
Island Colonization and Environmental Sustainability in the Postglacial Mediterranean


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Sustainability and Island Settlement
1.2. Conceptualizing Sustainability over Archaeological Timescales
2. Sustainability and the Mediterranean
3. Materials and Methods
3.1. Methodological Assumptions
3.1.1. Area
3.1.2. Isolation
3.1.3. Productivity
3.1.4. Biodiversity
3.2. Methodological Considerations
3.3. Analysis
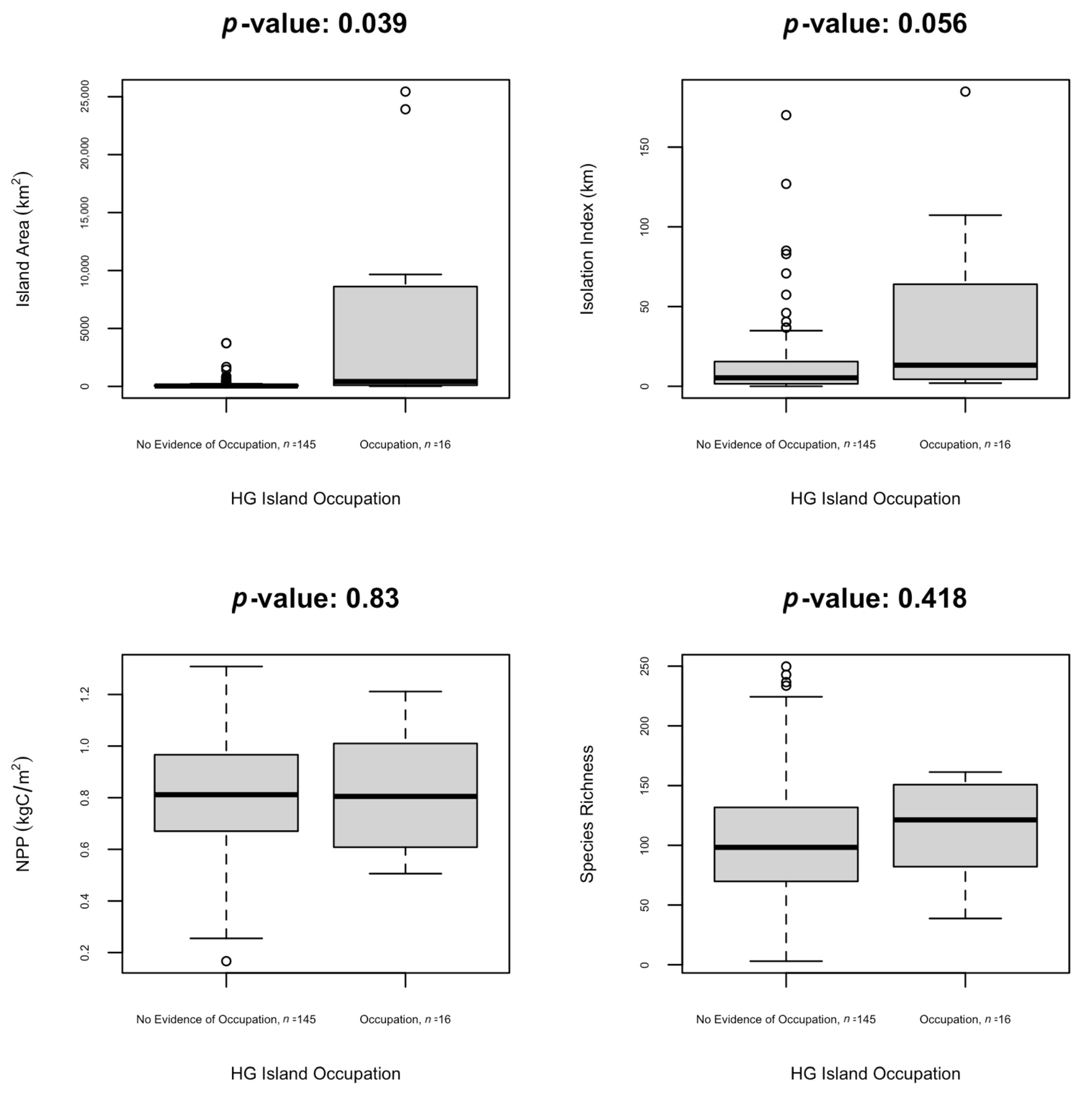
4. Results
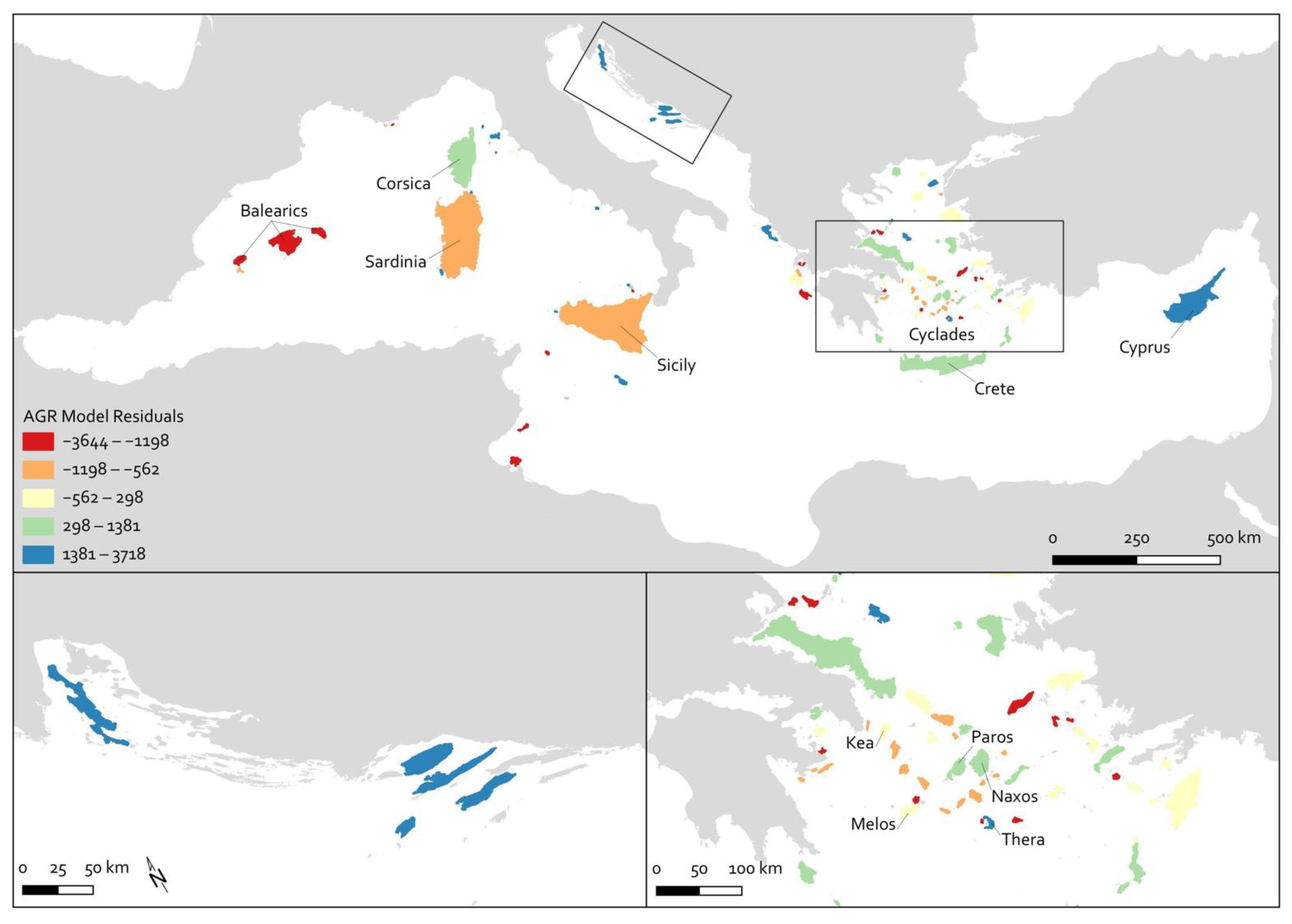
5. Discussion
5.1. Residuals in the Agropastoral Model
5.2. Area and Long-Term Sustainability
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erlandson, J.M.; Fitzpatrick, S.M. Oceans, islands, and coasts: Current perspectives on the role of the sea in human prehistory. J. Isl. Coast. Archaeol. 2005, 1, 5–32. [Google Scholar] [CrossRef]
- Fitzpatrick, S.M.; Anderson, A. Islands of isolation: Archaeology and the power of aquatic perimeters. J. Isl. Coast. Archaeol. 2008, 3, 4–16. [Google Scholar] [CrossRef]
- Keegan, W.F.; Diamond, J. Colonization of islands by humans: A biogeographical perspective. J. Archaeol. Method Theory 1987, 10, 49–92. [Google Scholar] [CrossRef]
- Leppard, T.P. The biophysical effects of Neolithic island colonization: General dynamics and sociocultural implications. Hum. Ecol. 2017, 45, 555–568. [Google Scholar] [CrossRef]
- Erlandson, J.M. The archaeology of aquatic adaptations: Paradigms for a new millennium. J. Archaeol. Res. 2001, 9, 287–350. [Google Scholar] [CrossRef]
- Rick, T.C.; Kirch, P.V.; Erlandson, J.M.; Fitzpatrick, S.M. Archeology, deep history, and the human transformation of island ecosystems. Anthropocene 2013, 4, 33–45. [Google Scholar] [CrossRef]
- Braje, T.J.; Leppard, T.P.; Fitzpatrick, S.M.; Erlandson, J.M. Archaeology, historical ecology and anthropogenic island ecosystems. Environ. Conserv. 2017, 44, 286–297. [Google Scholar] [CrossRef]
- Gaffney, D. Pleistocene water crossings and adaptive flexibility within the Homo genus. J. Archaeol. Res. 2020. [Google Scholar] [CrossRef]
- Vitousek, P.M. Oceanic islands as model systems for ecological studies. J. Biogeogr. 2002, 29, 573–582. [Google Scholar] [CrossRef]
- Fisher, C. Archaeology for sustainable agriculture. J. Archaeol. Res. 2020, 28, 393–441. [Google Scholar] [CrossRef]
- Boivin, N.; Zeder, M.; Fuller, D.; Crowther, A.; Larson, G.; Erlandson, J.; Denham, T.; Petraglia, M. Ecological consequences of human niche construction: Examining long-term anthropogenic shaping of global species distributions. Proc. Natl. Acad. Sci. USA 2016, 113, 6388–6396. [Google Scholar] [CrossRef] [PubMed]
- Braje, T.J. Evaluating the Anthropocene: Is there something useful about a geological epoch of humans? Antiquity 2016, 90, 504–518. [Google Scholar] [CrossRef]
- Athens, J.S. Rattus exulans and the catastrophic disappearance of Hawaiʻi’s native lowland forest. Biol. Invasions 2009, 11, 1489–1501. [Google Scholar] [CrossRef]
- Braje, T.J.; Rick, T.C.; Szpak, P.; Newsome, S.D.; McCain, J.M.; Elliott Smith, E.A.; Glassow, M.; Hamilton, S.L. Historical ecology and the conservation of large, hermaphroditic fishes in Pacific Coast kelp forest ecosystems. Sci. Adv. 2017, 3, e1601759. [Google Scholar] [CrossRef] [PubMed]
- Dugmore, A.J.; McGovern, T.H.; Vésteinsson, O.; Arneborg, J.; Streeter, R.; and Keller, C. Cultural adaptation, compounding vulnerabilities and conjunctures in Norse Greenland. Proc. Natl. Acad. Sci. USA 2012, 109, 3658–3663. [Google Scholar] [CrossRef] [PubMed]
- Kirch, P.V. Three islands and an archipelago: Reciprocal interactions between humans and island ecosystems in Polynesia. Earth Environ. Sci. Trans. R. Soc. Edinb. 2007, 98, 85–99. [Google Scholar] [CrossRef]
- Koch, A.; Brierley, C.; Maslin, M.M.; and Lewis, S.L. Earth System impacts of the European arrival and great dying in the Americas after 1492. Quat. Sci. Rev. 2019, 207, 13–36. [Google Scholar] [CrossRef]
- Marston, J.M. Modeling resilience and sustainability in ancient agricultural systems. J. Ethnobiol. 2015, 35, 585–605. [Google Scholar] [CrossRef]
- Curran, M.A. Wrapping our heads around sustainability. Sustainability 2009, 1, 5–13. [Google Scholar] [CrossRef]
- Kates, R.W.; Clark, W.C.; Corell, R.; Hall, J.M.; Jaeger, C.C.; Lowe, I.; McCarthy, J.J.; Schellnhuber, H.J.; Bolin, B.; Dickson, N.M.; et al. Sustainability science. Science 2001, 292, 641–642. [Google Scholar] [CrossRef] [PubMed]
- Stiner, M.C.; Munro, N.D.; Surovell, T.A. The tortoise and the hare: Small-game use, the broad-spectrum revolution, and Paleolithic demography. Curr. Anthropol. 2000, 41, 39–73. [Google Scholar] [CrossRef]
- Stiner, M.C.; Munro, N.D. On the evolution of diet and landscape during the Upper Paleolithic through Mesolithic at Franchthi Cave (Peloponnese, Greece). J. Hum. Evol. 2011, 60, 618–636. [Google Scholar] [CrossRef]
- Leppard, T.P. Process and dynamics of Mediterranean Neolithization (7000-5500 BC). J. Archaeol. Res. in press.
- Zeder, M.A. Domestication and early agriculture in the Mediterranean Basin: Origins, diffusion, and impact. Proc. Natl. Acad. Sci. USA 2008, 105, 11597–11604. [Google Scholar] [CrossRef] [PubMed]
- Rick, T.C.; Ontiveros, M.A.C.; Jerardino, A.; Mariotti, A.; Méndez, C.; Williams, A.N. Human-environmental interactions in Mediterranean climate regions from the Pleistocene to the Anthropocene. Anthropocene 2020. [Google Scholar] [CrossRef]
- Ramis, D.; Alcover, J.A. Revisiting the earliest human presence in Mallorca, western Mediterranean. Proc. Prehist. Soc. 2001, 67, 261–269. [Google Scholar] [CrossRef]
- Cherry, J.F.; Leppard, T.P. The Balearic Paradox: Why were the islands colonized so late? Pyrenae 2018, 49, 49–70. [Google Scholar]
- Dawson, H. Mediterranean Voyages: The Archaeology of Island Colonisation and Abandonment; Left Coast Press: Walnut Creek, CA, USA, 2014. [Google Scholar]
- Gjesfjeld, E.; Etnier, M.A.; Takase, K.; Brown, W.A.; Fitzhugh, B. Biogeography and adaptation in the Kuril Islands, Northeast Asia. World Arch. 2019, 51, 429–453. [Google Scholar] [CrossRef]
- Cherry, J.F. Pattern and process in the earliest colonization of the Mediterranean islands. Proc. Prehist. Soc. 1981, 47, 41–68. [Google Scholar] [CrossRef]
- Cherry, J.F. The initial colonisation of the West Mediterranean islands in the light of island biogeography and palaeogeography. In The Deya Conference of Prehistory: Early Settlement in the Western Mediterranean Islands and the Peripheral Areas; Waldren, W.H., Chapman, R., Lewthwaite, J., Kennard, R.C., Eds.; BAR International Series 229; British Archaeological Reports: Oxford, UK, 1984; pp. 7–22. [Google Scholar]
- Cherry, J.F. The first colonization of the Mediterranean islands: A review of recent research. J. Mediterr. Archaeol. 1990, 3, 145–222. [Google Scholar] [CrossRef]
- Cherry, J.F.; Leppard, T.P. Patterning and its causation in the pre-Neolithic colonization of the Mediterranean islands (Late Pleistocene to Early Holocene). J. Isl. Coast. Archaeol. 2018, 13, 191–205. [Google Scholar] [CrossRef]
- Renfrew, C.; Aspinall, A. Aegean obsidian and Franchthi Cave. In Excavations at Franchthi Cave, Greece (Fascicle 5); Jacobsen, T.W., Ed.; Indiana University Press: Bloomington, IN, USA, 1990; pp. 257–279. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. 2020. Available online: https://www.R-project.org/ (accessed on 17 March 2021).
- Global Administrative Areas. GADM Database of Global Administrative Areas, Version 2.8. 2020. Available online: www.gadm.org (accessed on 17 March 2021).
- Knodell, A.; Athanasoulis, D.; Tankosić, Ž.; Cherry, J.F.; Garonis, T.J.; Levine, E.; Nenova, D.; Öztürk, H. An island archaeology of uninhabited landscapes: Off-shore islets near Paros and Antiparos, Greece (The Small Cycladic Islands Project). J. Isl. Coast. Archaeol. 2020, 15. [Google Scholar] [CrossRef]
- Leppard, T.P. Adaptive responses to demographic fragility: Mitigating stochastic effects in early island colonization. Hum. Ecol. 2015, 43, 721–734. [Google Scholar] [CrossRef]
- Brose, U.; Ostling, A.; Harrison, K.; and Martinez, N.D. Unified spatial scaling of species and their trophic interactions. Nature 2004, 428, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Galiana, N.; Lurgi, M.; Claramunt-López, B.; Fortin, M.-J.; Leroux, S.; Cazelles, K.; Gravel, D.; Montoya, J.M. The spatial scaling of species interaction networks. Nat. Ecol. Evol. 2018, 2, 782–790. [Google Scholar] [CrossRef]
- Holt, R.D.; Lawton, J.H.; Polis, G.A.; Martinez, N.D. Trophic rank and the species-area relationship. Ecology 1999, 80, 1495–1504. [Google Scholar] [CrossRef]
- Roslin, T.; Várkonyi, G.; Koponen, M.; Vikberg, V.; Nieminen, M. Species-area relationships across four trophic levels: Decreasing island size truncates food chains. Ecography 2014, 37, 443–453. [Google Scholar] [CrossRef]
- Dahl, A. Island Directory; United Nations Environment Program: Nairobi, Kenya, 1991. [Google Scholar]
- Freeman, J.; Robinson, E.; Beckman, N.G.; Bird, D.; Baggio, J.A.; Anderies, J.M. The global ecology of human population density and interpreting changes in paleo-population density. J. Archaeol. Sci. 2020, 120, 105168. [Google Scholar] [CrossRef]
- Tallavaara, M.; Eronen, J.T.; Luoto, M. Productivity, biodiversity, and pathogens influence the global hunter-gatherer population density. Proc. Natl. Acad. Sci. USA 2018, 115, 1232–1237. [Google Scholar] [CrossRef] [PubMed]
- Running, S.; Zhao, M. MOD17A3HGF MODIS/Terra Net Primary Production Gap-Filled Yearly L4 Global 500m SIN Grid V006. NASA EOSDIS Land Processes DAAC. Available online: https://doi.org/10.5067/MODIS/MOD17A3HGF.006 (accessed on 25 September 2020).
- Eisenhauer, N.; Schielzeth, H.; Barnes, A.D.; Barry, K.; Bonn, A.; Brose, U.; Bruelheide, H.; Buchmann, N.; Buscot, F.; Ebeling, A. A multitrophic perspective on biodiversity-ecosystem functioning research. Adv. Ecol. Res. 2019, 61, 1–54. [Google Scholar]
- Jenkins, C.N.; Pimm, S.L.; Joppa, L.N. Global Patterns of Terrestrial Vertebrate Diversity and Conservation. Proc. Natl. Acad. Sci. USA 2013, 110, E2602–E2610. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.W.; Bettinger, R.L.; Codding, B.F.; Jones, T.L.; Schwitalla, A.W. Resource Scarcity Drives Lethal Aggression among Prehistoric Hunter-Gatherers in Central California. Proc. Natl. Acad. Sci. USA 2016, 113, 12120–12125. [Google Scholar] [CrossRef]
- Alcover, J.A.; Sans, A.; Palmer, M. The extent of extinctions of mammals on islands. J. Biogeogr. 1998, 25, 913–918. [Google Scholar] [CrossRef]
- Fattorini, S. Detecting biodiversity hotspots by Species-Area Relationships: A case study of Mediterranean beetles. Conserv. Biol. 2006, 20, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Gentile, G.; Argano, R. Island biogeography of the Mediterranean sea: The species–area relationship for terrestrial isopods. J. Biogeogr. 2005, 32, 1715–1726. [Google Scholar] [CrossRef]
- Di Maida, G. The earliest human occupation of Sicily: A review. J. Isl. Coast. Archaeol. 2020. [Google Scholar] [CrossRef]
- Leppard, T.P.; Runnels, C. Maritime hominin dispersals in the Pleistocene: Advancing the debate. Antiquity 2017, 91, 510–519. [Google Scholar] [CrossRef]
- Katsianis, M.; Bevan, A.; Styliaras, G.; Maniatis, Y. An Aegean history and archaeology written through radiocarbon dates. J. Open Archaeol. Data 2020, 8, 5. [Google Scholar] [CrossRef]
- Douka, K.; Efstratiou, N.; Hald, M.M.; Henriksen, P.S.; Karetsou, A. Dating Knossos and the arrival of the earliest Neolithic in the southern Aegean. Antiquity 2017, 91, 304–321. [Google Scholar] [CrossRef]
- Binder, D.; Lanos, P.; Angeli, L.; Gomart, L.; Guilaine, J.; Manen, C.; Maggi, R.; Muntoni, I.; Panelli, C.; Radi, G.; et al. Modelling the earliest north-western dispersal of Mediterranean Impressed Wares: New dates and Bayesian chronological model. Doc. Praehist. 2017, 44, 54–77. [Google Scholar] [CrossRef]
- Manen, C.; Perrin, T.; Guilaine, J.; Bouby, L.; Bréhard, S.; Briois, F.; Durand, F.; Marinval, P.; Vigne, J.-D. The Neolithic transition in the western Mediterranean: A complex and non-linear diffusion process—The radiocarbon record revisited. Radiocarbon 2019, 61, 531–571. [Google Scholar] [CrossRef]
- Kačar, S. Evidence of absence or absence of evidence? Searching for Late Mesolithic (Castelnovian) hunter-gatherers in the eastern Adriatic. J. Mediterr. Archaeol. 2020, 33. [Google Scholar] [CrossRef]
- van Andel, J.H.; Shackleton, J.C. Late Paleolithic and Mesolithic Coastlines of Greece and the Aegean. J. Field Archaeol. 1982, 9, 445–454. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Inference: A Practical Information-Theoretic Approach; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Capelli, C.; Starnini, E.; Cabella, R.; Piazza, M. The circulation of Early Neolithic pottery in the Mediterranean: A synthesis of new archaeometric data from the Impressed Ware culture of Liguria (north-west Italy). J. Archaeol. Sci. Rep. 2017, 16, 532–541. [Google Scholar] [CrossRef]
- Gabriele, M.; Convertini, F.; Verati, C.; Gratuze, B.; Jacomet, S.; Boschian, G.; Durrenmath, G.; Guilaine, J.; Lardeaux, J.-M.; Gomart, L.; et al. Long-distance mobility in the North-Western Mediterranean during the Neolithic transition using high resolution pottery sourcing. J. Archaeol. Sci. Rep. 2019, 28, 102050. [Google Scholar] [CrossRef]
- Broodbank, C.; Lucarini, G. The dynamics of Mediterranean Africa, ca. 9600–1000 bc: An interpretative synthesis of knowns and unknowns. J. Mediterr. Archaeol. 2019, 32, 195–267. [Google Scholar] [CrossRef]
- Keegan, W.F.; Fitzpatrick, S.M.; Sealey, K.S.; LeFebvre, M.L.; Sinelli, P.T. The role of small islands in marine subsistence strategies: Case studies from the Caribbean. Hum. Ecol. 2008, 36, 635–654. [Google Scholar] [CrossRef]
- Kirch, P.V.; Swift, J.A. New AMS radiocarbon dates and a re-evaluation of the cultural sequence of Tikopia Island, southeast Solomon Islands. J. Polyn. Soc. 2017, 126, 313–336. [Google Scholar] [CrossRef]
- Bocquet-Appel, J.-P. Paleoanthropological traces of a Neolithic Demographic Transition. Curr. Anthropol. 2002, 43, 637–650. [Google Scholar] [CrossRef]
- Mathias, R.A.; Bickel, C.A.; Beaty, T.H.; Petersen, G.M.; Hetmanski, J.B.; Liang, K.-Y.; Barnes, K.C. A study of contemporary levels and temporal trends in inbreeding in the Tangier Island, Virginia, population using pedigree data and isonymy. Am. J. Phys. Anthropol. 2000, 112, 29–38. [Google Scholar] [CrossRef]
- Soodyall, H.; Nebel, A.; Morar, B.; Jenkins, T. Genealogy and genes: Tracing the founding fathers of Tristan da Cunha. Eur. J. Hum. Genet. 2003, 11, 705–709. [Google Scholar] [CrossRef]
- Chiang, C.W.K.; Marcus, J.H.; Sidore, C.; Biddanda, A.; Al-Asadi, H.; Zoledziewska, M.; Pitzalis, M.; Busonero, F.; Maschio, A.; Pistis, G.; et al. Genomic history of the Sardinian population. Nat. Genet. 2018, 50, 1426–1434. [Google Scholar] [CrossRef]
- Marcus, J.H.; Posth, C.; Ringbauer, H.; Lai, L.; Skeates, R.; Sidore, C.; Beckett, J.; Furtwängler, A.; Olivieri, A.; Chiang, C.W.K.; et al. Genetic history from the Middle Neolithic to present on the Mediterranean island of Sardinia. Nat. Commun. 2020, 11, 939. [Google Scholar] [CrossRef]
- Allee, W.C. Animal Aggregations: A Study in General Sociology; University of Chicago Press: Chicago, IL, USA, 1931. [Google Scholar]
- Giovas, C.M.; Fitzpatrick, S.M. Prehistoric migration in the Caribbean: Past perspectives, new models and the ideal free distribution of West Indian colonization. World Archaeol. 2014, 46, 569–589. [Google Scholar] [CrossRef]
- Hanna, J.A.; Giovas, C.M. An islandscape IFD: Using the Ideal Free Distribution to predict Pre-Columbian settlements from Grenada to St. Vincent, Eastern Caribbean. Environ. Archaeol. 2019. [Google Scholar] [CrossRef]
- Kennett, D.J.; Anderson, A.; Winterhalder, B. The Ideal Free Distribution, food production, and the colonization of Oceania. In Behavioral Ecology and the Transition to Agriculture; Kennett, D.J., Winterhalder, B., Eds.; University of California Press: Berkeley, CA, USA, 2006; pp. 265–288. [Google Scholar]
- Shennan, S.; Downey, S.D.; Timpson, A.; Edinborough, K.; Colledge, S.; Kerig, T.; Manning, K.; Thomas, M.G. Regional population collapse followed initial agriculture booms in mid-Holocene Europe. Nat. Commun. 2013, 4, 2486. [Google Scholar] [CrossRef]
- Adloff, F.; Somot, S.; Sevault, F.; Jordà, G.; Aznar, R.; Déqué, M.; Herrmann, M.; Marcos, M.; Dubois, C.; Padorno, E.; et al. Mediterranean Sea response to climate change in an ensemble of twenty first century scenarios. Clim. Dyn. 2015, 45, 2775–2802. [Google Scholar] [CrossRef]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.-P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate change and interconnected risks to sustainable development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980. [Google Scholar] [CrossRef]
- Lionello, P.; Scarascia, L. The relation between climate change in the Mediterranean region and global warming. Reg. Environ. Chang. 2018, 18, 1481–1493. [Google Scholar] [CrossRef]
- Bennett, E.M.; Cramer, W.; Begossi, A.; Cundill, G.; Díaz, S.; Egoh, B.N.; Geijzendorffer, I.R.; Krug, C.B.; Lavorel, S.; Lazos, E.; et al. Linking biodiversity, ecosystem services, and human well-being: Three challenges for designing research for sustainability. Curr. Opin. Environ. Sustain. 2015, 14, 76–85. [Google Scholar] [CrossRef]
- Sandifer, P.A.; Sutton-Grier, A.E.; Ward, B.P. Exploring connections among nature, biodiversity, ecosystem services, and human health and well-being: Opportunities to enhance health and biodiversity conservation. Ecosyst. Serv. 2015, 12, 1–15. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plekhov, D.; Leppard, T.P.; Cherry, J.F. Island Colonization and Environmental Sustainability in the Postglacial Mediterranean. Sustainability 2021, 13, 3383. https://doi.org/10.3390/su13063383
Plekhov D, Leppard TP, Cherry JF. Island Colonization and Environmental Sustainability in the Postglacial Mediterranean. Sustainability. 2021; 13(6):3383. https://doi.org/10.3390/su13063383
Chicago/Turabian StylePlekhov, Daniel, Thomas P. Leppard, and John F. Cherry. 2021. "Island Colonization and Environmental Sustainability in the Postglacial Mediterranean" Sustainability 13, no. 6: 3383. https://doi.org/10.3390/su13063383
APA StylePlekhov, D., Leppard, T. P., & Cherry, J. F. (2021). Island Colonization and Environmental Sustainability in the Postglacial Mediterranean. Sustainability, 13(6), 3383. https://doi.org/10.3390/su13063383