
Variation in Plant Community Composition and Biomass to Macro and Micronutrients and Salinity across Egypt’s Five Major Coastal Lakes

 ,
, 
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
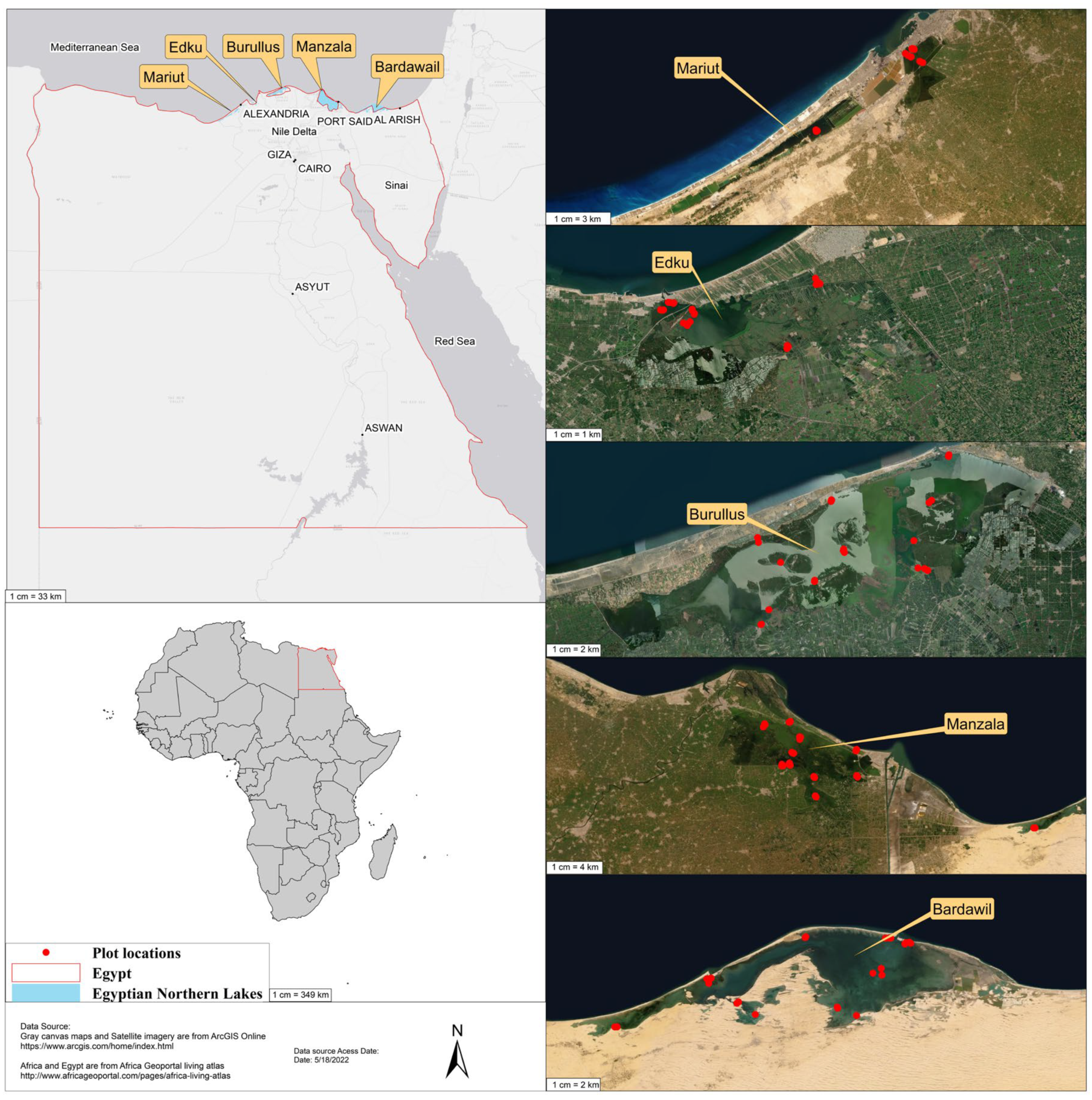
2.1. Study Area
2.2. Vegetation and Aboveground Biomass
2.3. Sediment, Plant, and Water Analyses
2.4. Data Analyses
3. Results
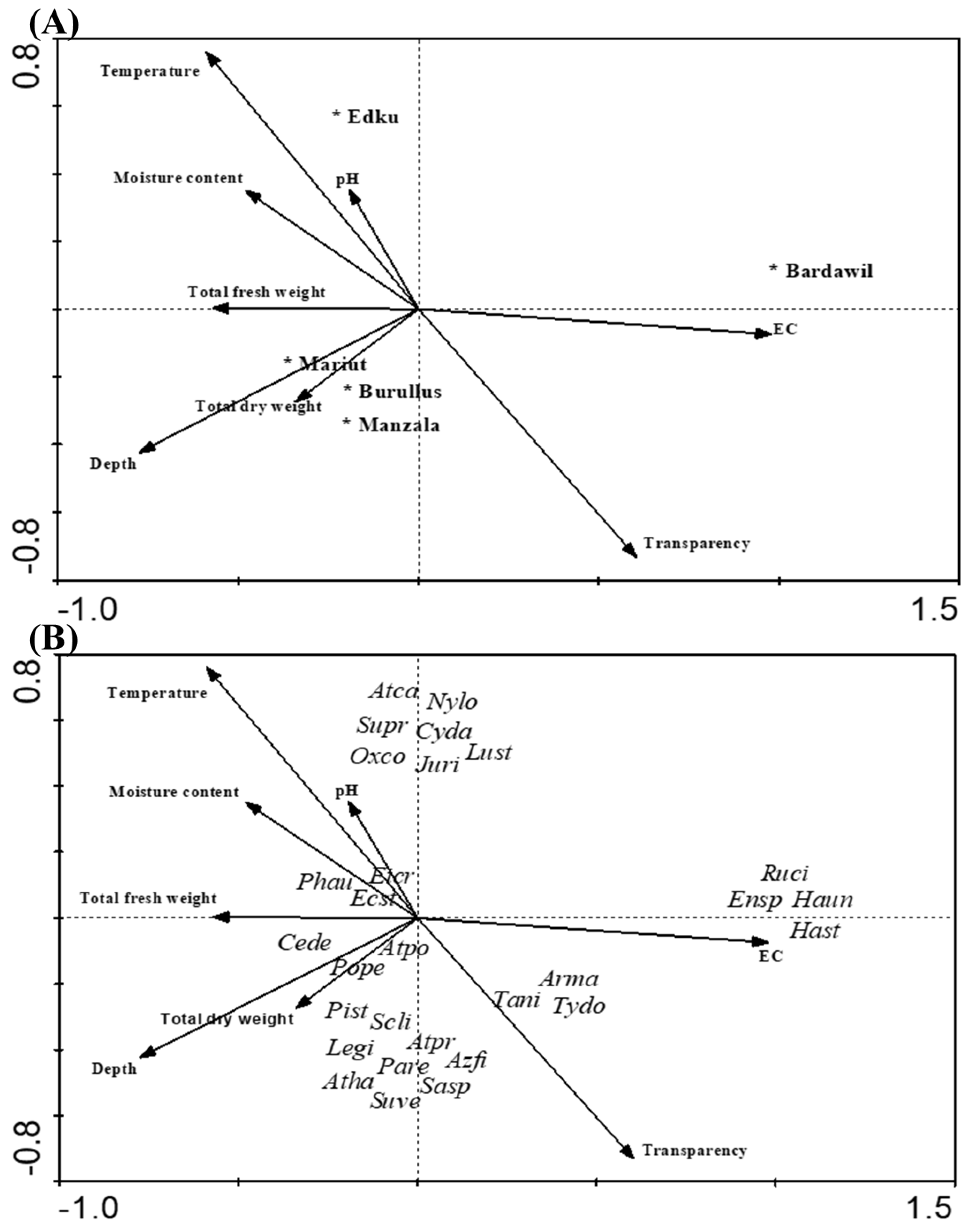
3.1. Vegetation Analyses
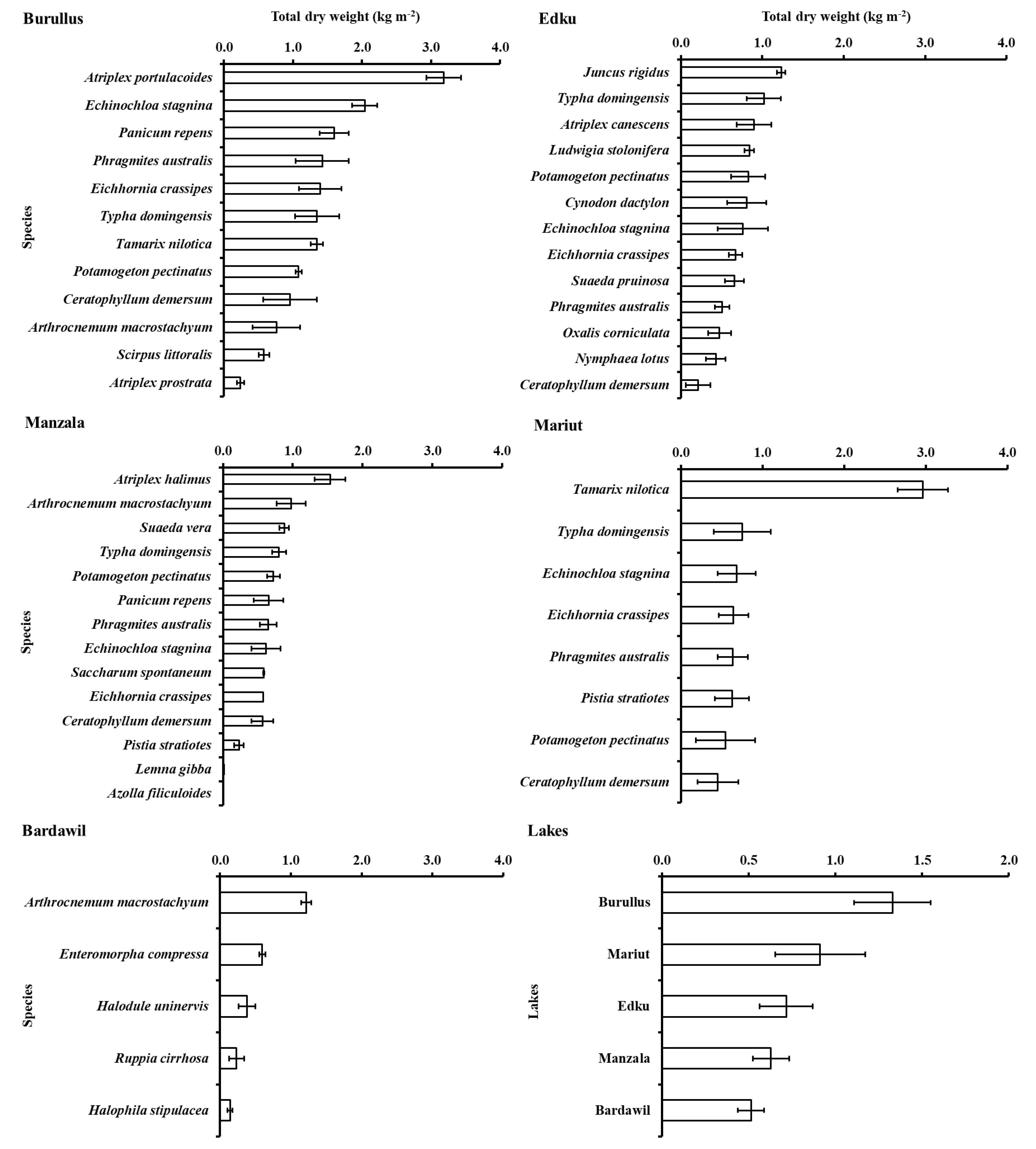
3.2. Aboveground Biomass
3.3. Macro and Micronutrient Analyses
3.3.1. Sediment Analyses
3.3.2. Water Analyses
3.3.3. Plant Material Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohamedein, L.; El-Sawy, M.; Bek, M. Sediment Contaminants in Northern Egyptian Coastal Lakes. In Egyptian Coastal Lakes and Wetlands: Part I; Negm, A.M., Bek, M.A., Abdel-Fattah, S., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 63–81. [Google Scholar]
- Shaltout, K.; Khalil, M. Lake Burullus: Burullus Protected Area; Publication of National Biodiversity Unit, Egyptian Environmental Affairs Agency (EEAA): Cairo, Egypt, 2005. [Google Scholar]
- Khalil, M.; Shaltout, K. Lake Bardawil and Zaranik Protected Area; Egyptian Environmental Affairs Agency: Cairo, Egypt, 2006. [Google Scholar]
- Maiorca, G.; Spampinato, G.; Caprio, A. Flora and vegetation of the la Vota coastal lakes (CW Calabria). Plant Sociol 2002, 39, 81–108. [Google Scholar]
- Jannuzzi, F.; Giudici, G.N.M.; Patrizio, S.; Massamormile, F.P. The Coastal Lakes of Campi Flegrei: Between Biodiversity and Anthropization; Firenze University Press: Firenze, Italy, 2020. [Google Scholar]
- Sebastiá-Frasquet, M.-T.; Aguilar-Maldonado, J.A.; Santamaría-del-Ángel, E.; Altur-Grau, V. Turbidity patterns in the Albufera lake, Spain, and their relation to irrigation cycles. WIT Trans. Ecol. Environ. (Online) 2019, 239, 173–180. [Google Scholar]
- Younis, A.M. Environmental impacts on Egyptian Delta Lakes’ biodiversity: A case study on Lake Burullus. In Egyptian Coastal Lakes and Wetlands: Part II; Springer International Publishing: Cham, Switzerland, 2018; pp. 107–128. [Google Scholar]
- Khalil, M.T. Physical and chemical properties of Egypt’s Coastal Wetlands; Burullus Wetland as a case study. In Egyptian Coastal Lakes and Wetlands: Part I; Springer International Publishing: Cham, Switzerland, 2018; pp. 83–101. [Google Scholar]
- Keshta, A.E.; Shaltout, K.H.; Baldwin, A.H.; El-Din, A.A.S. Sediment clays are trapping heavy metals in urban lakes: An indicator for severe industrial and agricultural influence on coastal wetlands at the Mediterranean coast of Egypt. Mar. Pollut. Bull. 2020, 151, 110816. [Google Scholar] [CrossRef]
- El Kafrawy, S.B.; Bek, M.; Negm, A.M. An overview of the Egyptian northern coastal lakes. In Egyptian Coastal Lakes and Wetlands: Part I; Negm, A.M., Bek, M.A., Abdel-Fattah, S., Eds.; Springer International Publishing: Cham, Switzerland, 2019; Volume Part I, pp. 3–17. [Google Scholar]
- El-Shinnawy, I. Al-Burullus Wetland’s Hydrological Study; MedWetCoast, Global Environmental Facility (GEF) and Egyptian Environmental Affairs Agency (EEAA): Cairo, Egypt, 2002.
- Fredston-Hermann, A.; Brown, C.J.; Albert, S.; Klein, C.J.; Mangubhai, S.; Nelson, J.L.; Teneve, L.; Wenger, A.; Gaines, S.D.; Halpern, B.S. Where Does River Runoff Matter for Coastal Marine Conservation? Front. Mar. Sci. 2016, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A global map of human impact on marine ecosystems. Science 2008, 319, 948–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halpern, B.S.; Frazier, M.; Potapenko, J.; Casey, K.S.; Koenig, K.; Longo, C.; Lowndes, J.S.; Rockwood, R.C.; Selig, E.R.; Selkoe, K.A.; et al. Spatial and temporal changes in cumulative human impacts on the world’s ocean. Nat. Commun. 2015, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Alongi, D.M. Coastal Ecosystem Processes; CRC Press: Boca Raton, FI, USA, 2020; p. 448. [Google Scholar]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Pollut. Bull. 2005, 50, 125–146. [Google Scholar] [CrossRef]
- Lapointe, B.E.; Barile, P.J.; Yentsch, C.S.; Littler, M.M.; Littler, D.S.; Kakuk, B. The relative importance of nutrient enrichment and herbivory on macroalgal communities near Norman’s Pond Cay, Exumas Cays, Bahamas: A “natural” enrichment experiment. J. Exp. Mar. Biol. Ecol. 2004, 298, 275–301. [Google Scholar] [CrossRef]
- Baldwin, A.H. Nitrogen and Phosphorus Differentially Affect Annual and Perennial Plants in Tidal Freshwater and Oligohaline Wetlands. Estuaries Coasts 2013, 36, 547–558. [Google Scholar] [CrossRef]
- Alongi, D.M. Macro- and Micronutrient Cycling and Crucial Linkages to Geochemical Processes in Mangrove Ecosystems. J. Mar. Sci. Eng. 2021, 9, 456. [Google Scholar] [CrossRef]
- Bedford, B.L.; Walbridge, M.R.; Aldous, A. Patterns in nutrient availability and plant diversity of temperate North American wetlands. Ecology 1999, 80, 2151–2169. [Google Scholar] [CrossRef]
- Zhang, D.J.; Qi, Q.; Tong, S.Z.; Wang, X.H.; An, Y.; Zhang, M.Y.; Lu, X.G. Soil Degradation Effects on Plant Diversity and Nutrient in Tussock Meadow Wetlands. J. Soil Sci. Plant Nutr. 2019, 19, 535–544. [Google Scholar] [CrossRef]
- Mylliemngap, W.; Barik, S.K. Plant diversity, net primary productivity and soil nutrient contents of a humid subtropical grassland remained low even after 50 years of post-disturbance recovery from coal mining. Environ. Monit. Assess 2019, 191, 17. [Google Scholar] [CrossRef]
- Seabloom, E.W.; Adler, P.B.; Alberti, J.; Biederman, L.; Buckley, Y.M.; Cadotte, M.W.; Collins, S.L.; Dee, L.; Fay, P.A.; Firn, J.; et al. Increasing effects of chronic nutrient enrichment on plant diversity loss and ecosystem productivity over time. Ecology 2021, 102, 11. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, S.; Ahmed, W.; Alatalo, J.M.; Mahmood, M.; Imtiaz, M.; Ditta, A.; Ali, E.F.; Abdelrahman, H.; Slaný, M.; Antoniadis, V.; et al. Herbal plants- and rice straw-derived biochars reduced metal mobilization in fishpond sediments and improved their potential as fertilizers. Sci. Total Environ. 2022, 826, 154043. [Google Scholar] [CrossRef]
- Cano-Ortiz, A.; Musarella, C.M.; Fuentes, J.C.P.; Gomes, C.J.P.; Quinto-Canas, R.; del Rio, S.; Cano, E. Indicative Value of the Dominant Plant Species for a Rapid Evaluation of the Nutritional Value of Soils. Agron. -Basel 2021, 11, 17. [Google Scholar] [CrossRef]
- Calabrese, G.; Perrino, E.V.; Ladisa, G.; Aly, A.; Tesfmichael Solomon, M.; Mazdaric, S.; Benedetti, A.; Ceglie, F.G. Short-term effects of different soil management practices on biodiversity and soil quality of Mediterranean ancient olive orchards. Org. Agric. 2015, 5, 209–223. [Google Scholar] [CrossRef]
- Shaltout, K.; Galal, T. Comparative study on the plant diversity of the Egyptian northern lakes. Egypt. J. Aquat. Res. 2006, 32, 254–270. [Google Scholar]
- Shaltout, K.H.; Galal, T.M.; El-Komi, T.M. Evaluation of the nutrient status of some hydrophytes in the water courses of Nile Delta, Egypt. Ecol. Mediterr. 2010, 36, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Galal, T.; Shaltout, K.; Hassan, L. The Egyptian Northeren Lakes Habitat Diversity, Vegetation and Economic Imoprtance; LAP LAMBERT Academic Publishing: Saarbrucken, Germany, 2012; p. 192. [Google Scholar]
- El-Bana, M.; Khedr, A.H.; Van Hecke, P.; Bogaert, J. Vegetation composition of a threatened hypersaline lake (Lake Bardawil), North Sinai. Plant Ecol. 2002, 163, 63–75. [Google Scholar] [CrossRef]
- Embabi, N.S.; Moawad, M.B. A semi-automated approach for mapping geomorphology of El Bardawil Lake, Northern Sinai, Egypt, using integrated remote sensing and GIS techniques. Egypt. J. Remote Sens. Space Sci. 2014, 17, 41–60. [Google Scholar] [CrossRef] [Green Version]
- Benninger, L.K.; Suayah, I.B.; Stanley, D.J. Manzala lagoon, Nile delta, Egypt: Modern sediment accumulation based on radioactive tracers. Environ. Geol. 1998, 34, 183–193. [Google Scholar] [CrossRef]
- Randazzo, G.; Stanley, D.J.; Di Geronimo, S.I.; Amore, C. Human-induced sedimentological changes in Manzala Lagoon, Nile Delta, Egypt. Environ. Geol. 1998, 36, 235–258. [Google Scholar] [CrossRef]
- Barakat, A.O.; Mostafa, A.; Wade, T.L.; Sweet, S.T.; El Sayed, N.B. Assessment of persistent organochlorine pollutants in sediments from Lake Manzala, Egypt. Mar. Pollut. Bull. 2012, 64, 1713–1720. [Google Scholar] [CrossRef]
- Eid, E.M.; Keshta, A.E.; Shaltout, K.H.; Baldwin, A.H.; El-Din, S.; Ahmed, A. Carbon sequestration potential of the five Mediterranean lakes of Egypt. Fundam. Appl. Limnol. 2017, 190, 87–96. [Google Scholar] [CrossRef]
- Boulos, L. Flora of Egypt Checklist; Al Hadara Publishing: Cairo, Egypt, 2009; Volume 1185494488. [Google Scholar]
- Raunkiaer, C. Plant Life Forms. Nature 1937, 140, 1035. [Google Scholar]
- Campbell, C.; Plank, C.O. Determination of total nitrogen in plant tissue by combustion. In Plant Analysis Reference Procedures for the Southern US Southern Coop; University of Georgia: Athens, GA, USA, 1992. [Google Scholar]
- Link, D.D.; Walter, P.J.; Kingston, H. Development and validation of the new EPA microwave-assisted leach method 3051A. Environ. Sci. Technol. 1998, 32, 3628–3632. [Google Scholar] [CrossRef]
- Kingston, H.M.; Haswell, S. Microwave-Enhanced Chemistry. Fundamentals, Sample Preparation and Applications; ACS Publications: Washington, DC, USA, 1997. [Google Scholar]
- Stefansson, A.; Gunnarsson, I.; Giroud, N. New methods for the direct determination of dissolved inorganic, organic and total carbon in natural waters by Reagent-Free (TM) Ion Chromatography and inductively coupled plasma atomic emission spectrometry. Anal. Chim. Acta 2007, 582, 69–74. [Google Scholar] [CrossRef]
- US-EPA. Methods for the Determination of Metals in Environmental Samples; US-EPA: Washington, DC, USA, 1994; p. 260.
- Allen, S.E. Chemical Analysis of Ecological Materials; Blackwell Scientific Publication: Oxford, UK, 1989. [Google Scholar]
- Hill, M.O. DECORANA-A FORTRAN Program for Detrended Correspondence Analysis and Reciprocal Averaging; Cornell University: Ithaca, NY, USA, 1979. [Google Scholar]
- Hill, M.O. TWINSPAN: A FORTRAN Program for Arranging Multivariate Data in an Ordered Two-Way Table by Classification of the Individuals and Attributes; Section of Ecology and Systematics; Cornell University: Ithaca, NY, USA, 1979; p. 90. [Google Scholar]
- Gauch, H.G.; Whittaker, R.H. Hierarchical-Classification of Community Data. J. Ecol. 1981, 69, 537–557. [Google Scholar] [CrossRef]
- Taylor, D. Vegetation Description and Analysis—A Practical Approach. Geogr. J. 1993, 159, 237. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing: Oxford, UK, 2004. [Google Scholar]
- Täckholm, V. Students’ Flora of Egypt, 2nd ed.; Cairo University Publishing: Beirut, Lebanon, 1974. [Google Scholar]
- Radford, E.A.; Catullo, G.; de Montmollin, B. Important Plant Areas of the South and East Mediterranean Region: Priority Sites for Conservation; International Union for Conservation of Nature: Gland, Switzerland, 2011. [Google Scholar]
- Shaltout, K.; Eid, E. Egypt. In Important Plant Areas of the South and East Mediterranean Region: Priority Sites for Conservation; Radford, E.A., Catullo, G., de Montmollin, B., Eds.; International Union for Conservation of Nature: Gland, Switzerland, 2011; pp. 40–43. [Google Scholar]
- Shaltout, K.H.; Al-Sodany, Y.M. Vegetation analysis of Burullus Wetland: A RAMSAR site in Egypt. Wetl. Ecol. Manag. 2008, 16, 421–439. [Google Scholar] [CrossRef]
- Abdel-Malik, S.; Khalil, M. Biological Diversity of Egypt: Aquatic Habitat Diversity; UNEP, Environmental Affairs Agency (EEAA): Cairo, Egypt, 1994; Volume 5. [Google Scholar]
- Fathi, A.A.; Abdelzahar, H.; El-naghy, M. Limnological studies on Edku Lake, Egypt. El-Minia Sci. Bull. 2000, 13, 44–60. [Google Scholar]
- Shakweer, L. Impacts of drainage water discharge on the water chemistry of Lake Edku. Egypt. J. Aquat. Res. 2006, 32, 264–282. [Google Scholar]
- Zhang, Y.Y.; Zhang, D.Y.; Barrett, S.C.H. Genetic uniformity characterizes the invasive spread of water hyacinth (Eichhornia crassipes), a clonal aquatic plant. Mol. Ecol. 2010, 19, 1774–1786. [Google Scholar] [CrossRef]
- Stohlgren, T.J.; Pyšek, P.; Kartesz, J.; Nishino, M.; Pauchard, A.; Winter, M.; Pino, J.; Richardson, D.M.; Wilson, J.; Murray, B.R.; et al. Globalization Effects on Common Plant Species. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 700–706. [Google Scholar]
- Harun, I.; Pushiri, H.; Amirul-Aiman, A.J.; Zulkeflee, Z. Invasive Water Hyacinth: Ecology, Impacts and Prospects for the Rural Economy. Plants-Basel 2021, 10, 23. [Google Scholar] [CrossRef]
- Wilson, J.R.; Holst, N.; Rees, M. Determinants and patterns of population growth in water hyacinth. Aquat. Bot. 2005, 81, 51–67. [Google Scholar] [CrossRef]
- Eid, E.M.; Shaltout, K.H. Population dynamics of Eichhornia crassipes (C. Mart.) Solms in the Nile Delta, Egypt. Plant Species Biol. 2017, 32, 279–291. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J.G. Wetlands; Wiley: Hoboken, NJ, USA, 2007. [Google Scholar]
- Newton, A.; Brito, A.C.; Icely, J.D.; Derolez, V.; Clara, I.; Angus, S.; Schernewski, G.; Inacio, M.; Lillebo, A.I.; Sousa, A.I.; et al. Assessing, quantifying and valuing the ecosystem services of coastal lagoons. J. Nat. Conserv. 2018, 44, 50–65. [Google Scholar] [CrossRef]
- Keshta, A.E.; Riter, J.C.A.; Shaltout, K.H.; Baldwin, A.H.; Kearney, M.; Sharaf El-Din, A.; Eid, E.M. Loss of Coastal Wetlands in Lake Burullus, Egypt: A GIS and Remote-Sensing Study. Sustainability 2022, 14, 4980. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) | ||||||||
| Burullus | Edku | Mariut | Manzala | |||||
| Edku | 0.48 | |||||||
| Mariut | 0.70 | 0.57 | ||||||
| Manzala | 0.62 | 0.44 | 0.64 | |||||
| Bardawil | 0.12 | 0.00 | 0.00 | 0.11 | ||||
| (B) | ||||||||
| Lake | Growth habit | Life forms | Total number of families | Total number of species | ||||
| Annuals | Perennials | Phanerophytes | Chamaephytes | Geophytes-helophytes | Hydrophytes | |||
| Burullus | 1 | 11 | 1 | 3 | 5 | 3 | 8 | 12 |
| Edku | 1 | 12 | 0 | 3 | 5 | 5 | 10 | 13 |
| Mariut | 0 | 8 | 1 | 0 | 3 | 4 | 7 | 8 |
| Manzala | 1 | 13 | 1 | 2 | 5 | 6 | 9 | 14 |
| Bardawil | 0 | 4 | 0 | 1 | 0 | 3 | 4 | 4 |
| Bardawil (n = 11) | Manzala (n = 11) | Burullus (n = 12) | Edku (n = 10) | Mariut (n = 10) | p Value | |
| Macronutrients in sediment (g kg−1) | ||||||
| P | 0.3 C ± 0.1 | 0.9 A ± 0.2 | 0.9 A ± 0.1 | 1.1 A ± 0.1 | 1.3 A ± 0.3 | 0.0023 |
| Ca | 34.4 C ± 9.8 | 109.1 B ± 25.7 | 94.3 B ± 17.6 | 68.9 CB ± 10.7 | 177.1 A ± 24.7 | <0.0001 |
| K | 2.5 A ± 0.6 | 2.6 A ± 0.5 | 3.3 A ± 0.4 | 4.0 A ± 0.4 | 3.4 A ± 0.5 | 0.2009 |
| Mg | 25.4 A ± 12.7 | 7.1 A ± 1.0 | 9.8 A ± 1.0 | 11.2 A ± 0.9 | 15.6 A ± 1.5 | 0.2117 |
| S | 4.9 A ± 1.8 | 8.0 A ± 1.4 | 8.1 A ± 1.1 | 5.9 A ± 0.8 | 8.2 A ± 1.2 | 0.2733 |
| Micronutrients in sediment (g kg−1) | ||||||
| Al | 9.7 C ± 2.6 | 15.5 BAC ± 2.9 | 18.9 BA ± 2.3 | 21.0 A ± 2.0 | 13.3 BC ± 2.5 | 0.0187 |
| Na | 9.6 A ± 2.1 | 3.0 B ± 0.3 | 4.7 B ± 0.6 | 3.8 B ± 0.4 | 4.5 B ± 0.7 | 0.0004 |
| Fe | 13.1 B ± 3.9 | 23.0 BA ± 4.0 | 29.8 A ± 3.7 | 35.0 A ± 3.2 | 24.4 BA ± 7.8 | 0.0262 |
| B | 0.10 A ± 0.05 | 0.02 B ± 0.002 | 0.02 B ± 0.002 | 0.02 B ± 0.002 | 0.03 B ± 0.01 | 0.0261 |
| Bardawil (n = 25) | Manzala (n = 39) | Burullus (n = 30) | Edku (n = 25) | Mariut (n = 19) | p value | |
| Macronutrients in water (mg L−1) | ||||||
| P | 0.03 B ± 0.005 | 0.2 B ± 0.04 | 0.2 B ± 0.08 | 0.2 B ± 0.02 | 0.7 A ± 0.18 | <0.0001 |
| N | 73.7 A ± 2.3 | 80.3 A ± 4.4 | 75.1 A ± 3.4 | 76.5 A ± 3.9 | 83.1 A ± 4.2 | 0.4988 |
| Mg | 1375.9 A ± 45.2 | 248.8 B ± 56.4 | 303.5 B ± 42.3 | 62.6 C ± 5.1 | 296.7 B ± 80.4 | <0.0001 |
| K | 779.7 A ± 19.9 | 96.0 B ± 16.8 | 136.4 B ± 13.4 | 40.8 C ± 3.5 | 101.6 B ± 20.9 | <0.0001 |
| Ca | 30.5 A ± 2.5 | 4.0 C ± 0.5 | 3.1 C ± 0.6 | 1.8 C ± 0.1 | 11.4 B ± 2.6 | <0.0001 |
| Micronutrients in water (mg L−1) | ||||||
| Na | 12,901.1 A ± 409.9 | 1656.2 B ± 377.3 | 2584.3 B ± 309.0 | 351.2 C ± 38.3 | 1752.6 B ± 447.3 | <0.0001 |
| Bardawil (n = 7) | Manzala (n = 21) | Burullus (n = 17) | Edku (n = 19) | Mariut (n = 15) | p value | |
| Macronutrients in plant material | ||||||
| C % | 23.5 B ± 1.8 | 33.1 A ± 1.0 | 35.2 A ± 0.8 | 32.6 A ± 1.1 | 34.0 A ± 1.3 | <0.0001 |
| N % | 1.3 C ± 0.2 | 2.2 A ± 0.2 | 1.7 BC ± 0.1 | 2.3 A ± 0.2 | 2.1 BA ± 0.2 | 0.0009 |
| CN ratio | 21.8 A ± 2.7 | 23.1 A ± 3.1 | 23.4 A ± 1.4 | 17.0 A ± 1.4 | 20.3 A ± 2.3 | 0.2197 |
| P (g kg−1) | 1.2 B ± 0.3 | 3.5 A ± 0.5 | 2.4 BA ± 0.4 | 3.1 A ± 0.4 | 2.9 A ± 0.5 | 0.0501 |
| K (g kg−1) | 18.5 A ± 3.9 | 21.9 A ± 4.0 | 16.1 A ± 1.4 | 20.1 A ± 3.2 | 24.4 A ± 4.7 | 0.5768 |
| Ca (g kg−1) | 44.3 A ± 9.6 | 18.3 A ± 4.5 | 26.9 A ± 9.0 | 34.7 A ± 13.3 | 19.8 A ± 5.0 | 0.4007 |
| Mg (g kg−1) | 11.3 A ± 1.7 | 5.5 B ± 0.6 | 6.1 B ± 0.7 | 6.9 B ± 1.2 | 5.3 B ± 0.8 | 0.0057 |
| S (g kg−1) | 7.4 BA ± 0.6 | 6.0 BA ± 0.6 | 5.3 B ± 0.8 | 5.0 B ± 0.3 | 8.5 A ± 1.3 | 0.0183 |
| Micronutrients in plant material (g kg−1) | ||||||
| Al | 4.7 A ± 1.5 | 2.7 B ± 0.6 | 1.6 B ± 0.5 | 2.6 B ± 0.5 | 1.0 B ± 0.3 | 0.0145 |
| Na | 36.1 A ± 11.1 | 21.3 A ± 5.6 | 26.0 A ± 7.5 | 16.4 A ± 5.7 | 16.9 A ± 2.2 | 0.38 |
| Fe | 7.7 A ± 3.0 | 3.7 B ± 0.8 | 3.4 B ± 1.1 | 3.5 B ± 0.8 | 1.4 B ± 0.4 | 0.0282 |
| B | 0.42 A ± 0.16 | 0.10 B ± 0.04 | 0.11 B ± 0.06 | 0.09 B ± 0.04 | 0.10 B ± 0.05 | 0.0151 |
| (A) Nutrients in sediment (g kg−1) | Al | B | Ca | Fe | K | Mg | Na | P |
| B | 0.1 ns | |||||||
| Ca | −0.1 ns | −0.1 ns | ||||||
| Fe | 0.8 *** | 0.0 ns | −0.1 ns | |||||
| K | 0.9 *** | 0.3 * | 0.0 ns | 0.7 *** | ||||
| Mg | 0.2 ns | 1.0 *** | 0.0 ns | 0.0 ns | 0.3 * | |||
| Na | 0.2 ns | 0.8 *** | −0.1 ns | 0.1 ns | 0.5 *** | 0.8 *** | ||
| P | 0.6 *** | −0.1 ns | 0.2 ns | 0.6 *** | 0.6 *** | 0.0 ns | 0.0 ns | |
| S | 0.6 *** | 0.3 * | 0.3 * | 0.6 *** | 0.6 *** | 0.4 ** | 0.4 ** | 0.5 *** |
| (B) Nutrients in water (mg L−1) | P | N | Mg | K | Na | |||
| N | 0.1 ns | |||||||
| Mg | −0.3 ** | 0.0 ns | ||||||
| K | −0.2 ** | −0.1 ns | 1.0 *** | |||||
| Na | −0.3 ** | 0.0 ns | 1.0 *** | 1.0 *** | ||||
| Ca | −0.2 * | −0.1 ns | 0.9 *** | 0.9 *** | 0.9 *** | |||
| (C) Nutrients in plant tissues (g kg−1) | C | N | P | K | Ca | Mg | S | |
| N | 0.1 ns | |||||||
| P | −0.2 * | 0.8 *** | ||||||
| K | −0.2 * | 0.5 *** | 0.7 *** | |||||
| Ca | −0.7 *** | −0.2 ns | 0.0 ns | −0.1 ns | ||||
| Mg | −0.8 *** | −0.2 ns | 0.0 ns | 0.0 ns | 0.6 *** | |||
| S | −0.2 * | 0.2 * | 0.1 ns | 0.0 ns | 0.2 ns | 0.3 ** | ||
| Na | −0.2 * | −0.3 ** | −0.3 ** | −0.1 ns | −0.1 ns | 0.4* ** | 0.1 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keshta, A.E.; Shaltout, K.H.; Baldwin, A.H.; Sharaf El-Din, A.; Eid, E.M. Variation in Plant Community Composition and Biomass to Macro and Micronutrients and Salinity across Egypt’s Five Major Coastal Lakes. Sustainability 2022, 14, 6180. https://doi.org/10.3390/su14106180
Keshta AE, Shaltout KH, Baldwin AH, Sharaf El-Din A, Eid EM. Variation in Plant Community Composition and Biomass to Macro and Micronutrients and Salinity across Egypt’s Five Major Coastal Lakes. Sustainability. 2022; 14(10):6180. https://doi.org/10.3390/su14106180
Chicago/Turabian StyleKeshta, Amr E., Kamal H. Shaltout, Andrew H. Baldwin, Ahmed Sharaf El-Din, and Ebrahem M. Eid. 2022. "Variation in Plant Community Composition and Biomass to Macro and Micronutrients and Salinity across Egypt’s Five Major Coastal Lakes" Sustainability 14, no. 10: 6180. https://doi.org/10.3390/su14106180
APA StyleKeshta, A. E., Shaltout, K. H., Baldwin, A. H., Sharaf El-Din, A., & Eid, E. M. (2022). Variation in Plant Community Composition and Biomass to Macro and Micronutrients and Salinity across Egypt’s Five Major Coastal Lakes. Sustainability, 14(10), 6180. https://doi.org/10.3390/su14106180