Abstract
Cadmium is an essential element for plant growth and development. Its accumulation in soil is more hazardous to human and animal health than to plants and microorganisms. A pot greenhouse experiment was conducted to determine the usability of Sinapis alba L. and Avena sativa L. for the phytoremediation of soil contaminated with cadmium and to verify cellulose viability in the remediation of soil under cadmium pressure in doses from 4 to 16 mg Cd2+ kg−1 soil d.m. (dry matter) The effect of cadmium on soil microbiome was investigated with the culture method and the variable region sequencing method. Sinapis alba L. and Avena sativa L. were found viable in the phytoremediation of soil contaminated with Cd2+. Avena sativa L. was more potent to accumulate Cd2+ in roots than Sinapis alba L. Although the fertilization of Cd2+- contaminated soil with cellulose stimulated the proliferation of microorganisms, it failed to mitigate the adverse effects of Cd2+ on bacterial diversity. Bacteria from the Sphingomonas, Sphingobium, Achromobacter, and Pseudomonas genera represented the core microbiome of the soils sown with two plant species, contaminated with Cd2+ and fertilized with cellulose. Stimulation of the growth and development of these bacteria may boost the efficacy of phytoremediation of cadmium-contaminated soils with Sinapis alba L. and Avena sativa L.
1. Introduction
Cadmium is a heavy metal posing severe threat to human and animal health. Because of its high toxicity, the risks stemming from its adverse impacts are considered globally [1,2,3,4]. Its levels recorded in uncontaminated soils range from 0.01 to 1 mg kg−1, with a global average of 0.36 mg kg−1 [5], and are observed to increase successively due to geological weathering of rocks [6] and anthropogenic activity, such as mining, metallurgy, fuel combustion, and soil amendment with phosphorus-based fertilizers [3,7,8]. The increasing environmental contamination with this element contributes to its accumulation in all links of the food chain [6,9,10].
Cadmium is toxic to both prokaryotic and eukaryotic organisms. Its toxicity is due to the binding of its free ions with atoms of sulfur, oxygen, and hydrogen present in tissues, which modifies the morphology of plant and animal cells. It may also disturb the circulation of such elements as: Fe, Cu, Mg, Ca, Zn, and Se [11]. The toxic effects of cadmium on microorganisms entail chiefly blocking the active centers of enzymes, reactions with -SH groups, competition with basic metabolites, and cell membrane damage [12]. In turn, its toxic effects on plants are triggered by disorders in the uptake of micro- and macroelements indispensable for proper body function. In plants, cadmium is bound by complexing compounds (metallothioneins and phytochelatins) and accumulated in vacuoles, cell membrane, cell compartments, Golgi apparatus, and endoplasmic reticulum [13].
Cadmium is very quickly transported throughout the trophic chain: soil—plant—man [14,15]. Its ions aggravate oxidative stress in plant cells because it is very easily absorbed by roots and transported to the aerial parts of plants, inhibiting their growth and potentially leading to their death [11,12,13,16]. The strongly adverse effects of cadmium on higher organisms are corroborated by the fact that it is included in the group 1 carcinogenic substances by the International Agency for Research on Cancer (IARC) [17].
Cadmium occurs in the environment mainly in the second oxidation state [18,19]. It is a white metal with bluish tint. Cadmium compounds feature various solubility in water, ranging from very good solubility (e.g., acetate, chloride, sulfate) to almost insolubility (e.g., oxide, sulfide). Similar to other heavy metals, considering its binding force, cadmium extends to seven geochemical fractions varying in mobility and, thereby, in bioavailability [18,20]. These fractions, beginning from the most available one to organisms, may be ordered as follows: ionic—occurring in soil water; ionic—adsorbed by the sorptive complex; carbonate; the one bound with manganese oxides; the one bound with iron oxides; organic; and the nondigestible one occurring in primary metals [21,22]. The amount of cadmium available to plants is determined by mobilization and immobilization processes, which are in turn dependent on the physicochemical properties of soil (pH, cation exchange capacity, fraction size distribution, and organic matter content and type), activities of soil microorganisms, and species of grown plants [19,20,23].
Root secretions may provide a source of nutrients to microorganisms which are more abundant and active in this rhizospheric zone of the soil than in the rootless one [24]. Soil microorganisms mineralize nutrients dissolved in the soil solution [25] and secrete their own metabolites (e.g., organic acids, siderophores, and biosurfactants), forming complexes with heavy metals which differ in stability and mobility [23,26,27]. They affect soil properties, such as sorption and desorption processes, which in turn determine the heavy metal accumulation in plant tissues. They may form a protective barrier preventing penetration of harmful substances to plant roots [28], and bolster physiological functions of plants by providing growth hormones and facilitating the uptake of iron, phosphorus, and potassium compounds [29].
Contaminated environments are most frequently recultivated via phytoextraction and phytostabilization. Hence, the choice of appropriate recultivation plant species for specified pollutants is of utmost importance [30,31]. Preparatory measures, such as liming, organic and mineral fertilization, and application of various sorbents, are strongly recommended before the exact recultivation [19,32,33,34].
In order to aid the phytostabilization of cadmium-contaminated soil, cellulose has been used in this study, as it is the most abundant organic compound in nature [35]. Cellulose is the major constituent of plant cell walls, a nonbranched polysaccharide containing 3000 to 4000 of D-glucose residues linked with β-1,4 glycosidic bonds [36]. It undergoes two-stage microbiological hydrolysis in the soil, firstly to cellobiose and then to glucose [37]. The rate of cellulose degradation is largely dependent on the activity of soil microorganisms for which it serves as a nutrient [38]. Its application allows capturing the microbiological variability of soil under strictly controlled conditions. After modification, cellulose can also serve as a good sorbent [39].
Considering the above, an experiment was established which combined remediation techniques applicable to soils contaminated with cadmium. The following research hypotheses were formulated: (1) crystalline cellulose stimulates the proliferation of autochthonous soil microorganisms, and (2) changes in the abundance and diversity of bacteria may increase Sinapis alba L. and Avena sativa L. tolerance to cadmium effects. Novelty of the study lies in the cultivation of two crops directly one after another, which is especially important in the case of soils contaminated with cadmium as its effects on the microbiome and the crop are long-standing. In addition, Sinapis alba L.—as the main crop—“cleans” the soil environment of cadmium by accumulating it in its roots and aerial parts [40], whereas Avena sativa L.—as the successive crop—serves as a phytosanitary plant that secrets an alkaloid, scopoletin, which prevents the development of fungi-inducing, take-all diseases of cereals [41].
Therefore, this study focuses on alternative techniques of remediation, including phytoremediation and phytostabilization of cadmium in soil, both deemed highly prospective owing to their simplicity, high efficacy, in situ applicability, and cost-effectiveness.
2. Materials and Methods
2.1. Soil
A pot experiment was conducted with soil having pHKCl 7.0, classified as sandy loam (<0.002 mm fraction—2.88%, 0.002–0.050 mm fraction—27.71%, and 0.050–2.000 mm fraction—69.41%). The soil material was collected from an area located in northeastern Poland, Tomaszkowo village, in the Mazurian Lake District (53.7161° N, 20.4167° E). In-depth characteristics of the sandy loam used in the experiment were provided in a work by Boros-Lajszner et al. [32].
2.2. Experimental Scheme
The study was carried out with the following independent variables: (1) extent of soil contamination with cadmium and (2) addition of cellulose as a substance mitigating adverse effects of cadmium. The soil was contaminated with cadmium in the form of an aqueous solution of cadmium chloride (CdCl2), in doses of 0, 4, 8, and 16 mg Cd2+ kg−1 soil d.m. Microcrystalline cellulose with a particle size of 90 μm (ACROS ORGANICS, Geel, Belgium, extra pure) was used in a dose of 15 g kg−1 soil d.m. As background, soil from all pots was fertilized with urea, monopotassium phosphate, potassium chloride, and magnesium sulfate in the following doses: N—160 mg, P—21 mg, K—73 mg, Mg—15 mg kg−1 soil d.m. Following study design, all components (cadmium chloride, cellulose, and fertilizers) were thoroughly mixed with soil (in the amount of 3.5 kg) and then the soil was transferred to pots. Analyses were carried out in 4 replications. White mustard (Sinapis alba L.) of Rota cultivar served as an experimental plant (8 plants per pot). Its aerial parts and roots were harvested after 40 days of growth. Next, a successive plant was sown, namely oat (Avena sativa L.) of Bingo cultivar (12 plants per pot). Oat crop was harvested after another 40 days. During white mustard and oat growth, the soil humidity was maintained at 50% of the maximum water-holding capacity. After the harvest of both crops, their aerial parts and roots were analyzed for Cd2+ content. Sinapis alba L. belongs to the family Brassicaceae. It is an annual plant characterized by a very high growth rate. It has been ranked third among the oil-producing crops in terms of acreage area in Poland. An additional advantage of Sinapis alba L. in its use as a phytoremediating plant is its high tolerance to soil contamination with cadmium [42,43]. Avena sativa L. belongs to the family Poaceae. It was selected as a successive crop because Avena sativa L. is a phytosanitary plant. This cereal species exerts two types of effects on soil: it significantly reduces weed infestation and prevents the development of take-all diseases in successive crops.
2.3. Soil and Plant Analysis
Soil graining was determined by the aerometric method [44,45], and soil pH was analyzed using the potentiometric method in an aqueous KCl solution with 1 mol dm−3 soil concentration [46]. Cadmium content was determined in the aerial parts and roots of Sinapis alba L. and Avena sativa L. by atomic absorption spectrometry following microwave mineralization, according to the Polish standard PN-EN 14084:2004(N) [47].
2.4. Indicators Valorizing the Usability of Plants for the Remediation of Cadmium-Contaminated Soil
The harvested crops of white mustard (Sinapis alba L.) and oats (Avena sativa L.) allowed for determining their resistance to cadmium based on the formula described by Orwin and Wardle [48]:
where:
RS—plant resistance to heavy metal;
C0—biomass yield in control soil (nonpolluted);
D0—difference between biomass yield produced on nonpolluted soil and soil polluted with heavy metal.
In addition, cadmium uptake by plants (D) was computed using the following formula:
and its redistribution in plants according to the following formula:
where:
YA—aboveground parts yield (g kg−1);
YR—root yield (g kg−1);
CA—heavy metal content in aboveground parts (µg g−1);
CR—heavy metal content in roots (µg g−1);
RA—redistribution of heavy metals in aboveground parts (%);
RR—redistribution of heavy metals in roots (%).
2.5. Microbiological Analyses
Microbiological analyses were carried out following the procedures described by Wyszkowska et al. [49]. The growth rate of individual groups of microorganisms and the counted number of grown colonies were used to compute the colony development index (CD) and ecophysiological diversity index (EP) of microorganisms in the cadmium-contaminated soil. The formulas have been described in the manuscript by De Leij et al. [42], Sarathchandra et al. [43], and Wyszkowska et al. [49].
In turn, the population counts of soil microorganism were used to calculate the index of their resistance (RS) to Cd2+ contamination according to the formula described by Orwin and Wardle [48] and the index of the impact of cadmium (IFCd) and cellulose (IFCe) on their abundance. Detailed description of particular indices was provided in the work by Boros-Lajszner et al. [50]. In addition, the control soil and soil contaminated with a cadmium dose of 16 mg Cd2+ kg−1 were analyzed for bacterial diversity with the variable region sequencing method. DNA was extracted using the Bead-Beat Micro Gravity kit (A & A Biotechnology, Gdansk, Poland). PCR was performed using Q5 Hotstat High- Fidelity DNA Polymerase (NEBNext, Ipswich, MA, USA). The metagenomic analysis, automatic data analysis, and preparation of the reference sequence database are presented in the paper by Borowik et al. [51].
2.6. Statistical Analyses
Data were processed statistically with the analysis of variance ANOVA at a significance level of p ≤ 0.05 using STATISTICA 13.1 software (StatSoft, Tulsa, OK, USA) [52]. The analyzed data had normal distribution and similar variance. Homogenous groups were computed using the post hoc Tukey test (HSD—Tukey’s honestly significant difference test).
Metagenomic data ≥1% were visualized with 4 types of computer software: bacteria phyla were visualized using STAMP 2.1.3. (St Lucia, Queensland, Australia) with two-way G-test for statistical hypotheses (w/Yates) + Fisher test [53]; bacterial classes, orders, and families—using an internet tool, Circos 0.68 [54]; bacterial genera—using Rstudio v1.2.5033 (JJ Allaire, Boston, MA, USA) with R project [55,56], gplots v3.6.2 library [57], and an internet tool for data set analysis InteractiveVenn (Irvine, CA, USA) [58]. In addition, bacteria biodiversity was determined based on the Shannon–Wiener index (H’) and Simpson index (D) [59,60].
3. Results
3.1. Effect of Cadmium and Cellulose on Plants
Sinapis alba L. (Figure 1) and Avena sativa L. (Figure 2) proved to be relatively resistant to soil contamination with cadmium doses from 4 to 16 mg Cd2+ kg−1 soil d.m. (Table 1). Figure 1 presents Sinapis alba L. at the BBCH 65 stage, i.e., at the full blossom stage when 50% of flowers are opened on the main inflorescence and older petals fall down. In turn, Figure 2 depicts Avena sativa L. at the BBCH 51 stage, i.e., at the panicle ejection stage.

Figure 1.
The effect of cadmium on the growth of Sinapis alba L. (a,e)—0 mg Cd2+; (b,f)—4 mg Cd2+; (c,g)—8 mg Cd2+; (d,h)—16 mg Cd2+.

Figure 2.
The effect of cadmium on the growth of Avena sativa L. (a,e)—0 mg Cd2+; (b,f)—4 mg Cd2+; (c,g)—8 mg Cd2+; (d,h)—16 mg Cd2+.

Table 1.
The index of resistance (RS) of Sinapis alba L. and Avena sativa L. to soil contamination with cadmium.
The plants’ response to the heavy metal tested varied depending on cellulose application to the soil. In the series without polysaccharide application, Avena sativa L. (mean RS of aerial parts = 0.786) was more resistant to cadmium presence in the soil environment than Sinapis alba L. (mean RS of aerial parts = 0.691). In the series with cellulose, the plants’ response was opposite, i.e., the mean RS of the aerial parts reached 0.902 for Sinapis alba L. and 0.724 for Avena sativa L. The response of roots for both tested plants to the tested doses of the heavy metal was stronger. The lower values of the resistance index (RS) of Sinapis alba L. and Avena sativa L. roots compared to the aerial parts are indicative of more destructive effect of the heavy metal on their roots.
Sinapis alba L. featured a significantly higher cadmium uptake from the soil contaminated with its dose of 16 mg Cd2+ kg−1 than Avena sativa L. (Table 2). Fertilization with cellulose caused a 54% decrease in cadmium uptake by Sinapis alba L. and a 28% decrease in its uptake by Avena sativa L. Values of redistribution indicators (RA and RR) prove that Sinapis alba L. accumulated 2.4-fold to 6.7-fold more cadmium in its roots than aerial parts, whereas Avena sativa L. accumulated even more cadmium in respective morphological parts, showing from 8.9-fold to 30.2-fold higher accumulation in roots than aerial parts.
Table 2.
Cadmium uptake (D) (μg kg−1 d.m.) by Sinapis alba L. and Avena sativa L. and its redistribution (%) in the aerial parts (RA) and roots (RR) of plants.
3.2. Effect of Cadmium and Cellulose on Soil Microorganisms
Cadmium significantly affected the population numbers of the analyzed microorganisms (Table 3). The highest counts of microorganisms were determined in the soil samples not contaminated with cadmium. Generally, higher counts of microorganisms were found in the control and contaminated soil sown with the successive crop, i.e., Avena sativa L., than in the soil with the main crop, i.e., Sinapis alba L. Soil contamination with the tested metal contributed to a significant decrease in the abundance of all soil microorganisms under study, as evidenced by the negative coefficient of correlation between the extent of soil contamination with cadmium and population numbers of microorganisms, ranging from −0.741 (actinobacteria in the soil sown with Avena sativa L.) to −0.999 (organotrophic bacteria in the soil sown with Avena sativa L.).
Table 3.
Microbial counts in the soil sown with Sinapis alba L. and Avena sativa L. contaminated with cadmium.
The adverse effect of cadmium on the communities of oligotrophic bacteria, actinobacteria, and fungi was also indicated by negative values of IFCd indices (Figure 3), which were observed to decrease along with an increasing Cd doses applied to the soil. In the soil sown with Sinapis alba L., the values of IFCd indices ranged from −0.262 (4 mg Cd2+ kg−1 soil d.m.) to −0.575 (16 mg Cd2+ kg−1 soil d.m.) for organotrophic bacteria, from −0.084 to −0.458 for actinobacteria, and from −0.048 to −0.624 for fungi, respectively. In the soil sown with Avena sativa L., the respective values ranged from −0.115 to −0.513 for organotrophic bacteria, from –0.614 to –0.678 for actinobacteria, and from −0.194 to −0.565 for fungi. The substantially higher IFCd values noted for organotrophs and fungi in the soil sown with Avena sativa L. prove that the impact of cadmium on their communities was weaker in the soil sown with the successive crop. In turn, the impact of cadmium on actinobacteria proliferation not only did not diminish with time of the experiment but increased, as indicated by the lower IFCd values determined in the soil sown with the successive plant. Population numbers of microorganisms were significantly determined not only by the extent of soil contamination and crop species but also by soil fertilization with cellulose (Figure 4). The polysaccharide tested significantly stimulated the proliferation of organotrophic bacteria, actinobacteria, and fungi in the noncontaminated soil and the soil contaminated with cadmium. The only exceptions were actinobacteria in the noncontaminated soil sown with Avena sativa L. and fungi in the soil contaminated with a cadmium dose of 4 mg Cd2+ kg−1 soil d.m. and sown with Avena sativa L. Fertilization with cellulose caused greater, positive changes in the population numbers of all groups of microorganisms in the soil sown with the main rather than successive crop. In the soil sown with Sinapis alba L., the mean value of the IFCe index reached 0.647 for organotrophic bacteria, 2.409 for actinobacteria, and 1.528 for fungi, whereas the respective values determined in the soil sown with Avena sativa L. were 0.383, 0.182, and 0.472.
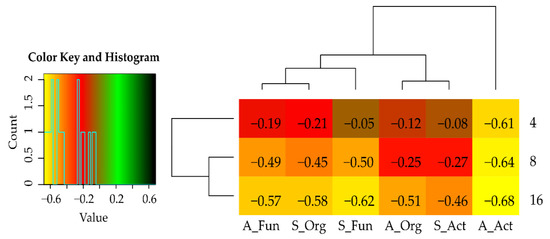
Figure 3.
Index of cadmium effect (IFCd) on microorganisms in the soil sown with Sinapis alba L. and Avena sativa L. S—Sinapis alba L.; A—Avena sativa L.; Org—organotrophic bacteria; Act—actinobacteria; Fun—fungi.
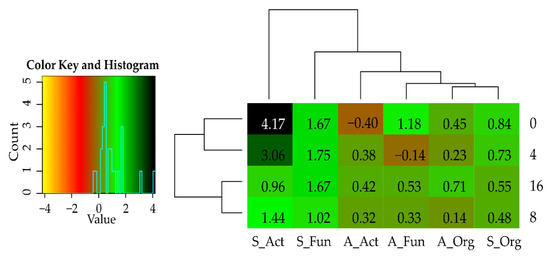
Figure 4.
Index of cellulose effect (IFCe) on microorganisms in the soil sown with Sinapis alba L. and Avena sativa L. Refer to the key in Figure 1.
The effects of the independent variables on the communities of organotrophic bacteria, actinobacteria, and fungi were very well mirrored by the index of their resistance (RS) to soil contamination with cadmium (Table 4). The analysis of RS index values allowed concluding that the organotrophic bacteria and fungi were more resistant to cadmium effects in the soil not fertilized with cellulose, regardless of crop species, whereas actinobacteria behaved the same way only in the soil sown with Sinapis alba L.
Table 4.
The index of resistance (RS) of microorganisms to soil contamination with cadmium and sown with Sinapis alba L. and Avena sativa L.
Soil contamination with cadmium triggered changes in soil microbiome diversity, as evidenced by the values of the colony development index (CD) and the physiological diversity index (EP) (Table 5 and Table 6). In the soil not fertilized with cellulose, the CD value computed for organotrophic bacteria was higher than the respective values determined for actinobacteria and fungi (Table 5). Regardless of Cd-contaminating dose, the CD values determined in the soil sown with Sinapis alba L. reached 49.634 for organotrophic bacteria, 21.520 for actinobacteria, and 30.613 for fungi, whereas the respective values noted in the soil sown with Avena sativa L. were 38.855, 21.232, and 35.611, respectively. Soil fertilization with cellulose exerted an inexplicit effect on CD values of the analyzed bacterial communities, as it decreased the CD value of organotrophic bacteria, increased that in fungi, and did not modify that in actinobacteria.
Table 5.
Colony development index (CD) of microorganisms in the soil contaminated with cadmium and sown with Sinapis alba L. and Avena sativa L.
Table 6.
Ecophysiological diversity index (EP) of microorganisms in the soil contaminated with cadmium and sown with Sinapis alba L. and Avena sativa L.
Regardless of crop species grown in pots, the cadmium contamination of soil not fertilized with cellulose diminished the ecophysiological diversity of organotrophic bacteria and actinobacteria, while increasing that in fungi (Table 6). The effect of cadmium on EP values of the analyzed microorganisms was more diverse in the polysaccharide-enriched soil. Generally, cadmium decreased the EP value of organotrophic bacteria in the soil sown with Sinapis alba L., and increased it in the soil sown with Avena sativa L. It also increased EP values computed for actinobacteria and fungi in soils sown with both crops tested. The analysis of data presented in Table 6 indicates that cellulose increased the ecophysiological diversity only in the case of organotrophic bacteria.
All independent variables (soil contamination with cadmium, soil fertilization with cellulose, and crop species) caused changes in the structure of the genetic diversity of bacteria at all taxonomic levels (Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12, Figure 13 and Figure 14). Of all identified phyla, the highest OTU was determined for Proteobacteria and Actinobacteria (Figure 5, Figure 6, Figure 7 and Figure 8). Proteobacteria turned out to be the prevailing phylum in the soil sown with Sinapis alba L., both in the noncontaminated and contaminated one as well as without and with cellulose addition (Figure 5).
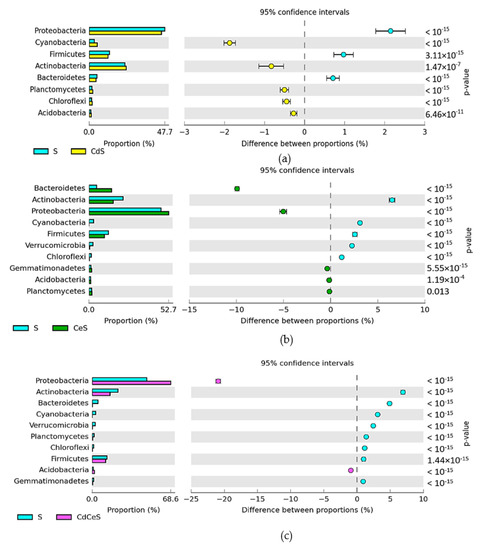
Figure 5.
A comparative analysis of the effects of soil contamination with cadmium (a), soil fertilization with cellulose (b), and soil contamination with cadmium + soil fertilization with cellulose (c) on changes in bacterial phyla in the soil sown with Sinapsis alba L. (S). Differences between ratios of bacterial phyla, OTU ≥ 1%. Cd—ion Cd2+; Ce—cellulose.
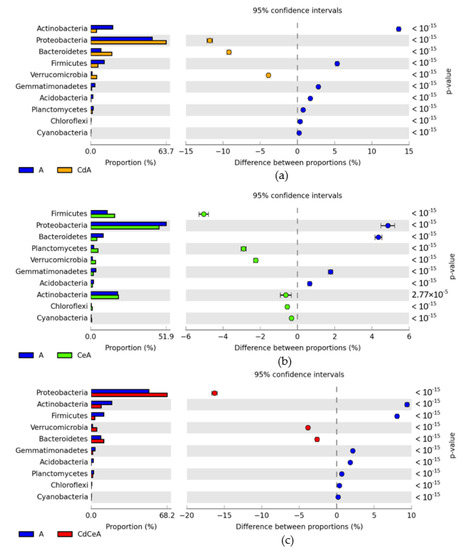
Figure 6.
A comparative analysis of the effects of soil contamination with cadmium (a), soil fertilization with cellulose (b), and soil contamination with cadmium + soil fertilization with cellulose (c) on changes in bacterial phyla in the soil sown with Avena sativa L. (A). Differences between ratios of bacterial phyla, OTU ≥ 1%. Cd—ion Cd2+; Ce—cellulose.
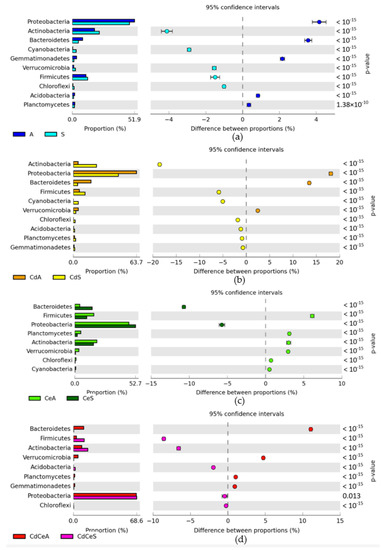
Figure 7.
A comparative analysis of the effects of crop species (a), soil contamination with cadmium (b), soil fertilization with cellulose (c), and soil contamination with cadmium + soil fertilization with cellulose (d) on changes in bacterial phyla in the soil. Cd—ion Cd2+; Ce—cellulose; S—Sinapis alba L.; A—Avena sativa L.
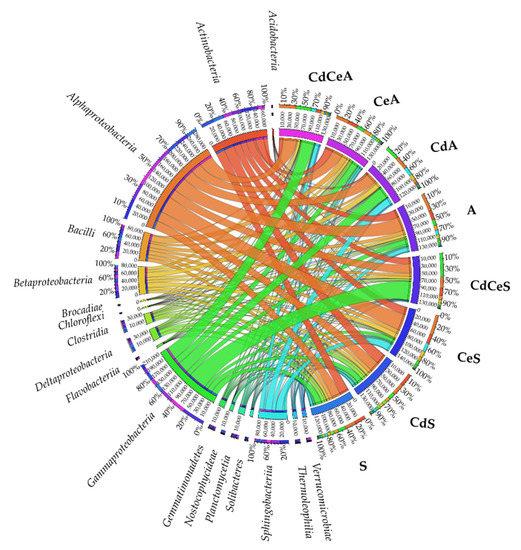
Figure 8.
Relative abundance of dominating bacterial classes in the samples of soil sown with Sinapis alba L. (S) and Avena sativa L. (A). Cd—ion Cd2+; Ce—cellulose.
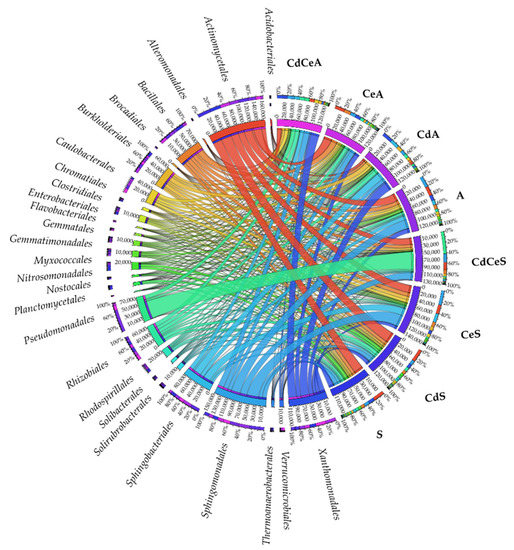
Figure 9.
Relative abundance of dominating bacterial orders in the samples of soil sown with Sinapis alba L. (S) and Avena sativa L. (A). Cd—ion Cd2+; Ce—cellulose.
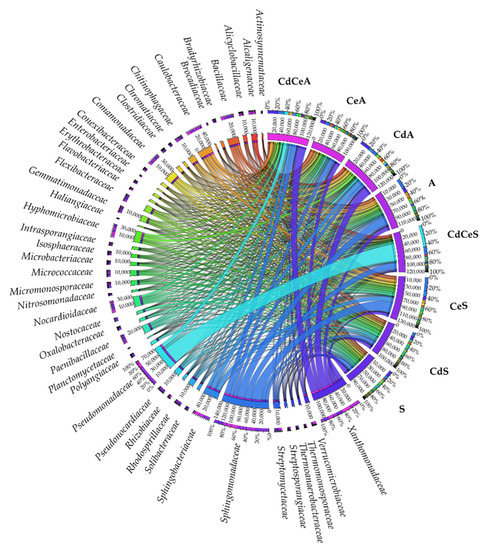
Figure 10.
Relative abundance of dominating bacterial families in the samples of soil sown with Sinapis alba L. (S) and Avena sativa L. (A). Cd—ion Cd2+; Ce—cellulose.
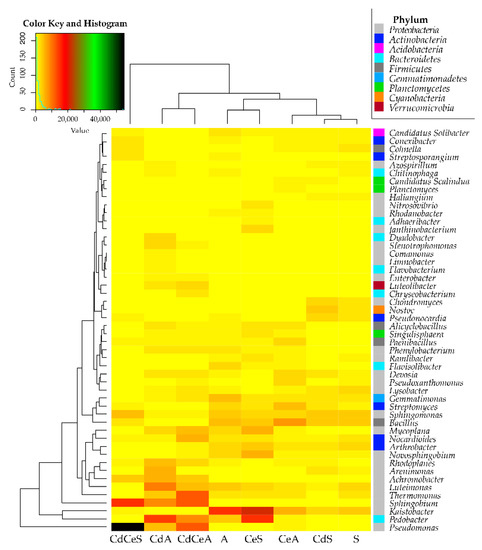
Figure 11.
Heat map showing relationships between bacterial genera in the samples of soil sown with Sinapis alba L. (S) and Avena sativa L. (A). Cd—ion Cd2+; Ce—cellulose.
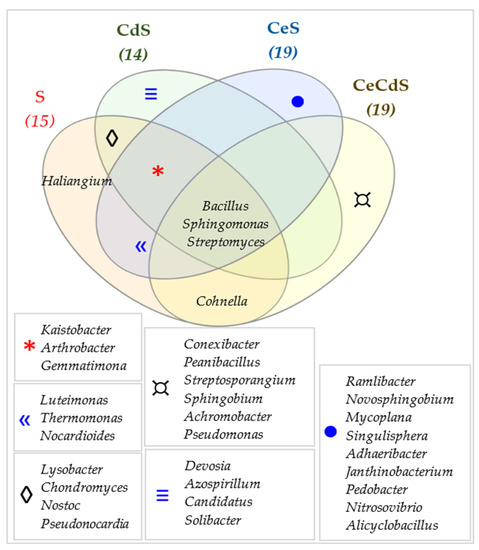
Figure 12.
Venn diagram presenting unique and common bacterial genera in the samples of soil sown with Sinapis alba L. (S). Cd—ion Cd2+; Ce—cellulose.
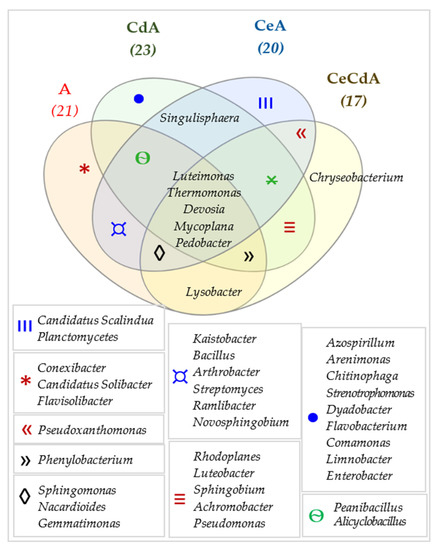
Figure 13.
Venn diagram presenting unique and common bacterial genera in the samples of soil sown with Avena sativa L. (A). Cd—ion Cd2+; Ce—cellulose.
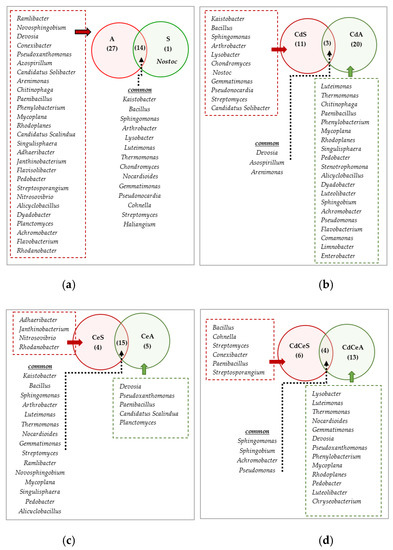
Figure 14.
A comparative analysis of the effects of crop species (a), soil contamination with cadmium (b), soil fertilization with cellulose (c), and soil contamination with cadmium + soil fertilization with cellulose (d) on changes in the unique and common bacterial genera in the soil. Cd—ion Cd2+; Ce—cellulose; S—Sinapis alba L.; A—Avena sativa L.
In the control soil not fertilized with cellulose and sown with Sinapis alba L. (S), the Proteobacteria accounted for 47.71%, whereas in the cadmium-contaminated soil (CdS) accounted for 45.56%. Cellulose application promoted Proteobacteria proliferation, boosting their contribution in the soil microbiome to 52.74% in CeS soil and to 68.61% in CdCeS soil. Contributions of the second ranked phylum, i.e., Actinobacteria, in the soil microbiome were as follows: S—22.63%, CdS—23.46%, CdCeS—16.11%, and CdCeS—15.70%. In the nonfertilized and noncontaminated soil sown with Avena sativa L. (A), Proteobacteria constituted 51.87%; in the cadmium-contaminated soil (CdA), they accounted for 63.65%; in the soil fertilized with cellulose (CeA), 47.01%; whereas, in the fertilized and contaminated soil (CdCeA) they accounted for 68.17% of all bacteria (Figure 4). Actinobacteria accounted for, respectively: 18.51% (A), 4.90% (CdA), 19.14% (CeA), and 9.11% (CdCeA). Soil contamination with cadmium triggered changes in the bacterial structure. It increased the OTU of Proteobacteria by 11.78% in the soil not fertilized with cellulose and by 16.30% in the fertilized soil.
Polysaccharide application to the soil sown with Avena sativa L. increased the difference in Proteobacteria proportion by 5%. Soil contamination with a cadmium dose of 16 mg Cd2+ kg−1 soil d.m. in the pots sown with Avena sativa L. (CdA) contributed to decreased abundance of Actinobacteria by 13.61% in the series without cellulose and by 9.4% (CdCeA) in the series with cellulose application (Figure 6). Other relatively abundant taxa identified in the noncontaminated and contaminated soil included Firmicutes and Bacteroidetes. A comparative analysis of the relative abundance of bacterial phyla in the samples of soil sown with Sinapis alba L. (S) and Avena sativa L. (A) allowed concluding that Avena sativa L. cultivation significantly reduced the OTU number of Proteobacteria (by 4.2%), while increasing that of Actinobacteria (by 4.2%) (Figure 7a). Exposure of the soil sown with Avena sativa L. to cadmium pressure decreased the relative abundance of Proteobacteria (by 18.1%) and Bacteroidetes (by 13.5%), and increased that in Actinobacteria (by 18.6%) (Figure 7b). Soil fertilization with cellulose (Figure 7c) promoted the proliferation of Bacteroidetes (OTU increase by 10.7%) and Proteobacteria (OTU increase by 5.7%), but suppressed that of Actinobacteria (by 3.0%). Cellulose application to the cadmium-contaminated soil diminished the relative abundance of Bacteroidetes (by 11.1%) and increased that in Firmicutes (by 8.6%) and Actinobacteria (by 6.6%) (Figure 7d).
Considering OTU ≥ 1%, the most representative at the class level turned out to be: Alphaproteobacteria, Gammaproteobacteria, and Betaproteobacteria from the phylum Proteobacteria; Actinobacteria from the phylum Actinobacteria; Bacilli from the phylum Firmicutes; and Sphingobacteriia from the phylum Bacteroidetes (Figure 8). The greatest differences among soil samples: A—43.785 OTU, CdA—32.744 OTU, CeA—32.760 OTU, CdCeA—42.843 OTU, S—27.133 OTU, CdS—30.325 OTU, CeS—50.320 OTU, and CdCeS—27.733 OTU, were observed in OTUs representing the class Alphaproteobacteria. Interestingly, in the soil sown with Sinapis alba L. and fertilized with cellulose (CeS) as well as in the cadmium-contaminated soil sown with Avena sativa L. (CdA), high OTU numbers were generated in the Betaproteobacteria and Sphingobacteriia classes.
Cadmium also affected the lower taxon, i.e., order (Figure 9). Actinomycetales from the phylum Actinobacteria turned out to be the prevailing order in the present study. Its abundance was high in all soil samples. Successive orders (in terms of abundance) representing the phylum Proteobacteria were predominated by Sphingomonadales and Xanthomonadales. High abundance was also demonstrated for the order Bacillales belonging to the phylum Firmicutes and the order Sphingobacteriales from the phylum Bacteroidetes. Worthy of attention is the order Pseudomonadales from the phylum Proteobacteria, which turned out to be abundant in the soil sown with Sinapis alba L. fertilized with cellulose and contaminated with cadmium (CdCeS).
The greatest changes identified in the structure of bacteria classified to families were observed for Xanthomonadaceae, Sphingomonadaceae, and Sphingobacteriaceae (Figure 10). The Pseudomonadaceae family bacteria turned out to be highly abundant in the soil samples contaminated with cadmium, fertilized with cellulose, and used to grow both crops studied (CdCeS and CdCeA).
The OTU number assigned to individual bacterial genera was found to depend on crop species grown, cellulose fertilization, and soil contamination with cadmium (Figure 11). The prevailing genus in the soil sown with Sinapis alba L. turned out to be: Kaistobacter, Bacillus, Sphingomonas Arthrobacter, Lysobacter, and Luteimonas. Soil contamination with cadmium led to diminished abundance of the Arthrobacter, Lysobacter and Luteimonas genus bacteria and increased abundance of Nostoc and Devosia, whereas cellulose application to the noncontaminated soil had the most beneficial effect on the Kaistobacter, Pedobacter, Mycoplana, and Novosphingobium genus bacteria. This positive impact of cellulose on the mentioned bacterial genera was substantially diminished by soil contamination with cadmium. Bacteria most abundant in the soil sown with Avena sativa L. were representatives of the following genera: Kaistobacter, Bacillus, Gemmatimonas, Luteimonas, Sphingomonas, and Streptomyces. Except for Luteimonas, all of them were adversely affected by cadmium. In contrast, soil contamination with cadmium promoted the development of bacteria from the following genera: Pedobacter, Sphingobium, Achromobacter, Pseudomonas, Arenimonas, Rhodoplanes, Luteimonas, and Thermomonas. Soil fertilization with cellulose modified the OTUs of individual bacterial genera, but did not mitigate the adverse effects of cadmium on Kaistobacter, Bacillus, Gemmatimonas, Sphingomonas, and Streptomyces.
The Venn analysis allowed identifying unique bacterial genera typical of particular soils under study (Figure 12, Figure 13 and Figure 14). In the case of the soil sown with Sinapis alba L., the prevailing genus identified in the noncontaminated and nonfertilized samples (S) turned out to be Haliangium; the prevailing genera in the cadmium-contaminated soil (CdS) included: Devosia, Azospirillum, Candidatus, and Solibacter; those in the soil with cellulose addition (CeS) included: Ramlibacter, Novosphingobium, Mycoplana, Singulisphaera, Adhaeribacter, Janthinobacterium, Pedobacter, Nitrosovibrio, and Alicyclobacillus; whereas, those identified in the soil additionally contaminated with cadmium (CeSCd) included: Conexibacter, Paenibacillus, Streptosporangium, Sphingobium, Achromobacter, and Pseudomonas.
The common genera for all soil samples included: Bacillus, Sphingomonas, and Streptomyces (Figure 12). In the case of the soil sown with Avena sativa L., Conexibacter, Candidatus, Solibacter, and Flavisolibacter were the unique genera in the noncontaminated soil samples (A); Candidatus, Scalindua, and Planctomyces—in the noncontaminated soil fertilized with cellulose (CeA); Azospirillum, Arenimonas, Chitinophaga, Stenotrophomonas, Dyadobacter, Flavobacterium, Comamonas, Limnobacter, and Enterobacter—in the cadmium-contaminated soil (CdA); and Chryseobacterium—in the soil fertilized with cellulose and contaminated with cadmium (CeCdA).
In turn, Luteimonas, Thermomonas, Devosia, Mycoplana, and Pedobacter turned out to be common genera for all soil samples. The first four belong to the phylum Proteobacteria, whereas the latter to the phylum Bacteroidetes (Figure 12). Figure 14 depicts unique bacterial genera and those common for crop species, for soils fertilized with cellulose, for those contaminated with cadmium, and for those contaminated with cadmium and fertilized with cellulose. In total, 14 genera were identified as common for both crops (A and S), i.e.: Kaistobacter, Bacillus, Sphingomonas, Arthrobacter, Lysobacter, Luteimonas, Thermomonas, Chondromyces, Nocardioides, Gemmatimonas, Pseudonocardia, Cohnella, Streptomyces, and Haliangium. Three common genera were identified in the soils contaminated with cadmium (CdA and CdS): Devosia, Azospirillum, and Arenimonas. The common microbiome of the soils sown with Sinapsis alba L. and Avena sativa L. fertilized with cellulose (CeA and CeS) included 15 genera: Kaistobacter, Bacillus, Sphingomonas, Arthrobacter, Luteimonas, Thermomonas, Nocardioides, Gemmatimonas, Streptomyces, Ramlibacter, Novosphingobium, Mycoplana, Singulisphaera, Pedobacter, and Alicyclobacillus, whereas that found in the soils contaminated with cadmium and fertilized with cellulose (CdCeA and CdCeS) included four genera: Sphingomonas, Sphingobium, Achromobacter, and Pseudomonas.
In summary of the above findings, it may be concluded that the changes observed in the genetic diversity of bacteria upon the influence of soil contamination with cadmium and its fertilization with cellulose, and upon cultivation of Sinapis alba L. and Avena sativa L. were reflected in the values of the Shannon–Wiener (Table 7) and Simpson (Table 8) indices.
Table 7.
Effect of cadmium (Cd) and cellulose (Ce) on the values of Shannon–Wiener diversity index of bacteria in the soil.
Table 8.
Effect of cadmium (Cd) and cellulose (Ce) on the values of Simpson diversity index of bacteria in the soil.
The Shannon–Wiener and Simpson indices are one of the most popular indices used to determine biodiversity. They provide information about population numbers and homogeneity of organisms constituting a given community. The higher their values are, the greater is the diversity. Cadmium exposure decreased the value of the Shannon–Wiener index in the soil sown with Avena sativa L., but did not modify it in the soil sown with Sinapis alba L. (Table 7). In turn, cellulose application to the noncontaminated soil used to grow Sinapis alba L. diminished the genetic diversity of bacteria at all taxonomic levels, while an opposite effect was observed in the soil sown with Avena sativa L. Cellulose application to the cadmium-contaminated soil did not improve bacterial diversity, regardless of crop species grown in the soil.
The Simpson’s index values confirm the effects of cadmium and cellulose on bacterial diversity (Table 8) indicated by Shannon–Wiener index values.
4. Discussion
4.1. Plant Response to Soil Contamination with Cadmium and Fertilization with Cellulose
Although cadmium is commonly believed to be a toxic element inhibiting plant growth and development [61], some authors [30,31,62] have concluded that its excess in the soil elicited a positive effect on the growth and development of grasses and legumes. In the study by Xu and Wang [63], Poa pratensis and Festuca arundinacea responded with a growth of aerial parts to cadmium doses of 50 and 100 mg Cd2+ kg−1, whereas Zhang et al. [64] reported the growth of aerial parts of Pennisetum americanum and Pennisetum purpureum in response to cadmium doses ranging from 1 to 8 mg Cd2+ kg−1. Both the main crop—Sinapis alba L. and the successive crop—Avena sativa L., tested in the present study, were relatively resistant to the applied cadmium doses. This resistance may be due to greater tolerance of certain species belonging to Poaceae and Brassicaceae families to the contamination with this heavy metal developed through, e.g., synthesis of protein substances (phytochelatins), which neutralize cadmium by its fixation [65,66]. The present study results and the findings reported by Deram et al. [67]; Aibibu et al. [68]; Zhang et al. [69]; Xu, Wang [63]; Guo et al. [70]; Zhang et al. [71]; Quezada Hinojosa et al. [72]; and Stanisławska-Głubiak et al. [73] prove that greater amounts of cadmium are accumulated in the roots than in aerial parts of plants. This may be due to the activation of defense mechanisms counteracting stress posed by cadmium presence in the soil, which is manifested by the inhibited transport of this heavy metal from roots to aerial parts [74]. Soil fertilization with cellulose mitigated the adverse effects of cadmium only in the case of Sinapis alba L., whereas the response of Avena sativa L. to cadmium was similar regardless of polysaccharide application. This is most likely due to variable physical stability resulting from various abnormalities, such as, e.g., collapses, microfibril kinks or voids (micropores), and the low capacity of unmodified cellulose to adsorb heavy metals [75,76,77]. Summarizing, it may be concluded that cellulose mitigated the adverse effect of cadmium on the growth of aerial parts only in the case of the main crop—Sinapis alba L., whereas the response of the successive crop—Avena sativa L., was similar, regardless of soil amendment with the polysaccharide.
4.2. Response of Microorganisms to Soil Contamination with Cadmium and Fertilization with Cellulose
Microorganisms not only play a key role in the course of biochemical processes in soil but may also affect the health status of plants by synthesizing phytohormones, increasing availability of nutrients, facilitating nutrient uptake by plants, and reducing toxicity of heavy metals in plants [28,29]. However, these functions may be impaired by various pollutants, such as heavy metals (including cadmium), pervading the natural environment and may be manifested by reduced abundance and diversity of microorganisms, which significantly affects the appropriate soil quality [6,9,10]. According to Zaborowska et al. [78] and Haddad et al. [35], cadmium may exert various effects on the growth and development of microorganisms; however, these effects are mainly determined by the contamination level. The present study results demonstrate that cadmium contributed to diminished abundance of organotrophic bacteria, actinobacteria, and fungi, as indicated by negative coefficients of correlation between the extent of soil contamination with cadmium and population numbers of the studied microorganisms, and by negative values of the cadmium effect index (IFCd). Cadmium not only affects the abundance of soil microbiota but may also trigger changes in their biodiversity [79]. Regardless of cadmium dose, in the case of the soil samples not amended without cellulose, higher values of the colony development index (CD) were determined for organotrophic bacteria and actinobacteria in pots sown with Sinapis alba L., and those of fungi in those sown with Avena sativa L. Soil fertilization with cellulose exerted various effects on the microorganisms as it decreased CD of bacteria, increased that in fungi, and had no significant effect on CD of actinobacteria. Values of the ecophysiological diversity index (EP) also changed upon cadmium influence. This heavy metal contributed to lower EP values computed for organotrophic bacteria and actinobacteria, and to higher EP values noted for fungi in the soil not fertilized with cellulose, whereas its effects varied in the fertilized soil. Boros-Lajszner et al. [50,80] reported similar results in their studies into the effect of nickel, cobalt, and cadmium on soil microbiome diversity. Similar conclusions were formulated by Wyszkowska et al. [49] in their experiment with heavy metals. Soil microorganisms usually respond rapidly to changes in the environment; hence, they are deemed to be viable indicators of soil quality and fertility [81,82]. In the present study, cellulose application to the soil increased the ecophysiological diversity of organotrophic bacteria. This increase was due to the fact that the organotrophs were predominated by r-strategists [42]. Organotrophic bacteria respond rapidly to the supply of nutrients. Hence, they play an important role in cellulose degradation to simple sugars [35,37].
Rhizospheres of Sinapis alba L. and Avena sativa L. were characterized by a specific composition of bacterial communities, which suggests that each species may develop a unique microbiome in the soil contaminated with cadmium [83]. Despite the above finding, the prevailing bacterial phyla identified in the soil used to grow both crop species were Proteobacteria and Actinobacteria, followed by Firmicutes and Bacteroidetes. This is consistent with findings reported by Hou et al. [84], Fu et al. [85], and Wang et al. [86]. According to Min et al. [87], bacteria belonging to these phyla play a key role in the restoration of microbial communities in the cadmium-contaminated soil. In the present study, the relative abundance of Proteobacteria and Actinobacteria was higher in the soil fertilized with cellulose. As posited by Li et al. [88], this may suggest that the higher organic matter content facilitates bacteria survival in extreme conditions [89]. Microorganisms which respond rapidly to changes in nutrients in the rhizosphere are fast-growing microorganisms (r-strategists) [90]. Actinobacteria—being k-strategists, are the so-called slow-growing microorganisms [91,92] and together with Firmicutes, Bacteroidetes and Acidobacterie may be actively engaged in organic matter transformation [93,94]. Therefore, a hypothesis may be advanced that the changes in bacteria abundance may increase tolerance to and accumulation of cadmium in the soil sown with Sinapis alba L. and Avena sativa L.
In the present study, the most tolerant to soil contamination with Cd2+ turned out to be Alphaproteobacteria, Betaproteobacteria, and Gammaproteobacteria from the phylum Proteobacteria together with Actinobacteria and Bacilli from the phylum Firmicutes. Additionally, Duan et al. [95] confirmed their high tolerance to contamination with Cd2+. Previous investigations by Liang et al. [96] and Borowik et al. [97] proved high resistance of the bacteria from Alphaproteobactera, Betaproteobactera, and Gammaproteobacteria to contaminants.
The present study results showed that soil contamination with cadmium led to a reduction in OTU numbers of bacteria representing the Luteimonas, Thermomonas, and Devosia genera from the phylum Proteobacteria, together with Nostoc bacteria belonging to the phylum Cyanobacteria. The bacteria from the genus Luteimonas and Thermomonas, and Nocardioides belonging to the phylum Actinobacteria, were also highly resistant to soil contamination with Cd2+. Bacteria from the genus Luteimonas were identified in soils amended with compost produced from food waste [98], in wastewater [99], in dump soils [100], and in fresh litter material at poultry farms [101], which suggests their high capability for degradation of insoluble large-molecular-weight organic matter. Likewise, Thermomonas genus bacteria are involved in degradation of sugar cane straw and goat manure [102]. These bacteria also colonize soils contaminated with heavy metals [50,80] and petroleum-based products [103,104,105,106], exhibiting a high bioremediating potential. In our previous studies [50,80], the Thermomonas genus bacteria also showed high tolerance to soil contaminated with Ni2+, Co2+, and Cd2+, and therefore should be perceived as potential microorganisms useful in bioaugmentation of soils contaminated with these heavy metals. Bacteria from the genus Nocardioides degrade aromatic amines, using them as a source of carbon, nitrogen, and energy. They boost their growth with vitamins, which are an indispensable micronutrient aiding the biodegradation process [107]. Microorganisms using C from the organic material take the greatest part in modeling ecosystems because they facilitate organic matter degradation and increase humus content by degrading cellulose, thereby reinforcing interactions between microorganisms [108]. In the present study, the fertilization of cadmium-contaminated soil with cellulose had definitely the most positive effect on the bacteria from Kaistobacter Mycoplana, and Novosphingobium genera belonging to the class Alphaproteobacteria, phylum Proteobacteria, together with Pedobacter belonging to the class Sphingobacteriia, phylum Bacteroidetes.
An advantage of the Kaistobacter genus bacteria is their potential for alleviating plant diseases, e.g., in tobacco [109]; Mycoplana genus bacteria were demonstrated to positively affect cultivation of orchard grass in petroleum-polluted soil [110], whereas Pedobacter bacteria enhanced tomato resistance to bacterial wilting [111]. This fundamental importance of reducing the risk of plant disease development in sustainable agriculture is suggested by a possible approach to modeling disease suppression rates [109,112]. The analysis of microorganisms at the genus level allowed identifying bacteria from the Sphingomonas, Sphingobium, Achromobacter, and Pseudomonas genera belonging to the phylum Proteobacteria, which were common for the soils sown with Sinapis alba L. and Avena sativa L. fertilized with cellulose and contaminated with cadmium. According to Altimira et al. [112] and Nilgiriwala et al. [113], Sphingomonas encode genes of multiple oxidases and enable Cd recovery from soil by means of enzymatic bioprecipitation. Likewise, the species belonging to the genus Pseudomonas are usually involved in the nitrogen cycle and contaminant degradation, and may enhance plant growth and development [114,115]. In turn, Gomez-Balderas et al. [116] demonstrated that soil contamination with Cd2+ might significantly diminish the relative abundance of Pseudomonas.
Qiong et al. [117] posited that bacteria response to cadmium contamination depended on soil type. The fluvoaquic soil severely contaminated with cadmium was mainly populated by Novosphingobium, Sphingobium, Pseudomonas, Rheinheimera, and Corynebacterium genera. Therefore, it is very important to continue research that would inform about the response of microorganisms in different types of soil and different regions, to identify the possibilities of improving the condition of contaminated soils.
5. Conclusions
Soil contamination with cadmium doses ranging from 4 to 16 mg Cd2+ kg−1 soil did not impair the growth and development of Sinapis alba L. and Avena sativa L., which proves that both crop species may be harnessed for the phytoremediation of soils contaminated with this metal. Given the greater capability of Avena sativa L. than Sinapis alba L. for Cd2+ accumulation, it was found more viable for soil remediation from the ecological standpoint as it reduces the risk of cadmium penetration into the food chain. Although Cd2+-contaminated soil fertilization with cellulose stimulated the proliferation of microorganisms, it did not mitigate the adverse effect of Cd2+ on bacterial diversity. Soil phytoremediation by Avena sativa L. may be aided by biostimulation with cellulose coupled with bioaugmentation using Chryseobacterium bacteria, whereas phytoremediation by Sinapis alba L., by bioaugmentation with the Sphingomonas, Sphingobium, Achromobacter, and Pseudomonas genera bacteria, which represented the unique microbiome in the soil contaminated with Cd2+ and fertilized with cellulose.
Author Contributions
J.W., E.B.-L. and J.K., conceptualization; J.W., E.B.-L., A.B. and J.K., methodology and formal analysis; E.B.-L., experiments; A.B. bioinformatic analysis and visualization of data; J.W., E.B.-L. and A.B., writing—review and editing; J.K., supervision. All authors contributed significantly to the discussion of the results and the preparation of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The results presented in this paper were obtained as part of a comprehensive study financed by the University of Warmia and Mazury in Olsztyn, Faculty of Agriculture and Forestry, Department of Soil Science and Microbiology (grant no. 30.610.006-110) and by a project financially supported by the Minister of Education and Science under the program entitled “Regional Initiative of Excellence” for the years 2019–2023, Project No. 010/RID/2018/19, amount of funding 12,000,000 PLN.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rajaganpathy, V.; Xavier, F.; Sreekumar, D.; Mandal, P.K. Heavy metal contamination in soil, water and fodder and their presence in livestock and products: A review. J. Environ. Sci. Technol. 2011, 4, 234–249. [Google Scholar] [CrossRef]
- Dhaliwal, S.S.; Taneja, P.K.; Singh, J.; Bhatti, S.S.; Singh, R. Cadmium accumulation potential of brassica species grown in metal spiked loamy sand soil. Soil Sediment. Contam. Int. J. 2020, 29, 638–649. [Google Scholar] [CrossRef]
- Setia, R.; Dhaliwal, S.S.; Kumar, V.; Singh, R.; Kukal, S.S.; Pateriya, B. Impact assessment of metal contamination in surface water of Sutlej River (India) on human health risks. Environ. Pollut. 2020, 265, 114907. [Google Scholar] [CrossRef]
- Setia, R.; Dhaliwal, S.S.; Singh, R.; Kumar, V.; Taneja, S.; Kukal, S.S.; Pateriya, B. Phytoavailability and human risk assessment of heavy metals in soils and food crops around Sutlej River, India. Chemosphere 2020, 263, 128321. [Google Scholar] [CrossRef]
- McLaughlin, M.J.; Smolders, E.; Zhao, F.J.; Grant, C.; Montalvo, D. Managing cadmium in agricultural systems. Adv. Agron. 2021, 166, 1–129. [Google Scholar] [CrossRef]
- Raza, A.; Habib, M.; Kakavand, S.N.; Zahid, Z.; Zahra, N.; Sharif, R.; Hasanuzzaman, M. Phytoremediation of cadmium: Physiological, biochemical, and molecular mechanisms. Biology 2020, 9, 177. [Google Scholar] [CrossRef]
- Bhatti, S.S.V.; Kumar, V.; Sambyal, J.; Singh, N.A.K. Comparative analysis of tissue compartmentalized heavy metal uptake by common forage crop: A field experiment. Catena 2018, 160, 185–193. [Google Scholar] [CrossRef]
- Kubier, A.; Wilkin, R.T.; Pichler, T. Cadmium in soils and groundwater: A review. Appl. Geochem. 2019, 108, 104388. [Google Scholar] [CrossRef]
- Moreno-Caselles, J.; Moral, R.; Perez-Espinosa, A.; Perez-Murcia, M.D. Cadmium accumulation and distribution in cucumber plant. J. Plant Nutr. 2000, 23, 243–250. [Google Scholar] [CrossRef]
- Watanabe, T.; Murata, Y.; Osaki, M. Amaranthus tricolor has the potential for phytoremediation of cadmium contaminated soils. Commum. Soil Sci. Plant Anal. 2009, 40, 3158–3169. [Google Scholar] [CrossRef]
- Ejnik, J.; Robinson, J.; Zhu, J.; Försterling, H.; FrankShaw, C.; Petering, D.H. Folding pathway of apo-metallothionein induced by Zn2+, Cd2+ and Co2+. J. Inorg. Biochem. 2002, 88, 144–152. [Google Scholar] [CrossRef]
- Pedro, F.; Feria-Cáceres, P.F.; Penagos-Velez, L.; Claudia, X.; Moreno-Herrera, C.X. Tolerance and cadmium (Cd) immobiliza-tion by native bacteria isolated in cocoa soils with increased metal content. Microbiol. Res. 2022, 13, 556–573. [Google Scholar] [CrossRef]
- Azizollahi, Z.; Ghaderian, S.M.; Ghotbi-Ravandi, A.A. Cadmium accumulation and its effects on physiological and biochemical characters of summer savory (Satureja hortensis L.). Int. J. Phytoremediat. 2019, 21, 1241–1253. [Google Scholar] [CrossRef]
- DalCorso, G.; Farinati, S.; Furini, A. Regulatory networks of cadmium stress in plants. Plant Signal. Behav. 2010, 5, 663–667. [Google Scholar] [CrossRef]
- Mendoza-Cozatl, D.G.; Butko, E.; Springer, F.; Torpey, J.W.; Komives, E.A.; Kehr, J.; Schroeder, J.I. Identification of high levels of phytochelatins, glutathione and cadmium in the phloem sap of Brassica napus. A role for thiol-peptides in the long-distance transport of cadmium and the effect of cadmium on iron translocation. Plant J. 2008, 54, 249–259. [Google Scholar] [CrossRef]
- Lux, A.; Martinka, M.; Vaculík, M.; White, P.J. Root responses to cadmium in the rhizosphere: A review. J. Exp. Bot. 2011, 62, 21–37. [Google Scholar] [CrossRef]
- Agents Classified by the IARC Monographs, Volumes 1–132. Available online: https://monographs.iarc.who.int/agents-classified-by-the-iarc/ (accessed on 12 August 2022).
- Conesa, H.M.; Wieser, M.; Gasser, M.; Hockmann, K.; Evangelou, M.W.H.; Studer, B. Effects of three amendments on extractability and fractionation of Pb, Cu, Ni and Sb in two shooting range soils. J. Hazard. Mater. 2010, 181, 845–850. [Google Scholar] [CrossRef]
- Bolan, N.B.; Kunhikrishnanc, A.; Thangarajana, R.; Kumpiened, J.; Parke, J.; Makinof, T.; Kirkhamg, M.B.; Scheckelh, K. Remediation of heavy metal(loid)s contaminated soils—To mobilize or to immobilize? J. Hazard. Mater. 2014, 266, 141–166. [Google Scholar] [CrossRef]
- Stritsis, B.; Steingrobe, B.; Claassen, N. Cadmium fractions in an acid sandy soil and Cd in soil solution as affected by plant growth. J. Plant Nutr. Soil Sci. 2014, 177, 431–437. [Google Scholar] [CrossRef]
- Oves, M.; Saghir Khan, M.; Zaidi, A.; Ahmad, E. Soil contamination, nutritive value, and human health risk assessment of heavy metals: An overview. In Toxicity of Heavy Metals to Legumes and Bioremediation; Zaidi, A., Wani, P.A., Khan, M.S., Eds.; Springer: Vienna, Austria, 2012; pp. 1–27. [Google Scholar] [CrossRef]
- Kabata-Pendias, A.; Pendias, H. Trace Elements in Soils and Plants; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar] [CrossRef]
- Li, M.; Mohamed, I.; Raleve, D.; Chen, W.; Huang, Q. Field evaluation of intensive compost application on Cd fractionation and phytoavailability in a mining-contaminated soil. Environ. Geochem. Health 2016, 38, 1193–1201. [Google Scholar] [CrossRef]
- Massaccesi, L.; Benucci, G.M.N.; Gigliotti, G.; Cocco, S.; Corti, G.; Agnelli, A. Rhizosphere effect of three plant species of environment under periglacial conditions (Majella Massif, central Italy). Soil Biol. Biochem. 2015, 89, 184–195. [Google Scholar] [CrossRef]
- Hütsch, B.W.; Augustin, J.; Merbach, W. Plant rhizodeposition—An important source for carbon turnover in soils. J. Plant. Nutr. Soil Sci. 2002, 165, 397–407. [Google Scholar] [CrossRef]
- Tahervand, S.; Jalali, M. Sorption, desorption, and speciation of Cd, Ni, and Fe by four calcareous soils as affected by pH. Environ. Monit. Assess. 2016, 188, 322. [Google Scholar] [CrossRef] [PubMed]
- Karimzadeh, L.; Nair, S.; Merkel, B.J. Effect of microbial siderophore DFOB on Pb, Zn, and Cd sorption onto zeolite. Aquat. Geochem. 2013, 19, 25–37. [Google Scholar] [CrossRef]
- Amabilis-Sosa, L.E.; Siebe, C.; Moeller-Chávez, G.; del Carmen Durán-Domínguez-de-Bazúa, M. Accumulation and distribution of lead and chromium in laboratory-scale constructed wetlands inoculated with metal-tolerant bacteria. Int. J. Phytoremed. 2015, 17, 1090–1096. [Google Scholar] [CrossRef]
- Lebeau, T.; Jesequel, K.; Braud, A. Bioaugmentation-assisted phytoextraction applied to metal-contaminated soils: State of the art and future prospects. In Microbes and Microbial Technology: Agricultural and Environmental Applications; Ahmad, I., Ahmad, F., Pichtel, J., Eds.; Microbes and Microbial Technology; Springer: New York, NY, USA, 2011. [Google Scholar] [CrossRef]
- Lange, B.; van der Ent, A.; Baker, A.J.M.; Echevarria, G.; Mahy, G.; Malaisse, F.; Meets, P.; Pourret, O.; Verbruggen, N.; Faucon, M.-P. Copper and cobalt accumulation in plants: A critical assessment of the current state of knowledge. New Phytol. 2017, 213, 537–551. [Google Scholar] [CrossRef]
- Sarwar, N.; Imran, M.; Shaheen, M.R.; Ishaque, W.; Kamran, M.A.; Matloob, A.; Rehim, A.; Hussain, S. Phytoremediation strategies for soils contaminated with heavy metals: Modifications and future perspectives. Chemosphere 2017, 171, 710–721. [Google Scholar] [CrossRef]
- Boros-Lajszner, E.; Wyszkowska, J.; Kucharski, J. Use of zeolite to neutralise nickel in a soil environment. Environ. Monit. Assess. 2018, 190, 54. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Gómez-Sagasti, M.T.; Alkorta, I.; Becerril, J.M.; Epelde, L.; Anza, M.; Garbisu, C. Microbial monitoring of the recovery of soil quality during heavy metal phytoremediation. Water Air Soil Pollut. 2012, 223, 3249–3262. [Google Scholar] [CrossRef]
- Haddad, S.A.; Lemanowicz, J.; Abd El-Azeim, M.M. Cellulose decomposition in clay and sandy soils contaminated with heavy metals. Int. J. Environ. Sci. Technol. 2019, 16, 3275–3290. [Google Scholar] [CrossRef]
- Chmolowska, D.; Hamda, N.; Laskowski, R. Cellulose decomposed faster in fallow soil than in meadow soil due to a shorter lag time. J. Soils Sediments 2017, 17, 299–305. [Google Scholar] [CrossRef]
- Xie, Y.; Fan, J.; Zhu, W.; Amombo, E.; Lou, Y.; Chen, L.; Fu, J. Effect of heavy metals pollution on soil microbial diversity and bermudagrass genetic variation. Front Plant Sci. 2016, 7, 755. [Google Scholar] [CrossRef] [PubMed]
- Tica, D.; Udovic, M.; Lestan, D. Immobilization of potentially toxic metals using different soil amendments. Chemosphere 2011, 85, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, L.; Luo, Z.; Shen, J.; Ni, Q.; Yao, J. Facile preparation of a cellulose-based bioadsorbent modified by hPEI in heterogeneous system for high-efficiency removal of multiple types of dyes. React. Funct. Polym. 2018, 125, 77–83. [Google Scholar] [CrossRef]
- Vasile, G.-G.; Tenea, A.-G.; Dinu, C.; Iordache, A.M.M.; Gheorghe, S.; Mureseanu, M.; Pascu, L.F. Bi-oavailability, accumulation and distribution of toxic metals (As, Cd, Ni and Pb) and their impact on Sinapis alba plant nutrient metabolism. Int. J. Environ. Res. Public Health 2021, 18, 12947. [Google Scholar] [CrossRef]
- Jan, R.; Asaf, S.; Numan, M.; Lubna; Kim, K.-M. Plant Secondary metabolite biosynthesis and transcriptional regulation in response to biotic and abiotic stress conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- De Leij, F.A.A.M.; Whips, J.; Lynch, J.M. The use of colony development for the characterization of bacterial communities in soil and on roots. Microb. Ecol. 1993, 7, 81–97. [Google Scholar] [CrossRef]
- Sarathchandra, S.U.; Burch, G.; Cox, N.R. Growth patterns of bacterial communities in the rhizoplane and rhizosphere of white clover (Trifolium repens L.) and perennial ryegrass (Lolium perenne L.) in long-term pasture. Appl. Soil Ecol. 1997, 6, 293–299. [Google Scholar] [CrossRef]
- PN-R-04032; In Soil and Mineral materials—Sampling and Determination of Particle Size Distribution. Polish Committee for Standardization: Warsaw, Poland, 1998.
- ISO 11464; Soil Quality—Pre-Treatment of Samples for Physico-Chemical Analysis. International Organization for Standardization: Geneva, Switzerland, 2006.
- ISO 10390; Soil Quality—Determination of PH. International Organization for Standardization: Geneva, Switzerland, 2005.
- PN-EN 14084: 2004 (N); Nickel, Cadmium and Cobalt Content In Above-Ground Parts and in Roots Determined by Flame Atomic Absorption Spectrometry and by Graphite Furnace Atomic Absorption Spectroscopy (FAAS and GFAAS) following Microwave Mineralization. International Organization for Standardization: Geneva, Switzerland, 2016. Available online: https://pzn.pkn.pl/kt/info/published/9000128800 (accessed on 20 April 2021).
- Orwin, K.H.; Wardle, D.A. New indices for quantifying the resistance and resilience of soil biota to exogenous disturbance. Soil Biol. Biochem. 2004, 36, 1907–1912. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Boros-Lajszner, E.; Kucharski, J. Calorific value of Festuca rubra biomass in the phytostabilization of soil contaminated with nickel, cobalt and cadmium which disrupt the microbiological and biochemical properties of soil. Energies 2022, 15, 3445. [Google Scholar] [CrossRef]
- Boros-Lajszner, E.; Wyszkowska, J.; Borowik, A.; Kucharski, J. The response of the soil microbiome to contamination with cadmium, cobalt an nickel in soil sown with Brassica napus. Minerals 2021, 11, 498. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J.; Kucharski, J. Bacteria and soil enzymes supporting the valorization of forested soils. Materials 2022, 15, 3287. [Google Scholar] [CrossRef] [PubMed]
- Dell Inc. Dell Statistica (Data Analysis Software System), version 13.1; Dell Inc.: Tulsa, OK, USA, 2016.
- Parks, D.H.; Tyson, G.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef]
- Krzywiński, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- R Studio Team. R Studio: Integrated Development for R; R Studio Inc.: Boston, MA, USA, 2019; Available online: http://www.rstudio.com/ (accessed on 10 May 2022).
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 10 May 2022).
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, M.; Moeller, S.; et al. Various R Programming Tools for Plotting Data. R Package Version 3.1.3. 2022. Available online: https://CRAN.R-project.org/package=gplots (accessed on 14 July 2022).
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Shannon, V.L.; Vanguelova, E.I.; Morison, J.I.L.; Shaw, L.J.; Clark, J.M. The contribution of deadwood to soil carbon dynamics in contrasting temperate forest ecosystems. Eur. J. For. Res. 2021, 141, 241–252. [Google Scholar] [CrossRef]
- Bibi, F.; Ali, Z. Measurement of diversity indices of avian communities at Taunsa Barrage Wildlife Sanctuary, Pakistan. J. Anim. Plant Sci. 2013, 23, 469–474. [Google Scholar]
- Nazar, R.; Iqbal, N.; Masood, A.; Khan, M.I.R.; Syeed, S.; Khan, N.A. Cadmium toxicity in plants and role of mineral nutrients in its alleviation. Am. J. Plant Sci. 2012, 3, 1476–1489. [Google Scholar] [CrossRef]
- Sobariu, D.L.; Fertu, D.I.T.; Diaconu, M.; Pavel, I.V.; Hlihor, R.-M.; Dragoi, E.N.; Curteanu, S.; Lenz, M.; Corvini, P.F.-X.; Gavrilescu, M. Rhizobacteria and plant symbiosis in heavy metals uptake and its implications for soil bioremediation. New Biotechnol. 2017, 39, 125–134. [Google Scholar] [CrossRef]
- Xu, P.; Wang, Z. Physiological mechanism of hypertolerance of cadmium in Kentucky bluegrass and tall fescue: Chemical forms and tissue distribution. Environ. Exp. Bot. 2013, 96, 35–42. [Google Scholar] [CrossRef]
- Zhang, X.; Xia, H.; Li, Z.; Zhung, P.; Gao, B. Potential of four grasses in remediation of Cd and Zn contaminated soils. Bioresour. Technol. 2010, 10, 2063–2066. [Google Scholar] [CrossRef] [PubMed]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Li, H.; Jiang, R.; Römheld, V.; Zhang, F.; Zhao, F.-J. The role of root hairs in cadmium acquisition by barley. Environ. Pollut. 2011, 159, 408–415. [Google Scholar] [CrossRef]
- Deram, A.; Denayer, F.O.; Petit, D.; Van Haluvyn, C. Seasonal variation of cadmium and zinc in Arrhenatherum elatius, a perennial grass species from highly contaminated soils. Environ. Pollut. 2006, 140, 62–70. [Google Scholar] [CrossRef]
- Aibibu, N.; Liu, Y.; Zeng, G.; Wang, X.; Chen, B.; Song, H.; Xu, L. Cadmium accumulation in Vetiveria zizanioides and its effects on growth, physiological and biochemical characters. Bioresour. Technol. 2010, 101, 6297–6303. [Google Scholar] [CrossRef]
- Zhang, S.; Lin, H.; Deng, L.; Gong, G.; Jia, Y.; Xu, X.; Li, T.; Li, Y.; Chen, H. Cadmium tolerance and accumulation characteristic Siegesbeckia orientalis L. Ecol. Eng. 2013, 5, 133–139. [Google Scholar] [CrossRef]
- Guo, Q.; Meng, L.; Mao, P.C.; Tian, X.X. An assessment of Agropyron cristatum tolerance to cadmium contaminated soil. Biol. Plant. 2014, 58, 174–178. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, B.; Xia, H. Effect of cadmium on growth, photosynthesis, mineral nutrition and metal accumulation of bana grass and vetiver grass. Ecotoxicol. Environ. Saf. 2014, 106, 102–108. [Google Scholar] [CrossRef]
- Quezada-Hinojosa, R.; Föllmi, K.B.; Gillet, F.; Matera, V. Cadmium accumulation in six common plant species associated with soils containing high geogenic cadmium concentrations at Le Gurnigel, Swiss Jura Mountains. Catena 2015, 124, 85–96. [Google Scholar] [CrossRef]
- Stanislawska-Glubiak, E.; Korzeniowska, J.; Kocon, A. Effect of the reclamation of heavy metal-contaminated soil on growth of energy willow. Pol. J. Environ. Stud. 2012, 21, 187–192. [Google Scholar]
- Masarovicova, E.; Kralova, K.; Kummerova, M. Principles of classification of medical plants as hyperaccumulators or excluders. Acta Physiol. Plant. 2010, 32, 823–829. [Google Scholar] [CrossRef]
- O’Connell, D.W.; Birkinshaw, C.; O’Dwyer, T.F. Heavy metal adsorbents prepared from the modification of cellulose: A review. Bioresour. Technol. 2008, 99, 6709–6724. [Google Scholar] [CrossRef] [PubMed]
- Sud, D.; Mahajan, G.; Kaur, M.P. Agricultural waste material as potential adsorbent for sequestering heavy metal ions from aqueous solutions—A review. Bioresour. Technol. 2008, 99, 6017–6027. [Google Scholar] [CrossRef]
- Hashim, M.A.; Mukhopadhyay, S.; Sahu, J.N.; Sengupta, B. Remediation technologies for heavy metal contaminated groundwater. J. Environ. Manag. 2011, 92, 2355–2388. [Google Scholar] [CrossRef]
- Zaborowska, M.; Kucharski, J.; Wyszkowska, J. Using basalt flour and brown algae to improve biological properties of soil contaminated with cadmium. Soil Water Res. 2015, 10, 181–188. [Google Scholar] [CrossRef]
- Wei, Y.; Zhao, Y.; Zhao, X.; Gao, X.; Zheng, Y.; Zuo, H.; Wei, Z. Roles of different humin and heavy-metal resistant bacteria from composting on heavy metal removal Yuquan. Bioresour. Technol. 2020, 296, 122375. [Google Scholar] [CrossRef]
- Boros-Lajszner, E.; Wyszkowska, J.; Borowik, A.; Kucharski, J. Energetic value of Elymus elongatus L. and Zea mays L. grown on soil polluted with Ni2+, Co2+, Cd2+, and Sensitivity of rhizospheric bacteria to heavy metals. Energies 2021, 14, 4903. [Google Scholar] [CrossRef]
- Abd El-Azeim, M.M.; Mohamed, W.S.; Hammam, A.A. Soil physiochemical properties in relation to heavy metals status of agricultural soils in El-Minia Governorate, Egypt. J. Soil Sci. Agric. Eng. Mansoura Univ. 2016, 7, 423–431. [Google Scholar] [CrossRef]
- Salman, S.A.; Elnazer, A.A.; El Nazer, H.A. Integrated mass balance of some heavy metals fuxes in Yaakob village, south Sohag, Egypt. Int. J. Environ. Sci. Technol. 2017, 14, 1011–1018. [Google Scholar] [CrossRef]
- Zeng, P.; Guo, Z.; Cao, X.; Xiao, X.; Liu, Y.; Shi, L. Phytostabilization potential of ornamental plants grown in soil contaminated with cadmium. Int. J. Phytoremed. 2018, 20, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Wang, K.; Liu, T.; Wang, H.; Lin, Z.; Qian, J.; Lu, L.; Tian, S. Unique rhizosphere micro-characteristics facilitate phytoextraction of multiple metals in soil by the hyperaccumulating plant Sedum alfredii. Environ. Sci. Technol. 2017, 51, 5675–5684. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.D.; Zhou, L.; Chen, P.; Du, Q.; Pang, T.; Song, C.; Wang, X.C.; Liu, W.G.; Yang, W.Y.; Yong, T.W. Effects of maize-soybean relay intercropping on crop nutrient uptake and soil bacterial community. J. Integr. Agric. 2019, 18, 2006–2018. [Google Scholar] [CrossRef]
- Wang, M.; Li, S.; Chen, S.; Meng, N.; Li, X.; Zheng, H.; Zhao, C.; Wang, D. Manipulation of the rhizosphere bacterial community by biofertilizers is associated with mitigation of cadmium phytotoxicity. Sci. Total Environ. 2019, 649, 413–421. [Google Scholar] [CrossRef]
- Min, X.; Wang, Y.; Chai, L.; Yang, Z.; Liao, Q. High-resolution analyses reveal structural diversity patterns of microbial communities in Chromite Ore Processing Residue (COPR) contaminated soils. Chemosphere 2017, 183, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sun, M.; Zhang, H.; Xu, N.; Sun, G. Use of mulberry–soybean intercropping in salt–alkali soil impacts the diversity of the soil bacterial community. Microb. Biotechnol. 2016, 9, 293–304. [Google Scholar] [CrossRef]
- Acosta-Martinez, V.; Bell, C.W.; Morris, B.E.L.; Zak, J.; Allen, V.G. Long-term soil microbial community and enzyme activity responses to an integrated cropping-livestock system in a semi-arid region. Agric. Ecosyst. Environ. 2010, 137, 231–240. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Borowik, A.; Olszewski, J.; Kucharski, J. Soil Bacterial Community and soil enzyme activity depending on the cultivation of Triticum aestivum, Brassica napus, and Pisum sativum ssp. arvense. Diversity 2019, 11, 246. [Google Scholar] [CrossRef]
- Maron, P.A.; Sarr, A.; Kaisermann, A.; Lévêque, J.; Mathieu, O.; Guigue, J.; Karimi, B.; Bernard, L.; Dequiedt, S.; Terrat, S.; et al. High microbial diversity promotes soil ecosystem functioning. Appl. Environ. Microbiol. 2018, 84, 1–13. [Google Scholar] [CrossRef]
- Pascault, N.; Ranjard, L.; Kaisermann, A.; Bachar, D.; Christen, R.; Terrat, S.; Mathieu, O.; Lévêque, J.; Mougel, C.; Henault, C.; et al. Stimulation of different functional groups of bacteria by various plant residues as a driver of soil priming effect. Ecosystems 2013, 16, 810–822. [Google Scholar] [CrossRef]
- Liu, X.; Huang, Z.; Havrilla, C.A.; Liu, Y.; Wu, G. Plant litter crust role in nutrients cycling potentials by bacterial communities in a sandy land ecosystem. Land Degrad. Dev. 2021, 32, 3194–3203. [Google Scholar] [CrossRef]
- Eichorst, S.A.; Kuske, C.R.; Schmidt, T.M. Influence of plant polymers on the distribution and cultivation of bacteria in the phylum Acidobacteria. Appl. Environ. Microbiol. 2011, 77, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.; Liu, Y.; Zhang, H.; Chen, G.; Song, J. Cadmium pollution impact on the bacterial community of haplic cambisols in Northeast China and inference of resistant genera. J. Soil Sci. Plant Nutr. 2020, 20, 1156–1170. [Google Scholar] [CrossRef]
- Liang, J.; Tang, S.Q.; Gong, J.L.; Zeng, G.M.; Tang, W.W.; Song, B.; Zhang, P.; Yang, Z.X.; Luo, Y. Responses of enzymatic activity and microbial communities to biochar/compost amendment in sulfamethoxazole polluted wetland soil. J. Hazard. Mater. 2020, 385, 121533. [Google Scholar] [CrossRef] [PubMed]
- Borowik, A.; Wyszkowska, J.; Kucharski, M.; Kucharski, J. Implications of soil pollution with diesel oil and BP petroleum with active technology for soil health. Int. J. Environ. Res. Public Health 2019, 16, 2474. [Google Scholar] [CrossRef]
- Young, C.C.; Kämpfer, P.; Chen, W.M.; Yen, W.S.; Arun, A.B.; Lai, W.A.; Shen, F.T.; Rekha, P.D.; Lin, K.Y.; Chou, J.H. Luteimonas composti sp. nov., a moderately thermophilic bacterium isolated from food waste. Int. J. Syst. Evol. Microbiol. 2007, 57, 741–744. [Google Scholar] [CrossRef]
- Siddiqi, M.Z.; Yeon, J.M.; Choi, H.; Lee, J.H.; Kim, S.Y.; Wee, J.H.; Im, W.T. Luteimonas granuli sp. nov., isolated from granules of the wastewater treatment plant. Curr. Microbiol. 2020, 77, 2002–2007. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, J.; Ma, J.; Xu, N.; Xin, F.; Zhang, W.; Zhang, H.; Dong, W.; Jiang, M. Luteimonas wenzhouensis sp. nov., a chitinolytic bacterium isolated from a landfill soil. Curr. Microbiol. 2021, 78, 383–388. [Google Scholar] [CrossRef]
- Wang, L.; Lilburn, M.; Yu, Z. Intestinal Microbiota of Broiler Chickens As Affected by Litter Management Regimens. Front. Microbiol. 2016, 7, 593. [Google Scholar] [CrossRef]
- Tayyab, M.; Islam, W.; Arafat, Y.; Pang, Z.; Zhang, C.; Lin, Y.; Waqas, M.; Lin, S.; Lin, W.; Zhang, H. Effect of sugarcane straw and goat manure on soil nutrient transformation and bacterial communities. Sustainability 2018, 10, 2361. [Google Scholar] [CrossRef]
- Galitskaya, P.; Biktasheva, L.; Blagodatsky, S.; Selivanovskaya, S. Response of bacterial and fungal communities to high petroleum pollution in different soils. Sci. Rep. 2021, 11, 164. [Google Scholar] [CrossRef] [PubMed]
- Novakovskiy, A.B.; Kanev, V.A.; Markarova, M.Y. Long-term dynamics of plant communities after biological remediation of oil-contaminated soils in Far North. Sci. Rep. 2021, 11, 4888. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Lv, J.-L.; Cai, Y.-L.; Yao, Y.-Y.; Zhang, K.; Ma, C.; Li, J.-X.; Ren, X.-Y.; Hu, J.-J.; Zhao, J.-H. Study on gaseous chlorobenzene treatment by a bio-trickling filter: Degradation mechanism and microbial community. Processes 2022, 10, 1483. [Google Scholar] [CrossRef]
- Ruan, M.; Zhang, Y.; Chai, T. Rhizosphere soil microbial properties on tetraena mongolica in the arid and semi-arid regions, China. Int. J. Environ. Res. Public Health 2020, 17, 5142. [Google Scholar] [CrossRef]
- Wang, X.; Tian, L.; Li, Y.; Zhong, C.; Tian, C. Effects of exogenous cellulose-degrading bacteria on humus formation and bacterial community stability during composting. Bioresour. Technol. 2022, 359, 127458. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, S.; Jiang, Q.; Bai, Y.; Shen, G.; Li, S.; Ding, W. Using community analysis to explore bacterial indicators for disease suppression of tobacco bacterial wilt. Sci. Rep. 2016, 6, 36773. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J.; Kucharski, J. Microbiological study in petrol-spiked soil. Molecules 2021, 26, 2664. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Lu, Y.; Lin, W.; Tian, J.; Cai, K. Biochar suppresses bacterial wilt of tomato by improving soil chemical properties and shifting soil microbial community. Microorganisms 2019, 7, 676. [Google Scholar] [CrossRef]
- Estendorfer, J.; Stempfhuber, B.; Vestergaard, G.; Schulz, S.; Rillig, M.C.; Joshi, J.; Schröder, P.; Schloter, M. Definition of core bacterial taxa in different root compartments of Dactylis glomerata, grown in soil under different levels of land use intensity. Diversity 2020, 12, 392. [Google Scholar] [CrossRef]
- Altimira, F.; Yáñez, C.; Bravo, G.; González, M.; Rojas, L.; Seeger, M. Characterization of copper-resistant bacteria and bacterial communities from copper-polluted agricultural soils of central Chile. BMC Microbiol. 2012, 12, 193. [Google Scholar] [CrossRef]
- Nilgiriwala, K.S.; Alahari, A.; Rao, A.S.; Apte, S.K. Cloning and overexpression of alkaline phosphatase PhoK from Sphingomonas sp. strain BSAR-1 for bioprecipitation of uranium from alkaline solutions. Appl. Environ. Microbiol. 2008, 74, 5516–5523. [Google Scholar] [CrossRef] [PubMed]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Lalucat, J.; Bennasar, A.; Bosch, R.; Garcia-Valdes, E.; Palleroni, N.J. Biology of Pseudomonas stutzeri. Microbiol. Mol. Biol. Rev. 2006, 70, 510–547. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Balderas, C.D.C.; Cochet, N.; Bert, V.; Tarnaud, E.; Sarde, C. 16S rDNA analysis of bacterial communities associated with the hyperaccumulator Arabidopsis halleri grown on a Zn and Cd polluted soil. Eur. J. Soil Biol. 2014, 60, 16–23. [Google Scholar] [CrossRef]
- Qiong, W.; Fengshan, P.; Xiaomeng, X.; Rafiq, M.T.; Xiao’e, Y.; Bao, C.; Ying, F. Cadmium level and soil type played a selective role in the endophytic bacterial community of hyperaccumulator Sedum alfredii Hance. Chemosphere 2021, 263, 127986. [Google Scholar] [CrossRef]














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).