
Effects of Heat Stress on Growth, Physiology of Plants, Yield and Grain Quality of Different Spring Wheat (Triticum aestivum L.) Genotypes

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Material, Design, and Site
2.2. Data Acquisition
2.3. Morpho-Phonological Traits
2.4. Physiological and Biochemical Traits of Plants
2.5. Statistical Analysis
3. Results
3.1. Analysis of Variance (ANOVA)
3.2. General Combining Ability (GCA)
3.3. Specific Combining Ability (SCA)
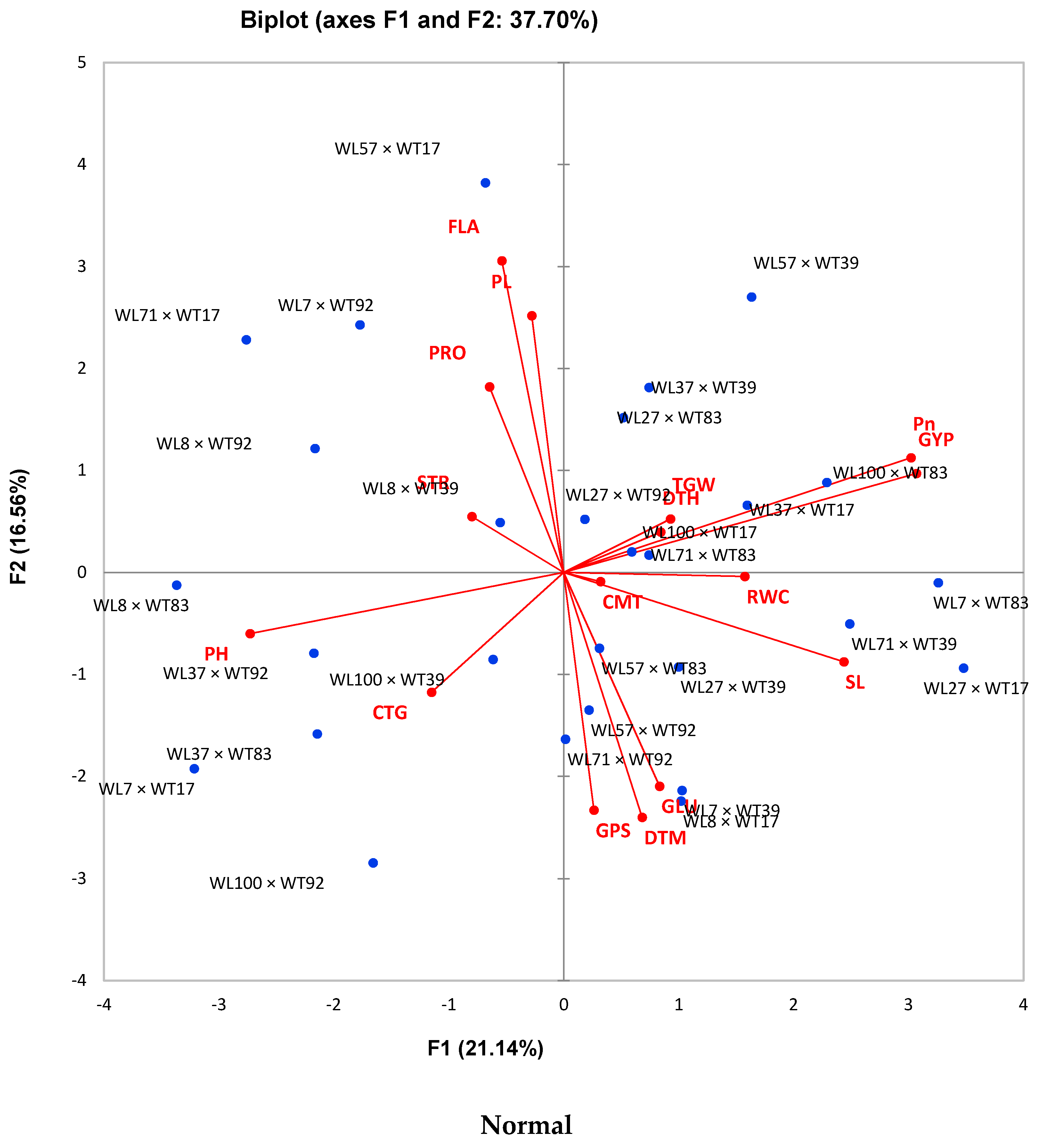
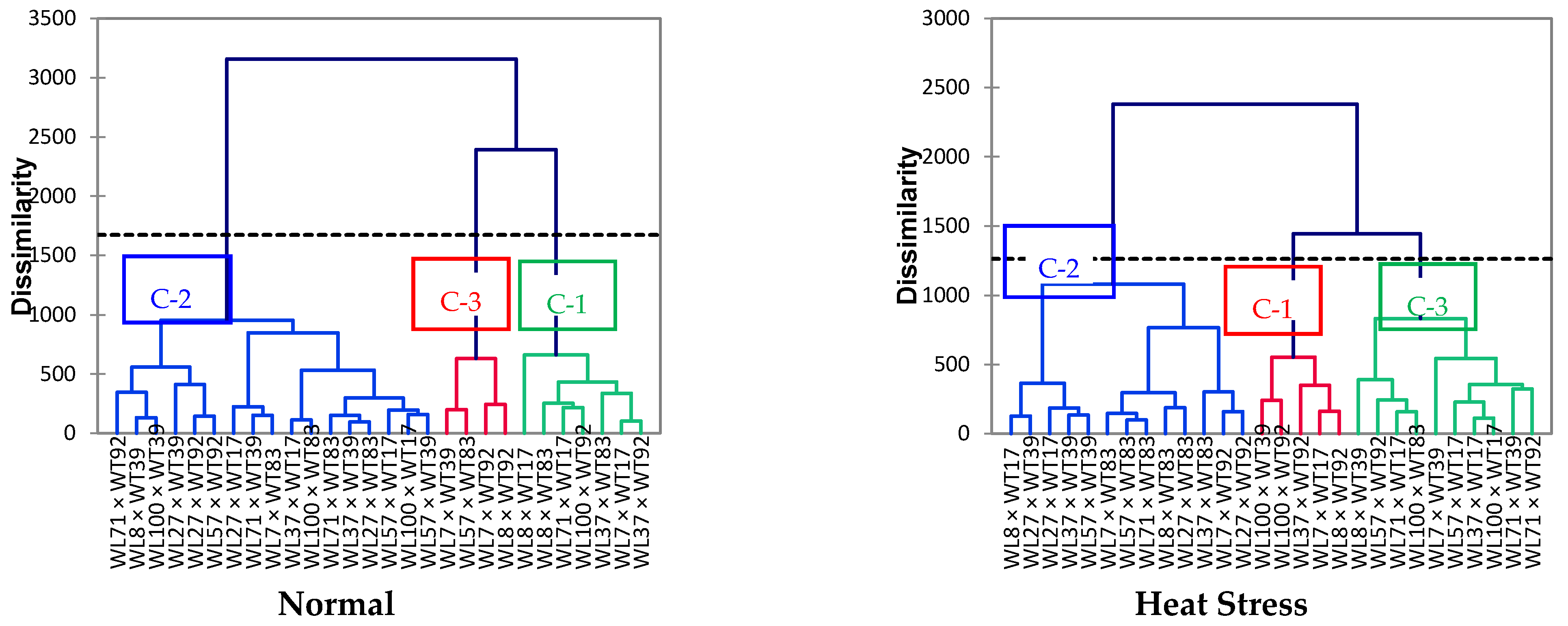
3.4. Correlation, Profile Plot, and Biplot Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef]
- Braun, H.J.; Atlin, G.; Payne, T.; Reynolds, M.P. Climate Change and Crop Production; CABI: Wallingford, UK, 2010; pp. 115–138. [Google Scholar]
- FAO Food and Agriculture Organization of United Nations 2020. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 10 January 2021).
- Sharma, I.; Tyagi, B.S.; Singh, G.; Venkatesh, K.; Gupta, O.P. Enhancing wheat production—A global perspective. Indian J. Agric. Sci. 2015, 85, 3–13. [Google Scholar]
- Poudel, P.B.; Poudel, M.R. Heat stress effects and Tolerance in wheat: A review. J. Biol. Today’s World 2020, 379, 217. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Narayanan, S.; Erdayani, E.; Prasad, P.V.V. Effects of high temperature stress during anthesis and grain filling periods on photosynthesis, lipids and grain yield in wheat. BMC Plant Biol. 2020, 20, 268. [Google Scholar] [CrossRef] [PubMed]
- Akter, N.; Rafiqul Islam, M. Heat stress effects and management in wheat. A review. Agron. Sustain. Dev. 2017, 37, 1–7. [Google Scholar] [CrossRef]
- Talukder, A.S.M.H.M.; McDonald, G.K.; Gill, G.S. Effect of short-term heat stress prior to flowering and early grain set on the grain yield of wheat. Field Crop. Res. 2014, 160, 54–63. [Google Scholar] [CrossRef]
- Kaushal, N.; Bhandari, K.; Siddique, K.H.M.; Nayyar, H. Food crops face rising temperatures: An overview of responses, adaptive mechanisms, and approaches to improve heat tolerance. Cogent Food Agric. 2016, 2. [Google Scholar] [CrossRef]
- Balla, K.; Karsai, I.; Bónis, P.; Kiss, T.; Berki, Z.; Horváth, Á.; Mayer, M.; Bencze, S.; Veisz, O. Heat stress responses in a large set of winter wheat cultivars (Triticum aestivum L.) depend on the timing and duration of stress. PLoS ONE 2019, 14, e0222639. [Google Scholar] [CrossRef]
- Rahman, M.A.; Chikushi, J.; Yoshida, S.; Karim, A.J.M.S. Growth and Yield components of Wheat genotypes exposed to High temperature stress under control environmrnt. Bangladesh J. Agric. Res. 2009, 34, 361–372. [Google Scholar]
- Vignjevic, M.; Wang, X.; Olesen, J.E.; Wollenweber, B. Traits in Spring Wheat Cultivars Associated with Yield Loss Caused by a Heat Stress Episode after Anthesis. J. Agron. Crop Sci. 2014. [Google Scholar] [CrossRef]
- Shah, N.H.; Paulsen, G.M. Interaction of drought and high temperature on photosynthesis and grain-filling of wheat. Plant Soil 2003, 257, 219–226. [Google Scholar] [CrossRef]
- Asseng, S.; Ewert, F.; Martre, P.; Rötter, R.P.; Lobell, D.B.; Cammarano, D.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W.; et al. Rising temperatures reduce global wheat production. Nat. Clim. Chang. 2015, 5, 143–147. [Google Scholar] [CrossRef]
- Lobell, D.B.; Burke, M.B.; Tebaldi, C.; Mastrandrea, M.D.; Falcon, W.P.; Naylor, R.L. Prioritizing climate change adaptation needs for food security in 2030. Science 2008, 319, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Ristic, Z.; Bukovnik, U.; Prasad, P.V.V. Correlation between Heat Stability of Thylakoid Membranes and Loss of Chlorophyll in Winter Wheat under Heat Stress. Crop Sci. 2007, 47, 12. [Google Scholar] [CrossRef]
- Romanus, K.G.; Hussein, S.; Mashela, W.P. Combining ability analysis and association of yield and yield components among selected cowpea lines. Euphytica 2008, 162, 205–210. [Google Scholar] [CrossRef]
- Arndt, S.; Irawan, A.; Sanders, G.J. Apoplastic water fraction and rehydration techniques introduce significant errors in measurements of relative water content and osmotic potential in plant leaves. Physiol. Plant 2015. [Google Scholar] [CrossRef]
- Naveed, M.; Ahsan, M.; Akram, H.M.; Aslam, M.; Ahmed, N. Measurement of cell membrane thermo-stability and leaf temperature for heat tolerance in maize (Zea mays L.): Genotypic variability and inheritance pattern. Maydica 2016, 61, 7. [Google Scholar]
- Steel, R.G.D.; Torrie, J.H.; Dicky, D.A. Principles and Procedures of Statistics: A Biometrical Approach, 3rd ed.; McGraw Hill, Inc.: New York, NY, USA, 1997; pp. 352–358. [Google Scholar]
- Kempthorne, O. An Introduction to Genetic Statistics; John Wiley Sons, Inc.: New York, NY, USA, 1957. [Google Scholar]
- Shaukat, S.; Khan, A.S.; Hussain, M.; Kashif, M.; Ahmed, N. Selection of Superior parents and cross combination for quality traits in Bread Wheat (Triticum aestivum L.) under Normal and Heat stressed conditions. Pak. J. Agric. Sci 2018, 55, 801–807. [Google Scholar] [CrossRef]
- Yousaf, M.I.; Hussain, K.; Hussain, S.; Ghani, A.; Arshad, M.; Mumtaz, A.; Hameed, R.A. Characterization of indigenous and exotic maize hybrids for grain yield and quality traits under heat stress. Int. J. Agric. Biol. 2018, 20, 333–337. [Google Scholar] [CrossRef]
- Hossain, K.M.W.; Raffi, S.A. Evaluation of exotic wheat (Triticum aestivum L.) genotypes for heat tolerance on the basis of physiological phenotyping. J. Bangladesh Agric. Univ. 2018, 16, 457–463. [Google Scholar] [CrossRef]
- Qaseem, M.F.; Qureshi, R.; Shaheen, H. Effects of Pre-Anthesis Drought, Heat and Their Combination on the Growth, Yield and Physiology of diverse Wheat (Triticum aestivum L.) Genotypes Varying in Sensitivity to Heat and drought stress. Sci. Rep. 2019, 9, 6955. [Google Scholar] [CrossRef] [PubMed]
- Ghani, A.; Yousaf, M.I.; Arshad, M.; Hussain, K.; Hussain, D.; Hussain, A.; Shehzad, A. YH-5427: A Highly Productive, Heat tolerant, Stalk Rot and Lodging resistace, Yellow Maize Hybrid of Punjab, Pakistan. Int. J. Biol. Biotechnol. 2020, 17, 561–570. [Google Scholar]
- Fasahat, P. Principles and Utilization of Combining Ability in Plant Breeding. Biom. Biostat. Int. J. 2016, 4. [Google Scholar] [CrossRef]
- Fellahi, Z.E.A.; Hannachi, A.; Bouzerzour, H.; Boutekrabt, A. Line × Tester Mating Design Analysis for Grain Yield and Yield Related Traits in Bread Wheat (Triticum aestivum L.). Int. J. Agron. 2013, 2013, 9. [Google Scholar] [CrossRef]
- Al-Ashkar, I.; Alotaibi, M.; Refay, Y.; Ghazy, A.; Zakri, A.; Al-Doss, A. Selection criteria for high-yielding and early-flowering bread wheat hybrids under heat stress. PLoS ONE 2020, 15, e0236351. [Google Scholar] [CrossRef]
- Bnejdi, F.; Gazzah, M.E. Epistasis and genotype-by-environment interaction of grain protein content in durum wheat. Genet. Mol. Biol. 2010, 130, 125–130. [Google Scholar] [CrossRef]
- Farooq, J.; Khaliq, I.; Ali, M.A.; Kashif, M.; Rehman, A.; Naveed, M.; Ali, Q.; Nazeer, W.; Farooq, A. Inheritance pattern of yield attributes in spring wheat at grain filling stage under different temperature regimes. Aust. J. Crop Sci. 2011, 5, 1745–1753. [Google Scholar]
- Abbasi, S.; Baghizadeh, A.; Mohammadi-nejad, G. Genetic analysis of traits related to grain yield in wheat under drought stress condition by generation mean analysis. Iran. J. Genet. Plant Breed. 2013, 2, 42–46. [Google Scholar]
- Singh, M.K.; Sharma, P.K.; Tyagi, B.S.; Singh, G. Combining ability analysis for yield and protein content in bread wheat (Triticum aestivum). Indian J. Agric. Sci. 2014, 84, 328–336. [Google Scholar]
- Okechukwu, E.C.; Agbo, C.U.; Uguru, M.I.; Ogbonnaya, F.C. Germplasm evaluation of heat tolerance in bread wheat in Tel Hadya, Syria. Chil. J. Agric. Res. 2016, 76, 9–17. [Google Scholar] [CrossRef]
- Sharma, V.I.; Dodiya, N.S.; Dubey, R.B.; Khan, R.U. Combining ability analysis in bread wheat (Triticum aestivum (L.) Em. Thell) under Different Environmental Conditions. Bangladesh J. Bot. 2019, 48, 85–93. [Google Scholar] [CrossRef]
- Sareen, S.; Bhusal, N.; Singh, G.; Tyagi, B.S.; Tiwari, V.; Singh, G.P.; Sarial, A.K. Genetics of Grain Yield and its Components in Wheat under Heat Stress. Cereal Res. Commun. 2018, 46, 448–459. [Google Scholar] [CrossRef]
- Ataei, R.; Gholamhoseini, M.; Kamalizadeh, M. Genetic analysis for quantitative traits in bread wheat exposed to irrigated and drought stress conditions. Int. J. Exp. Bot. 2017, 86, 228–235. [Google Scholar]
- Grover, G.; Sharma, A.; Gill, H.S.; Srivastava, P.; Bains, N.S. Rht8 gene as an alternate dwarfing gene in elite Indian spring wheat cultivars. PLoS ONE 2018, 13, e0199330. [Google Scholar] [CrossRef]
- Divashuk, M.G.; Bespalova, L.A.; Vasilyev, A.V.; Fesenko, I.A.; Puzyrnaya, O.Y.; Karlov, G.I. Reduced height genes and their importance in winter wheat cultivars grown in southern Russia. Euphytica 2013, 190, 137–144. [Google Scholar] [CrossRef]
- Yousaf, M.I.; Maize, A.M.; Maize, W.A. Contribution of Spike-Related Traits for Grain Yield in Spring Wheat. J. Agric. Basic Sci. 2017, 2, 2710–2714. [Google Scholar]
- Menshawy, A.M. Evaluation of some early bread wheat genotypes under different sowing dates. 1. Earliness characters. Egypt. J. Plant Breed. 2007, 11, 25–40. [Google Scholar]
- Khalid, M.U.; Akhtar, N.; Arshad, M.; Yousaf, M.I. Characterization of Maize inbred lines for Grain Yield and related traits under Heat Stress conditions. Int. J. Biol. Biotechnol. 2020, 17, 367–375. [Google Scholar]
- Mumtaz, A.; Hussain, D.; Saeed, M.; Arshad, M.; Yousaf, M.I. Stability and adaptability of sorghum hybrids elucidated with genotype-environment interaction biplots. Turkish J. Field Crop. 2019, 24, 155–163. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal/Field Conditions | Tunnel Conditions | ||||
|---|---|---|---|---|---|
| Month | Temperature (Min./Max.) | Relative Humidity | Precipitation (mm) | Temperature (Min./Max.) | Relative Humidity |
| November | 10/28 °C | 62 | 13 | ||
| December | 9/21 °C | 63 | 5 | 13/26 °C | 75 |
| January | 6/20 °C | 43 | 24 | 9/23 °C | 54 |
| February | 9/20 °C | 30 | 29 | 12/24 °C | 41 |
| March | 14/28 °C | 39 | 5 | 17/32 °C | 50 |
| April | 21/36 °C | 40 | 31 | 25/40 °C | 48 |
| May | 32 | 27/44 °C | 43 | ||
| Sr. # | Parentage | Line/Tester | Sr. # | Parentage | Line/Tester |
|---|---|---|---|---|---|
| 1 | WL7 | L1 | 1 | WT17 | T1 |
| 2 | WL8 | L2 | 2 | WT39 | T2 |
| 3 | WL27 | L3 | 3 | WT83 | T3 |
| 4 | WL37 | L4 | 4 | WT92 | T4 |
| 5 | WL57 | L5 | |||
| 6 | WL71 | L6 | |||
| 7 | WL100 | L7 | |||
| Sr. # | Cross Name | Parentage | Sr. # | Cross Name | Parentage |
| 1 | WL7 × WT17 | L1 × T1 | 15 | WL7 × WT83 | L1 × T3 |
| 2 | WL8 × WT17 | L2 × T1 | 16 | WL8 × WT83 | L2 × T3 |
| 3 | WL27 × WT17 | L3 × T1 | 17 | WL27 × WT83 | L3 × T3 |
| 4 | WL37 × WT17 | L4 × T1 | 18 | WL37 × WT83 | L4 × T3 |
| 5 | WL57 × WT17 | L5 × T1 | 19 | WL57 × WT83 | L5 × T3 |
| 6 | WL71 × WT17 | L6 × T1 | 20 | WL71 × WT83 | L6 × T3 |
| 7 | WL100 × WT17 | L7 × T1 | 21 | WL100 × WT83 | L7 × T3 |
| 8 | WL7 × WT39 | L1 × T2 | 22 | WL7 × WT92 | L1 × T4 |
| 9 | WL8 × WT39 | L2 × T2 | 23 | WL8 × WT92 | L2 × T4 |
| 10 | WL27 × WT39 | L3 × T2 | 24 | WL27 × WT92 | L3 × T4 |
| 11 | WL37 × WT39 | L4 × T2 | 25 | WL37 × WT92 | L4 × T4 |
| 12 | WL57 × WT39 | L5 × T2 | 26 | WL57 × WT92 | L5 × T4 |
| 13 | WL71 × WT39 | L6 × T2 | 27 | WL71 × WT92 | L6 × T4 |
| 14 | WL100 × WT39 | L7 × T2 | 28 | WL100 × WT92 | L7 × T4 |
| Sr.# | Trait Name | Sr. # | Trait Name |
|---|---|---|---|
| 1 | Cell membrane thermostability (CMT) | 9 | Days to maturity (DTM) |
| 2 | Canopy temperature at grain filling stage (CTG) | 10 | Grains per spike (GPS) |
| 3 | Relative water Content (RWC) | 11 | 1000-grain weight (TGW, g) |
| 4 | Plant height (PH, cm) | 12 | Protein (%) |
| 5 | Flag leaf area (FLA, cm2) | 13 | Starch (%) |
| 6 | Peduncle length (PL, cm) | 14 | Gluten (%) |
| 7 | Spike length (SL, cm) | 15 | Net photosynthetic rate (Pn) |
| 8 | Days to heading (DTH) | 16 | Grain yield per plant (GY, g) |
| Mean Squares under Optimal Sowing Conditions | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source of Variation | df | CMT | CTG | RWC | PH | FLA | PL | SL | DTH | DTM | GPS | TGW | Pn | Protein% | Starch% | Gluten% | GYP |
| Replications (R) | 2 | 21.9 ** | 19.5 ** | 11.8 ** | 29.7 ** | 1.4 NS | 2.8 NS | 25.9 ** | 0.3 NS | 7.8 ** | 15.9** | 15.9** | 0.12** | 4.8** | 0.29 NS | 2.1 NS | 0.31 NS |
| Testers (T) | 3 | 152 ** | 5.9 ** | 74.3 ** | 245 ** | 5.2 ** | 28.4 ** | 2.6 ** | 33.8 ** | 49.2 ** | 74.6 ** | 111 ** | 208 ** | 1.6 NS | 1.75 NS | 10.5 ** | 48.1 ** |
| Lines (L) | 6 | 140 ** | 1.6 NS | 9.1 ** | 224 ** | 14.6 ** | 20.7 ** | 6.1 ** | 12.2 ** | 53.4 ** | 33.7 ** | 19.6 ** | 61.1 ** | 4.5 ** | 2.72 * | 8.2 ** | 26.0 ** |
| L × T | 18 | 211 ** | 2.4 ** | 46.8 ** | 76.2 ** | 26.2 ** | 18.5 ** | 9.8 ** | 36.1 ** | 69.9 ** | 55.1 ** | 28.3 ** | 134 ** | 5.4 ** | 2.84 ** | 18.9 ** | 34.2 ** |
| Error | 57 | 0.351 | 0.73 | 0.727 | 0.05 | 0.95 | 1.11 | 0.074 | 0.51 | 0.88 | 0.36 | 0.28 | 0.003 | 1.016 | 1.13 | 1.1 | 0.99 |
| Mean Squares under Heat Stress Conditions | |||||||||||||||||
| Replications | 2 | 20.1 ** | 17.5 ** | 20.9 ** | 13.4 NS | 22.9 ** | 21.6 ** | 22.4 ** | 19.3 ** | 0.3 NS | 0.5 NS | 1.5 NS | 11.6 NS | 0.2 NS | 2.8 NS | 13.1 ** | 0.8 NS |
| Testers (T) | 3 | 92.3 ** | 9.6 ** | 24.3 ** | 617 ** | 5.7 ** | 23.1 ** | 6.6 ** | 59.1 ** | 125 ** | 20.8 ** | 417 ** | 117 ** | 2.2 NS | 4.8 ** | 16.1 ** | 17.4 ** |
| Lines (L) | 6 | 40.1 ** | 4.3 ** | 23.6 ** | 72.0 ** | 4.8 ** | 8.2 ** | 3.5 ** | 42.9 ** | 21.5 ** | 15.1 ** | 188 ** | 66.8 ** | 6.2 ** | 1.2 NS | 7.6 ** | 17.8 ** |
| L × T | 18 | 109 ** | 5.3 ** | 25.7 ** | 84.5 ** | 13.8 ** | 16.7 ** | 9.4 ** | 40.5 ** | 66.5 ** | 40.9 ** | 73.6 ** | 50.9 ** | 5.8 ** | 1.4 NS | 23.9 ** | 10.7 ** |
| Error | 57 | 0.29 | 0.40 | 0.37 | 16.6 | 0.16 | 0.27 | 0.27 | 0.33 | 1.00 | 1.04 | 1.10 | 6.52 | 1.24 | 1.20 | 0.81 | 1.11 |
| Lines/Testers | CMT | CTG | RWC | PH | FLA | PL | SL | DTH | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | |
| WL7 | 5.60 ** | 0.18 | 0.35 | 0.29 | −0.17 | 0.91 ** | −1.60 ** | 0.53 | −1.15** | −0.78 ** | 0.78 * | 0.47 ** | −0.01 | −0.16 | 0.60 ** | 0.47 ** |
| WL8 | −1.75 ** | −1.20 ** | −0.58 * | −0.17 | −0.24 | 0.40 * | 7.93 ** | −0.14 | −0.29 | 0.17 | −0.20 | −1.17 ** | −0.42** | −0.05 | −1.02 ** | −2.51 ** |
| WL27 | −0.09 | −1.86 ** | 0.20 | 0.47 * | 1.16 ** | 1.52 ** | −5.73** | −1.59 | −0.37 | −0.48 ** | −0.66 * | −0.23 | 1.48 ** | 0.38 * | −0.13 | 2.57 ** |
| WL37 | −3.91 ** | −0.16 | −0.16 | 0.73 ** | 0.10 | 1.36 ** | 3.01 ** | 4.90 ** | −0.60* | 0.13 | 0.09 | 0.69 ** | 0.20 * | 0.31 * | −0.18 | 1.06 ** |
| WL57 | 3.39 ** | −1.44 ** | 0.02 | −0.81** | 0.55 * | −1.67 ** | −0.78** | −1.89 | 2.22 ** | 1.22 ** | 1.78 ** | 1.10 ** | −0.56** | −0.27 | 1.90 ** | 1.53 ** |
| WL71 | −0.23 | 1.01 ** | 0.45 | −0.74 ** | 0.25 | −0.67 ** | −1.12** | −2.31 | 0.57* | −0.26 * | 0.60 | −0.87 ** | −0.56** | −0.93 ** | −1.00 ** | −1.64 ** |
| WL100 | −3.00 ** | 3.46 ** | −0.29 | 0.22 | −1.65 ** | −1.84 ** | −1.70** | 0.50 | −0.38 | 0.00 | −2.40 ** | 0.01 | −0.13 | 0.73 ** | −0.16 | −1.47 ** |
| Std. Error | 0.171 | 0.158 | 0.247 | 0.183 | 0.246 | 0.175 | 0.061 | 1.175 | 0.304 | 0.114 | 0.076 | 0.150 | 0.207 | 0.149 | 0.271 | 0.166 |
| WT17 | −3.88 ** | −0.88 ** | 0.62 ** | 0.99 ** | 1.68 ** | −0.01 | 1.75 ** | −1.53 | 0.11 | 0.68 ** | −0.09 | −0.22 | 0.20 ** | −0.52 ** | 1.54 ** | 0.76 ** |
| WT39 | 1.64 ** | −0.46 ** | −0.66 ** | −0.13 | 1.27 ** | 1.47 ** | −5.03** | −6.73** | 0.25 | 0.13 | −1.61 ** | −1.32 ** | 0.07 | 0.41 ** | 0.44 ** | −0.39 ** |
| WT83 | 0.25 | −1.71 ** | −0.10 | −0.42 ** | −0.52 ** | −0.36 ** | 0.83 ** | 2.34 * | −0.73** | −0.45** | 0.78 ** | 0.37 ** | −0.51** | −0.44 ** | −1.37 ** | 1.77 ** |
| WT92 | 1.98 ** | 3.05 ** | 0.14 | −0.44 ** | −2.43 ** | −1.10 ** | 2.46 ** | 5.92 ** | 0.36 | −0.36** | 0.92 ** | 1.17 ** | 0.25 ** | 0.55 ** | −0.61 ** | −2.14 ** |
| Std. Error | 0.129 | 0.119 | 0.186 | 0.138 | 0.186 | 0.132 | 0.046 | 0.888 | 0.212 | 0.086 | 0.230 | 0.113 | 0.058 | 0.113 | 0.157 | 0.126 |
| Lines/Testers | DTM | GPS | TGW | Protein% | Starch% | Gluten% | Pn | GYP | ||||||||
| NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | |
| WL7 | 1.63 ** | 2.35 ** | 2.10 ** | 0.70 * | −0.37 * | −0.20 | 0.50 | 0.75 * | 0.65 * | −0.32 | 0.52 | 0.59 * | −1.93** | −1.83 * | −0.37 | −1.00 ** |
| WL8 | −1.68 ** | 0.67 * | 2.28 ** | −0.13 | −0.27 | −0.44 | 0.18 | 0.09 | −0.25 | −0.11 | −1.02 ** | −0.94 ** | −3.06** | −2.37 ** | −2.30 ** | −0.99 ** |
| WL27 | 2.24 ** | −1.97 ** | −2.09 ** | 0.98 ** | 0.72 ** | 5.17 ** | 0.01 | 0.29 | 0.44 | 0.60 | −0.32 | −1.01 ** | 3.22 ** | 2.60 ** | 2.10 ** | 1.18 ** |
| WL37 | 2.03 ** | −0.54 | −0.45 * | −1.35 ** | 0.93 ** | 0.87 ** | 0.56 | 0.52 | 0.10 | 0.16 | −0.32 | 0.01 | −1.45** | −0.95 | −1.28 ** | −1.03 ** |
| WL57 | 0.20 | 0.17 | −1.14 ** | 0.69 * | 1.35 ** | 3.85 ** | 0.38 | 0.30 | 0.04 | 0.10 | −0.79 * | −0.10 | 1.79 ** | 3.83 ** | 0.58* | 2.06 ** |
| WL71 | −1.14 ** | 0.07 | 0.41 * | 0.85 ** | 0.16 | −2.55 ** | −0.55 | −0.67 * | −0.19 | −0.16 | 0.72 * | 0.24 | 1.32 ** | −1.43 | 1.08 ** | −0.39 |
| WL100 | −3.27 ** | −0.75 * | −1.11 ** | −1.75 ** | −2.52 ** | −6.71 ** | −1.09** | −1.27** | −0.79 * | −0.27 | 1.22 ** | 1.22 ** | 0.10 ** | 0.15 | 0.19 | 0.18 |
| Std. Error | 0.271 | 0.238 | 0.173 | 0.295 | 0.153 | 0.303 | 0.291 | 0.321 | 0.307 | 0.316 | 0.301 | 0.259 | 0.015 | 0.737 | 0.288 | 0.304 |
| WT17 | −2.14 ** | −2.93** | 2.34 ** | 1.48 ** | 1.37 ** | −1.97 ** | 0.27 | 0.38 | 0.11 | −0.17 | −0.98 ** | −1.25 ** | 1.04 ** | 1.64 ** | 0.48* | 0.59* |
| WT39 | 0.12 | −0.90** | −1.97 ** | −0.36 | −2.96 ** | −2.47 ** | 0.20 | 0.16 | −0.09 | 0.05 | −0.04 | 0.23 | 1.46 ** | 2.42 ** | 0.81 ** | 0.95 ** |
| WT83 | 1.47 ** | 2.66 ** | −0.97 ** | −0.66 ** | 2.21 ** | 6.68 ** | −0.19 | −0.30 | −0.35 | −0.51 * | 0.38 | 0.23 | 2.17 ** | −2.07 ** | 0.97 ** | −0.64 ** |
| WT92 | 0.55 ** | 1.17 ** | 0.60 ** | −0.46 * | −0.62 ** | −2.24 ** | −0.28 | −0.24 | 0.33 | 0.63 * | 0.63 ** | 0.79 ** | −4.67** | −1.98 ** | −2.25 ** | −0.91 ** |
| Std. Error | 0.205 | 0.219 | 0.131 | 0.223 | 0.115 | 0.229 | 0.220 | 0.243 | 0.232 | 0.239 | 0.228 | 0.196 | 0.011 | 0.557 | 0.218 | 0.229 |
| Crosses | CMT | CTG | RWC | PH | FLA | PL | SL | DTH | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | |
| WL7 × WT17 | −11.47** | −2.61** | 1.92 ** | 1.28 ** | −1.08* | 2.53 ** | 8.42 ** | 7.61 ** | 1.12 | −1.89** | −2.06 ** | −2.28 ** | −1.57 ** | −0.23 | −0.83 * | −2.57 ** |
| WL8 × WT17 | 1.47 ** | −5.44 ** | 0.29 | 0.65 | 5.09 ** | 0.04 | 2.21 ** | 0.18 | −3.64 ** | 1.75 ** | −0.31 | −0.54 | 1.30 ** | 2.08 ** | −0.79 | 2.56 ** |
| WL27 × WT17 | 5.47 ** | −2.08** | −0.90 | −2.10 ** | 1.79 ** | −0.26 | −4.79 ** | 1.67 | −0.56 | −0.45 | −2.55 ** | 2.71 ** | 2.94 ** | 0.35 | −2.02 ** | −5.15 ** |
| WL37 × WT17 | −5.34 ** | 2.31 ** | −0.01 | −0.25 | −1.16* | 1.22 ** | −6.47 ** | −3.62 | −3.47** | −2.97** | 0.74 | 1.29 ** | 1.01 ** | 0.85 ** | 2.45 ** | 1.35 ** |
| WL57 × WT17 | −3.04 ** | 8.99 ** | −0.29 | 0.91* | 0.36 | 0.45 | −0.58 ** | −5.90* | 3.79 ** | 1.90 ** | 0.90 | −2.61 ** | −2.00 ** | −1.73 ** | 0.40 | −0.11 |
| WL71 × WT17 | 2.73 ** | −6.19** | −0.71 | −0.84* | −7.62 ** | −5.19 ** | 4.92 ** | 1.50 | 2.51 ** | 1.48 ** | 4.06 ** | 4.20 ** | −1.05 ** | −0.46 | −0.66 | −0.34 |
| WL100 × WT17 | 10.17 ** | 5.03 ** | −0.30 | 0.34 | 2.62 ** | 1.19 ** | −3.70 ** | −1.44 | 0.26 | 0.18 | −0.79 | −2.76 ** | −0.62 ** | −0.86 ** | 1.46 ** | 4.26 ** |
| WL7 × WT39 | 15.04 ** | 6.84 ** | −1.47 ** | −1.90 ** | 0.95 | 2.51 ** | −6.62 ** | −16.85** | −3.87** | −0.84 | −3.17 ** | 0.66 * | 0.94 ** | 1.14 ** | 3.84 ** | 4.59 ** |
| WL8 × WT39 | −2.52 ** | −2.30** | −1.46 ** | −0.53 | −5.97** | −3.98 ** | −9.87 ** | −7.51 ** | 1.47* | −1.86** | −0.90 | −1.49 ** | 0.67 ** | 0.00 | −4.16 ** | −4.44 ** |
| WL27 × WT39 | 6.41 ** | −3.82** | −0.95 | 0.13 | 2.04 ** | 2.85 ** | −6.42 ** | −6.09* | −1.62** | −0.33 | −2.77 ** | −3.38 ** | −0.93 ** | 1.43 ** | 0.63 | 0.85 * |
| WL37 × WT39 | 4.34 ** | 0.37 | −0.93 | −0.41 | −1.24* | 0.08 | −8.75 ** | −5.67* | 2.67 ** | 0.13 | −0.49 | 0.63 * | 0.64 ** | 2.72 ** | −1.52 ** | −5.02 ** |
| WL57 × WT39 | 1.34 ** | −4.21** | −2.90 ** | −1.65 ** | 2.21** | 3.77 ** | −5.67 ** | 0.16 | −0.21 | 1.24 ** | −0.23 | 1.13 ** | −1.90 ** | −0.83 ** | −2.63 ** | −2.53 ** |
| WL71 × WT39 | 9.01 ** | 2.38 ** | −0.52 | 0.24 | −1.26* | 1.85 ** | −8.77 ** | −2.25 | 1.50 ** | −0.43 | −1.10 | −4.55 ** | 1.21 ** | 2.87 ** | 2.31 ** | 3.06 ** |
| WL100 × WT39 | 5.03 ** | 3.67 ** | −0.72 | −3.77 ** | 0.41 | 3.25 ** | −1.37 ** | 1.86 | 1.06 | −1.69** | −1.99 ** | −0.66 * | −1.53 ** | −0.77 * | −6.19 ** | −4.60 ** |
| WL7 × WT83 | −1.56 ** | −4.00** | −2.01 ** | −0.20 | 0.51 | −0.08 | −7.56 ** | 0.89 | −0.15 | 4.09 ** | 0.79 | 0.18 | 2.03 ** | 1.98 ** | −1.68 ** | −0.66 |
| WL8 × WT83 | −8.74 ** | 3.03 ** | 1.10* | −1.10 ** | −2.74 ** | 1.32 ** | 3.56 ** | 3.40 | −0.90 | −1.60** | −2.17 ** | 0.82 ** | −2.15 ** | −2.16 ** | 2.77 ** | −1.68 ** |
| WL27 × WT83 | −6.02 ** | 8.01 ** | 0.51 | 0.61 | −0.06 | −1.17 ** | 3.68 ** | −3.03 | 2.18 ** | 0.87 ** | 0.39 | 0.64 * | −2.33 ** | −1.49 ** | −4.31 ** | 0.84 * |
| WL37 × WT83 | 7.59 ** | 5.23 ** | 0.70 | 0.11 | −2.88** | −2.60 ** | 3.83 ** | 0.38 | −2.42** | −1.78** | −1.43* | −3.16 ** | 0.02 | −0.31 | −2.19 ** | 3.35** |
| WL57 × WT83 | 13.52 ** | −1.91** | 0.42 | −1.49 ** | 0.58 | 0.34 | 0.31* | 1.14 | −0.63 | −1.87** | −0.97 | −1.42 ** | 1.43 ** | 1.90 ** | −1.15 ** | −1.83 ** |
| WL71 × WT83 | 2.06 ** | −5.21** | 0.14 | −0.36 | 3.79 ** | 2.55 ** | 2.61 ** | 3.37 | −0.16 | −0.81** | −0.81 | 0.49 | −0.28 | −1.30 ** | 0.19 | −3.42 ** |
| WL100 × WT83 | −6.86 ** | −5.14 ** | −0.85 | 2.43 ** | 0.80 | −0.35 | −6.43 ** | −6.15* | 2.08 ** | 1.10** | 4.19** | 2.47 ** | 1.29 ** | 1.38 ** | 6.38 ** | 3.40 ** |
| WL7 × WT92 | 3.51 ** | 0.19 | 0.28 | −0.31 | −0.79 | −3.48 ** | −1.03 ** | 3.16 | 3.05 ** | −1.90** | 2.92 ** | 0.36 | −1.52 ** | −1.95 ** | −2.43 ** | −2.53** |
| WL8 × WT92 | 15.31** | 5.12 ** | −1.21* | −0.15 | 3.21 ** | 4.09 ** | −2.68 ** | −1.26 | 3.21 ** | 1.18 ** | 1.86 ** | 0.11 | 0.06 | 1.01 ** | 1.08 * | 2.41 ** |
| WL27 × WT92 | −0.35 | −1.69** | 0.07 | 0.24 | −4.19 ** | 0.05 | 0.75 ** | 2.26 | 0.15 | −0.63** | 3.41** | −1.06 ** | 0.19 | 0.65 * | 4.60 ** | 2.31 ** |
| WL37 × WT92 | −1.07 ** | −7.49** | −1.04* | −0.58 ** | 4.87 ** | 2.78 ** | 4.62 ** | 3.72 | 3.37 ** | 4.08 ** | −0.35 | 0.15 | −1.80 ** | −2.33 ** | 0.16 | −0.83 * |
| WL57 × WT92 | −6.30 ** | −2.45** | 1.50 ** | 1.10 ** | −3.55 ** | −3.08 ** | −0.84 ** | −0.59 | −2.81** | −1.80** | −1.23* | 1.81 ** | 2.34 ** | 1.61 ** | 2.28 ** | 3.31 ** |
| WL71 × WT92 | −8.28 ** | 9.45 ** | −0.19 | −0.17 | 4.68 ** | 2.27 ** | −5.54 ** | −7.82 ** | −3.70** | −0.79** | −3.67 ** | −1.24 ** | −0.01 | −0.17 | −2.94 ** | −0.45 |
| WL100 × WT92 | −2.82 ** | −3.13** | 0.59 | −0.13 | −4.24 ** | −2.61 ** | 4.72 ** | 0.53 | −3.26** | −0.13 | −2.94 ** | −0.14 | 0.73 ** | 1.19 ** | −2.75 ** | −4.21 ** |
| S. E | 0.342 | 0.316 | 0.493 | 0.365 | 0.492 | 0.349 | 0.122 | 2.350 | 0.561 | 0.227 | 0.607 | 0.300 | 0.152 | 0.299 | 0.414 | 0.332 |
| Wheat Crosses | DTM | GPS | TGW | Protein% | Starch% | Gluten% | Pn | GYP | ||||||||
| NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | NT | HT | |
| WL7 × WT17 | 2.85 ** | 0.76 | 0.09 | 1.41 * | −2.6 ** | −3.22** | −1.90** | −2.03** | 1.67 ** | −0.76 | 0.59 | −0.70 | −10.2** | −6.38** | −5.41** | −3.3 ** |
| WL8 × WT17 | 6.36 ** | −0.02 | 1.97 ** | 1.90 ** | 4.06 ** | 5.68 ** | −1.63** | −1.30* | −1.46 * | 0.08 | 2.82 ** | 3.69 ** | 5.67 ** | −0.05 | 2.68 ** | 1.59 * |
| WL27 × WT17 | −0.71 | −2.0** | 7.43 ** | 3.82 ** | 1.41 ** | −2.73** | −0.61 | −0.66 | −0.04 | −0.04 | 0.38 | −1.36 * | 4.12 ** | 3.82 * | 2.07 ** | 1.15 |
| WL37 × WT17 | −1.24 * | −0.23 | −1.16** | 0.27 | −1.76** | −1.65** | −0.40 | −0.07 | −0.55 | −0.20 | 0.24 | 0.73 | 8.47 ** | 3.96 ** | 4.84 ** | 1.46 * |
| WL57 × WT17 | −5.1 ** | −4.9** | −4.52** | −3.80** | 1.61 ** | −0.69 | 1.03 | 1.46 ** | −0.65 | −0.31 | −2.15** | −0.34 | −0.61** | −0.50 | −0.44 | −0.19 |
| WL71 × WT17 | −3.27** | 2.6 * | −4.18** | −4.97** | −4.21** | −2.21** | 2.15 ** | 1.45 ** | 0.66 | 0.96 | −0.62 | −1.54 ** | −8.65** | −0.97 | −3.73** | −0.99 |
| WL100 × WT17 | 1.14 * | 3.7 ** | 0.37 | 1.37 ** | 1.48 ** | 4.82 ** | 1.35 * | 1.15 | 0.36 | 0.26 | −1.26 * | −0.48 | 1.22 ** | 0.12 | −0.01 | 0.32 |
| WL7 × WT39 | 5.17 ** | 6.6 ** | 2.20 ** | 1.21 ** | −4.58** | −5.25** | −0.11 | −0.03 | −0.17 | 1.23 | 0.40 | 1.12 * | −1.77** | 0.90 | −1.12 | 0.51 |
| WL8 × WT39 | 4.57 ** | 3.6 ** | −4.04 ** | −3.79** | −6.66** | −7.26** | 0.13 | −0.31 | 0.67 | 0.86 | 0.09 | 0.62 | 3.40 ** | 5.25 ** | 1.85 ** | 1.65 ** |
| WL27 × WT39 | 4.32 ** | 3.1 ** | −6.76** | 1.72 ** | −2.53** | 4.13 ** | 0.72 | 0.45 | −0.43 | −0.78 | 4.14 ** | 6.07 ** | −8.82** | −3.83 * | −3.75** | −1.5 * |
| WL37 × WT39 | −1.34 * | 3.8 ** | −5.42** | −1.83** | 1.10 ** | 5.43 ** | 0.74 | 1.31 * | 0.23 | 0.37 | −0.80 | −0.33 | 4.77 ** | 5.19 ** | 1.82 ** | 2.85 ** |
| WL57 × WT39 | −5.38** | −2.0** | −8.56** | −2.81** | −4.41** | 7.44 ** | −0.15 | −0.41 | 0.41 | 0.61 | 0.84 | 0.85 | 2.26 ** | 1.89 | 0.85 | 0.65 |
| WL71 × WT39 | 6.93 ** | −1.9 * | −3.03** | −4.92** | −6.91** | −5.05** | −1.12 | −1.52 * | −0.18 | 0.05 | 4.23 ** | 4.66 ** | 6.73 ** | −5.45** | 4.04 ** | −2.3 ** |
| WL100 × WT39 | 1.48 ** | 1.02 * | −4.54 ** | −2.43** | −6.36** | −2.97** | −0.66 | −1.09 | −1.9 ** | −0.78 | −2.35** | −2.65** | −3.59** | 1.51 | −1.39** | 0.69 |
| WL7 × WT83 | 1.16 * | −0.99 | 1.44 ** | 1.40 ** | 1.47 ** | 3.02 ** | 0.55 | 0.41 | −1.49 * | −0.58 | 2.23 ** | 2.91 ** | 9.40 ** | 1.74 | 5.32 ** | 1.04 |
| WL8 × WT83 | −5.30** | 1.8 ** | −2.76** | −0.93 | −0.04 | 1.96 ** | 0.64 | 0.43 | 0.27 | 0.19 | −0.38 | −0.76 | −7.53** | −2.59 | −3.65** | −1.03 |
| WL27 × WT83 | −4.17** | 4.8 ** | −1.21** | −2.12** | −1.44 ** | −4.58** | 0.84 | 1.00 | 0.51 | 0.37 | −1.14 | −0.78 | 3.29 ** | −2.55 | 1.95 ** | −0.24 |
| WL37 × WT83 | 1.77 ** | −9.9** | −1.20** | −3.58** | −1.81** | −0.12 | 1.41 * | 0.90 | −0.03 | 0.15 | 1.80 ** | 1.45 ** | −9.35** | −4.38** | −4.67** | −2.1 ** |
| WL57 × WT83 | 5.98 ** | 4.5 ** | 5.73 ** | 0.57 | −2.97** | −4.10** | −1.76** | −2.18** | 0.10 | 0.08 | −1.69** | −2.82** | −3.76** | 0.30 | −1.65** | −0.36 |
| WL71 × WT83 | 1.34 * | 1.3 ** | −0.81* | 5.10 ** | 1.95 ** | 5.47 ** | −0.69 | 0.04 | −0.98 | −0.79 | −3.17** | −3.11** | 2.88 ** | 5.58 ** | −0.05 | 2.27 ** |
| WL100 × WT83 | −0.78 | −1.6 ** | −1.19** | −0.42 | 2.84 ** | −1.64 ** | −1.00 | −0.59 | 1.62 * | 0.58 | 2.35 ** | 3.10 ** | 5.07 ** | 1.90 | 2.76 ** | 0.42 |
| WL7 × WT92 | −6.93** | −4.4 ** | −8.03** | −5.85** | 1.37 ** | 4.95 ** | 1.39 * | 1.42 * | −0.22 | 0.33 | −2.29** | −1.85** | 3.01 ** | 4.52 ** | 1.54 | 2.14 ** |
| WL8 × WT92 | −3.38** | −3.4 ** | 0.53 | 0.99 | −1.69** | −0.88 | 0.80 | 0.96 | 0.32 | −0.91 | −1.60 * | −2.08** | −1.10** | −1.83 | −0.55 | −1.8 ** |
| WL27 × WT92 | 2.81 ** | −3.9** | −3.77** | −5.26** | −1.78** | 2.68 ** | −1.02 | −1.02 | −0.23 | 0.67 | −2.44 ** | −2.45** | 1.84 ** | 3.34 * | 0.06 | 1.03 |
| WL37 × WT92 | 3.06 ** | 8.4 ** | 3.48 ** | 3.31** | −1.86** | −4.17** | −1.82** | −2.36** | 0.15 | −0.10 | −0.31 | −0.37 | −3.46** | −3.99** | −1.66** | −1.8 ** |
| WL57 × WT92 | 6.79 ** | 4.5** | 3.04 ** | 4.20 ** | 1.43 ** | −3.15** | 0.82 | 0.90 | −0.06 | −0.16 | 3.94 ** | 3.79 ** | 2.53 ** | −0.91 | 1.57 ** | 0.25 |
| WL71 × WT92 | −2.75 ** | −0.1** | 3.71 ** | 2.96 ** | 4.83 ** | 1.29 * | −0.41 | −0.20 | 0.30 | 0.00 | 0.50 | 1.46** | −0.54 ** | 1.62 | 0.07 | 1.34 * |
| WL100 × WT92 | 0.40 | −1.2** | 1.05 ** | −0.35 | −2.29 ** | −0.72 | 0.24 | 0.30 | −0.26 | 0.17 | 2.19 ** | 1.50 ** | −2.27** | −2.75 | −1.03 | −1.07 |
| S. E | 0.54 | 0.58 | 0.346 | 0.589 | 0.305 | 0.606 | 0.582 | 0.643 | 0.614 | 0.631 | 0.602 | 0.519 | 0.030 | 1.474 | 0.576 | 0.607 |
| Mean Values of Four Classes of Wheat Hybrid Crosses under Optimal Conditions | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Class | CMT | CTG | RWC | PH | FLA | PL | SL | DTH | DTM | GPS | TGW | PRO | STR | GLU | Pn | GYP |
| 1 | 43.7 | 25.7 | 60.3 | 111.6 | 27.5 | 41.4 | 12.9 | 98.2 | 130.1 | 57.4 | 39.8 | 13.7 | 55.0 | 35.1 | 23.4 | 15.10 |
| 2 | 45.8 | 25.0 | 62.5 | 99.1 | 28.9 | 41.7 | 14.3 | 99.4 | 130.0 | 54.9 | 40.6 | 13.8 | 54.8 | 34.5 | 33.8 | 20.55 |
| 3 | 62.9 | 24.9 | 61.9 | 103.2 | 28.8 | 43.5 | 13.7 | 99.7 | 130.7 | 58.1 | 38.9 | 14.2 | 55.3 | 33.1 | 26.5 | 16.83 |
| Mean Values of Four Classes of Wheat Hybrid Crosses under Heat Stress Conditions | ||||||||||||||||
| 1 | 57.9 | 24.7 | 63.3 | 101.5 | 28.9 | 31.6 | 13.1 | 84.1 | 113.1 | 48.0 | 36.9 | 12.3 | 53.5 | 34.4 | 15.3 | 9.06 |
| 2 | 53.7 | 24.8 | 62.7 | 96.2 | 28.5 | 31.6 | 12.9 | 88.4 | 112.1 | 47.9 | 50.3 | 13.7 | 53.8 | 34.3 | 20.4 | 11.93 |
| 3 | 58.4 | 25.5 | 61.0 | 89.7 | 28.7 | 31.4 | 12.9 | 89.0 | 112.1 | 48.0 | 39.5 | 13.5 | 53.8 | 35.7 | 20.5 | 11.83 |
| Treatments | Class No. | Name of Wheat Crosses/Genotypes |
|---|---|---|
| Optimal | Class 1 | WL7 × WT17, WL8 × WT17, WL71 × WT17, WL8 × WT83, WL37 × WT83, WL37 × WT92, WL100 × WT92 |
| Class 2 | WL27 × WT17, WL37 × WT17, WL57 × WT17, WL100 × WT17, WL8 × WT39, WL27 × WT39, WL37 × WT39, WL57 × WT39, WL71 × WT39, WL100 × WT39, WL7 × WT83, WL27 × WT83, WL71 × WT83, WL100 × WT83, WL27 × WT92, WL57 × WT92, WL71 × WT92 | |
| Class 3 | WL7 × WT39, WL57 × WT83, WL7 × WT92, WL8 × WT92 | |
| Heat Stress | Class 1 | WL7 × WT17, WL100 × WT39, WL8 × WT92, WL37 × WT92, WL100 × WT92 |
| Class 2 | WL8 × WT17, WL27 × WT17, WL27 × WT39, WL37 × WT39, WL57 × WT39, WL7 × WT83, WL8 × WT83, WL27 × WT83, WL37 × WT83, WL57 × WT83, WL71 × WT83, WL7 × WT92, WL27 × WT92 | |
| Class 3 | WL37 × WT17, WL57 × WT17, WL71 × WT17, WL100 × WT17, WL7 × WT39, WL8 × WT39, WL71 × WT39, WL100 × WT83, WL57 × WT92, WL71 × WT92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riaz, M.W.; Yang, L.; Yousaf, M.I.; Sami, A.; Mei, X.D.; Shah, L.; Rehman, S.; Xue, L.; Si, H.; Ma, C. Effects of Heat Stress on Growth, Physiology of Plants, Yield and Grain Quality of Different Spring Wheat (Triticum aestivum L.) Genotypes. Sustainability 2021, 13, 2972. https://doi.org/10.3390/su13052972
Riaz MW, Yang L, Yousaf MI, Sami A, Mei XD, Shah L, Rehman S, Xue L, Si H, Ma C. Effects of Heat Stress on Growth, Physiology of Plants, Yield and Grain Quality of Different Spring Wheat (Triticum aestivum L.) Genotypes. Sustainability. 2021; 13(5):2972. https://doi.org/10.3390/su13052972
Chicago/Turabian StyleRiaz, Muhammad Waheed, Liu Yang, Muhammad Irfan Yousaf, Abdul Sami, Xu Dong Mei, Liaqat Shah, Shamsur Rehman, Liu Xue, Hongqi Si, and Chuanxi Ma. 2021. "Effects of Heat Stress on Growth, Physiology of Plants, Yield and Grain Quality of Different Spring Wheat (Triticum aestivum L.) Genotypes" Sustainability 13, no. 5: 2972. https://doi.org/10.3390/su13052972
APA StyleRiaz, M. W., Yang, L., Yousaf, M. I., Sami, A., Mei, X. D., Shah, L., Rehman, S., Xue, L., Si, H., & Ma, C. (2021). Effects of Heat Stress on Growth, Physiology of Plants, Yield and Grain Quality of Different Spring Wheat (Triticum aestivum L.) Genotypes. Sustainability, 13(5), 2972. https://doi.org/10.3390/su13052972