
Purification Effects on β-HCH Removal and Bacterial Community Differences of Vertical-Flow Constructed Wetlands with Different Vegetation Plantations

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
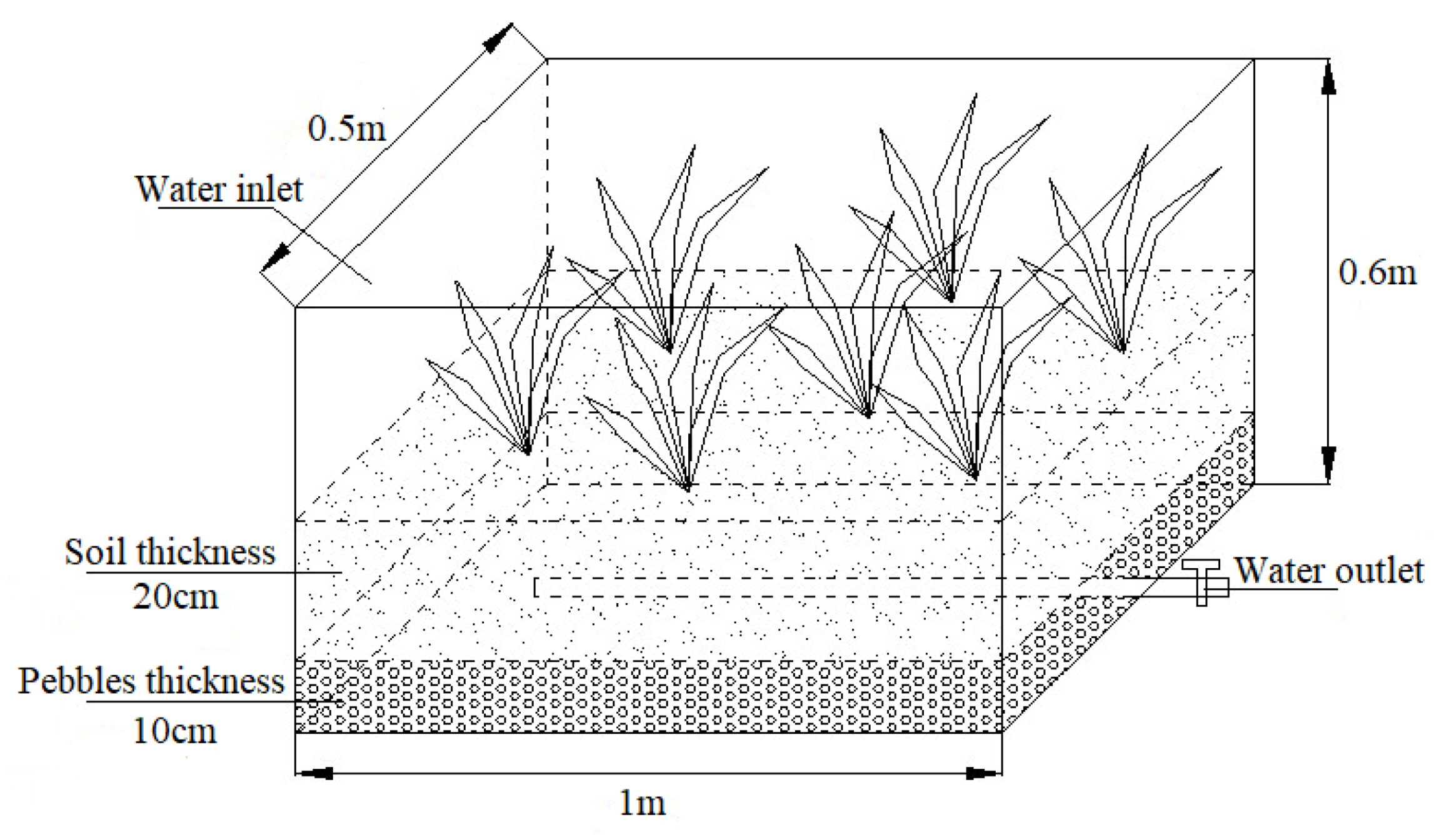
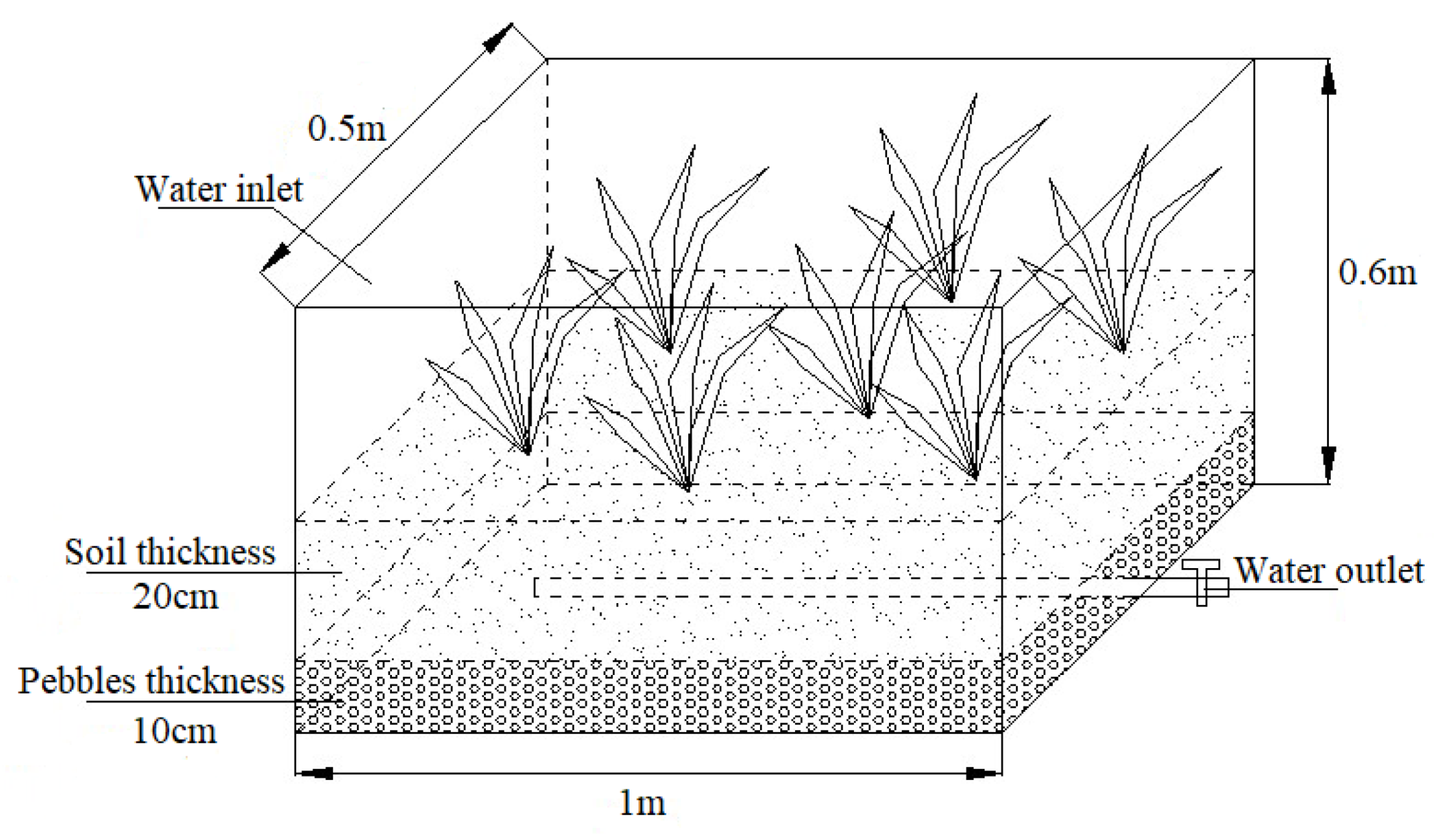
2.1. Experimental Setup and Operating Conditions
2.2. Sample Collection
2.3. Instrumentation and Chemicals
2.4. Sample Analysis
2.5. Data Analysis
3. Results and Discussion
3.1. β-HCH Removal
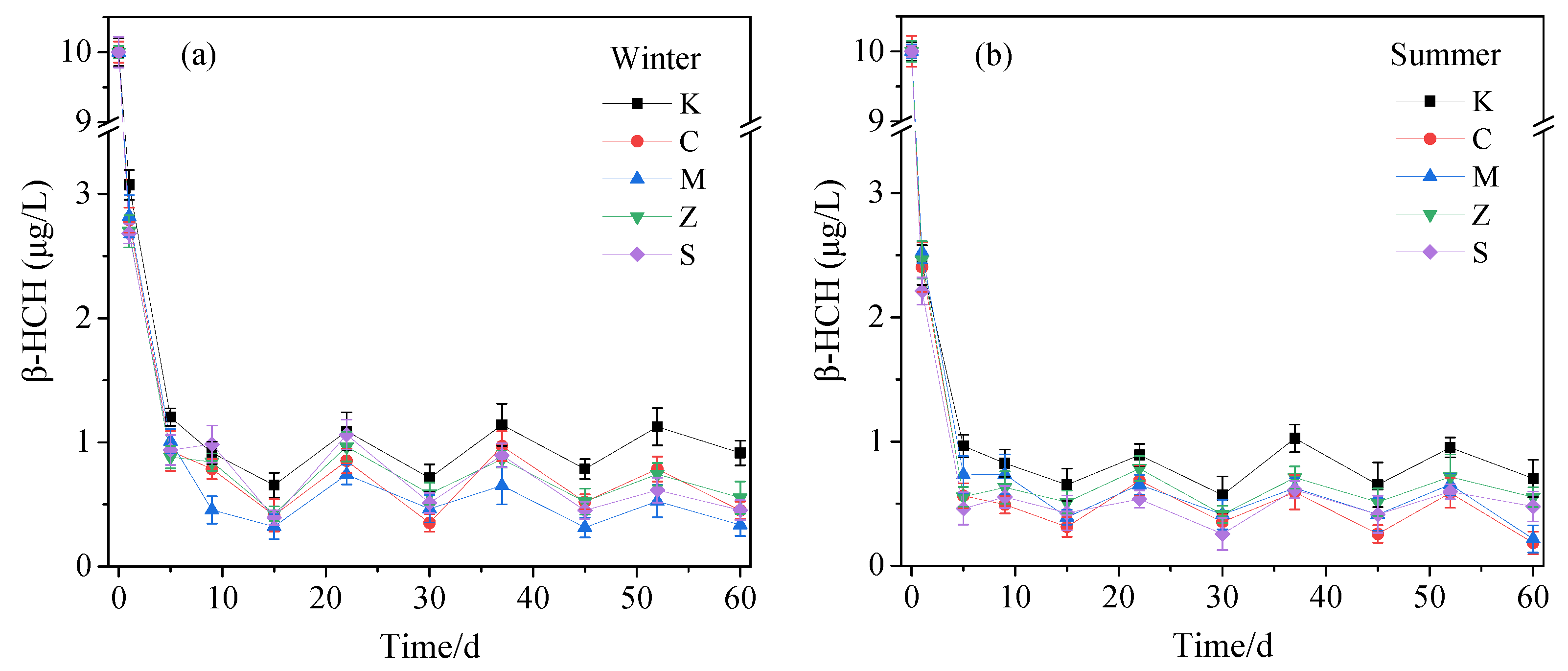
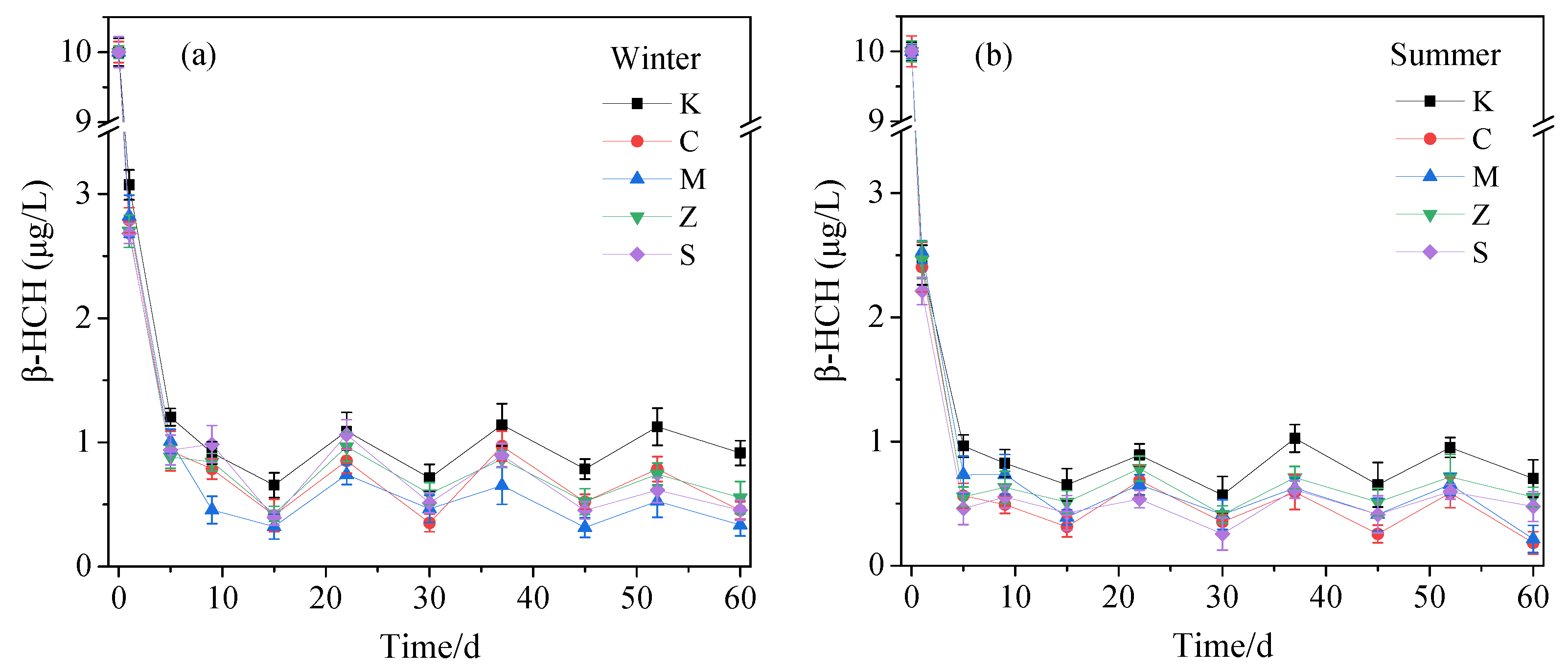
3.1.1. Removal of β-HCH in Water
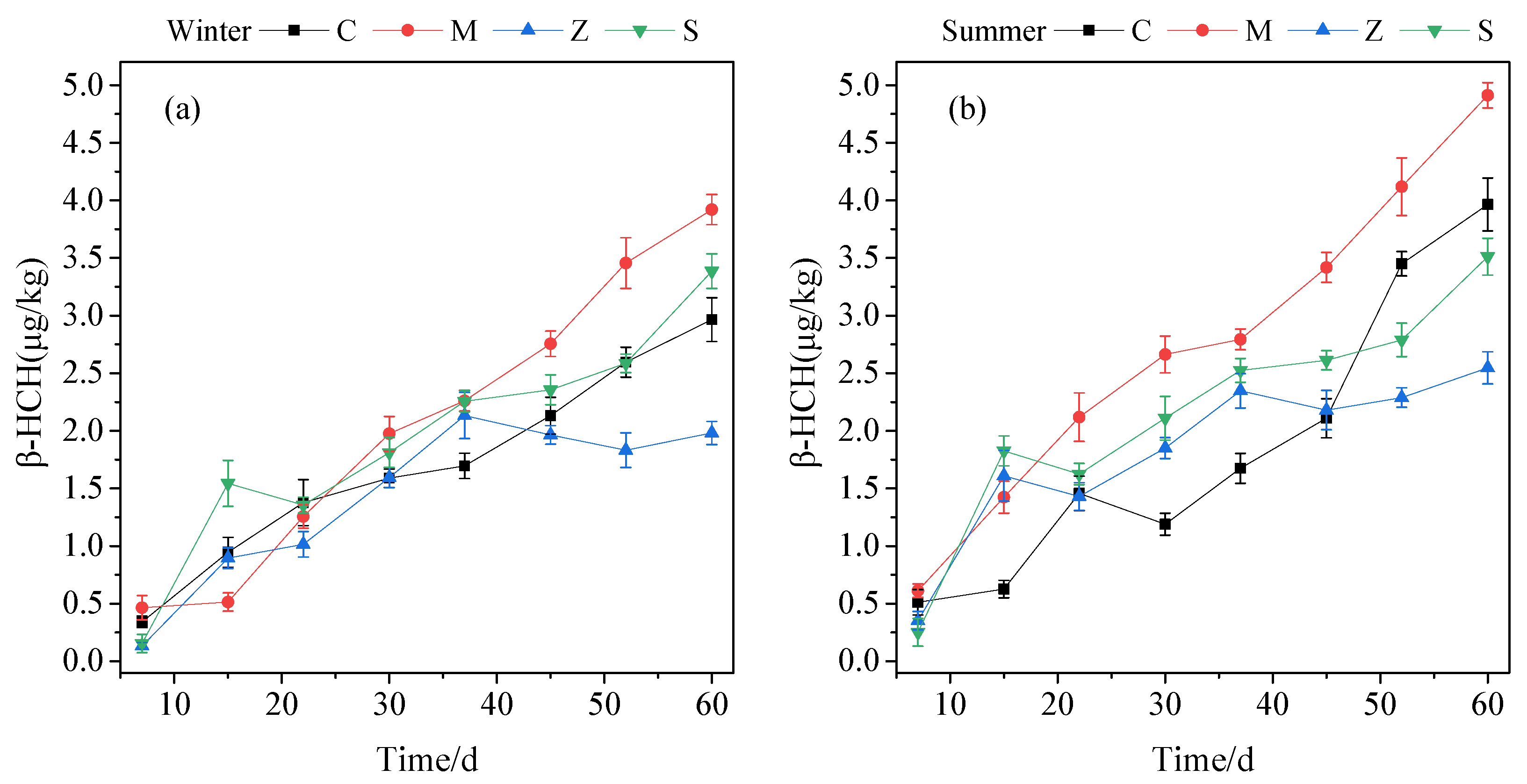
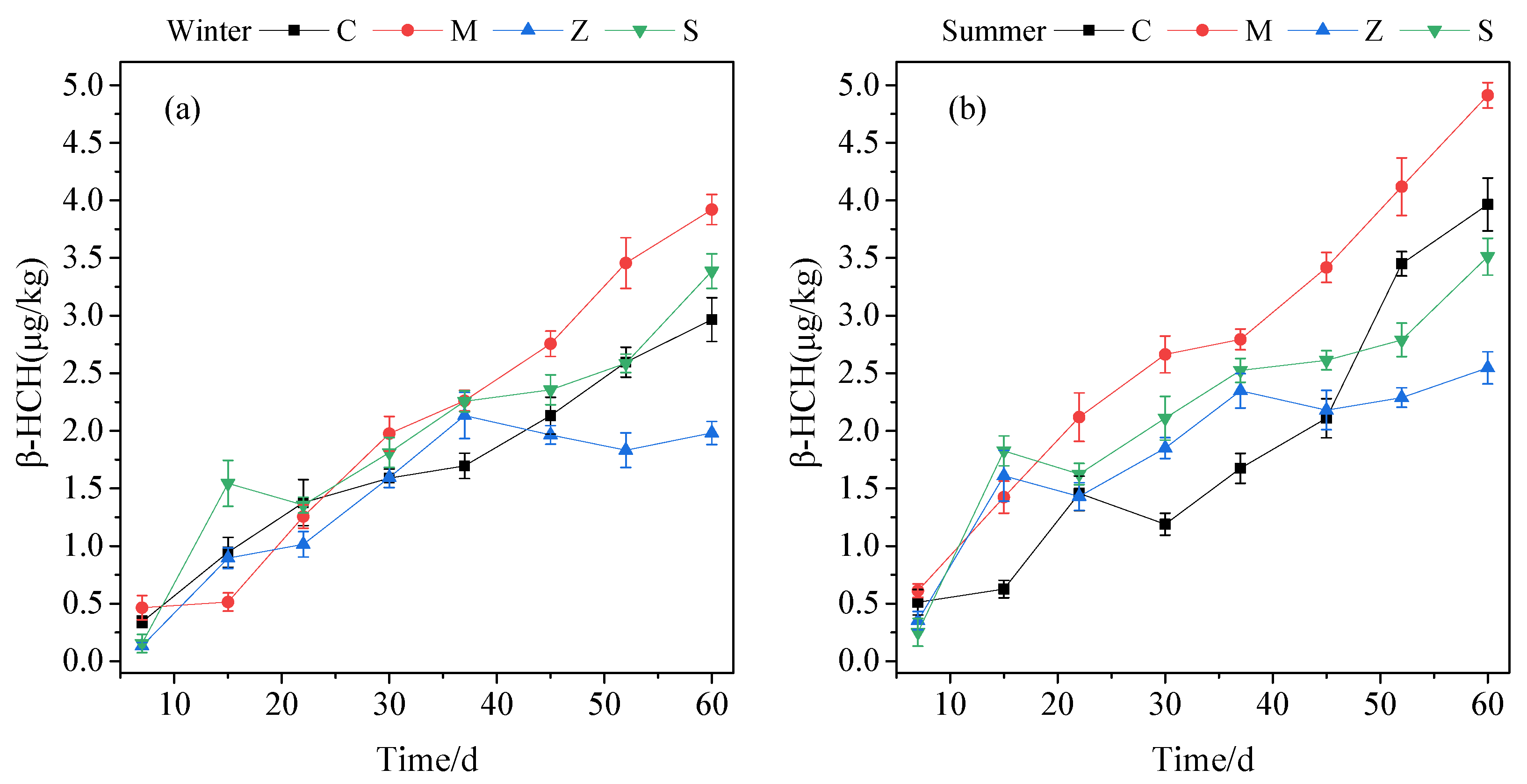
3.1.2. Plant Uptake
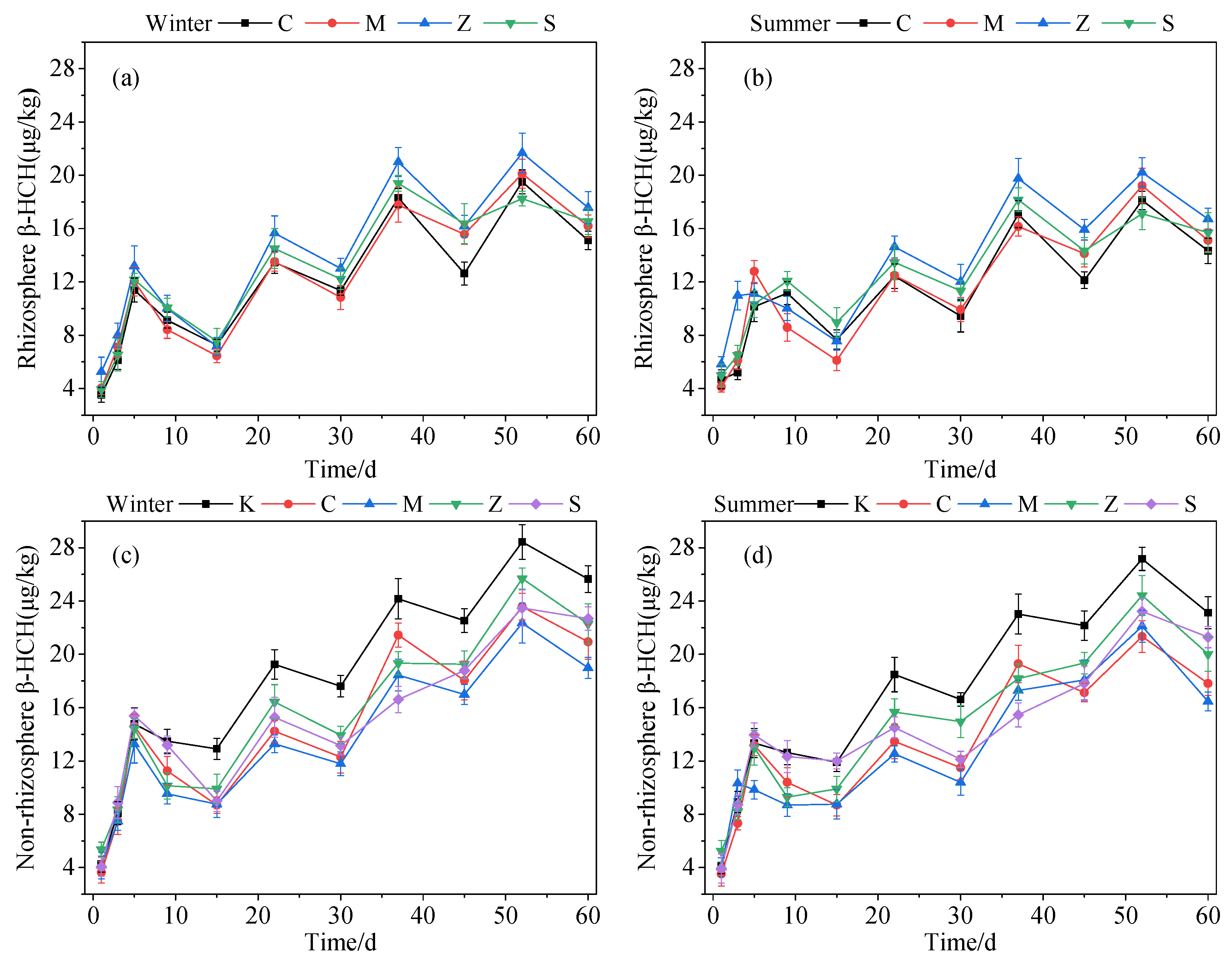
3.1.3. Substrate Sorption
3.2. Bacterial Community Structure
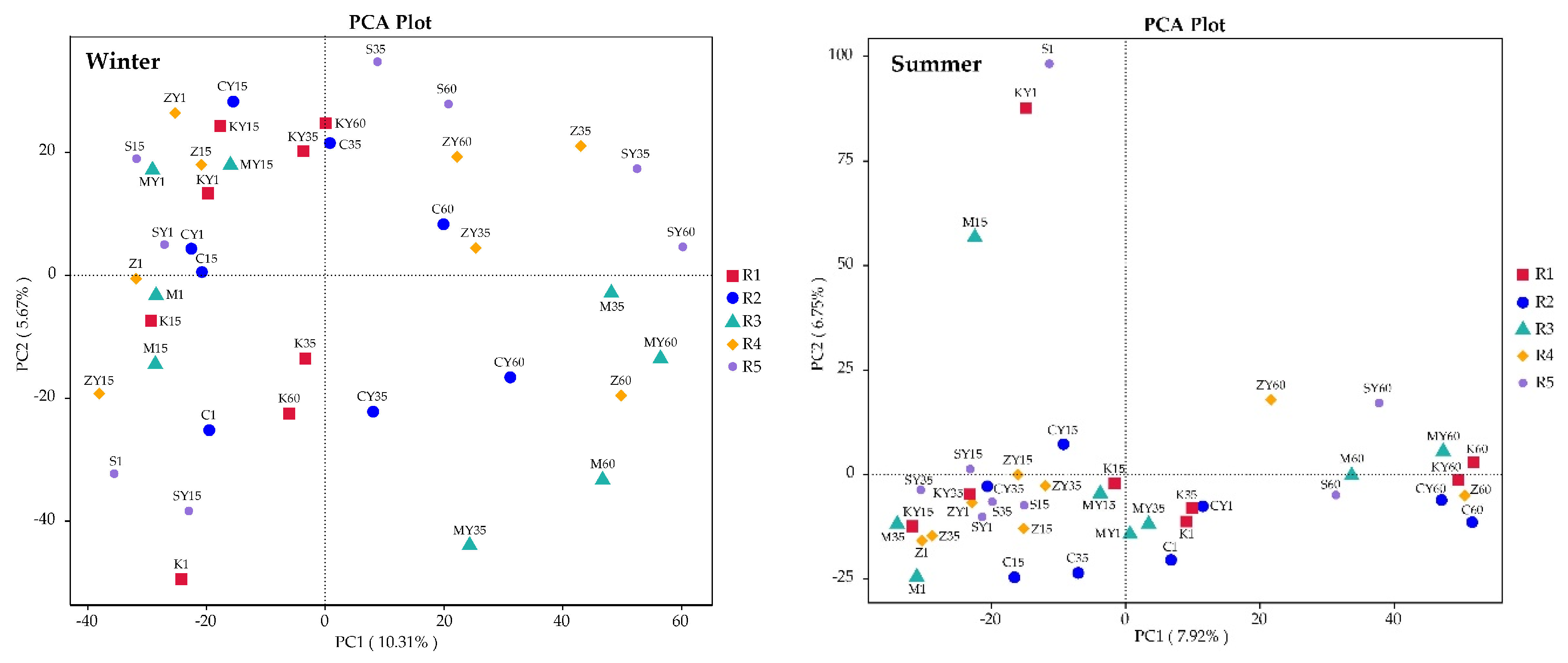
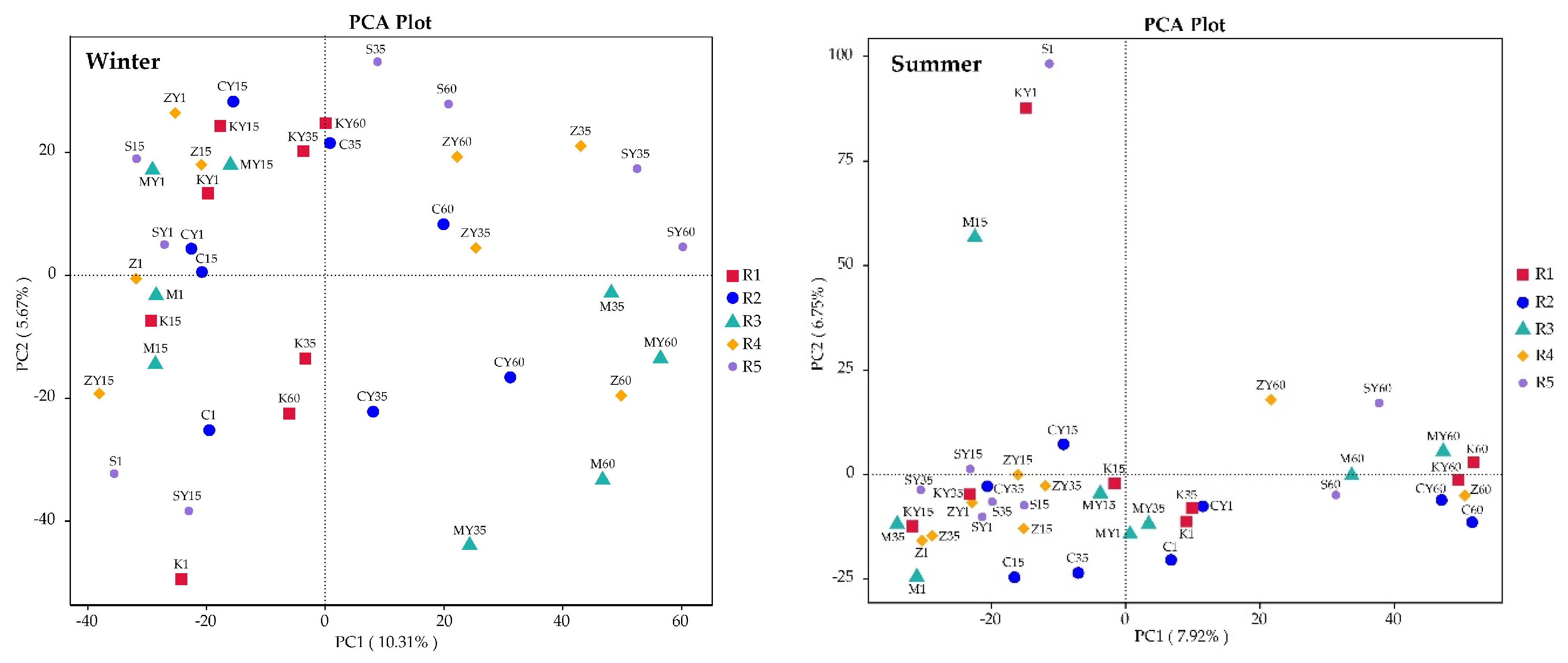
3.2.1. Alterations in Abundance and Diversity of Bacterial Community Caused by β-HCH
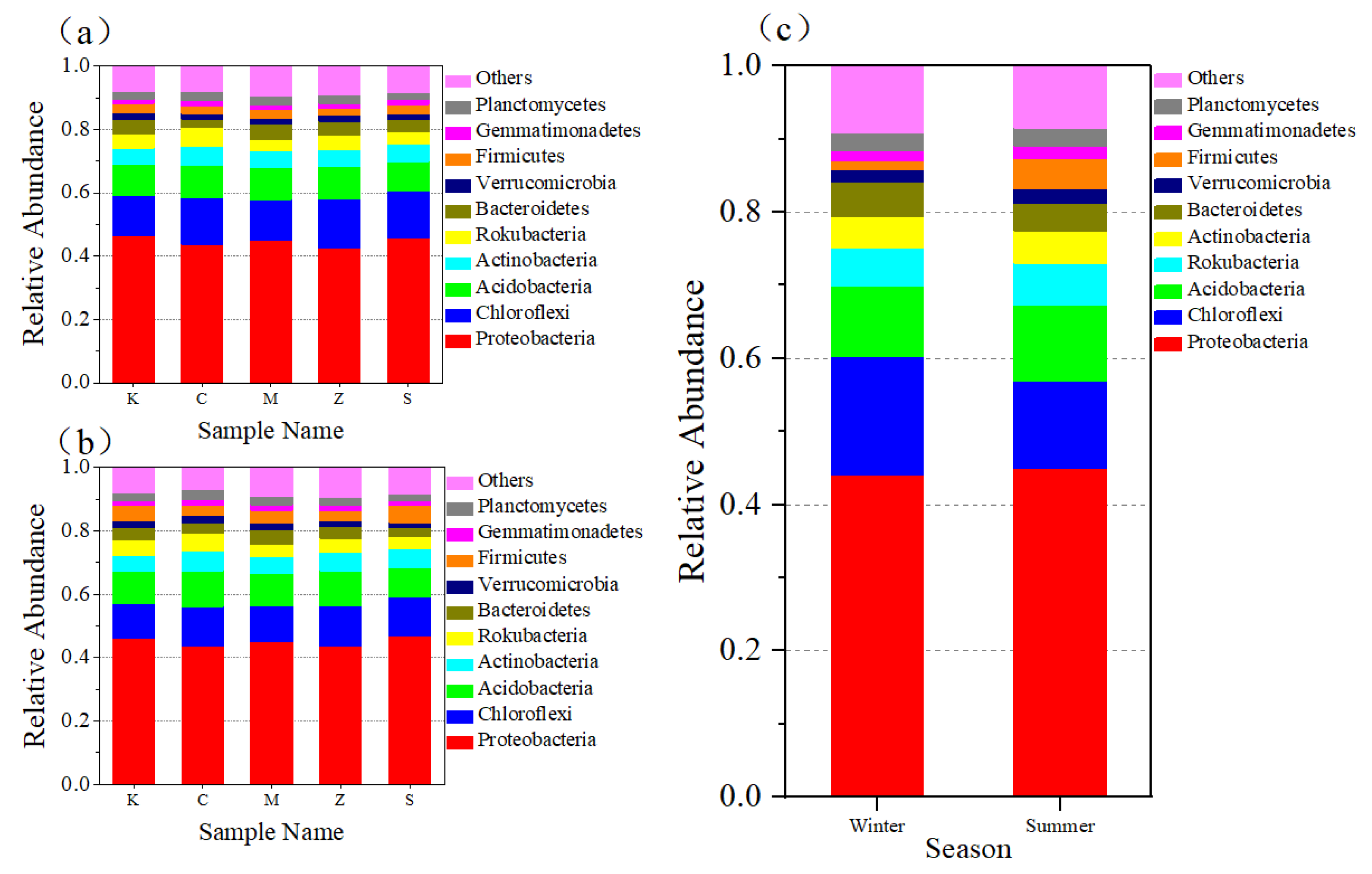
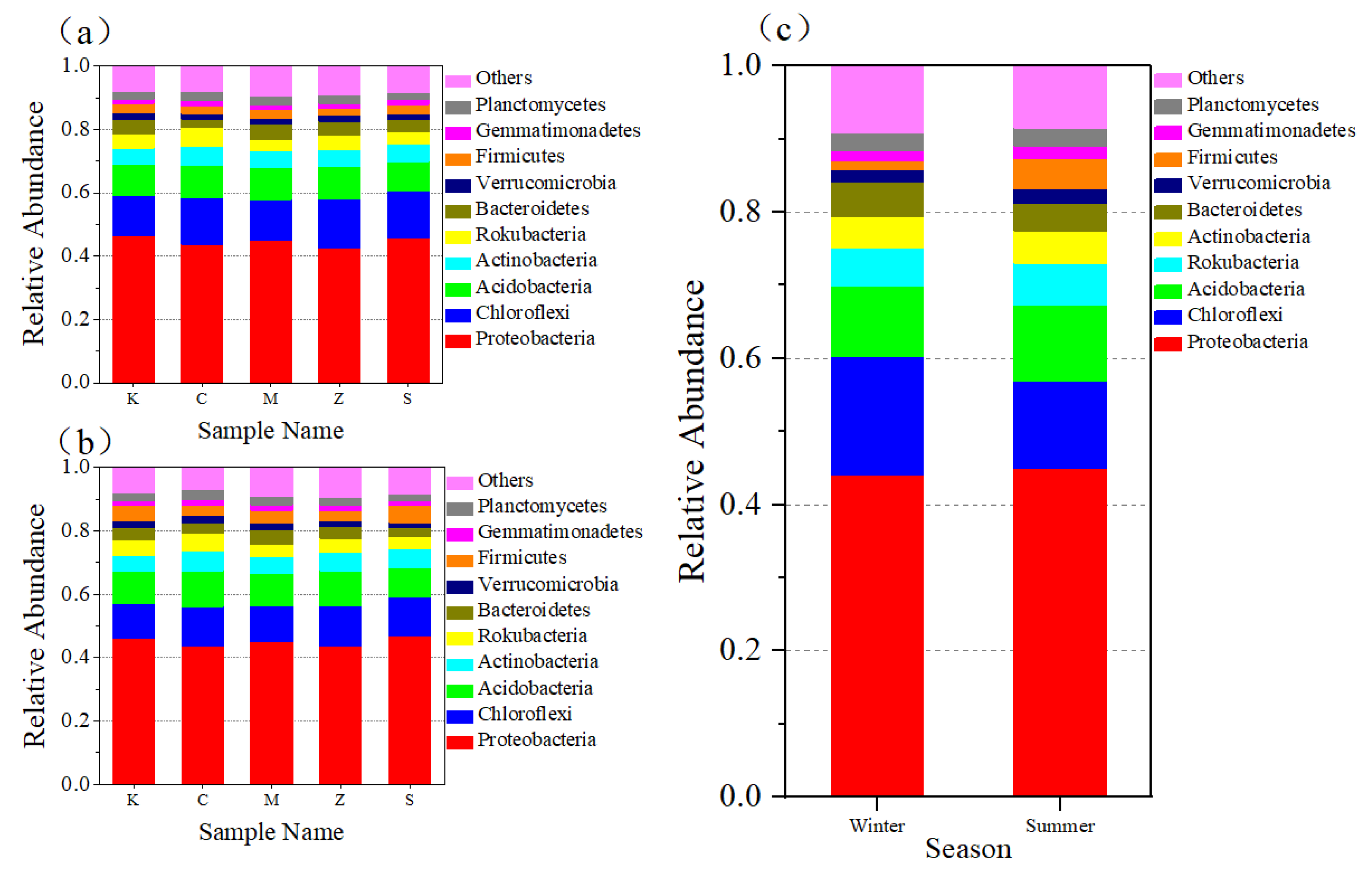
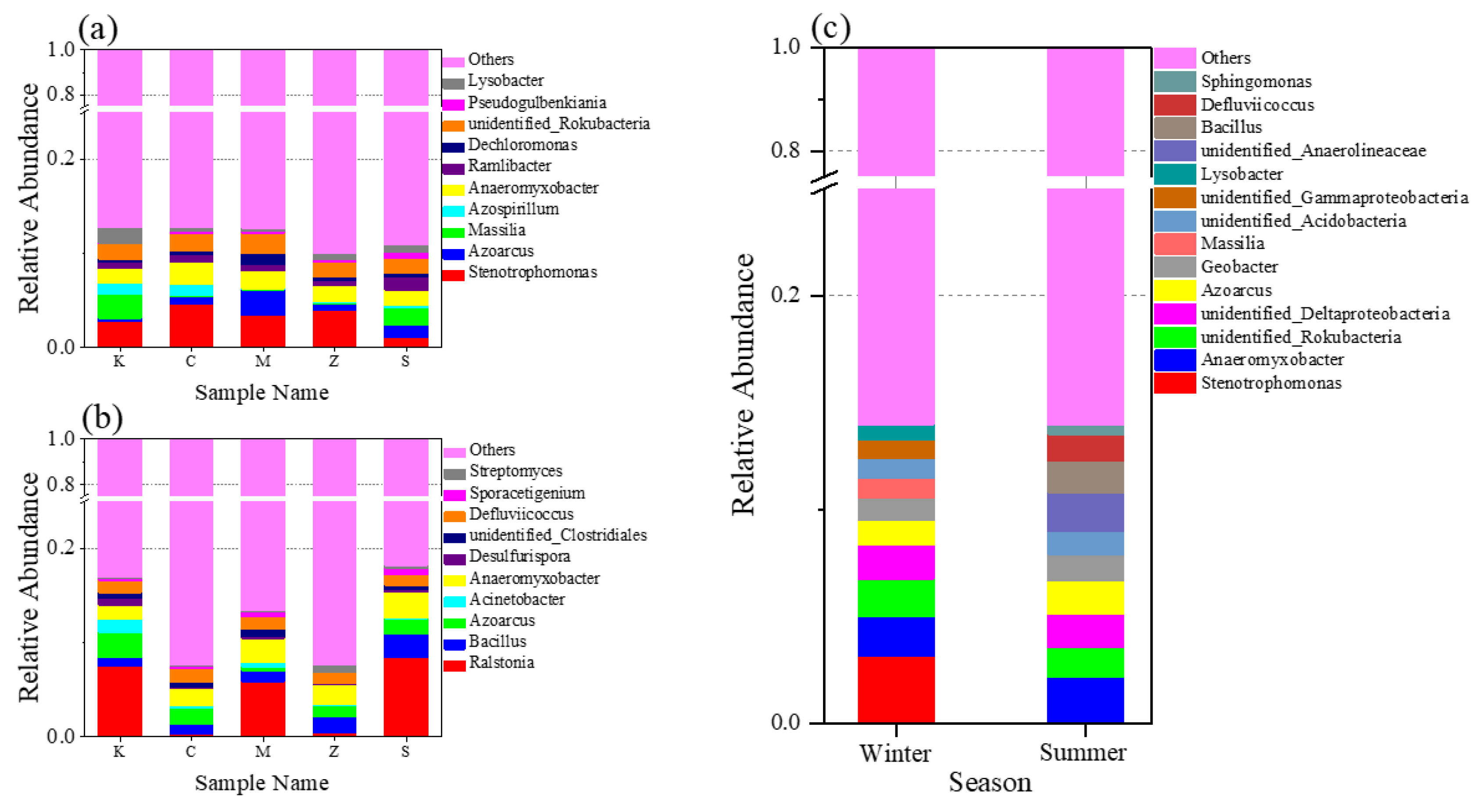
3.2.2. Bacterial Community at the Phylum Level in Both Seasons and in Different Plants
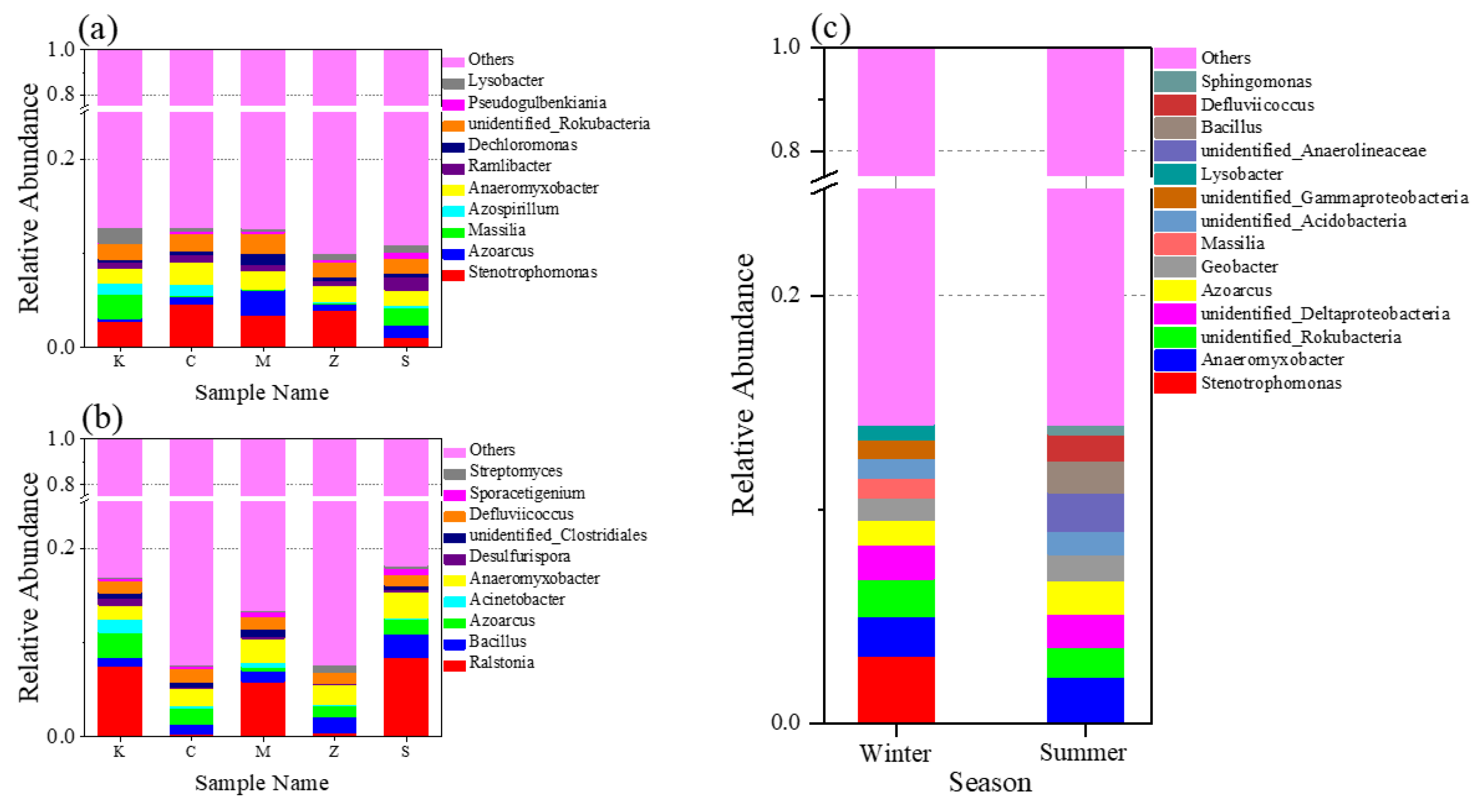
3.2.3. Bacterial Community at the Genus Level in Both Seasons and in Different Plants
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dong:, J.; Gao, H.; Wang, S.; Yao, H.; Ma, M. Simulation of the transfer and fate of HCHs since the 1950s in Lanzhou, China. Ecotoxicol. Environ. Saf. 2009, 72, 1950–1956. [Google Scholar] [CrossRef]
- Tian, C.; Ma, J.; Liu, L.; Jia, H.; Xu, D.; Li, Y.-F. A modeling assessment of association between East Asian summer monsoon and fate/outflow of α-HCH in Northeast Asia. Atmos. Environ. 2009, 43, 3891–3901. [Google Scholar] [CrossRef]
- Wu, Z.; Lin, T.; Hu, L.; Li, Y.; Guo, Z. Semi-centennial sediment records of HCHs and DDTs from the East China marginal seas: Role of lateral transport in catchment. Chemosphere 2021, 263, 128100. [Google Scholar] [CrossRef]
- Vijgen, J.; de Borst, B.; Weber, R.; Stobiecki, T.; Forter, M. HCH and lindane contaminated sites: European and global need for a permanent solution for a long-time neglected issue. Environ. Pollut. 2019, 248, 696–705. [Google Scholar] [CrossRef]
- Wu, X.; Chen, A.; Yuan, Z.; Kang, H.; Xie, Z. Atmospheric organochlorine pesticides (OCPs) and polychlorinated biphenyls (PCBs) in the Antarctic marginal seas: Distribution, sources and transportation. Chemosphere 2020, 258, 127359. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Shivanandappa, T. Stereospecificity in the cytotoxic action of hexachlorocyclohexane isomers. Chem. Biol. Interact. 2010, 183, 34–39. [Google Scholar] [CrossRef]
- Bhatt, P.; Kumar, M.S.; Chakrabarti, T. Fate and Degradation of POP-Hexachlorocyclohexane. Crit. Rev. Environ. Sci. Technol. 2009, 39, 655–695. [Google Scholar] [CrossRef]
- Srivastava, V.; Srivastava, T.; Kumar, M.S. Fate of the persistent organic pollutant (POP)Hexachlorocyclohexane (HCH) and remediation challenges. Int. Biodeterior. Biodegrad. 2019, 140, 43–56. [Google Scholar] [CrossRef]
- Kumari, K.; Tripathy, J.; Mohapatra, P.; Verma, S.; Das, B.; Raina, V.; Ray, L. Evaluation of toxicity of HCH isomers and its degradation metabolites on mammalian cell line and zebra fish embryos. New Biotechnol. 2018, 44, S151. [Google Scholar] [CrossRef]
- Hernandez, R.; Lash, J.P.; Burrows, B.; Wilund, K.R.; Mattix-Kramer, H.J.; Peralta, C.; Durazo-Arvizu, R.A.; Talavera, G.A.; Penedo, F.J.; Khambaty, T.; et al. The association of positive affect and cardiovascular health in Hispanics/Latinos with chronic kidney disease: Results from the Hispanic Community Health Study/Study of Latinos (HCHS/SOL). Prev. Med. Rep. 2019, 15, 100916. [Google Scholar] [CrossRef] [PubMed]
- Shah, H.K.; Sharma, T.; Banerjee, B.D. Organochlorine pesticides induce inflammation, ROS production, and DNA damage in human epithelial ovary cells: An in vitro study. Chemosphere 2020, 246, 125691. [Google Scholar] [CrossRef]
- Vijgen, J.; Abhilash, P.C.; Li, Y.F.; Lal, R.; Forter, M.; Torres, J.; Singh, N.; Yunus, M.; Tian, C.; Schaffer, A.; et al. Hexachlorocyclohexane (HCH) as new Stockholm Convention POPs--a global perspective on the management of Lindane and its waste isomers. Environ. Sci. Pollut. Res. Int. 2011, 18, 152–162. [Google Scholar] [CrossRef]
- Liu, F.F.; Fan, J.; Du, J.; Shi, X.; Zhang, J.; Shen, Y. Intensified nitrogen transformation in intermittently aerated constructed wetlands: Removal pathways and microbial response mechanism. Sci. Total Environ. 2019, 650 Pt 2, 2880–2887. [Google Scholar] [CrossRef]
- Cryder, Z.; Wolf, D.; Carlan, C.; Gan, J. Removal of urban-use insecticides in a large-scale constructed wetland. Environ. Pollut. 2020, 268 Pt A, 115586. [Google Scholar] [CrossRef]
- Hickey, A.; Arnscheidt, J.; Joyce, E.; O’Toole, J.; Galvin, G.; M, O.C.; Conroy, K.; Killian, D.; Shryane, T.; Hughes, F.; et al. An assessment of the performance of municipal constructed wetlands in Ireland. J. Environ. Manag. 2018, 210, 263–272. [Google Scholar] [CrossRef]
- Irwin, N.B.; Irwin, E.G.; Martin, J.F.; Aracena, P. Constructed wetlands for water quality improvements: Benefit transfer analysis from Ohio. J. Environ. Manag. 2018, 206, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Burgos, V.; Araya, F.; Reyes-Contreras, C.; Vera, I.; Vidal, G. Performance of ornamental plants in mesocosm subsurface constructed wetlands under different organic sewage loading. Ecol. Eng. 2017, 99, 246–255. [Google Scholar] [CrossRef]
- Tanner, C.C.; Nguyen, M.L.; Sukias, J.P.S. Nutrient removal by a constructed wetland treating subsurface drainage from grazed dairy pasture. Agric. Ecosyst. Environ. 2005, 105, 145–162. [Google Scholar] [CrossRef]
- Park, J.; Cho, K.H.; Lee, E.; Lee, S.; Cho, J. Sorption of pharmaceuticals to soil organic matter in a constructed wetland by electrostatic interaction. Sci. Total Environ. 2018, 635, 1345–1350. [Google Scholar] [CrossRef] [PubMed]
- Stottmeister, U.; Wiessner, A.; Kuschk, P.; Kappelmeyer, U.; Kastner, M.; Bederski, O.; Muller, R.A.; Moormann, H. Effects of plants and microorganisms in constructed wetlands for wastewater treatment. Biotechnol. Adv. 2003, 22, 93–117. [Google Scholar] [CrossRef]
- Yan, Q.; Xu, Y.; Yu, Y.; Zhu, Z.W.; Feng, G. Effects of pharmaceuticals on microbial communities and activity of soil enzymes in mesocosm-scale constructed wetlands. Chemosphere 2018, 212, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, X.; He, X.; Zhang, S.; Liang, R.; Shen, J. Effects of root exudates on denitrifier gene abundance, community structure and activity in a micro-polluted constructed wetland. Sci. Total Environ. 2017, 598, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-J.; Tian, Y.-H.; Zhang, Y.; Song, B.-R.; Li, H.-C.; Chen, Z.-H. Effects of root organic exudates on rhizosphere microbes and nutrient removal in the constructed wetlands. Ecol. Eng. 2016, 92, 243–250. [Google Scholar] [CrossRef]
- Abdolali, A.; Guo, W.S.; Ngo, H.H.; Chen, S.S.; Nguyen, N.C.; Tung, K.L. Typical lignocellulosic wastes and by-products for biosorption process in water and wastewater treatment: A critical review. Bioresour. Technol. 2014, 160, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Tursi, A.; Chidichimo, F.; Bagetta, R.; Beneduci, A. BTX removal from open aqueous systems by modified cellulose fibers and evaluation of competitive evaporation kinetics. Water 2020, 12, 3415. [Google Scholar] [CrossRef]
- Wojciechowska, E. Removal of persistent organic pollutants from landfill leachates treated in three constructed wetland systems. Water Sci. Technol. 2013, 68, 1164. [Google Scholar] [CrossRef]
- Tursi, A.; Chatzisymeon, E.; Chidichimo, F.; Beneduci, A.; Chidichimo, G. Removal of endocrine disrupting chemicals from water: Adsorption of bisphenol-A by biobased hydrophobic functionalized cellulose. Int. J. Environ. Res. Public Health 2018, 15, 2419. [Google Scholar] [CrossRef] [Green Version]
- Tursi, A.; Beneduci, A.; Chidichimo, F.; Vietro, N.D.; Chidichimo, G. Remediation of hydrocarbons polluted water by hydrophobic functionalized cellulose. Chemosphere 2018, 201, 530–539. [Google Scholar] [CrossRef]
- Guo, W.; Zhang, H.; Huo, S. Organochlorine pesticides in aquatic hydrophyte tissues and surrounding sediments in Baiyangdian wetland, China. Ecol. Eng. 2014, 67, 150–155. [Google Scholar] [CrossRef]
- Tang, X.; Yang, Y.; Tao, R.; Chen, P.; Dai, Y.; Jin, C.; Feng, X. Fate of mixed pesticides in an integrated recirculating constructed wetland (IRCW). Sci. Total Environ. 2016, 571, 935–942. [Google Scholar] [CrossRef]
- Dordio, A.V.; Carvalho, A.J. Organic xenobiotics removal in constructed wetlands, with emphasis on the importance of the support matrix. J. Hazard. Mater. 2013, 252–253, 272–292. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.Q.; Gersberg, R.M.; Zhu, J.; Hua, T.; Jinadasa, K.B.; Tan, S.K. Batch versus continuous feeding strategies for pharmaceutical removal by subsurface flow constructed wetland. Environ. Pollut. 2012, 167, 124–131. [Google Scholar] [CrossRef]
- Torralba-Sanchez, T.L.; Kuo, D.T.F.; Allen, H.E.; Di Toro, D.M. Bioconcentration factors and plant-water partition coefficients of munitions compounds in barley. Chemosphere 2017, 189, 538–546. [Google Scholar] [CrossRef]
- Vymazal, J.; Brezinova, T. The use of constructed wetlands for removal of pesticides from agricultural runoff and drainage: A review. Environ. Int. 2015, 75, 11–20. [Google Scholar] [CrossRef]
- Romeh, A.A. Phytoremediation of water and soil contaminated with imidacloprid pesticide by Plantago Major, L. Int. J. Phytoremediat. 2010, 12, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Meng, P.; Pei, H.; Hu, W.; Shao, Y.; Li, Z. How to increase microbial degradation in constructed wetlands: Influencing factors and improvement measures. Bioresour. Technol. 2014, 157, 316–326. [Google Scholar] [CrossRef]
- Truu, M.; Juhanson, J.; Truu, J. Microbial biomass, activity and community composition in constructed wetlands. Sci. Total Environ. 2009, 407, 3958–3971. [Google Scholar] [CrossRef]
- Balázs, H.E.; Schmid, C.A.O.; Podar, D.; Hufnagel, G.; Radl, V.; Schröder, P. Development of microbial communities in organochlorine pesticide contaminated soil: A post-reclamation perspective. Appl. Soil Ecol. 2020, 150, 103467. [Google Scholar] [CrossRef]
- Al-Ani, M.A.M.; Hmoshi, R.M.; Kanaan, I.A.; Thanoon, A.A. Effect of pesticides on soil microorganisms. J. Phys. Conf. Ser. 2019, 1294, 072007. [Google Scholar] [CrossRef]
- Wołejko, E.; Jabłońska-Trypuć, A.; Wydro, U.; Butarewicz, A.; Łozowicka, B. Soil biological activity as an indicator of soil pollution with pesticides—A review. Appl. Soil Ecol. 2020, 147, 103356. [Google Scholar] [CrossRef]
- Rangasamy, K.; Athiappan, M.; Devarajan, N.; Samykannu, G.; Parray, J.A.; Aruljothi, K.N.; Shameem, N.; Alqarawi, A.A.; Hashem, A.; Abd Allah, E.F. Pesticide degrading natural multidrug resistance bacterial flora. Microb. Pathog. 2018, 114, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Behera, P.; Mohapatra, M.; Adhya, T.K.; Suar, M.; Pattnaik, A.K.; Rastogi, G. Structural and metabolic diversity of rhizosphere microbial communities of Phragmites karka in a tropical coastal lagoon. Appl. Soil Ecol. 2018, 125, 202–212. [Google Scholar] [CrossRef]
- Ansola, G.; Arroyo, P.; Saenz de Miera, L.E. Characterisation of the soil bacterial community structure and composition of natural and constructed wetlands. Sci. Total Environ. 2014, 473–474, 63–71. [Google Scholar] [CrossRef]
- Regar, R.K.; Gaur, V.K.; Bajaj, A.; Tambat, S.; Manickam, N. Comparative microbiome analysis of two different long-term pesticide contaminated soils revealed the anthropogenic influence on functional potential of microbial communities. Sci. Total Environ. 2019, 681, 413–423. [Google Scholar] [CrossRef]
- Benimeli, C.S.; Amoroso, M.J.; Chaile, A.P.; Castro, G.R. Isolation of four aquatic streptomycetes strains capable of growth on organochlorine pesticides. Bioresour. Technol. 2003, 89, 133–138. [Google Scholar] [CrossRef]
- Fuentes, M.S.; Benimeli, C.S.; Cuozzo, S.A.; Amoroso, M.J. Isolation of pesticide-degrading actinomycetes from a contaminated site: Bacterial growth, removal and dechlorination of organochlorine pesticides. Int. Biodeterior. Biodegrad. 2010, 64, 434–441. [Google Scholar] [CrossRef]
- Sineli, P.E.; Tortella, G.; Dávila Costa, J.S.; Benimeli, C.S.; Cuozzo, S.A. Evidence of α-, β- and γ-HCH mixture aerobic degradation by the native actinobacteria Streptomyces sp. M7. World J. Microbiol. Biotechnol. 2016, 32, 81. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xu, L.; Huang, T.; Yan, M.; Liu, K.; Miao, Y.; He, H.; Li, S.; Sekar, R. Combined effects of seasonality and stagnation on tap water quality: Changes in chemical parameters, metabolic activity and co-existence in bacterial community. J. Hazard. Mater. 2021, 403, 124018. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Yi, H.; Chen, S.; Zhang, Z.; Cui, K.; Bing, Y.; Zhuo, Q.; Li, B.; Xie, S.; Guo, Q. Influences of plant type on bacterial and archaeal communities in constructed wetland treating polluted river water. Environ. Sci. Pollut. Res. Int. 2016, 23, 19570–19579. [Google Scholar] [CrossRef]
- Sharma, M.; Khurana, H.; Singh, D.N.; Negi, R.K. The genus Sphingopyxis: Systematics, ecology, and bioremediation potential-A review. J. Environ. Manag. 2021, 280, 111744. [Google Scholar] [CrossRef]
- Xu, Y.; Niu, L.; Qiu, J.; Zhou, Y.; Lu, H.; Liu, W. Stereoselective accumulations of hexachlorocyclohexanes (HCHs) are correlated with Sphingomonas spp. in agricultural soils across China. Environ. Pollut. 2018, 240, 27–33. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | OTU | Shannon | Chao1 | Ace | Goods Coverage |
|---|---|---|---|---|---|
| Winter 1 | 3143 | 9.151 | 3534 | 3577 | 0.986 |
| Winter 15 | 3114 | 9.319 | 3830 | 3632 | 0.985 |
| Winter 35 | 3088 | 8.913 | 3447 | 3500 | 0.987 |
| Winter 60 | 3173 | 9.240 | 3545 | 3584 | 0.987 |
| Summer 1 | 3039 | 8.802 | 3516 | 3636 | 0.981 |
| Summer 15 | 3255 | 9.297 | 3899 | 4006 | 0.978 |
| Summer 35 | 3453 | 9.721 | 4242 | 4332 | 0.975 |
| Summer 60 | 3343 | 9.547 | 4429 | 4347 | 0.975 |
| Sample Name | OTU | Shannon | Chao1 | Ace | Goods Coverage |
|---|---|---|---|---|---|
| K | 3145 | 9.199 | 3984 | 3793 | 0.982 |
| C | 3293 | 9.514 | 4480 | 4048 | 0.980 |
| M | 3301 | 9.790 | 4023 | 3933 | 0.981 |
| Z | 3284 | 9.356 | 4025 | 3929 | 0.981 |
| S | 3097 | 9.143 | 3954 | 3781 | 0.981 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Q.; Zeng, H.; Liang, Y.; Qin, L.; Peng, G.; Huang, L.; Song, X. Purification Effects on β-HCH Removal and Bacterial Community Differences of Vertical-Flow Constructed Wetlands with Different Vegetation Plantations. Sustainability 2021, 13, 13244. https://doi.org/10.3390/su132313244
Chen Q, Zeng H, Liang Y, Qin L, Peng G, Huang L, Song X. Purification Effects on β-HCH Removal and Bacterial Community Differences of Vertical-Flow Constructed Wetlands with Different Vegetation Plantations. Sustainability. 2021; 13(23):13244. https://doi.org/10.3390/su132313244
Chicago/Turabian StyleChen, Qing, Honghu Zeng, Yanpeng Liang, Litang Qin, Guangsheng Peng, Liangliang Huang, and Xiaohong Song. 2021. "Purification Effects on β-HCH Removal and Bacterial Community Differences of Vertical-Flow Constructed Wetlands with Different Vegetation Plantations" Sustainability 13, no. 23: 13244. https://doi.org/10.3390/su132313244
APA StyleChen, Q., Zeng, H., Liang, Y., Qin, L., Peng, G., Huang, L., & Song, X. (2021). Purification Effects on β-HCH Removal and Bacterial Community Differences of Vertical-Flow Constructed Wetlands with Different Vegetation Plantations. Sustainability, 13(23), 13244. https://doi.org/10.3390/su132313244