Abstract
Although mycorrhizae applications have been widely used to improve the establishment and growth of agricultural crops, there have been no studies on their application in field-grown nectarine trees. In this work, a commercial arbuscular mycorrhizal fungus (AMF; Glomus iranicum var. tenuihypharum) was applied by means of fertigation to inoculate young “Flariba” nectarine trees grown in south-eastern Spain to evaluate its effect on plant water status, and vegetative and reproductive growth. Using minirhizotrons to measure the root dynamics over a complete growing season, revealed that AMF substantially increased root growth (51% increase compared with untreated trees), while no changes in plant water status or canopy development were noted. The productive response improved in inoculated trees, as demonstrated by a significantly higher yield, fruit size, number of fruits per tree and greater crop load efficiency values than in untreated trees. Given that the same amount of irrigation solution was applied in both treatments, the irrigation water use efficiency increased by 19.5% in AMF compared with untreated trees. The findings of this study suggest that a simple inoculation of AMF can be considered a good practice in semi-arid agro-systems to firmly establish efficient young nectarine trees and enhance their adaptation to field conditions.
1. Introduction
Nectarines and peaches (Prunus persica L. Batsch) are, jointly, the fourth most important fruit crop in the world, and in Spain, both Prunus species are important crops for export to European markets [1]. Indeed, Spain is the second largest producer in the Mediterranean Basin after Italy, with an average annual production of 1854 Mt in the period 2014–2018 [2]. However, the water scarcity faced by the Mediterranean area means that water-conserving irrigation techniques need to be improved [3,4,5].
While advances in irrigation and fertilization technologies have optimized water and nutrient consumption in general, it is the morphology and architecture of the root system that determine the ability of plants to take up water and nutrients [6], affecting the shoot growth rate and the relative size of canopies [7]. This underlines the importance of promoting the development of an efficient root system that encourages sufficient mineral nutrition and a suitable water balance [8].
Among the options proposed in recent years to favor the functioning of the root system is the use of arbuscular mycorrhizal fungi (AMF), soil microorganisms that have been seen to establish a symbiotic relationship with 80–90% of vascular plant species and 90% of agricultural plant species [9], including most cereals, vegetables, and horticultural plants.
AMF can improve plant establishment and host plant performance by transferring nutrients and increasing root water absorption [10]. According to Bucher et al. [11], effective symbiosis allows the plants to explore a greater volume of soil. Moreover, mycorrhizal fungi produce plant hormones and changes in root exudate composition [12] that improves the structure and quality of soils and promote better rooting [9].
Several studies have shown that inoculation with AMF positively affects several aspects of plant life, e.g., nutrition, growth, stress tolerance, and disease resistance [13,14,15,16,17], although, the exact response of mycorrhizal plants depends on the plant species and the fungus used [18] and also the kind of stress the plants must confront [19].
The alleviating effect of AMF symbiosis on the plant response to water scarcity in Mediterranean areas is normally a result of the positive effects that the AMF have on the uptake and transport of water and nutrients, thus ensuring the adequate hydration of plant tissues, increasing the photosynthetic capacity, and promoting growth [10,20,21,22,23]. Furthermore, Zheng et al. [24] reported that AMF application protected the plant against water stress caused by flooding through the accumulation of proline, which is also associated with an improved root architecture.
Since the root system plays such a crucial role in plant development, it is essential to understand its dynamics and the competition that exists with the aerial part. However, studying the roots under field conditions is difficult because the soil limits their accessibility for observation and because the methods proposed to date are tedious, requiring careful counts and frequent measurements at the same site [25,26]. In this respect, the glass walls method allows root growth to be observed in situ, the minirhizotron technique—a modification of this method—being particularly well suited to studying rooting in natural conditions [26,27]. The possibility of using this technique, coupled with electronic equipment for the acquisition of images from inside the tube and image analysis technology, has greatly facilitated the procedure for undertaking the in situ measurements of roots and recording their growth in order to evaluate root length density (RLD), which acts as an indicator of the ability of plants to absorb soil water [10,28]. New software for capturing and displaying root images has been developed to study root distribution and root dynamics in annual plants [29,30] and fruit trees [21,26,31,32,33,34] but few studies have attempted to evaluate the response of plant roots to different soil management practices.
Among other things, this means that the plant root relationship with mycorrhizal colonization, which, to date, has mostly been carried out by means of destructive methods, needs further study. The main objective of this study was to investigate the effects of mycorrhizal colonization on the vegetative growth of a young nectarine tree orchard, using the minirhizotron technique and with special emphasis on root dynamics. Fruit growth, yield, and plant water status were also evaluated throughout a complete growing season. The potential effect of AMF as a cultural practice for enhancing the in-field establishment of young nectarine trees is discussed.
2. Materials and Methods
2.1. Experimental Conditions
The experiment was performed at the CEBAS-CSIC experimental field station in Santomera, Murcia, Spain (38°06′ N, 1°02′ W, 110 m altitude) over one complete growing season (from May to May the following year). The study material consisted of a 0.5 ha plot of two-year old early-maturing nectarine trees (Prunus persica L. Batsch, cv. “Flariba”) grafted on GxN-15 rootstock, trained to an open-center canopy and spaced 6.5 m × 3.5 m. The phenological stages of this cultivar are depicted in [35]. Briefly, flowering started at the end of January, followed by leaf development before harvesting in early-May.
The soil, classified as Lithic xeric haploxeroll, is stony and shallow, with a clay-loam texture (clay fraction: 41% illite, 17% smectite, and 30% palygorskite). It has a low organic matter content (1.30%), and a cationic exchange capacity of 97.9 mmol kg−1. The available soil water content was 140 mm m−1 and bulk density was 1430 kg m−3. The volumetric soil water content at field capacity and wilting point were 0.29 and 0.14 m3 m−3, respectively.
Meteorological data (air temperature, relative humidity, solar radiation, wind speed and rainfall) were recorded by an automatic weather station located at the CEBAS-CSIC experimental station (http://www.cebas.csic.es/general_spain/est_meteo.html (accessed on 20 May 2012)), which reads values every 5 min and records the average every 15 min. Reference crop evapotranspiration (ET0) and vapor pressure deficit (VPD) were calculated on a daily basis.
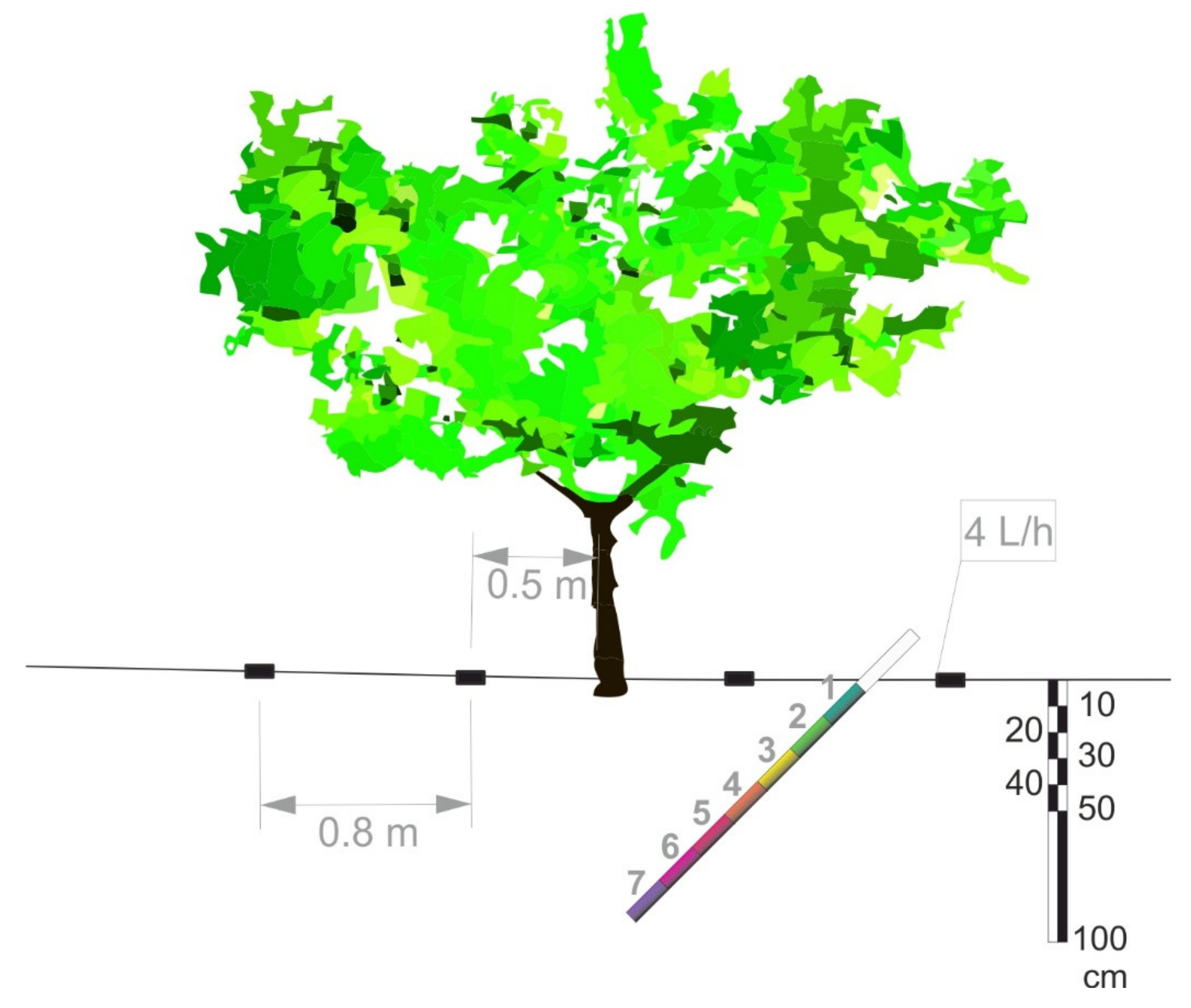
Crop management (including pest control) was that commonly carried out in commercial fruit orchards of the area. The soil was not tilled and weeds were eliminated by chemical treatment. Trees were drip irrigated using one drip-line per tree row with four pressure-compensated emitters (4 L h−1) per tree located 0.5 and 1.3 m from the tree trunk at both sides of the tree (Figure 1). Irrigation was scheduled weekly to satisfy maximum (100%) crop evapotranspiration (ETc) requirements, which was estimated by multiplying daily reference crop evapotranspiration (ET0), calculated using the Penman-Monteith equation [36], by the locally determined crop coefficients (Kc) for early-maturing Prunus sp. [37], corrected by the plant cover factor (Kr) for young fruit trees [38]. Irrigation water (from the Tagus-Segura transfer system), with an average electrical conductivity (EC25 °C) of 1.3 dS m−1, was applied during the night, as needed. The annual amount of irrigation water applied, measured with in-line water meters, was 4430 m3 ha−1. Seasonal fertilizer applications were 83–56–109 kg ha−1 of N, P2O5 and K2O, respectively, applied through the drip irrigation system.
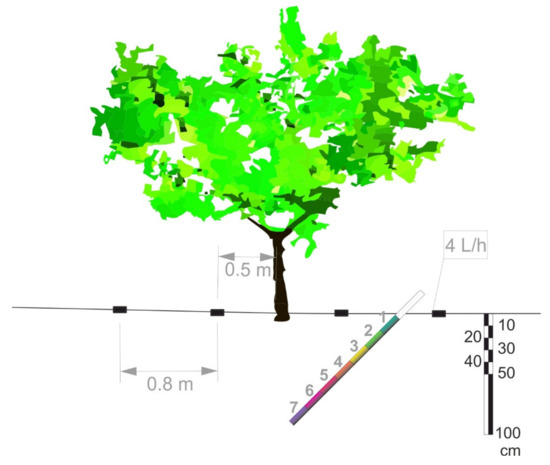
Figure 1.
Relative position of 1.8 m long minirhizotron methacrylate transparent tube (inner diameter 0.06 m) with respect to nectarine tree trunk, drip irrigation line (black rectangles indicate the four emitters) in the 0–100 cm soil profile.
2.2. Treatments
Two treatments were considered in this study: one involving inoculation with arbuscular mycorrhizal fungus (hereafter, AMF treatment) and one control (hereafter, untreated treatment). The commercial AMF, consisted of Glomus iranicum var. tenuihypharum (a mixture of spores, mycorrhizal root fragments and rhizospheric soil) [39], and was applied in spring (20 April 2011) through the irrigation system at a rate of 3 kg ha−1.
Treatments were arranged in a completely randomized design with six replications, each consisting of six trees. The central four were used for measurements and the rest were considered as border trees.
2.3. Measurements
2.3.1. Plant Water Status
Plant water status was evaluated every 7–14 days throughout the growing season by measuring midday stem water potential (Ψstem, MPa) using a pressure chamber (Soil Moisture Equip. Corp. Model 3000, 153 Santa Barbara, CA, USA). Measurements were taken at around 12 h solar time in fully expanded leaves selected from near the tree trunk in the north-facing part of the tree. The leaves were covered with aluminum foil bags for at least 2 h prior to the measurements, following the recommendations of Hsiao [40]. Six leaves, one per tree and replicate, from both treatments were used.
2.3.2. Root Measurements
Root dynamics was evaluated using the minirhizotron method. For this, transparent plastic tubes were installed in December 2010, at a 45° angle, using an adapted hydraulic soil-coring machine, 0.9 m from the tree trunk, midway between the two emitters (Figure 1), on one representative tree in four out of the six replications (n = 4) in both treatments.
The tubes were 1.8 m long with outer and inner diameters of 70 and 64 mm, respectively. The total length of each buried tube was 1.4 m, so that it reached a total depth of 1 m (Figure 1). The part of the tube protruding from the soil surface was covered with isolating material to prevent light from entering the tube and the tube from becoming heated.
Root images (0.26 m × 0.20 m) were captured twice per month, starting on May 2011, using a CI-600 Root Growth Scanning System, which consisted of a rotating, linear scan head connected to a laptop computer. Linear and non-distorted scanned color images captured with the system were analyzed using the CI-400CIAS Computer Imaging Analysis Software provided with the WinRHIZOTM Tron software v.2008 (Regent Instruments Inc., Quebec, QC, Canada).
The number of root points, and length and diameter of roots were determined at seven depths: 0–10, 10–25, 25–40, 40–55, 55–70, 70–85, and 85–100 cm) (Figure 1). Roots were classified into three diameter classes: very fine (<0.5 mm), fine (0.5–2.0 mm), and coarse (>2 mm).
Root length density (RLD) was calculated according to the formula proposed by Upchurch [28], based on Newman’s line intersection method, which uses the number of root points which intersect the minirhizotron tube within the view frame:
where NR is the number of roots, A is the minirhizotron frame area observed by the scanner (439.82 cm2), and d is the outer diameter of the minirhizotron tube (7 cm). This method calculates the expected value of root length within the soil volume occupied by the tube, as if the tube were not present [41]. RLD was expressed as total root length per unit of sampled soil volume (cm of root per cm3 of soil).
RLD = (NR · d)/(A · d)
2.3.3. Aboveground Measurements
Fruits
Weekly from early March until harvest, the equatorial fruit diameter was measured in 100 fruits randomly selected from the four experimental trees of each replication (n = 6) of each treatment, using a digital caliper (0–150 ± 0.01 mm; Mitutoyo, CD-15D, Kawasaki, Japan).
Nectarine fruits from the untreated border trees were sampled frequently during the fruit growing season, making the following measurements in individual fruits (n = 120): suture, cheek-to-cheek (equatorial) and longitudinal (height) diameters measured with a digital caliper, fresh mass with a scales (Mettler PE 360 DeltaRange, Greifensee, Switzerland, with an accuracy of ± 0.001 g), and dry mass, after 24–48 h at 60 °C in a ventilated oven (Model 2005142, J.P. Selecta, Madrid, Spain).
Vegetative Growth
The shoot length was measured twice monthly on four tagged shoots per tree, one from each compass direction, on six trees (one tree per replication, n = 6) of each treatment from March to November, using a tape measure.
Trunk diameter was measured on a monthly basis with a forest caliper (Codimex-C 100 cm, Warszawa, Poland in all experimental trees, at a marked location 0.3 m from the soil surface. Trunk cross-sectional area (TCSA) was estimated as being equivalent of a circle.
Growth rates for root, shoot, trunk and fruits were calculated as increase in length or diameter per day and also as percentage of total/annual growth.
Pruning dry mass was determined in six trees, one per replicate and treatment (n = 6). Winter pruning was determined during dormancy, eliminating the shoots to maintain the vase tree architecture, and summer pruning was determined just after harvest, eliminating the water sprouts. The eliminated shoots were weighed in the field and samples were taken to the lab to obtain dry mass, after 48 h 65 °C in a ventilated oven.
Canopy tree cover was estimated in summer with zenithal images, which were analyzed following the procedure indicated in Conesa et al. [35].
Plant biomass partitioning, i.e., above-ground (including fruits) and root portions [42], was estimated as dry weight mass contribution, following Carvajal et al. [43].
Yield, Fruit Quality and Efficiencies at Harvest
Yield at harvest was evaluated on two picking dates (2 and 9 May 2012), weighing on a scales (0–6000 ± 2 g, Scaltec, Model SSH 92, Dania Beach, FL, U.S.A.) and counting the total number of fruits per tree in all the experimental trees. A sample of 10 fruits from each replication at the two picking dates was taken to the lab in insulated bags to measure the following: equatorial and longitudinal diameters, using a digital caliper; fresh mass, with a scales; total soluble solids, with a hand-held refractometer (Atago ATC-1, Tokyo, Japan), expressing the values as °Brix; and external fruit color, with a Konica Minolta Chroma Meter CR-10, Osaka, Japan). The results of the last parameter were expressed as CIEL*a*b* chromatic coordinates (L* = lightness, a* = red-green component), b* = blue-yellow component), from which values of chromaticity or Chrome (C*) and hue angle (h°) were calculated as:
C* = √(a*2 + b*2)
h° = arc tan (b*/a*)
Irrigation water use efficiency (IWUE, kg cm−3) was determined as the ratio between yield and total irrigation water applied. Production efficiency (PE, kg cm−2) was calculated as the ratio between yield and TCSA. Crop load efficiency (CLE, fruits cm−2) was determined as the ratio of number of fruits per tree to TCSA.
2.4. Statistical Analysis
Data were analyzed using the SPSS 20 (IBM, Armonk, NY, USA). Means were compared with the least significant difference test at a confidence level of 95% (LSD0.05).
3. Results
3.1. Meteorological Data and Plant Water Status
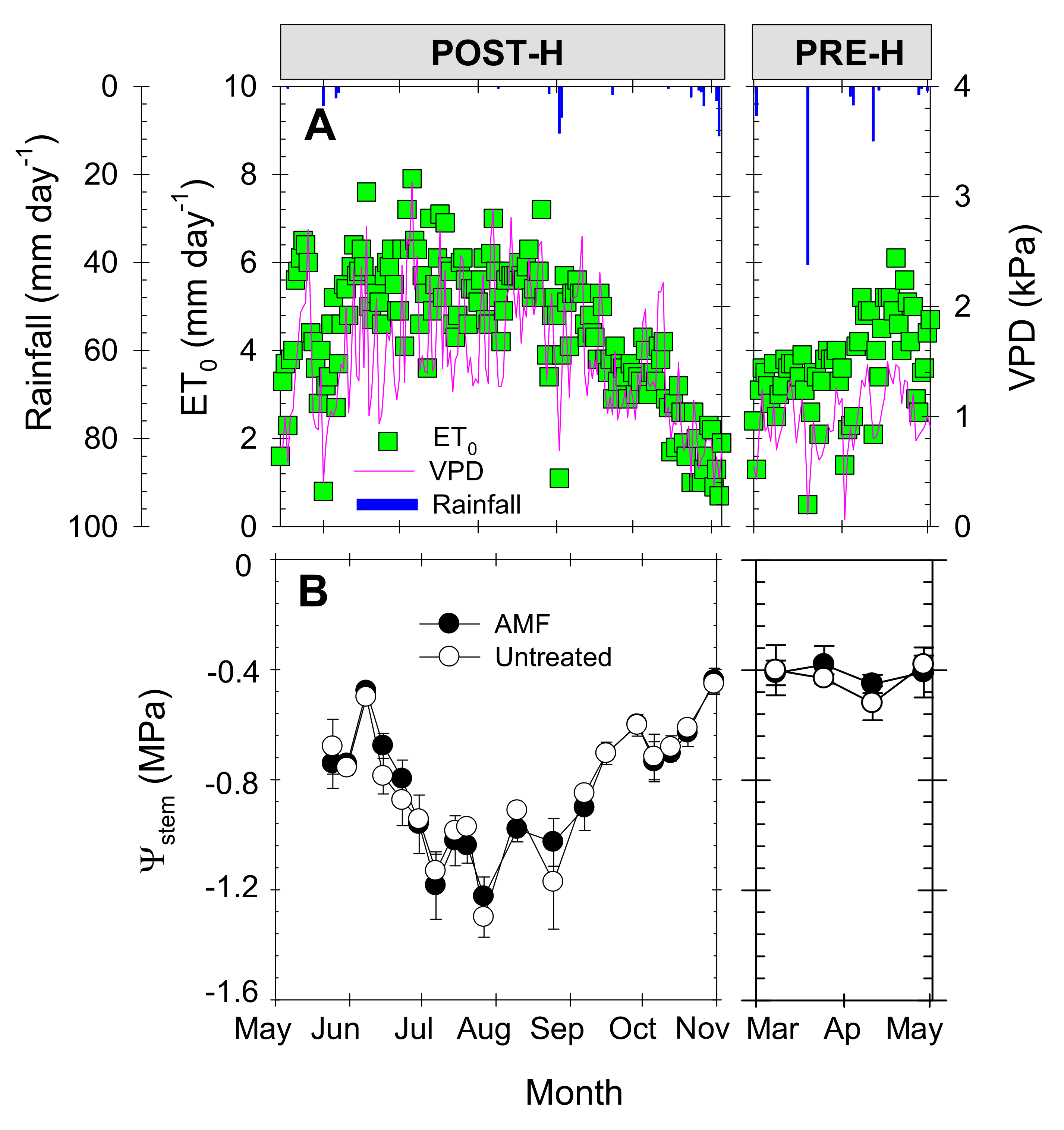
Agro-meteorological conditions of the study area are typical of a Mediterranean semi-arid climate, with hot dry summers (maximum air temperature 38.7 °C) and mild winters (average temperature 8 °C). Annual rainfall and reference crop evapotranspiration (ET0) values (≈250 and 1320 mm, respectively) were based on the average values of the five previous years. Total rainfall and ET0 for the studied growing season (2011/2012) were 246 and 1287 mm, respectively (Figure 2A). Daily ET0 registered the highest values in June (8 mm), and the lowest in November (0.2 mm). September was the wettest month with a total rainfall of 91 mm. Mean daily VPD values ranged from 3.1 kPa in July to 0.2 kPa in November 2011 (Figure 2A).
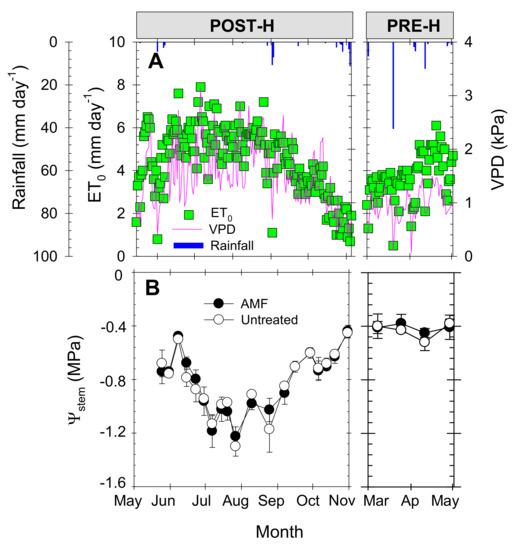
Figure 2.
Seasonal evolution of (A) daily reference crop evapotranspiration (ET0, green squares), rainfall (vertical blue bars), and vapor pressure deficit (VPD, purple line); and (B) midday stem water potential (Ψstem) in arbuscular mycorrhizal fungus (AMF)-inoculated (black circles) and untreated (white circles) young “Flariba” nectarine trees. Each point is the mean ± standard error (SE) of 6 leaves (one per replication). Phenological periods indicated by POST-H (post-harvest) and PRE-H (pre-harvest).
The seasonal trend of midday stem water potential (Ψstem) values exhibited a non-flat pattern, the lowest values of around −1.3 MPa being recorded during the summer (July), reflecting the period of greatest evaporative demand. As expected for early-maturing cultivars, the best plant water status was found during the pre-harvest period, when the, Ψstem value averaged −0.38 MPa (Figure 2B). Comparing the AMF and untreated young “Flariba” nectarine trees, no significant differences were observed in Ψstem at any time during the growing season, with mean values of −0.82 and −0.42 MPa during the post-harvest and pre-harvest periods, respectively.
3.2. Roots vs. Aerial Growth Pattern
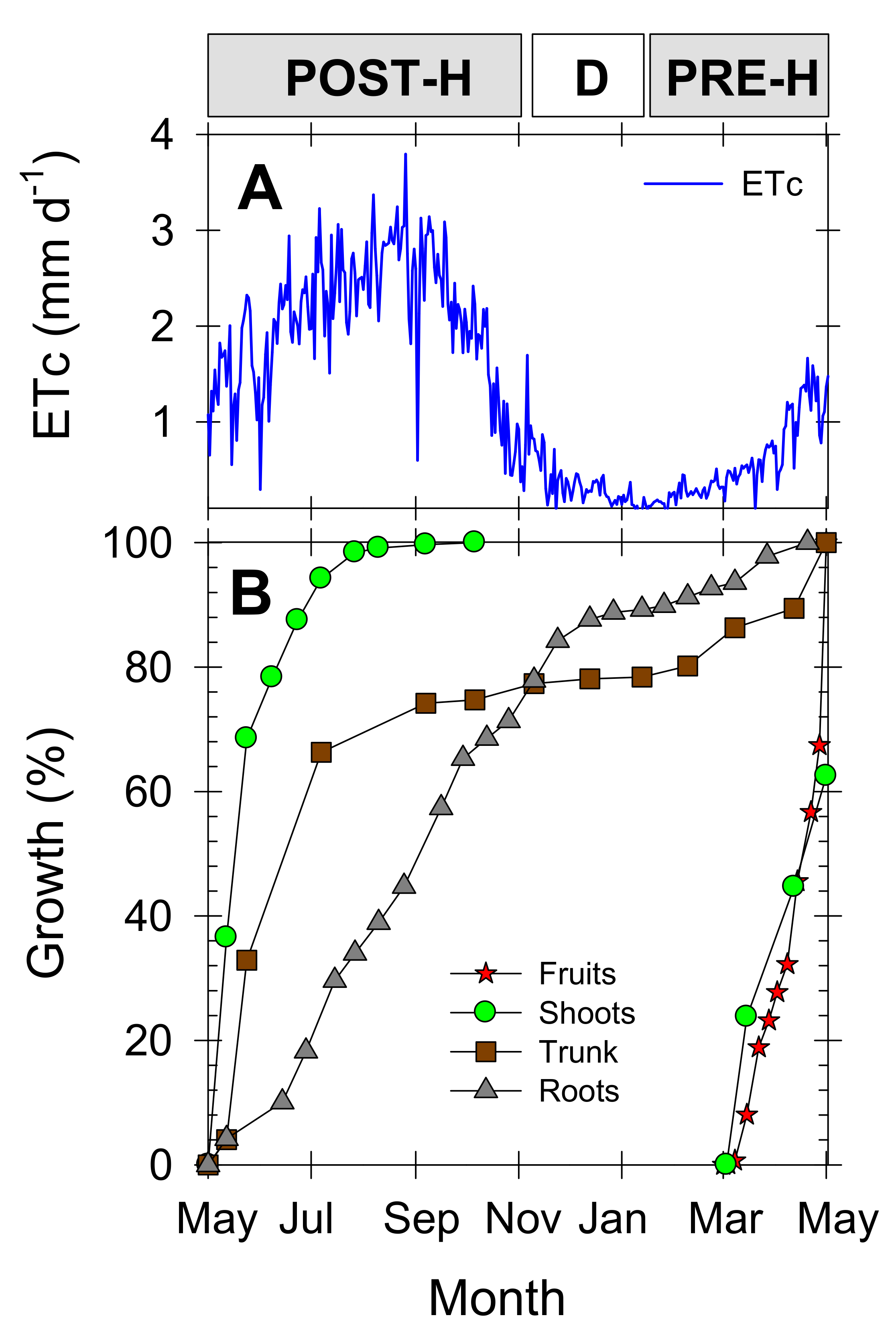
During the studied growing season, seasonal crop evapotranspiration (ETc) was 470 mm, coinciding with the total amount of irrigation water applied to the young “Flariba” nectarine trees (Figure 3A). Maximum daily ETc values were observed during the summer, decreasing from October onwards, and increasing again at the beginning of March (pre-harvest) during the active vegetative and reproductive growth period.
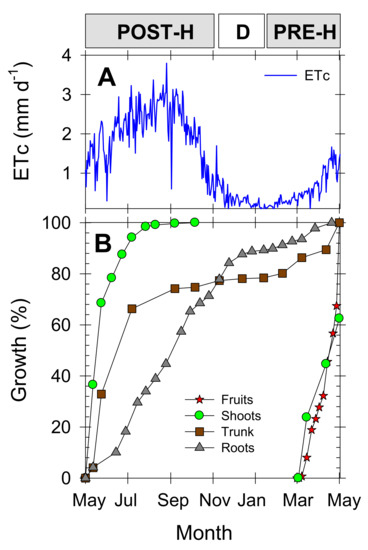
Figure 3.
(A) crop evapotranspiration (ETc), and (B) fruit diameter (red stars), shoot length (green circles), trunk diameter (brown squares), and root length (grey triangles) of young “Flariba” nectarine trees during the experimental period, expressed as percentage of seasonal growth. Phenological periods indicated by POST-H (post-harvest), D (dormancy) and PRE-H (pre-harvest).
To describe a typical phenological series for early-maturing nectarine cultivars, Figure 3B shows the seasonal growth pattern of the different nectarine tree organs (roots, fruits, shoots and trunk) in untreated trees, expressed as a percentage of the total growth at the end of the growing season (considered from May to May the following year). Active vegetative growth occurred in the summer months, during the postharvest period, when trunk growth was seen to be more advanced with respect to that of the roots. Roots continued to grow throughout the season until dormancy, when their growth rate fell slightly. The short fruit growth period of the early-maturing nectarine cultivar coincided with initial shoot growth. The fruits were harvested by early May, by which time the shoots had reached about 60% of their final development (Figure 3B). Root growth was low during the fruit growth period (Figure 3B), coinciding with the period of lowest water demand (Figure 3A).
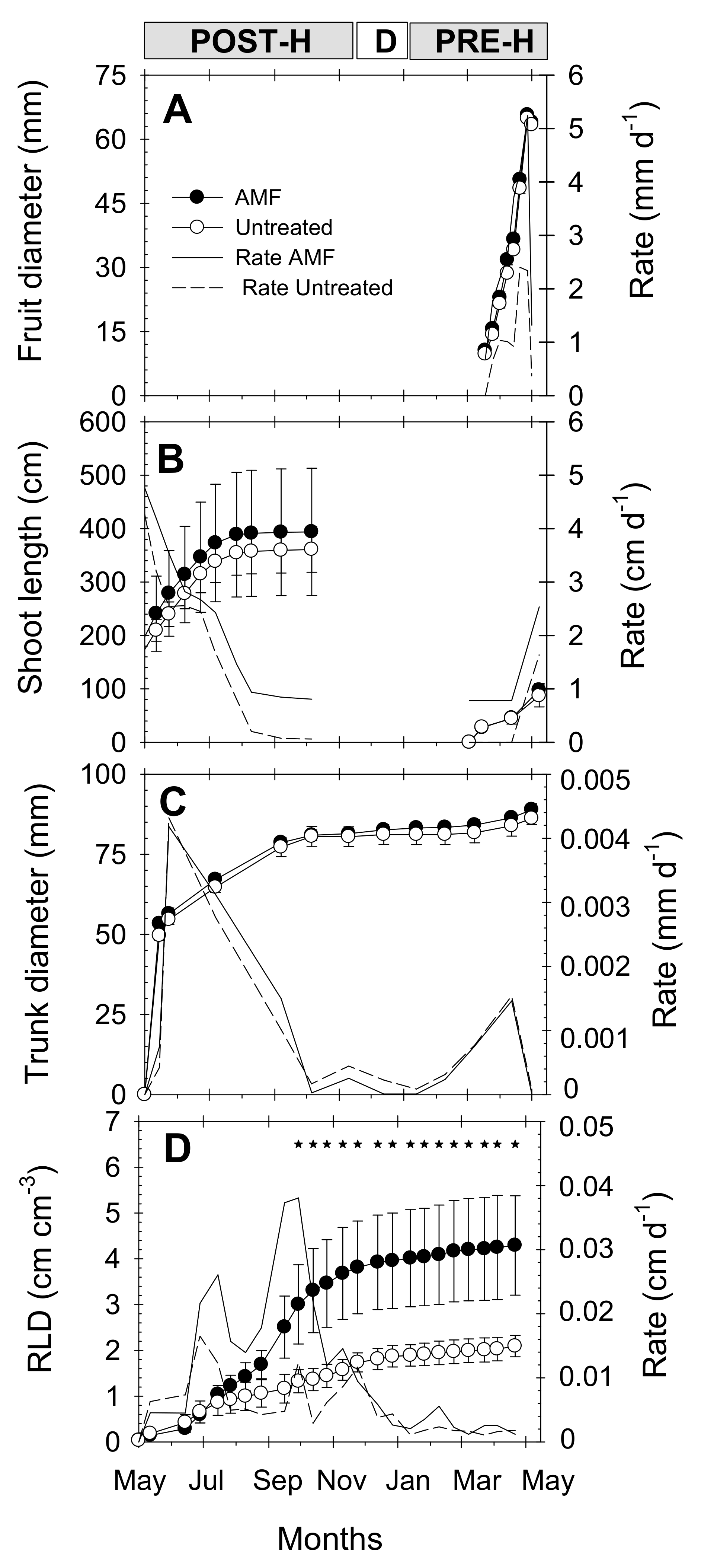
The growth dynamics of fruits, shoots and trunk of young “Flariba” nectarine trees were similar in AMF and untreated trees, with slightly higher absolute values at the end of the season in the AMF trees (Figure 4A,C). Root length density (RLD) values, which took into account all root diameters throughout the soil profile (0–100 cm depth), increased continuously during the growing season in both treatments (Figure 4D). However, clear differences were observed from August, and RLD was significantly higher in AMF than in untreated trees from October onwards, with 51% higher values in AMF inoculated trees than in untreated ones at the end of the season (Figure 4D).
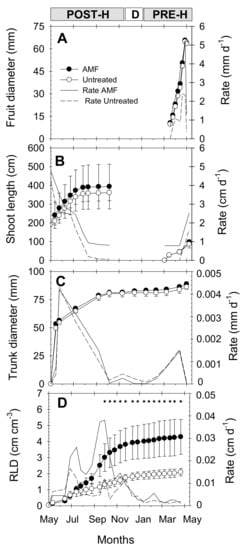
Figure 4.
Seasonal growth (symbols) and growth rate (lines) of (A) fruit diameter, (B) shoot length, (C) trunk diameter, and (D) root length density (RLD) in 0–100 cm soil profile in AMF inoculated (black circles, solid line) and untreated (white circles, dashed line) young “Flariba” nectarine trees. Bars on data point are ± SE of the mean (100 fruits, 16 shoots, 24 trunks and 4 minirhizotrons). Asterisks indicate significant differences in RLD values according to LSD0.05 test. Phenological periods indicated by POST-H (post-harvest), D (dormancy) and PRE-H (pre-harvest).
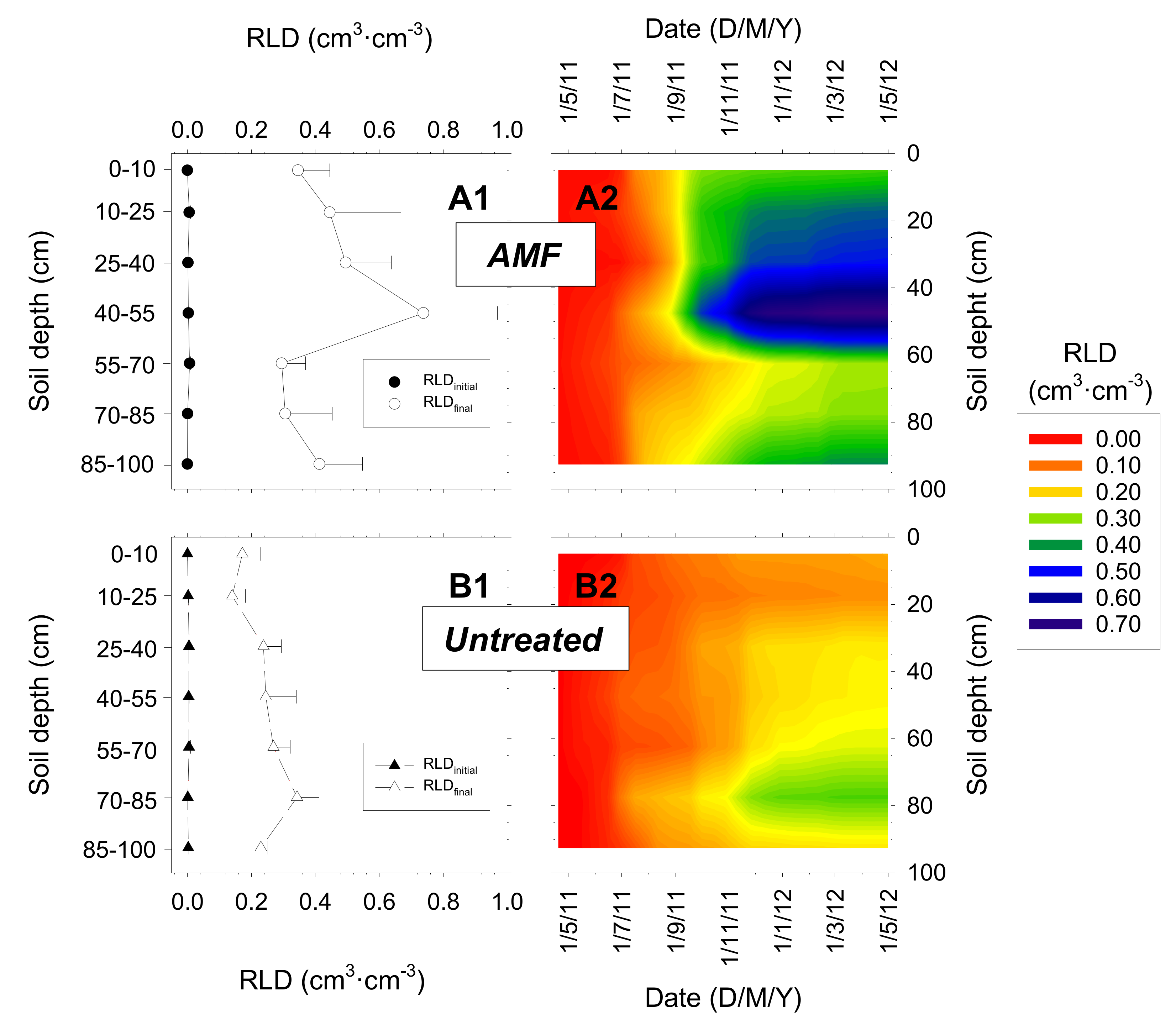
The root distribution in the soil profile throughout the experimental period (one entire growing season) indicated that RLD, which started from almost nil values, increased differently in both treatments (Figure 5). The roots were distributed homogeneously in the soil profile in the untreated trees, and the highest root growth was detected at deep soil layers (0.4 cm cm−3 at 70–85 cm) (Figure 5B). However, in the AMF trees, root growth was high throughout the soil profile, the highest RLD values occurring at 45–70 cm soil depth (main active root zone) root growth was low in the most superficial soil layer (Figure 5A). Mean RLD in the soil profile was 45% higher in the AMF nectarine trees than in untreated trees (0.27 and 0.15 cm cm−3, respectively) (Figure 5).
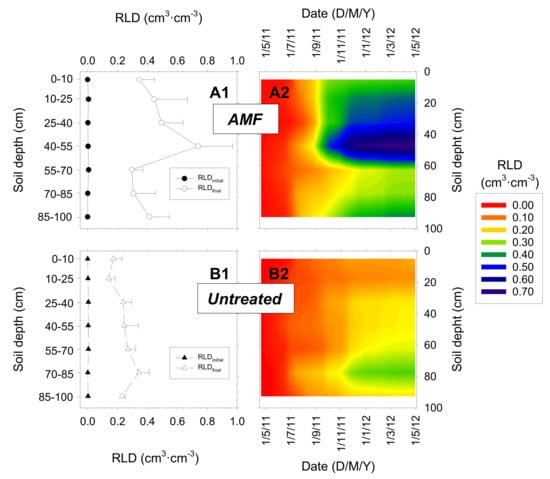
Figure 5.
Root length density (RLD) profiles during the growing season in: (A) AMF inoculated (circles) and (B) untreated (triangles) young “Flariba” nectarine trees, (1) at the beginning (black symbols, RLDinitial) and the end (white symbols, RLDfinal) of the experimental period and (2) throughout the growing season (n = 23 measurements). Horizontal bars on data point are ± SE of the mean (n = 4).
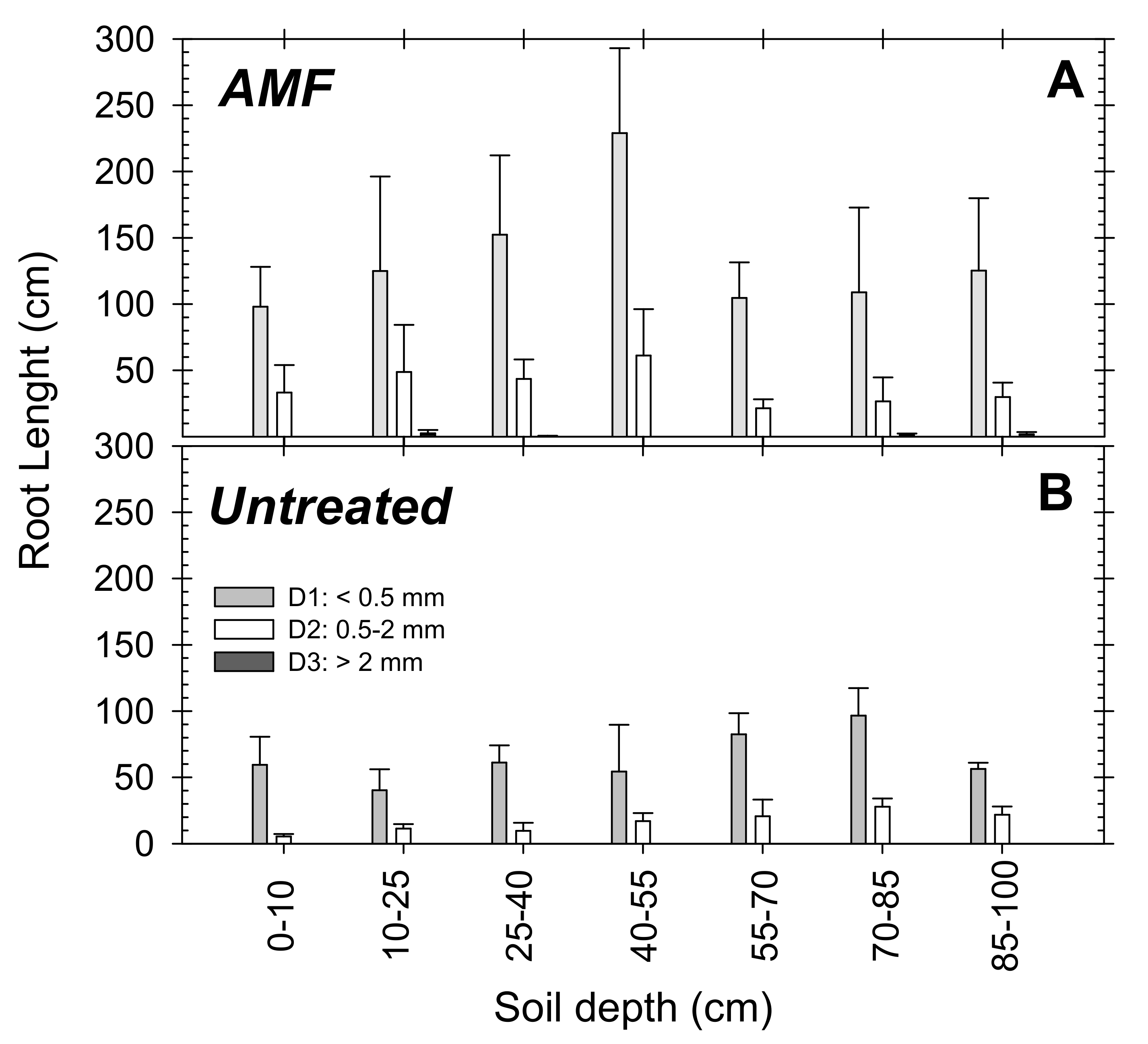
The root data of the young “Flariba” nectarine trees covered all diameters, but when they were analyzed and divided into the main three groups (D1 < 0.5 mm; D2: 0.5–2 mm; D3 > 2 mm), most corresponded to the very fine category (D1) (Figure 6). In the AMF inoculated trees 70% of roots were classified as very fine roots (D1) and 28% as fine (D2). Coarse roots (D3) were only observed in the AMF trees, where they represented 2% of the total roots observed for the treatment (Figure 6A). The root length data of untreated trees were divided into 60% very fine (D1) and 40% fine roots (D2), and no coarse roots were observed (Figure 6B).
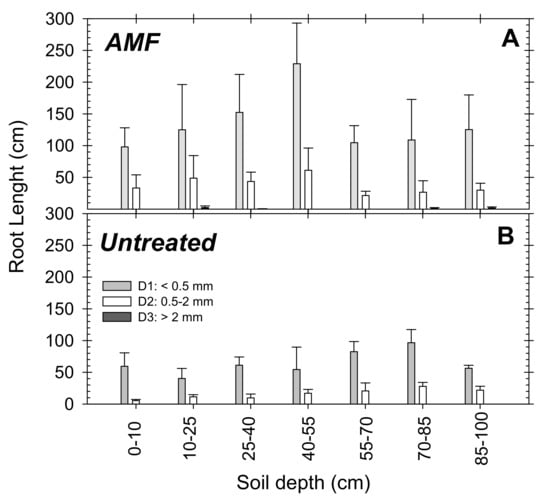
Figure 6.
Root length at the different soil depths in (A) AMF inoculated and (B) untreated young “Flariba” nectarine trees at the end of the growing season (30 April) for each root diameter: grey bars D1 < 0.5 mm; white bars D2: 0.5–2 mm; black bars D3 > 2 mm. Bars are the mean + SE (n = 4).
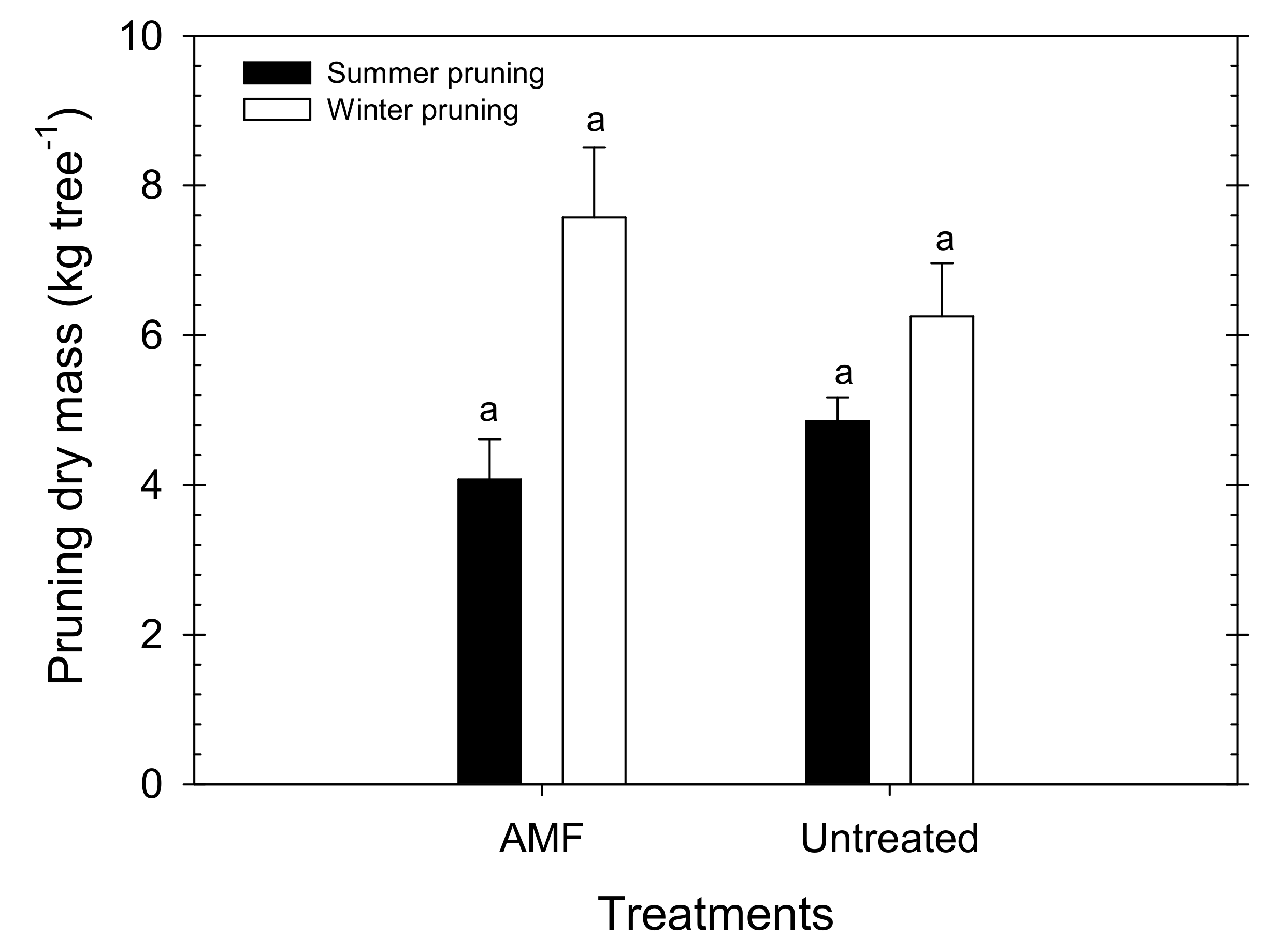
The total pruned weight was similar in AMF and untreated trees, with mean values of 11.64 and 11.10 kg of dry mass per tree, respectively (Figure 7). Although there were no significant differences between treatments, the summer pruning (mainly consisting of eliminating water sprouts) was slightly higher in the untreated trees, while the winter pruning needs were higher in the AMF trees.
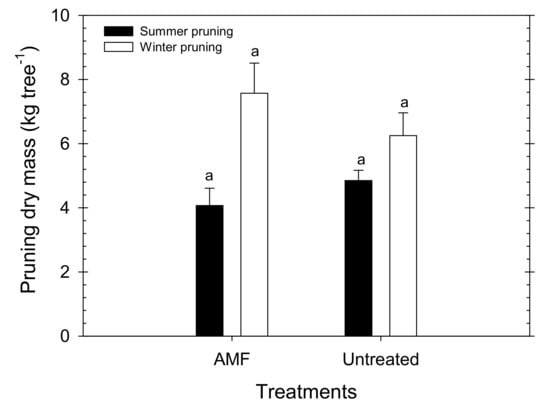
Figure 7.
Summer (black bars) and winter (white bars) pruning dry mass in AMF inoculated and untreated young “Flariba” nectarine trees. Bars are the mean ± SE (n = 6). Letter ‘a’ indicates no significant differences between treatments for each time of pruning.
3.3. Yield Components and Productive Efficiencies
Table 1 presents the effects of AMF inoculation in young “Flariba” nectarine trees on the main yield components. Total yield and the number of fruits were significantly higher (by around 20%) in the AMF trees with respect to untreated ones. Since the same amount of water was applied (4430 m3 ha−1) in both treatments, this implied a 19.5% increase in the irrigation water use efficiency (IWUE) for the AMF treatment. Moreover, the crop load efficiency (CLE) was also 20% higher in the AMF trees, as both treatments had similar canopy areas. Fruit mass, trunk cross sectional area (TCSA) and the productive efficiency (PE) values were similar in both treatments (Table 1).

Table 1.
Yield components in AMF inoculated and untreated young “Flariba” nectarine trees.
3.4. Fruit Quality
The physico-chemical fruit quality traits that were evaluated only showed significant differences in fruit diameter, with higher values recorded in AMF trees than in untreated trees (Table 2). At the first picking date, lightness and Hue angle (h°) values were also higher in AMF trees, but these significant differences had disappeared by the second picking date. Total soluble solid (°Brix) values were similar in both treatments.
Table 2.
Fruit quality at two picking dates in AMF inoculated and untreated young “Flariba” nectarine trees.
The nectarine fruit allometric relationships were derived as fresh to dry mass (y = 0.4063 e0.0471x; r2= 0.63, p ≤ 0.01); equatorial diameter to dry mass (y = 5.9112 e0.0182x; r2 = 0.58, p ≤ 0.01); and equatorial diameter to fresh mass (y = 0.025x2 + 0.002x − 0.26; r2 = 0.98, p ≤ 0.001) ratios.
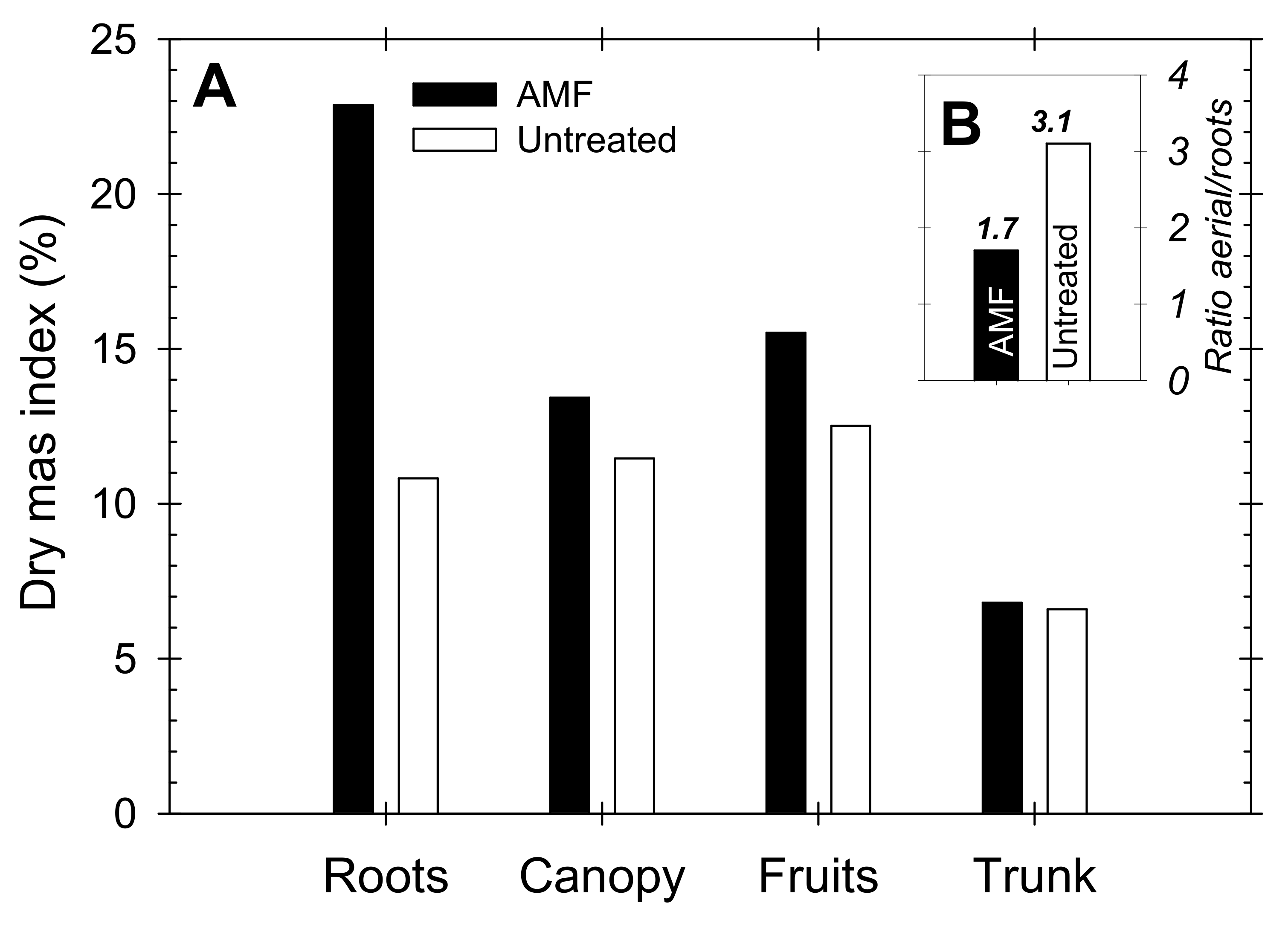
Based on the estimations made by Carvajal et al. [41], the estimated dry mass contribution of the different organs of the nectarine trees were (in g per tree): roots = 8.54 (32%); canopy (shoots + leaves) = 5.87 (22%); fruits = 9.60 (36%); and trunk = 2.83 (11%). These values were weighted with respect to the experimental data obtained at the end of the season (30 April 2012) in both AMF and untreated trees, and the results are depicted in Figure 8. The calculated dry mass index pointed to a higher percentage of roots, canopy and fruits in the AMF trees than in untreated trees, whereas the corresponding trunk values were similar in both treatments. Indeed, the aerial/root portion ratios were 1.7 and 3.1 for the AMF and untreated trees, respectively, reflecting biomass partitioning in favor of roots in the AMF inoculated trees.
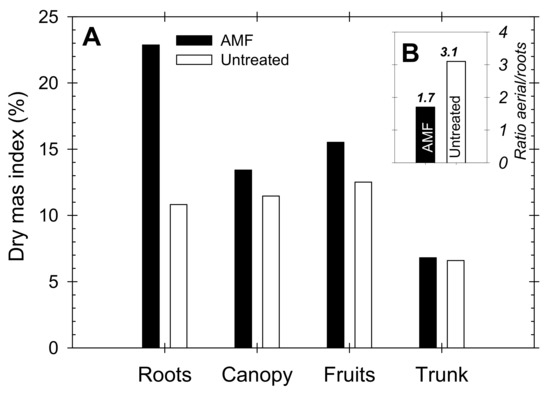
Figure 8.
(A) Dry mass index (%) of roots, canopy, fruits and trunk, and (B) Aerial/root partitioning in AMF inoculated (black bars) and untreated (white bars) young “Flariba” nectarine trees at the end of the growing season.
4. Discussion
Two benefits that arise from using the commercial AMF Glomus iranicum var. tenuihypharum in young “Flariba” nectarine trees are considered in this study: (i) the significant (51%) increase in the root system, as indicated by the higher RLD values measured with minirhizotrons, by the end of the season, and (ii) the improved crop productivity response at harvest compared with untreated trees. Hence, the yield and number of fruits per tree significantly increased (by ≈20%) in the inoculated trees despite the similar tree size (canopy and trunk area). Furthermore, the crop load efficiency (CLE) and the irrigation water use efficiency (IWUE) also increased by ≈20% in the AMF trees. All the above data confirm the beneficial effect of applying AMF to stimulate field-grown young fruit tree crops.
The AMF successfully colonized the roots of the drip irrigated “Flariba” nectarine trees, as indicated by the root data (Figure 4D, Figure 5 and Figure 6), although data on the percentage of colonization in soil samples taken in summer were not available due to a technical issue. Bucher et al. [11] indicated that the effective symbiosis of arbuscular mycorrhiza fungi allows the plants to explore a greater volume of soil by producing extra-matricial mycelia that extend from colonized tree root systems into the soil. Lû et al. [12] found that AMF inoculation had a positive influence on the soil microbe balance as a result of improvements in soil aggregate stability and changes root exudate composition. Additionally, Calvet et al. [15] observed the inhibition of nematode roots colonization after AMF inoculation in the peach-almond hybrid GF-677.
In our study, the mean seasonal RLD values for the AMF inoculated and untreated trees were 0.4 cm cm−3 and 0.2 cm root cm−3 soil, respectively (Figure 5). Furthermore, there was a significantly higher percentage of very fine roots, with a diameter of <0.5 mm (Figure 6) in AMF inoculated trees. It is known that AMF tend to colonize tap and lateral roots as it is easier to penetrate young growing roots [44] than thicker roots (>0.5 mm diameter), which are more lignified as they have an anchoring role. Indeed, the fine roots are considered more active than coarse roots since they contribute more to nutrient uptake [45]. Hence, the increase in RLD as well as the percentage of very fine roots in AMF trees was indicative of the successful mycorrhiza roots colonization. In peach seedlings grown in pots, Alvarado-Raya [46] described that AMF inoculation resulted in a root colonization > 70% of the total root length.
The minirhizotron technique was seen to be a useful tool for studying young “Flariba” nectarine root dynamics (Figure 4, Figure 5 and Figure 6). Similarly, Abrisqueta et al. [26] demonstrated the feasibility of using minirhizotrons in adult field-grown peach trees for studying root dynamics and distribution. These authors indicated that peach roots grew continuously during the growing season, although more slowly during the last stage of fruit growth.
Our results showed no significant differences between AMF and untreated trees in terms of aerial vegetative growth even though the absolute values of the canopy area and volume were higher in the AMF inoculated plants (Table 1). Similar results were obtained for the dry mass index, which reached 1.7 in AMF and 3.1 in the untreated trees (Figure 8), indicating the positive effect of AMF application on biomass partitioning in favor of roots with respect to the aerial portion.
Root colonization is associated with improved host nutrition, particularly of P [20,47,48,49]. Abbott and Robson [50] indicated that phosphate absorption was one of the most important factors involved in the increase in growth detected in AMF inoculated plants. Indeed, Baldi et al. [51] observed an increase in the leaf phosphorus concentration, while no change was noted in other leaf nutrients, after AMF application in field-grown nectarine trees. The activity of the enzyme fungal alkaline phosphatase (ALP), which was identified in AMF, was seen to facilitate nutrient uptake (including P) and photosynthesis processes, all would contribute to the improved growth noted in inoculated plants [20,52,53]. Kobe et al. [54] reported that non-structural carbohydrates dominate root growth since fine root production seems to increase with N availability [55]. AMF have also been said to increase the use of different forms of N by plants [56] and to induce N uptake for direct transfer to the host root [57].
Nectarine tree root growth was seen to decrease during the rapid fruit growth period, which acted as powerful photo-assimilate sinks (Figure 4A,D). When the fruits were harvested, shoots had reached approximately 60% of their final length. Roots and trunk follow the same pattern of growth with maximum rates during the postharvest period (Figure 4C). This asynchrony in aerial vs. root growth has been reported in other deciduous fruit trees, such as almond [58], peach [26] and apricot [59]. In this sense, Chalmers and van den Ende [60] indicated that the growth of the fruit and roots is competitively inter-related throughout the life of the peach tree, while the aerial vegetative growth (i.e., leaf, trunk and shoot) is independent. Furthermore, root physiology of one-year-old compost-grown nectarine trees was influenced more than aerial biomass by the changes in growing conditions [61].
In our study, nectarine fruits at the two harvests were classified as “Extra class” according to EEC directive 3596/90 [62], which enforces a minimum diameter of 56 mm as the minimum diameter for nectarine fruit to be considered as such. The total yield and the crop load were significantly (20%) higher in AMF inoculated trees (Table 1). These positive changes as a result of AMF inoculation were probably linked to the increase in photo-assimilate production, nutrient translocation, as well as hormone production, all of which increase fruit production [63]. Some authors have reported a good relationship between yield components and mycorrhizal spore population in other fruit species such as plum and cherry [64], apple [65,66] and peach [67]. Fruit diameter was also significantly higher in AMF trees compared with un-inoculated plants (Table 2). Shresta et al. [68] reported that fruit size was greater in ‘Satsuma’ mandarin trees inoculated with different species of AMF, resulting in higher yields. These findings lead to the conclusion that the photo-assimilates produced are destined to fruit production rather than vegetative growth. Nicolás et al. [23] observed that AMF favored root development in ‘Crimson Seedless’ table grapes by mobilizing the apex starch reserves, leading to a higher concentration of starch in the inner part of the root.
The presence of AMF in the root zone has been described as a mechanism to protect the plant from adverse environmental stresses, such as water deficit, flooding or diseases [13,17,22,59,69]. Colonization leads to further soil exploration by new roots, leading to an increase in water uptake by the roots, which translates into a better plant water status [70], as assessed from Ψstem [23]. However, our results did not show any significant differences in the seasonal Ψstem values in either AMF or untreated trees, which might be attributed to the young character of the cultivar studied (Figure 2A). Conesa et al. [71] quantified the effect of mycorrhiza inoculated Citrus roots as an improvement of 0.4–0.7 MPa. Additionally, the mean Ψstem values observed during the growing season (−0.42 and −0.82 MPa in the pre- and post-harvest periods, respectively) were characteristic of well-watered nectarine trees [72,73,74], and coincided with the irrigation applied, which was based on 100% of computed ETc values.
Finally, AMF inoculation increased IWUE and also CLE values by 20% (Table 1) which highlights the essential role of AMF application in semi-arid areas, where water resources are limited [14,18].
5. Conclusions
The commercial AMF selected, Glomus iranicum var. tenuihypharum, was capable of colonizing the ‘Flariba’ nectarine roots satisfactorily, increasing the root length by around 50% with respect to the untreated trees when measured one year after inoculation. There were also positive effects on yield components and productive efficiencies such as yield, number of fruits per tree, crop load efficiency and irrigation water use efficiency, which were significantly (≈20%) higher in AMF inoculated trees. However, aerial biomass and plant water status remained unchanged.
Hence, the inducement of such symbiosis might be recommended to increase the sustainability of agricultural practices in semi-arid areas threatened by the consequences of climate change, since the application of AMF was seen to improve the establishment of nectarine trees in field conditions during their early years by enhancing root system development. Nonetheless, in spite of the positive findings observed, more research is needed to confirm the beneficial effects of AMF inoculation in adult nectarine trees.
Author Contributions
Conceptualization, M.C.R.-S. and J.V.; methodology and formal analysis, J.V., W.C. and L.L.-M.; data curation, W.C. and L.L.-M.; writing—original draft preparation, M.R.C.; writing—review and editing, M.R.C., M.C.R.-S., J.V., W.C., L.L.-M.; project administration and funding acquisition, M.C.R.-S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Spanish Economy and Competitiveness Ministry (MINECO)-FEDER (AGL2013-49047-C2-2R), the Spanish State Research Agency (PID2019-106226RB-C21/AEI/10.13039/501100011033) and the Seneca Foundation of Region of Murcia (Spain) under the Excellence Group Program (19903/GERM/15) projects. M.R. Conesa received a postdoctoral financial support from the Juan de la Cierva Spanish program (FJCI-2017-32045).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Iglesias, I.; Echeverría, G. Differential effect of cultivar and harvest date on nectarine colour, quality and consumer acceptance. Sci. Hortic. 2009, 120, 41–50. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Food and Agriculture Organization Statistical Data 2020. Available online: http://www.fao.org/faostat/en/?#data/QC (accessed on 15 March 2021).
- Lionello, P.; Malanotte-Rizzoli, P.; Boscolo, R.; Alpert, P.; Artale, V.; Li, L.; Luterbacher, J.; May, W.; Trigo, R.M.; Tsimplis, M.; et al. The Mediterranean climate: An overview of the main characteristics and issues. In Mediterranean Climate Variability; Lionello, P., Malanotte-Rizzoli, P., Boscolo, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2006; Volume 4, pp. 1–26. [Google Scholar]
- Fereres, E.; Soriano, M.A. Deficit irrigation for reducing agricultural water use. J. Exp. Bot. 2006, 58, 147–159. [Google Scholar] [CrossRef] [Green Version]
- English, M.J.; Solomon, K.H.; Hoffman, G.J. A Paradigm Shift in Irrigation Management. J. Irrig. Drain. Eng. 2002, 128, 267–277. [Google Scholar] [CrossRef]
- Passioura, J.B. Water transport in and to roots. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1988, 39, 245–256. [Google Scholar] [CrossRef]
- Vamerali, T.; Saccomani, M.; Bona, S.; Mosca, G.; Guarise, M.; Ganis, A. A comparison of root characteristics in relation to nutrient and water stress in two maize hybrids. Plant Soil 2003, 157–167. [Google Scholar] [CrossRef]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and crop responses: A review. Biol. Agric. Hortic. 2014, 31, 1–17. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. (Eds.) Colonization of roots and anatomy of arbuscular mycorrhiza. In Mycorrhizal Symbiosis; Academic Press: London, UK, 2008; pp. 42–90. [Google Scholar]
- Sánchez-Blanco, M.J.; Álvarez, S.; Ortuño, M.F.; Ruiz-Sánchez, M.C. Root System Response to Drought and Salinity: Root Distribution and Water Transport. In Root Engineering, Soil Biology; Springer: Berlin/Heidelberg, Germay, 2014; pp. 325–352. [Google Scholar] [CrossRef]
- Bucher, M.; Wegmüller, S.; Drissner, D. Chasing the structures of small molecules in arbuscular mycorrhizal signaling. Curr. Opin. Plant Biol. 2009, 12, 500–507. [Google Scholar] [CrossRef]
- Lǚ, L.-H.; Zou, Y.-N.; Wu, Q.-S. Mycorrhizas Mitigate Soil Replant Disease of Peach Through Regulating Root Exudates, Soil Microbial Population, and Soil Aggregate Stability. Commun. Soil Sci. Plant Anal. 2019, 50, 909–921. [Google Scholar] [CrossRef]
- Chitarra, W.; Pagliarani, C.; Maserti, B.; Lumini, E.; Siciliano, I.; Cascone, P.; Schubert, A.; Gambino, G.; Balestrini, R.; Guerrieri, E. Insights on the Impact of Arbuscular Mycorrhizal Symbiosis On Tomato Tolerance to Water Stress. Plant Physiol. 2016, 171, 1009–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of Arbuscular Mycorrhizal Fungi on Plant Growth and Performance: Importance in Biotic and Abiotic Stressed Regulation. Diversity 2020, 12, 370. [Google Scholar] [CrossRef]
- Calvet, C.; Pinochet, J.; Hernández-Dorrego, A.; Estaún, V.; Camprubi, A. Field microplot performance of the peach-almond hybrid GF-677 after inoculation with arbuscular mycorrhizal fungi in a replant soil infested with root-knot nematodes. Mycorrhiza 2001, 10, 295–300. [Google Scholar] [CrossRef]
- Lenoir, I.; Fontaine, J.; Sahraoui, A.L.-H. Arbuscular mycorrhizal fungal responses to abiotic stresses: A review. Phytochemistry 2016, 123, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, J.M.R.; Azcon, R.; Gomez, M. Effects of arbuscular-mycorrhizal glomus species on drought tolerance: Physiological and nutritional plant responses. Appl. Environ. Microbiol. 1995, 61, 456–460. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Fernández, C.; Aroca, R.; Barea, J.M. Influence of arbuscular mycorrhizal fungi and water regime on the development of endemic Thymus species in dolomitic soils. Appl. Soil Ecol. 2011, 48, 31–37. [Google Scholar] [CrossRef]
- Posta, K.; Duc, N.H. Benefits of Arbuscular Mycorrhizal Fungi Application to Crop Production under Water Scarcity. 2020. Available online: https://www.intechopen.com/books/droughtdetection-and-solutions/benefits-of-arbuscular-mycorrhizal-fungi-application-to-crop-production-underwater-scarcity (accessed on 15 March 2020).
- Franco, J.A.; Abrisqueta, J.M. A comparison between minirhizotron and soil coring methods of estimating root distribution in young almond trees under trickle irrigation. J. Hortic. Sci. 1997, 72, 797–805. [Google Scholar] [CrossRef]
- Gómez-Bellot, M.J.; Ortuño, M.F.; Nortes, P.A.; Vicente-Sánchez, J.; Martín, F.F.; Bañón, S.; Sánchez-Blanco, M.J. Protective effects of Glomus iranicum var. tenuihypharum on soil and Viburnum tinus plants irrigated with treated wastewater under field conditions. Mycorrhiza 2015, 25, 399–409. [Google Scholar] [CrossRef]
- Nicolás, E.; Maestre-Valero, J.F.; Alarcón, J.J.; Pedrero, F.; Vicente-Sánchez, J.; Bernabé, A.; Fernández, F. Effectiveness and persistence of arbuscular mycorrhizal fungi on the physiology, nutrient uptake and yield of Crimson Seedless grapevine. J. Agric. Sci. 2015, 153, 1084–1096. [Google Scholar] [CrossRef] [Green Version]
- Zheng, F.-L.; Liang, S.-M.; Chu, X.-N.; Yang, Y.-L.; Wu, Q.-S. Mycorrhizal fungi enhance flooding tolerance of peach through inducing proline accumulation and improving root architecture. Plant Soil Environ. 2020, 66, 624–631. [Google Scholar] [CrossRef]
- Bland, W.L.; Mesarch, M.A. Counting error in the line-intercept method of measuring root length. Plant Soil 1990, 125, 155–157. [Google Scholar] [CrossRef]
- Abrisqueta, I.; Conejero, W.; López-Martínez, L.; Vera, J.; Sánchez, M.C.R. Root and aerial growth in early-maturing peach trees under two crop load treatments. Span. J. Agric. Res. 2017, 15, e0803. [Google Scholar] [CrossRef] [Green Version]
- Taylor, H.M. Minirhizotron Observation Tubes: Methods and Applications for Measuring Rhizosphere Dynamics; American Society of Agronomy: Madison, WI, USA, 1987; 143p. [Google Scholar]
- Upchurch, D.R. Conversion of minirhizotron-root intersections to root length density. In Minirhizotron Observation Tubes: Methods and Applications for Measuring Rhizosphere Dynamics; Taylor, H.M., Ed.; Wiley: Hoboken, NJ, USA, 1987; pp. 51–65. [Google Scholar]
- Machado, R.M.A.; Oliveira, M.D.R.G. Comparison of tomato root distributions by minirhizotron and destructive sampling. Plant Soil 2003, 255, 375–385. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Hammer, R.D.; Blanchar, R.W. Minirhizotron quantification of soybean root growth as affected by reduced A horizon in soil. J. Plant Nutr. Soil Sci. 2003, 166, 708–711. [Google Scholar] [CrossRef]
- Abrisqueta, J.; Mounzer, O.; Álvarez, S.; Conejero, W.; García-Orellana, Y.; Tapia, L.; Vera, J.; Abrisqueta, I.; Sánchez, M.C.R. Root dynamics of peach trees submitted to partial rootzone drying and continuous deficit irrigation. Agric. Water Manag. 2008, 95, 959–967. [Google Scholar] [CrossRef]
- Wells, C.E.; Glenn, D.M.; Eissenstat, D.M. Changes in the risk of fine-root mortality with age: A case study in peach, Prunus persica (Rosaceae). Am. J. Bot. 2002, 89, 79–87. [Google Scholar] [CrossRef]
- Bernier, P.Y.; Robitaille, G. A plane intersect method for estimating fine root productivity of trees from minirhizotron images. Plant Soil 2004, 265, 165–173. [Google Scholar] [CrossRef]
- Fernández, J.E.; Moreno, F.; Martín-Aranda, J.; Fereres, E. Olive-tree root dynamics under different soil water regimes. Agric. Mediterr. 1992, 122, 225–235. [Google Scholar]
- Conesa, M.R.; Martínez-López, L.; Conejero, W.; Vera, J.; Ruiz-Sánchez, M. Summer pruning of early-maturing Prunus persica: Water implications. Sci. Hortic. 2019, 256, 108539. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guidelines for Computing Crop Water Requirements; Food and Agriculture Organization of the United Nations: Rome, Italy, 1998; Available online: http://www.fao.org/docrep/x0490e/x0490e00.htm (accessed on 20 April 2011).
- Abrisqueta, I.; Abrisqueta, J.; Tapia, L.; Munguía, J.; Conejero, W.; Vera, J.; Ruiz-Sánchez, M. Basal crop coefficients for early-season peach trees. Agric. Water Manag. 2013, 121, 158–163. [Google Scholar] [CrossRef]
- Fereres, E.; Martinich, D.A.; Aldrich, T.M.; Castel, J.R.; Holzapfel, E.; Schulbach, H. Drip irrigation saves money in young almond orchards. Calif. Agric. 1982, 36, 12–13. [Google Scholar]
- Mokrini, F.; Waeyenberge, L.; Viaene, N.; Moens, M. First Report of the Cereal Cyst Nematode Heterodera latipons on Wheat in Morocco. Plant Dis. 2012, 96, 774. [Google Scholar] [CrossRef]
- Hsiao, T.C. Measurement of tree water status. In Irrigation of Agricultural Crops. Agronomy Monograph; Steward, B.A., Nielsen, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1990; pp. 243–279. [Google Scholar]
- Merrill, S.D.; Upchurch, D.R. Converting Root Numbers Observed at Minirhizotrons to Equivalent Root Length Density. Soil Sci. Soc. Am. J. 1994, 58, 1061–1067. [Google Scholar] [CrossRef]
- IPCC (Intergovernmental Panel on Climate Change). Good Practice Guidance for Land Use, Land-Use Change and Forestry; Penman, J., Gytarsky, M., Hiraishi, T., Krug, T., Kruger, D., Pipatti, R., Buendía, L., Miwa, K., Ngara, T., Tanabe, K., et al., Eds.; IPCC: Geneva, Switzerland, 2003. [Google Scholar]
- Carvajal, M.; Mota, C.; Alcaráz-López, C.; Iglesias, M.; Martínez-Ballesta, M.C. Investigación sobre la absorción de CO2 por los cultivos más representativos de la Región de Murcia. In Etiquetado de Carbono en Las Explotaciones y Productos Agrícolas: La Iniciativa Agricultura Murciana Como Sumidero de CO2; Consejería de Agricultura y Agua (CARM): Murcia, Spain, 2010; pp. 65–92. ISBN 978-84-693-6838-1. [Google Scholar]
- Hooker, J.E.; Munro, M.; Atkinson, D. Vesicular-arbuscular mycorrhizal fungi induced alteration in poplar root system morphology. Plant Soil 1992, 145, 207–214. [Google Scholar] [CrossRef]
- Alarcón, A.L.; Gómez-Bellot, M.J.; Bernabe, A.J.; Calvo, G.; Martín, F.F. Changes in root architecture and productivity of melon (Cucumis melo L. cv. Hispano Nunhems) promoted by Glomus iranicum var. tenuihypharum. J. Hortic. Sci. Biotechnol. 2019, 95, 364–373. [Google Scholar] [CrossRef]
- Alvarado-Raya, H.E. Peach seedling growth with mycorrhiza and vermicompost. Tecnociencia 2017, 9, 48–57. [Google Scholar]
- Giovanetti, M.; Mosse, B. An evolution of techniques to measure vesicular-arbuscular infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular mycorrhizal fungi in alleviation of salt stress: A review. Ann. Bot. 2009, 104, 1263–1280. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.S.; Li, G.H.; Zou, Y.N. Roles of arbuscular mycorrhizal fungi on growth and nutrient acquisition of peach (Prunus persica L. Batsch) seedlings. J. Anim. Plant. Sci. 2011, 21, 746–750. [Google Scholar]
- Abbott, L.; Robson, A. Factors influencing the occurrence of vesicular-arbuscular mycorrhizas. Agric. Ecosyst. Environ. 1991, 35, 121–150. [Google Scholar] [CrossRef]
- Baldi, E.; Amadei, P.; Pelliconi, F.; Tosell, M. Use of Tricho derma spp. and arbuscular mycorrhizal fungi to increase soil beneficial population of bacteria in a nectarine commercial orchard: Effect on root growth, nutrient acquisition and replanting disease. J. Plant Nutr. 2015, 39, 1147–1155. [Google Scholar] [CrossRef]
- Gianinazzi, S.; Gianinazzi-Pearson, V.; Tisserant, B.; Lemoine, M.C. Protein Activities as Potential Markers of Functional Endomycorrhizae in Planta. In Mycorrhizas in Ecosystems; Read, D.J., Lewis, D.H., Fitter, A.H., Alexander, I.J., Eds.; CAB International: Wallingford, UK, 1992; pp. 333–339. [Google Scholar]
- Amaya-Carpio, L.; Davies, F.; Fox, T.; He, C. Arbuscular mycorrhizal fungi and organic fertilizer influence photosynthesis, root phosphatase activity, nutrition, and growth of Ipomoea carnea ssp. Fistulosa. Photosynthetica 2009, 47, 1–10. [Google Scholar] [CrossRef]
- Kobe, R.K.; Iyer, M.; Walters, M.B. Optimal partitioning theory revisited: Nonstructural carbohydrates dominate root mass responses to nitrogen. Ecology 2010, 91, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Pregitzer, K.S.; Hendrick, R.L.; Fogel, R. The demography of fine roots in response to patches of water and nitrogen. New Phytol. 1993, 125, 575–580. [Google Scholar] [CrossRef] [Green Version]
- Hodge, A.; Campbell, C.; Fitter, A.H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature 2001, 413, 297–299. [Google Scholar] [CrossRef] [Green Version]
- Johansen, A.; Finlay, R.D.; Olsson, P.A. Nitrogen metabolism of external hyphae of the arbuscular mycorrhizal fungus Glomus intraradices. New Phytol. 1996, 133, 705–712. [Google Scholar] [CrossRef]
- Ross, N.W.; Catlin, P.B. Rootstocks and root physiology. In Almond Orchard Management; Micke, W., Kester, D., Eds.; Division of Agricultural Sciences, University of California: Berkeley, CA, USA, 1978; pp. 25–29. [Google Scholar]
- Pastor, A.P.; Sánchez, M.C.R.; Domingo, R.; Torrecillas, A. Growth and phenological stages of Búlida apricot trees in south-east Spain. Agronomie 2004, 24, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Chalmers, D.J.; Van den Ende, B. Productivity of peach trees. Factors affecting dry weight distribution during tree growth. Ann. Bot. 1975, 39, 423–432. [Google Scholar] [CrossRef]
- Sorrenti, G.; Muzzi, E.; Toselli, M. Root growth dynamic and plant performance of nectarine trees amended with biochar and compost. Sci. Hortic. 2019, 257, 108710. [Google Scholar] [CrossRef]
- European Union. Commission Regulation (EC) No 1221/2008 of 5 December 2008. Amending Regulation (EC) No 1580/2007 laying down implementing rules of Council Regulations (EC) No 2200/96, (EC) No 2201/96 and (EC) No 1182/2007 in the fruit and vegetable sector as regards marketing standards. Off. J. Eur. Union 2008, 51, 80. [Google Scholar]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.; Bonini, P.; Colla, G. Plant- and Seaweed-Based Extracts Increase Yield but Differentially Modulate Nutritional Quality of Greenhouse Spinach through Biostimulant Action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Slawomir, S.; Aleksander, S. The influence of mycorrhizal fungi on the growth and yield of plum and sour cherry trees. J. Fruit Ornam. Plant Res. 2010, 18, 71–77. [Google Scholar]
- Sharma, S.D.; Bhutani, V.P.; Dohroo, N.P. Occurrence of VAM fungi under old apple orchards. J. Indian Soc. Soil Sci. 1998, 46, 143–144. [Google Scholar]
- Sharma, S.; Sharma, N.; Sharma, C.; Sood, R.; Singh, R. Studies on Correlations Between Endomycorrhizal and Azotobacter Population With Growth, Yield And Soil Nutrient Status Of Apple (Malus domestica Borkh) Orchards in Himachal Pradesh. Acta Hortic. 2005, 283–287. [Google Scholar] [CrossRef]
- Awasthi, R.P.; Godara, R.K.; Kaith, N.S. Correlation between VA-mycorrhizae spore number, root colonization, Azotobacter population and fruit yield of July Elberta peach. J. Hill Res. 1999, 12, 1–4. [Google Scholar] [CrossRef]
- Shrestha, Y.H.; Ishii, T.; Matsumoto, I.; Kadoya, K. Effects of Vesicular-Arbuscular Mycorrhizal Fungi on Satsuma Mandarin Tree Growth and Water Stress Tolerance and on Fruit Development and Quality. J. Jpn. Soc. Hortic. Sci. 1996, 64, 801–807. [Google Scholar] [CrossRef]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; De Pascale, S.; Bonini, P.; Colla, G. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Azcón, R. Hyphal contribution to water uptake in mycorrhizal plants as affected by the fungal species and water status. Physiol. Plant. 1995, 95, 472–478. [Google Scholar] [CrossRef]
- Conesa, M.; Espinosa, P.; Pallarés, D.; Pérez-Pastor, A. Influence of Plant Biostimulant as Technique to Harden Citrus Nursery Plants before Transplanting to the Field. Sustainability 2020, 12, 6190. [Google Scholar] [CrossRef]
- Naor, A. Irrigation Scheduling and Evaluation of Tree Water Status in Deciduous Orchards. Hortic. Rev. 2010, 111–165. [Google Scholar] [CrossRef]
- Conesa, M.R.; Conejero, W.; Vera, J.; Agulló, V.; García-Viguera, C.; Ruiz-Sánchez, M.C. Irrigation management practices in nectarine fruit quality at harvest and after cold storage. Agric. Water Manag. 2021, 243, 106519. [Google Scholar] [CrossRef]
- López, G.; Echeverria, G.; Bellvert, J.; Mata, M.; Behboudian, M.H.; Girona, J.; Marsal, J. Water stress for a short period before harvest in nectarine: Yield, fruit composition, sensory quality, and consumer acceptance of fruit. Sci. Hortic. 2016, 211, 1–7. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).