function of Soil Density and Water Content. Soil Sci. Soc. Am. J. 2005, 69, 215–226. [Google Scholar] [CrossRef]
- Maupin, C.; Struve, D.K. Red Oak Transplanted to Different Bulk Density Soils Have Similar Water Use Characteristics. J. Arboric. 1997, 23, 233–238. [Google Scholar] [CrossRef]
- Jordan, D.; Ponder, F.; Hubbard, V.C. Effects of Soil Compaction, Forest Leaf Litter and Nitrogen Fertilizer on Two Oak Species and Microbial Activity. Appl. Soil Ecol. 2003, 23, 33–41. [Google Scholar] [CrossRef]
- Bejarano, M.D.; Villar, R.; Murillo, A.M.; Quero, J.L. Effects of Soil Compaction and Light on Growth of Quercus Pyrenaica Willd. (Fagaceae) Seedlings. Soil Tillage Res. 2010, 110, 108–114. [Google Scholar] [CrossRef]
- Kormanek, M.; Głąb, T.; Banach, J.; Szewczyk, G. Effects of Soil Bulk Density on Sessile Oak Quercus Petraea Liebl. Seedlings. Eur. J. For. Res 2015, 134, 969–979. [Google Scholar] [CrossRef]
- Kormanek, M.; Banach, J.; Ryba, M. Influence of Substrate Compaction in Nursery Containers on the Growth of Scots Pine (Pinus sylvestris L.) Seedlings. For. Res. Pap. 2013, 74, 307–314. [Google Scholar] [CrossRef]
- Kormanek, M.; Banach, J.; Leńczuk, D. Determination of the Impact of Soil Compaction on Growth Performance and Quality of Seedlings of European Beech Fagus sylvatica L. Grown in the Laboratory Conditions; SAGE Publishing: Thousand Oaks, CA, USA, 2013; pp. 67–78. [Google Scholar]
- Puértolas, J. Effects of Nutritional Status and Seedling Size on Field Performance of Pinus Halepensis Planted on Former Arable Land in the Mediterranean Basin. Forestry 2003, 76, 159–168. [Google Scholar] [CrossRef]
- Haase, D. Morphological and Physiological Evaluations of Seedling Quality. In National Proceedings: Forest and Conservation Nursery Associations—2006; Proc. RMRS-P-50; USDA Forest Service: Fort Collins, CO, USA, 2007; pp. 3–8. [Google Scholar]
- Grossnickle, S.C. Why Seedlings Survive: Influence of Plant Attributes. New For. 2012, 43, 711–738. [Google Scholar] [CrossRef]
- Ivetić, V.; Skorić, M. The Impact of Seeds Provenance and Nursery Production Method on Austrian Pine (Pinus Nigra Arn.) Seedlings Quality. Ann. For. Res. 2013, 3, 297–305. [Google Scholar]
- Johnson, J.D.; Cline, M.L. Seedling Quality of Southern Pines. In Forest Regeneration Manual; Duryea, M.L., Dougherty, P.M., Eds.; Forestry Sciences; Springer: Dordrecht, The Netherlands, 1991; Volume 36, pp. 143–159. ISBN 978-0-7923-0960-4. [Google Scholar]
- Roller, K.J. Suggested Minimum Standards for Containerized Seedlings in Nova Scotia; Information Report M-X-69; Maritimes Forest Research Centre: Fredericton, NB, Canada, 1977; pp. 1–18. [Google Scholar]
- PN-R-67025 Sadzonki Drzew i Krzewów do Upraw Leśnych i na Plantacje; Polski Komitet Normalizacyjny: Warszawa, Poland, 1999; ISBN 978-83-236-2771-5.
- Skrzyszewska, K.; Banach, J.; Bownik, G. Wpływ sposobu przedsiewnego przygotowania żołędzi i terminu siewu na kiełkowanie nasion i wzrost sadzonek dębu szypułkowego. Sylwan 2019, 163, 716–725. [Google Scholar] [CrossRef]
- Makita, N.; Hirano, Y.; Mizoguchi, T.; Kominami, Y.; Dannoura, M.; Ishii, H.; Finér, L.; Kanazawa, Y. Very Fine Roots Respond to Soil Depth: Biomass Allocation, Morphology, and Physiology in a Broad-Leaved Temperate Forest. Ecol. Res. 2011, 26, 95–104. [Google Scholar] [CrossRef]
- Farahnak, M.; Mitsuyasu, K.; Hishi, T.; Katayama, A.; Chiwa, M.; Jeong, S.; Otsuki, K.; Sadeghi, S.M.M.; Kume, A. Relationship between Very Fine Root Distribution and Soil Water Content in Pre- and Post-Harvest Areas of Two Coniferous Tree Species. Forests 2020, 11, 1227. [Google Scholar] [CrossRef]
- TIBCO Software Inc. Statistica (Data Analysis Software System); TIBCO Software Inc.: Sydney, NSW, Australia, 2017. [Google Scholar]
- Tworkoski, T.; Burger, J.; Wm, D. Soil Texture and Bulk Density Affect Early Growth of White Oak Seedlings. Tree Plant. Note 1983, 34, 22–25. [Google Scholar]
- Grossnickle, S.C. Ecophysiology of Northern Spruce Species: The Performance of Planted Seedlings; NRC Research Press: Ottawa, ON, Canada, 2000; ISBN 978-0-660-17959-9. [Google Scholar]
- Wrzesiński, P. The Influence of Seedling Density in Containers on Morphological Characteristics of European Beech. For. Res. Pap. 2015, 76, 304–310. [Google Scholar] [CrossRef][Green Version]
- Kormanek, M.; Banach, J.; Leńczuk, D. Influence of Soil Compaction on the Growth of Silver Fir (Abies alba Mill.) under a Forest Canopy. EQ 2015, 22, 47–54. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Importance of Root Growth in Overcoming Planting Stress. New For. 2005, 30, 273–294. [Google Scholar] [CrossRef]
- Ivetić, V.; Grossnickle, S.; Škorić, M. Forecasting the Field Performance of Austrian Pine Seedlings Using Morphological Attributes. Iforest-Biogeosci. For. 2016, 10, 99–107. [Google Scholar] [CrossRef]
- Rodríguez-Ortiz, J.C.; Carballo-Méndez, F.d.J.; Preciado-Rangel, P.; Rodríguez-Fuentes, H.; Lozano-Cavazos, C.J. Broccoli Seedling Production in Response to Recognised Organic Inputs. Int. J. Agric. Biol. 2021, 26, 8. [Google Scholar]
- Zisa, R.R.; Halverson, H.G.; Stout, B.B. Establishment and Early Growth of Conifers on Compact Soils in Urban Areas; Forest Service Research Paper; Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Amherst, MA, USA, 1980; Volume NE-451, pp. 1–8. [Google Scholar]
- Misra, R.K.; Gibbons, A.K. Growth and Morphology of Eucalypt Seedling-Roots, in Relation to Soil Strength Arising from Compaction. Plant Soil 1996, 182, 1–11. [Google Scholar] [CrossRef]
- Mósena, M.; Dillenburg, L.R. Early Growth of Brazilian Pine (Araucaria Angustifolia [Bertol.] Kuntze) in Response to Soil Compaction and Drought. Plant Soil 2004, 258, 293–306. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).