
Carbon Cycling in Mangrove Ecosystem of Western Bay of Bengal (India)


Abstract
1. Introduction
2. Materials and Methods
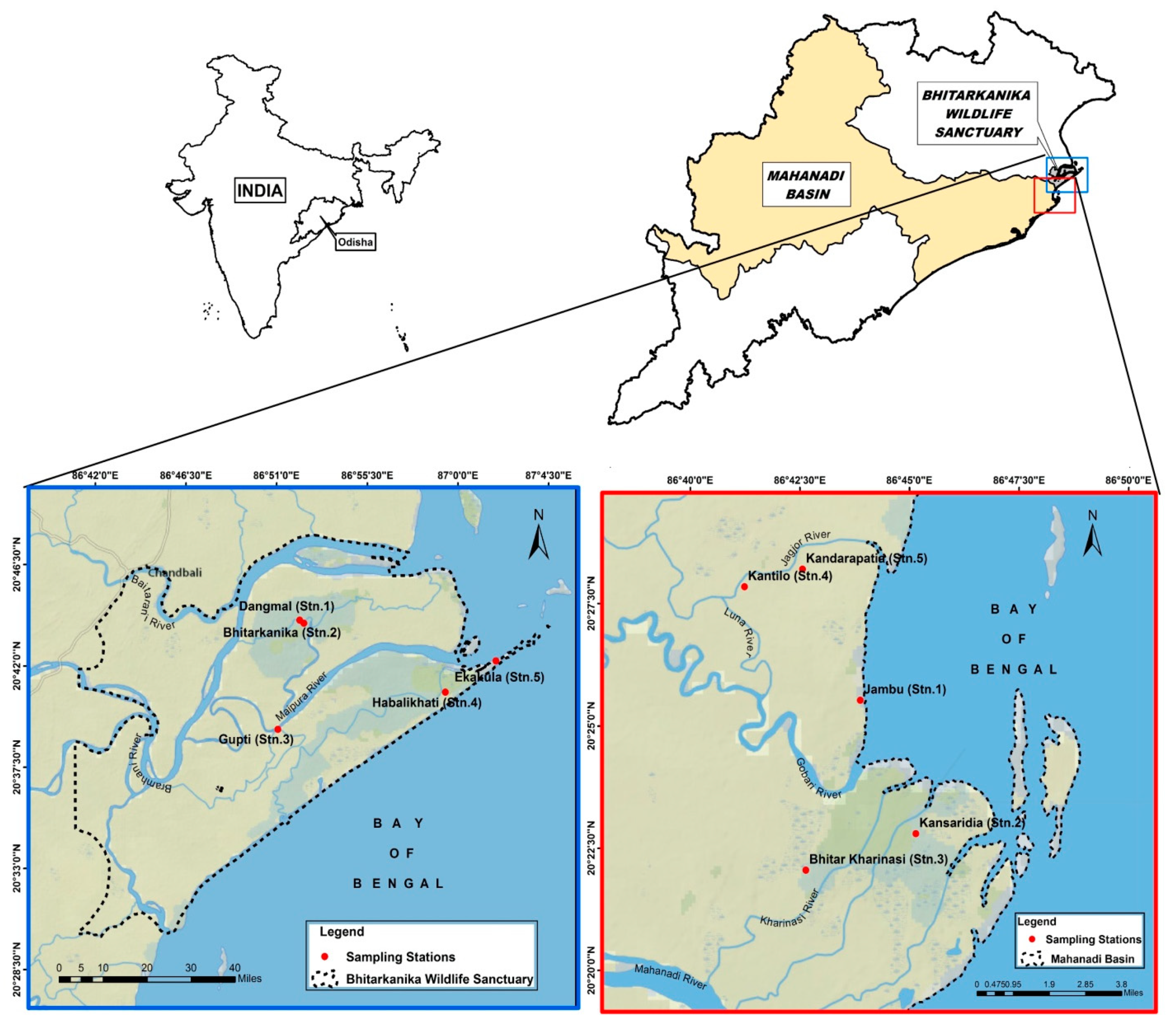
2.1. Study Area
2.2. Estimation of above Ground Biomass and above Ground Carbon
2.3. Analysis of Soil Organic Carbon (SOC)
2.4. Analysis of Dissolved Inorganic Carbon (DIC)
2.5. Statistical Analyses
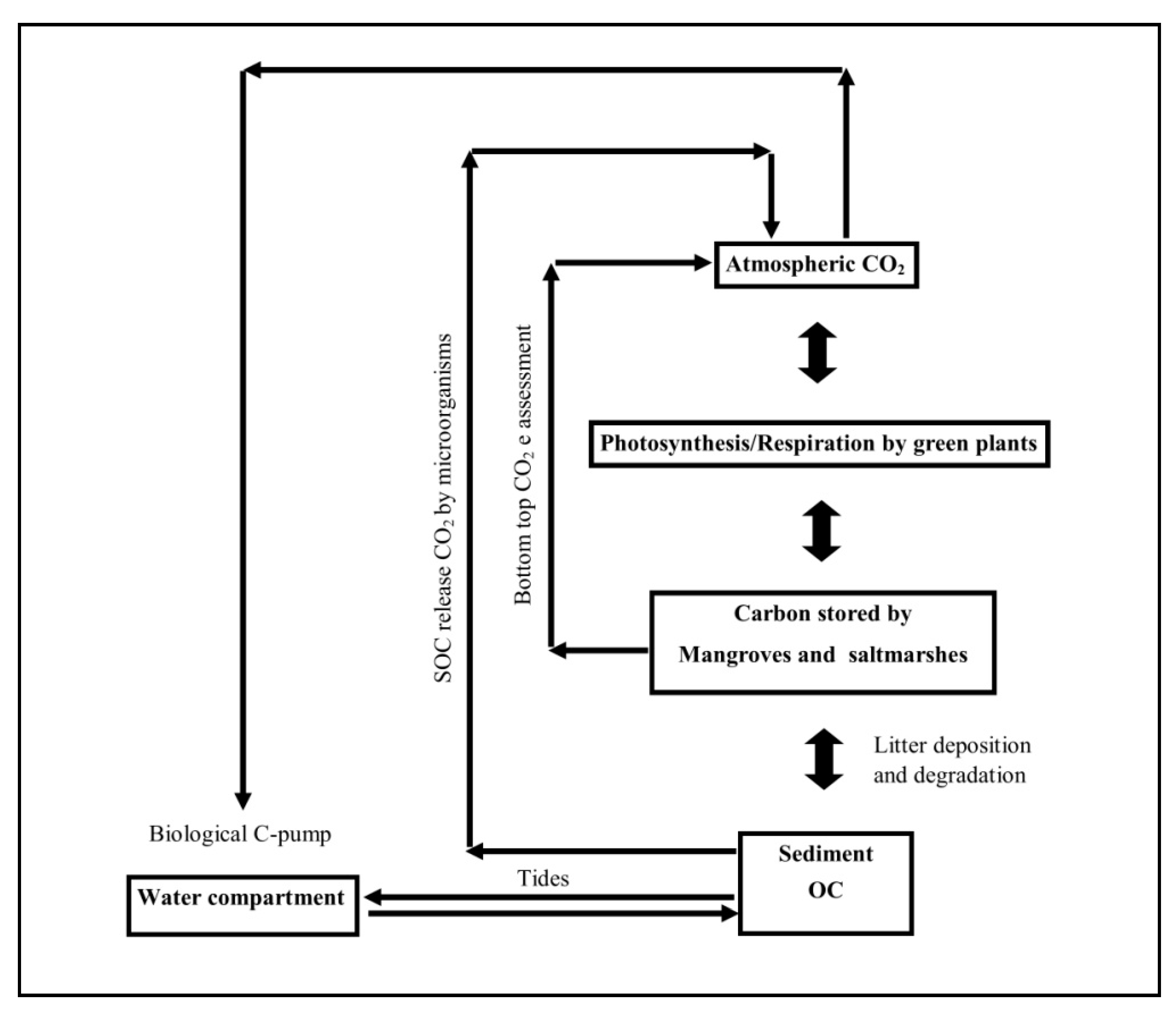
3. Results and Discussion
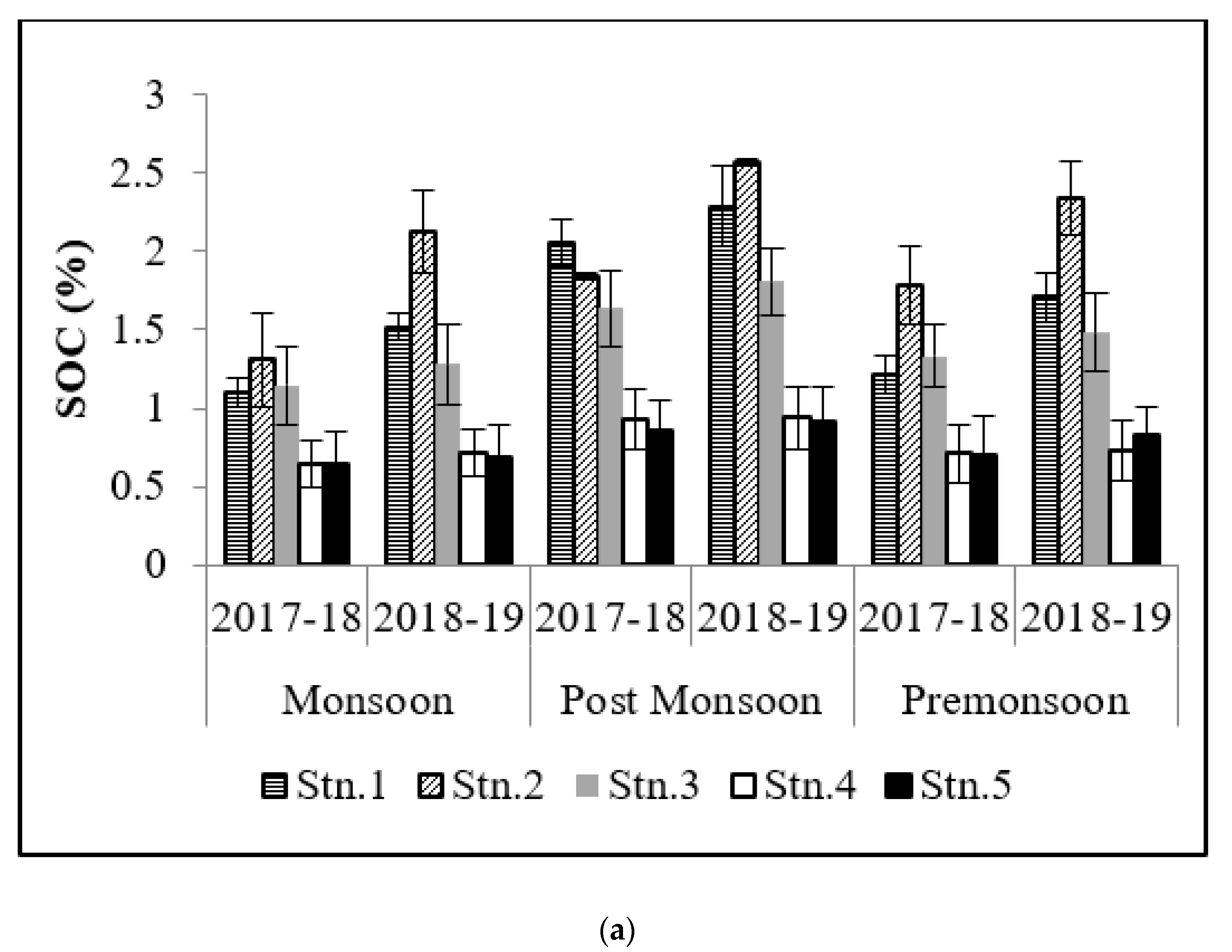
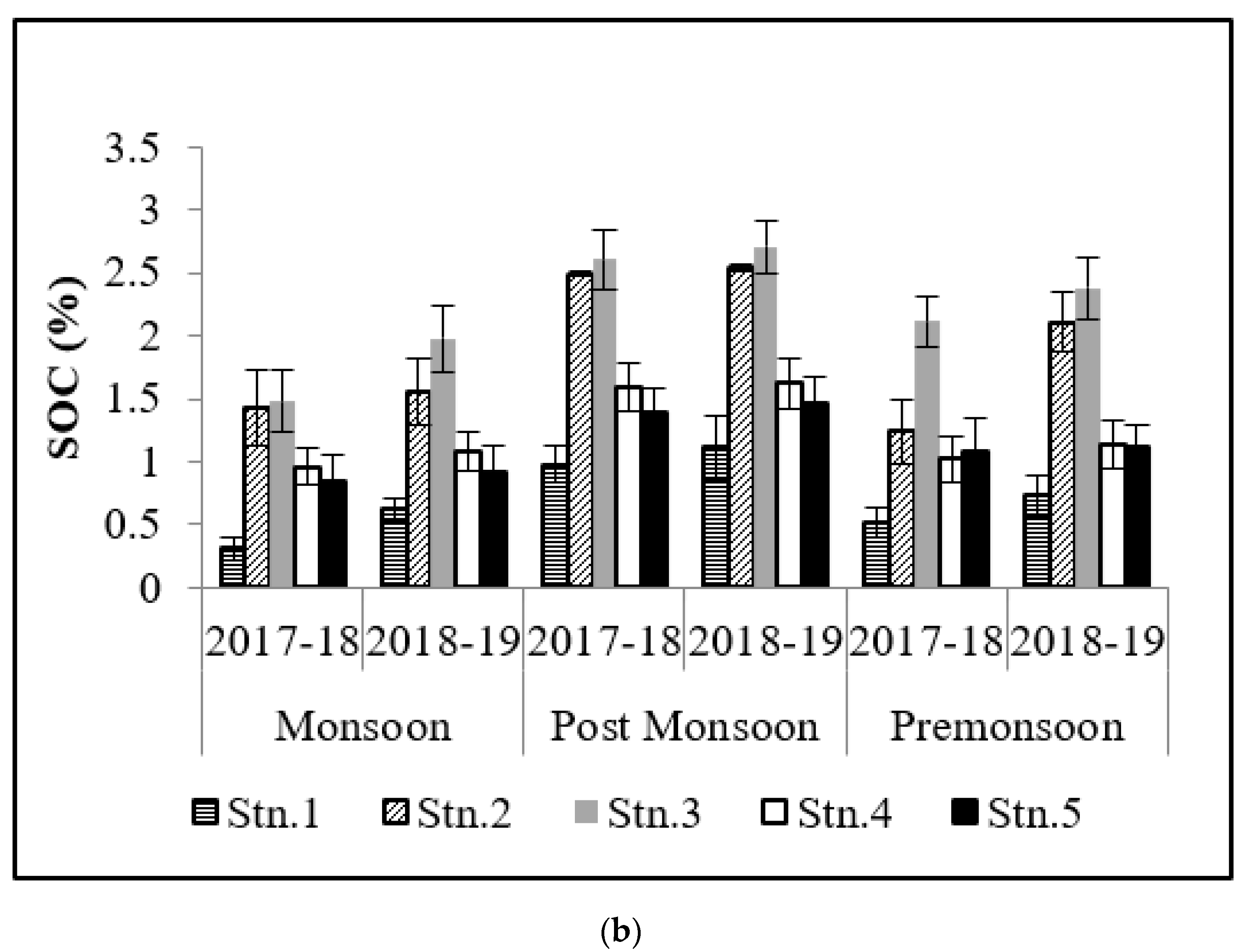
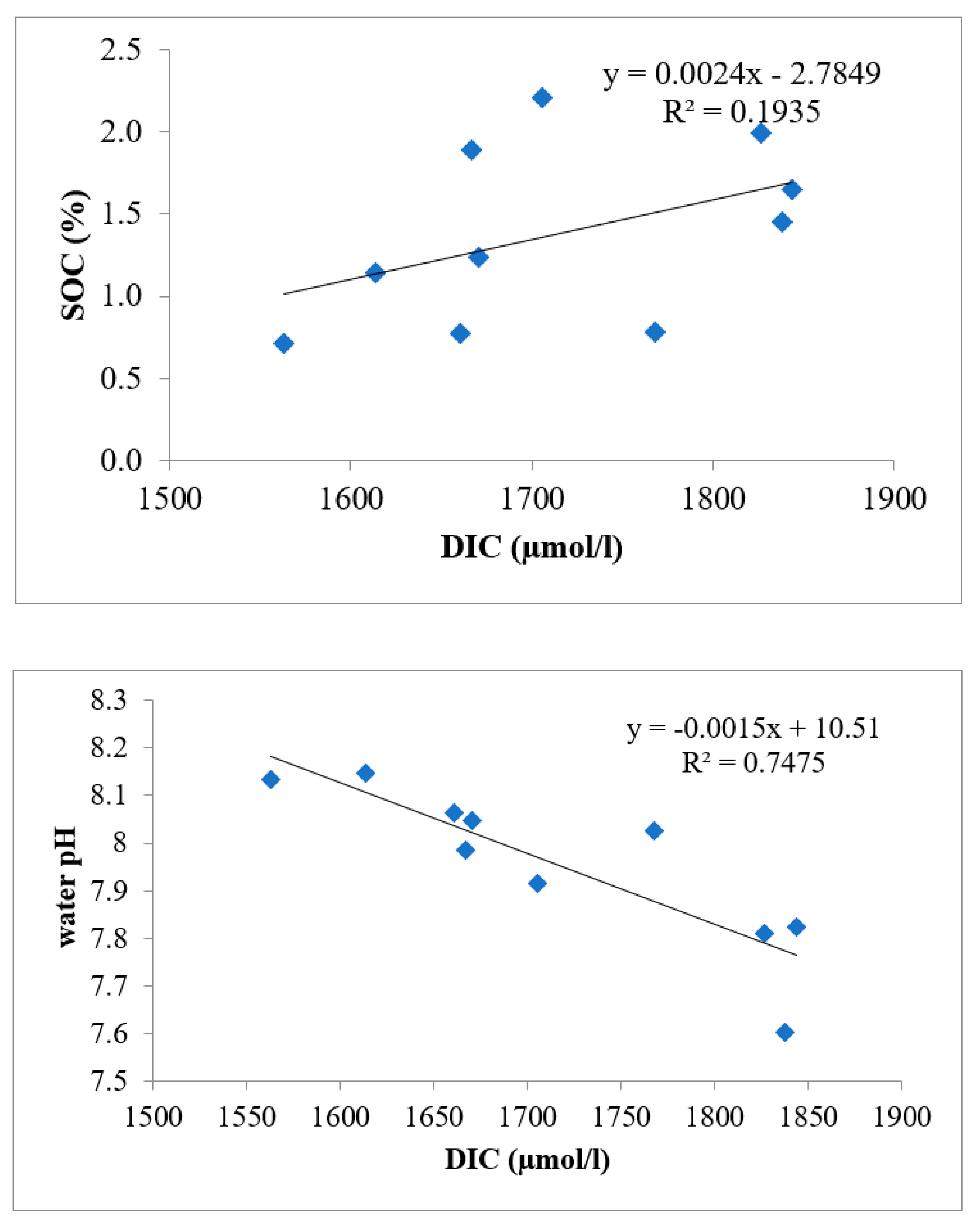
3.1. Soil Organic Carbon (SOC)
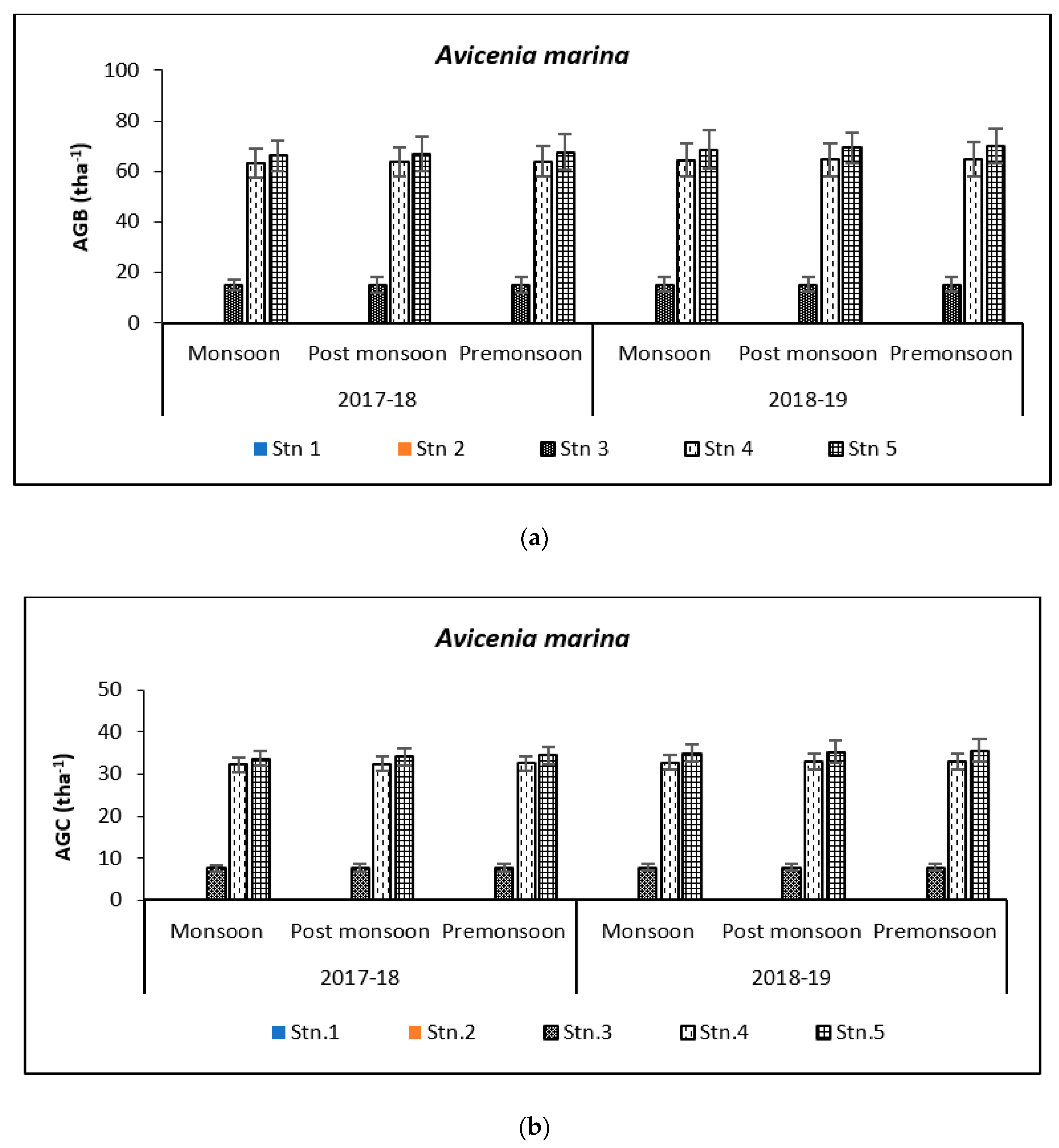
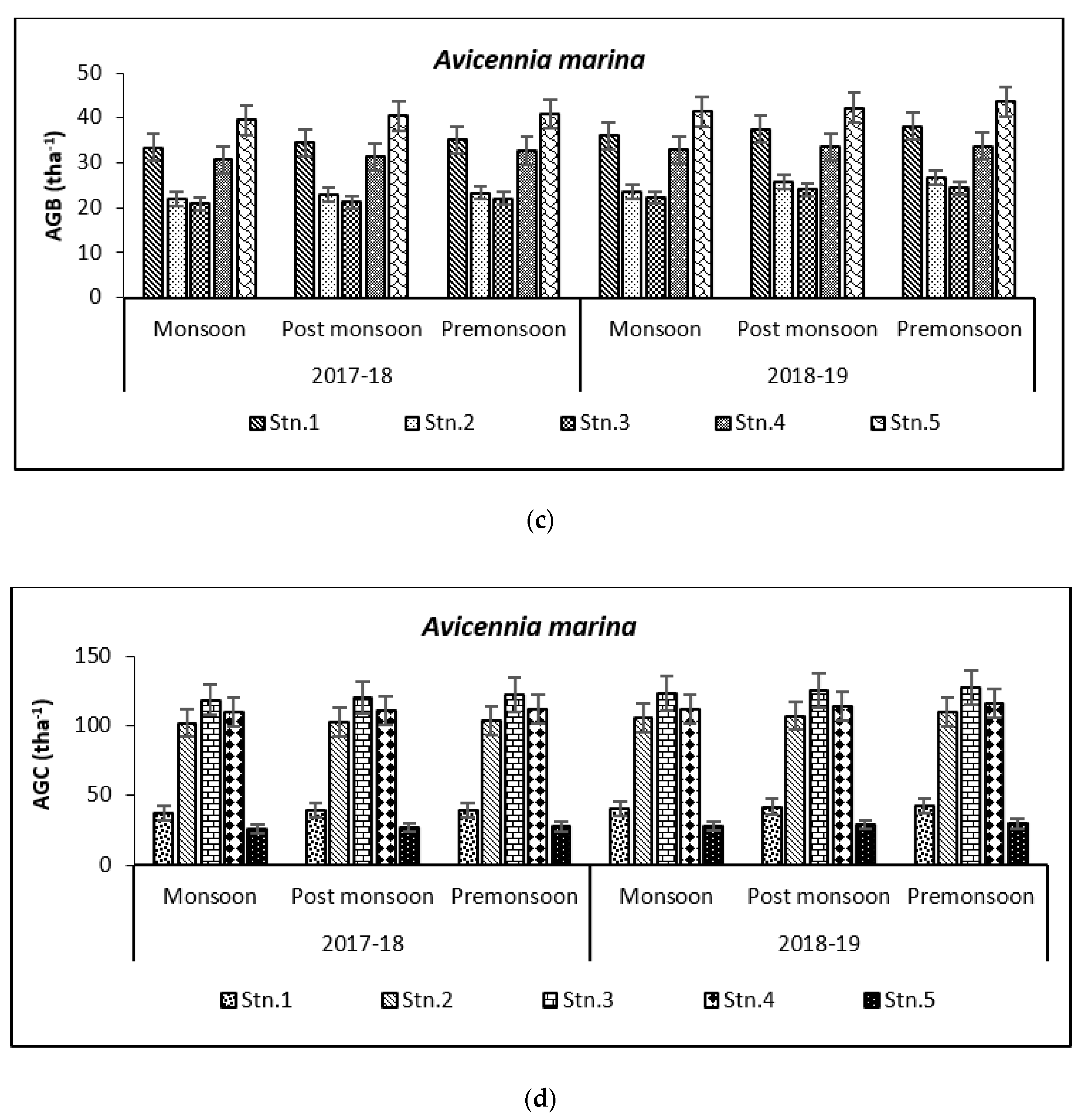
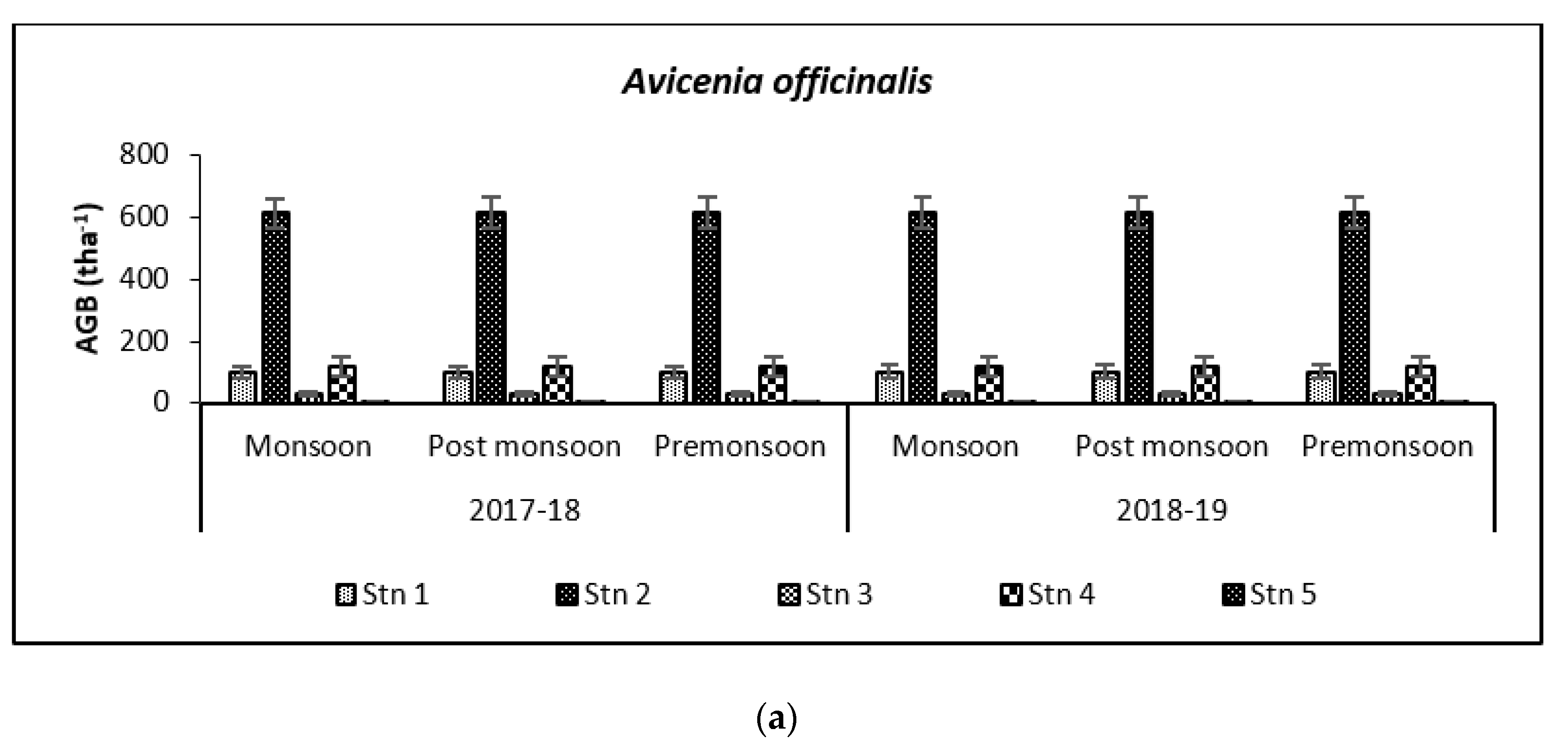
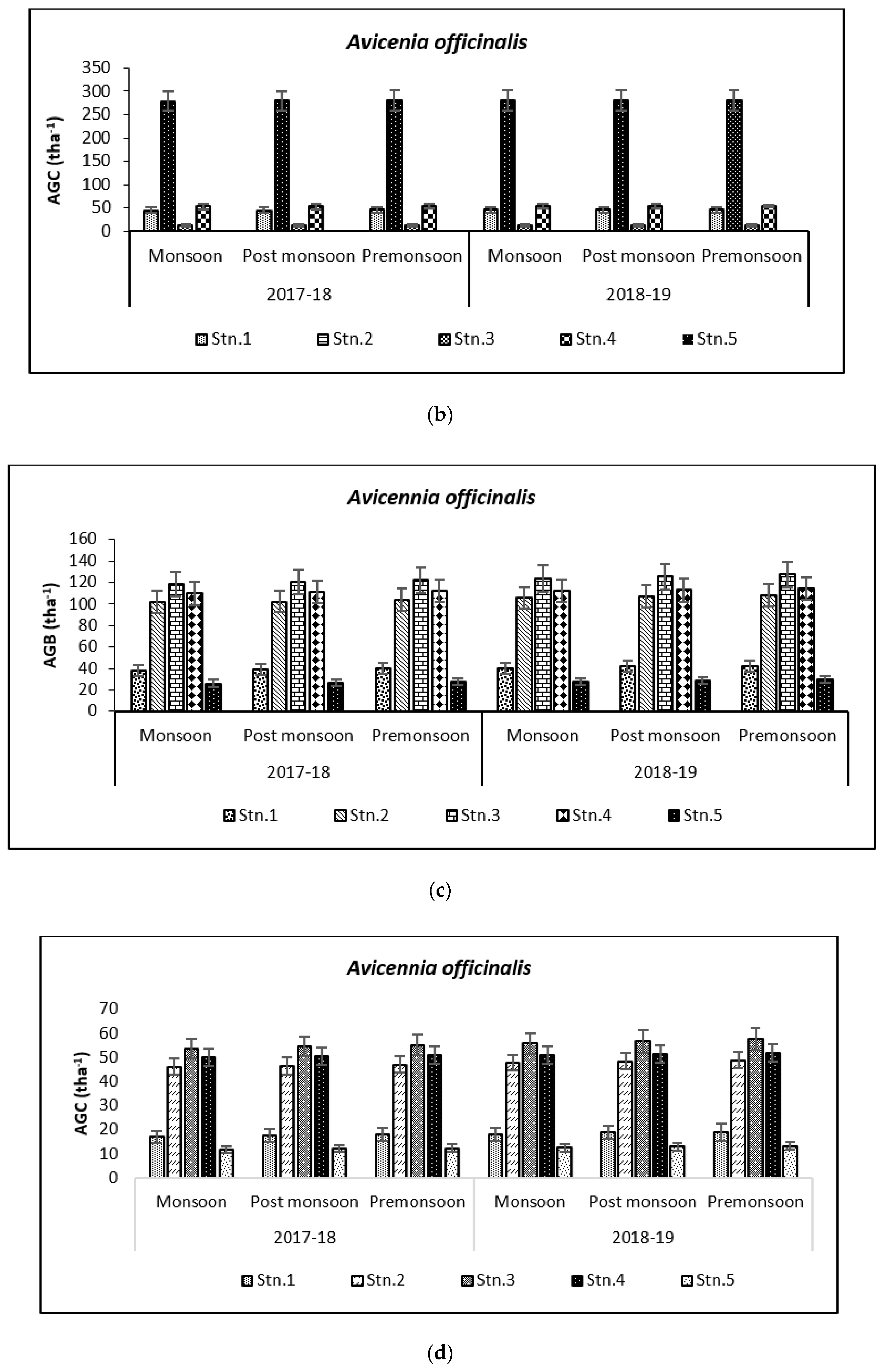
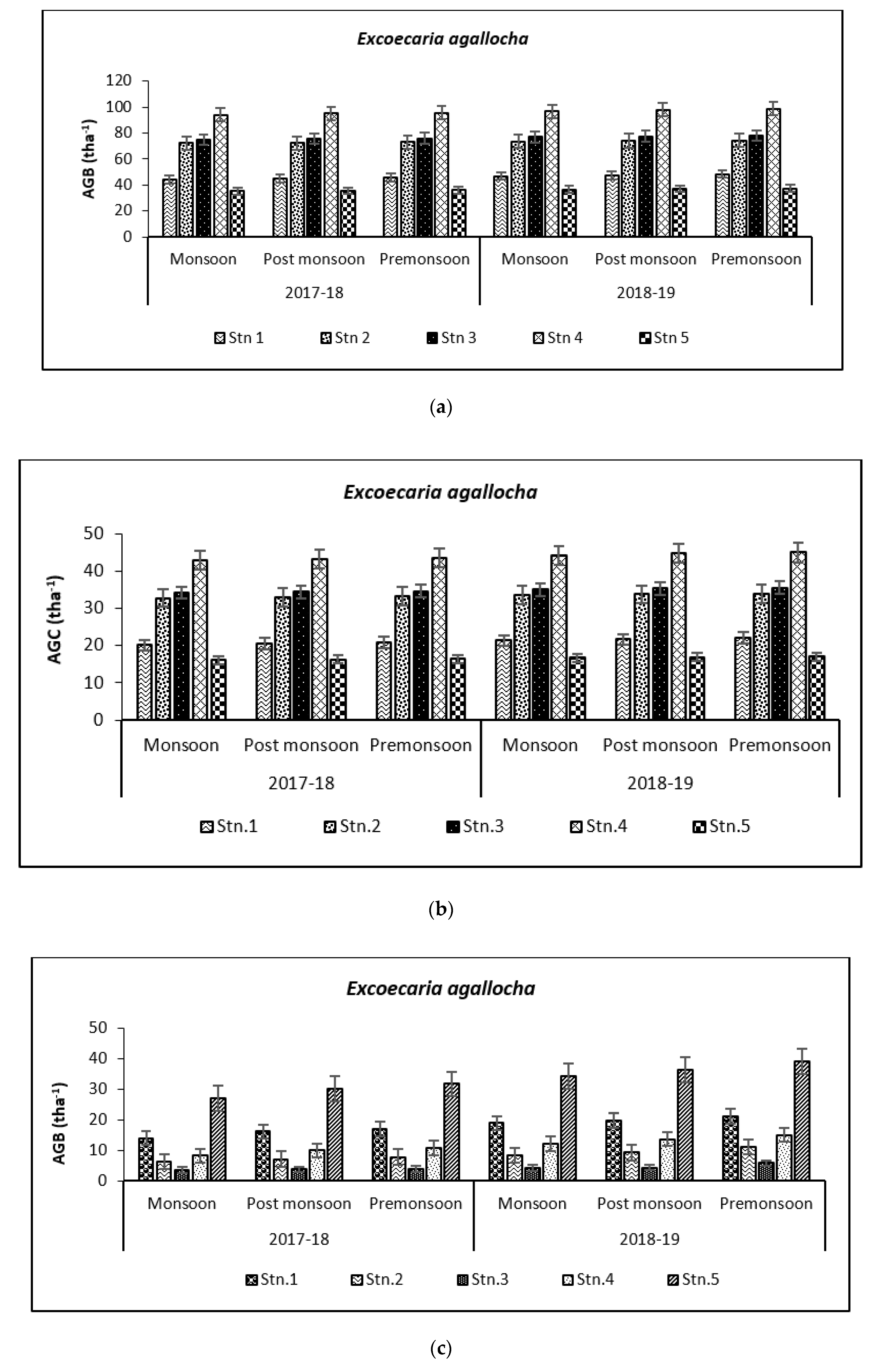
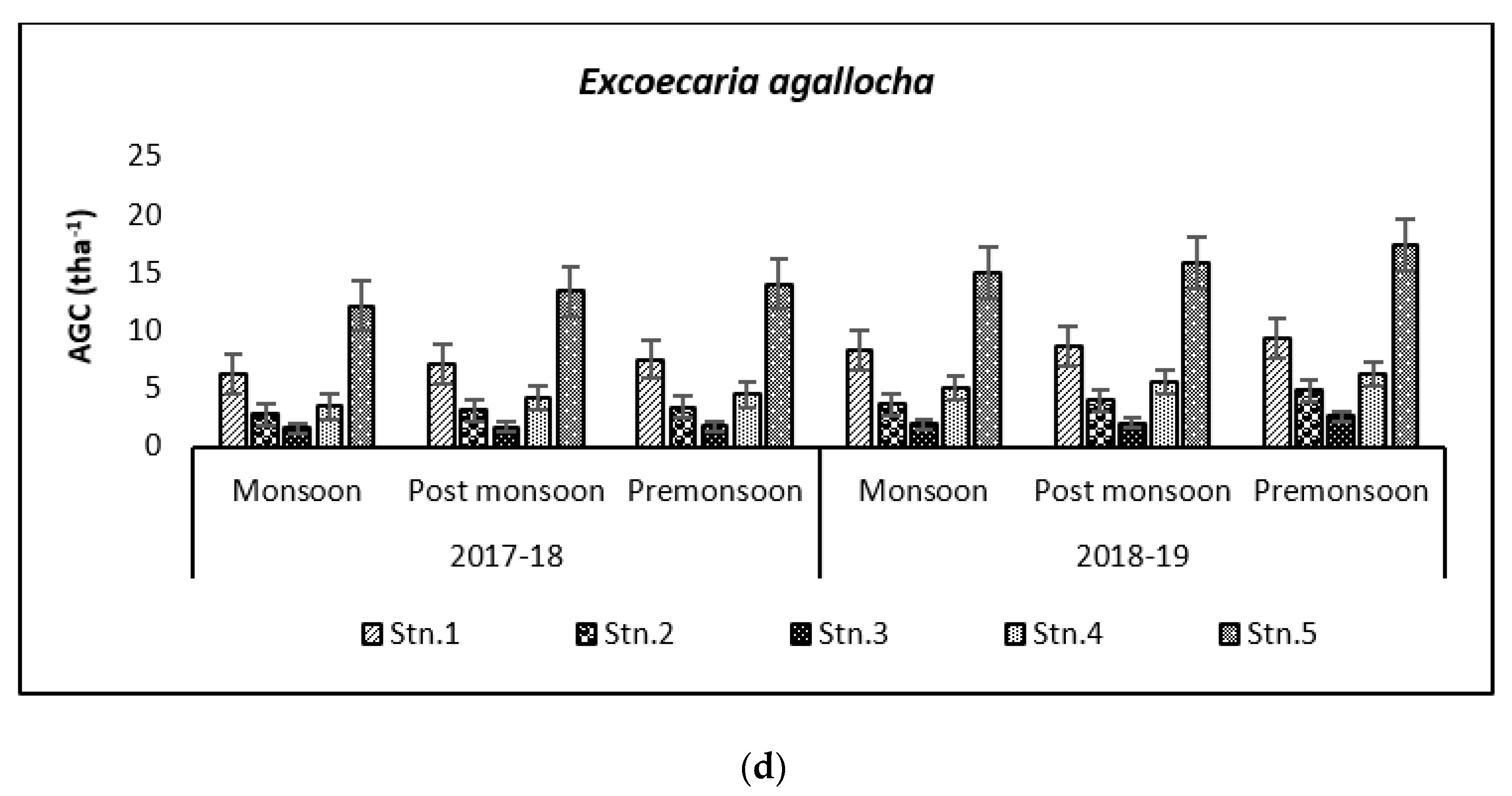
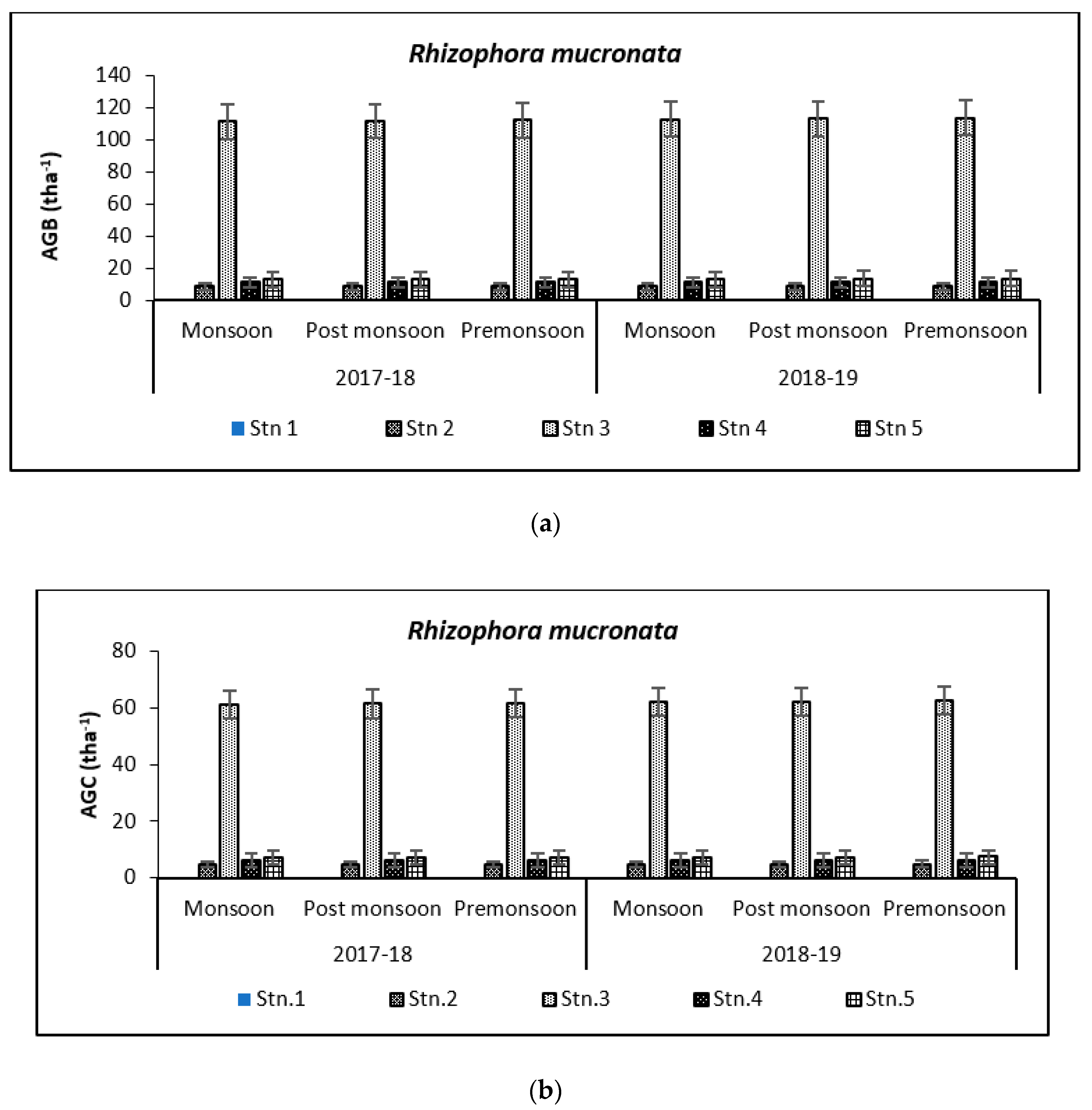
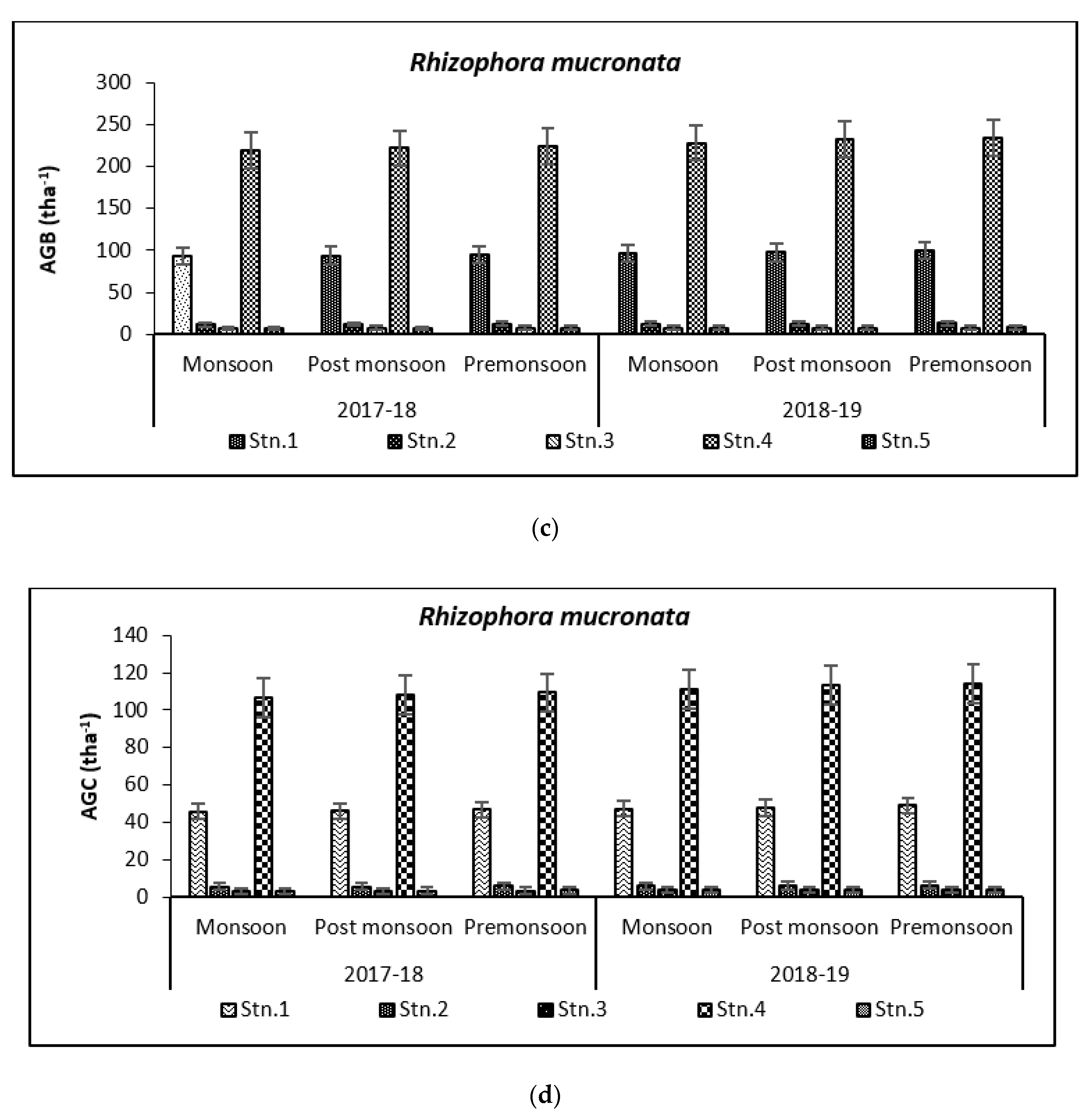
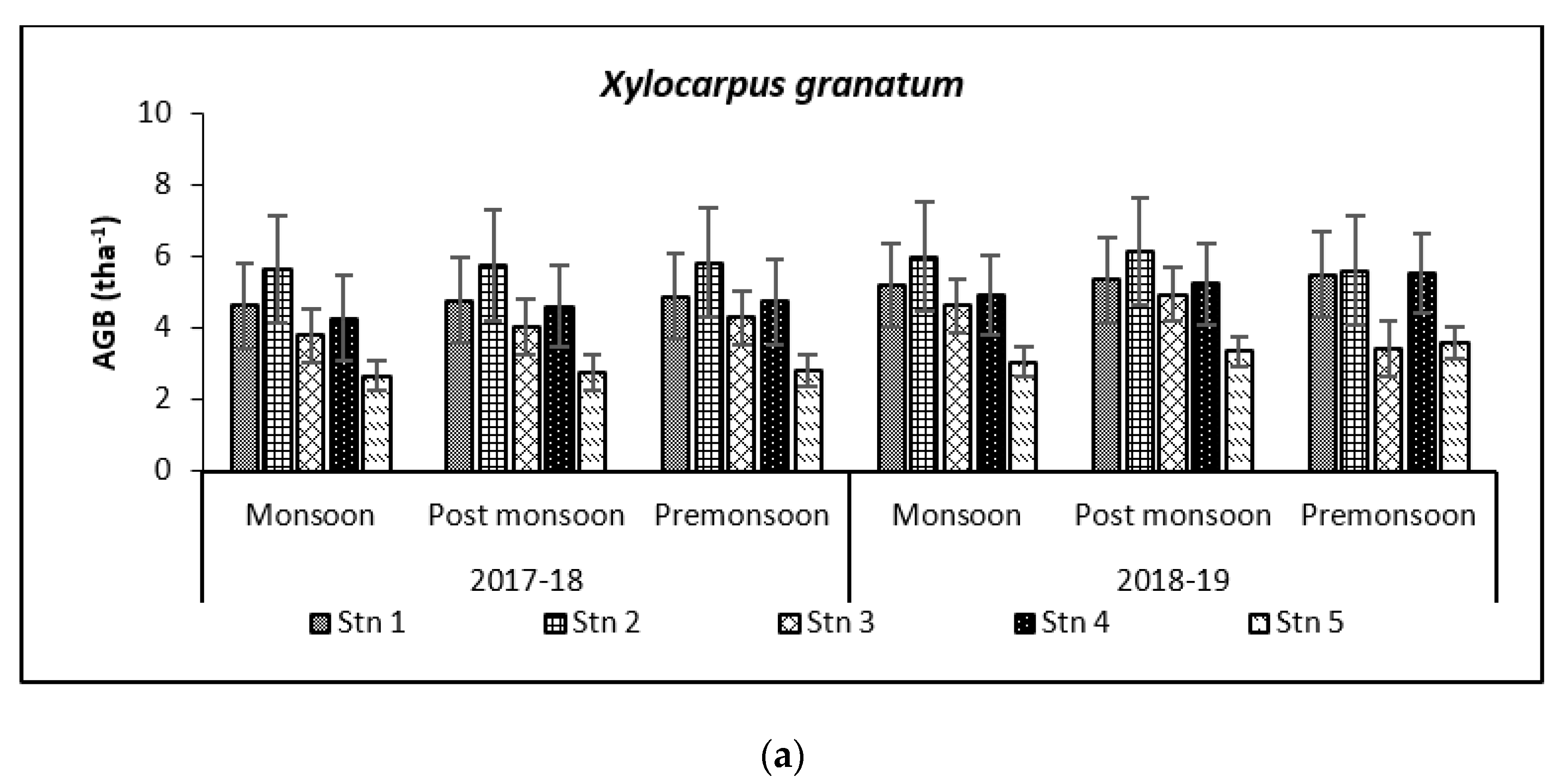
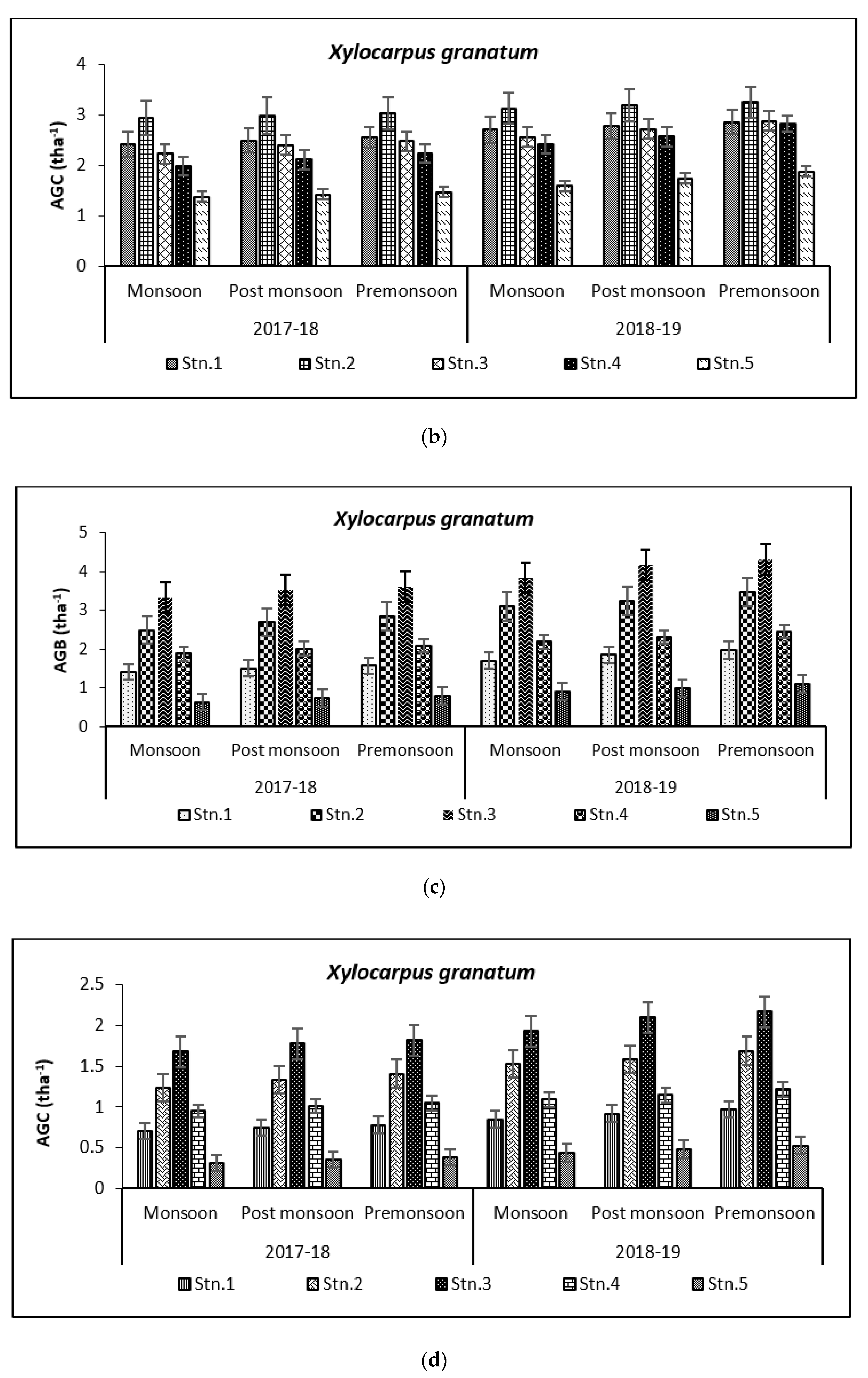
3.2. Carbon Storage in Mangroves (Species Wise)
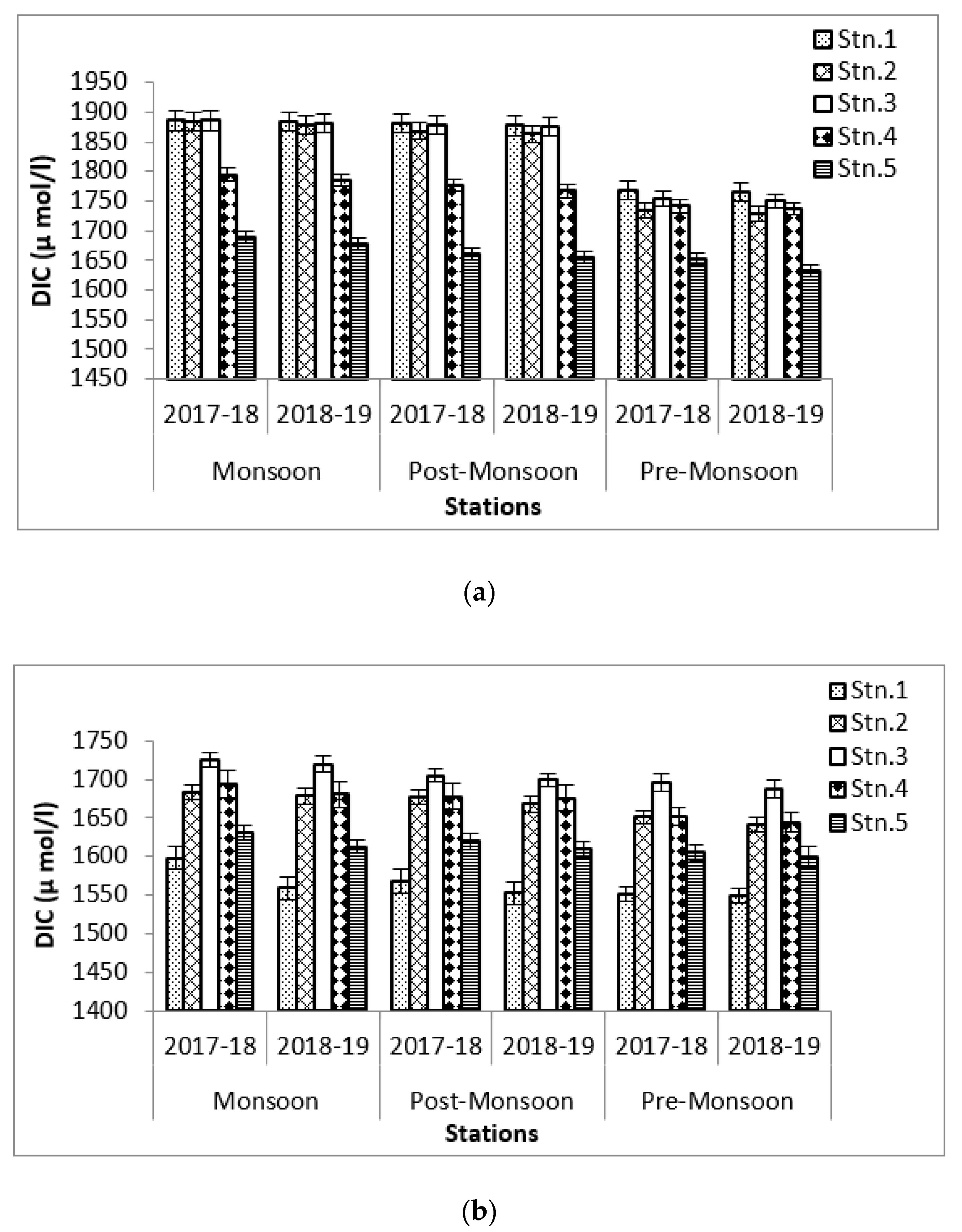
3.3. Carbon Storage in Aquatic Medium
3.4. Prospects in Carbon Storage (Whole Ecosystem)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The IUCN Red List of Threatened Species. Available online: http://www.iucnredlist.org. (accessed on 4 January 2019).
- Duke, N.C. Mangrove floristics and biogeography. In Tropical Mangrove Ecosystems; Coastal and Marine Estuarine Series 41; Robertson, A.I., Alonghi, D.M., Eds.; American Geophysical Union: Washington, DC, USA, 1992; pp. 63–100. [Google Scholar]
- Donato, C.D.; Kauffman, J.B.; Murdiarso, D.; Kurni-Anto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Mathew, G.; Jeyabaskaran, R.; Prema, D. Mangrove ecosystems in India and their conservation. In Coastal Fishery Resources of India-Conservation and Sustainable Utilization; Society of Fisheries Technologists: Kerala, India, 2010; pp. 186–196. [Google Scholar]
- Basha, S.K.M.; Indira Priyadarsini, A.; John Paul, M. Mangrove diversity of south east coast of Andhra Pradesh, India. Int. J. Eng. Res. Technol. 2018, 7, 333–337. [Google Scholar]
- Naskar, K.; Mandal, R. Ecology and Biodiversity of Indian Mangrove (Part-1: Global Status); Daya Publishing House: Delhi, India, 1999; pp. 1–356. [Google Scholar]
- Forest Survey of India (Ministry of Environment and Forests) Dehradun. State Forest Report; Forest Survey of India: Uttarakhand, India, 2017. [Google Scholar]
- Sahu, S.C.; Suresh, H.S.; Murthy, I.K.; Ravindranath, N.H. Mangrove area assessment in India: Implications of loss of mangroves. J. Earth Sci. Clim. Chang. 2016, 6, 1–7. [Google Scholar]
- Banerjee, K.; Chowdhury, M.R.; Sengupta, K.; Sett, S.; Mitra, A. Influence of anthropogenic and natural factors on the mangrove soil of Indian Sundarbans wetland. Arch. Environ. Sci. 2012, 6, 80–91. [Google Scholar]
- Sahoo, K.; Dhal, N.K. Potential microbial diversity in mangrove ecosystems: A review. Indian J. Mar. Sci. 2003, 38, 249–256. [Google Scholar]
- Abu Hena, M.K.; Ashrafu, M.A.K. Coastal and estuarine resources of Bangladesh: Management and conservation issues. Maejo Intern J. Sci. Technol. 2009, 3, 313–342. [Google Scholar]
- Duarte, C.M.; Losada, I.J.; Hendriks, I.E.; Mazarrasa, I.; Marba, N. The role of coastal plant communities for climate change mitigation and adaptation. Nat. Clim. Chang. 2013, 3, 961–968. [Google Scholar] [CrossRef]
- Mitra, A.; Sundaresan, J. How to Study Stored Carbon in Mangroves; CSIR-National Institute of Science Communication: New Delhi, India; Information Resources (NISCAIR): New Delhi, India, 2016; ISBN 978-81-7236-349-9. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and proposed modification of the chromic acid tritation method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Edmond, J.M. High precision determination of titration alkalinity and total carbon dioxide content of seawater by potentiometric titration. Deep-Sea Res. 1970, 17, 737–750. [Google Scholar]
- Goyet, C.; Beauverger, C.; Brunet, C.; Poisson, A. Distribution of carbon dioxide partial pressure in surface waters of the southwest Indian Ocean. Tellus 1991, 43B, 1–11. [Google Scholar]
- D.O.E. Handbook of Methods for Analysis of the Various Parameters of the Carbon Dioxide System in Sea Water, 2nd ed.; Dickson, A.G., Goyet, C., Eds.; ORNL/CDIAC/USDOE: Washington, DC, United States, 1994; p. 74. [Google Scholar]
- Das, S.; Ganguly, D.; Maiti, T.K.; Mukherjee, A.; Jana, T.K.; De, T.K. A Depth-wise diversity of free living N2 fixing and Nitrifying Bacteria and its seasonal variation with nitrogen containing nutrients in the mangrove sediments of Sundarbans, WB, India. Open J. Mar. Sci. 2013, 3, 112–119. [Google Scholar] [CrossRef][Green Version]
- Reddy, H.R.V.; Hariharan, V. Distribution of nutrients in the sediments of the Netravathi Gurupur estuary, Mangalore. Indian J. Fish. 1986, 33, 123–126. [Google Scholar]
- Sverdrup, H.U.; Johnson, M.W.; Fleming, R.H. The Oceans, Their Physics, Chemistry, and General Biology; Printice Hall, Inc.: Hoboken, NJ, USA, 1942; p. 1087. [Google Scholar]
- Mitra, A.; Sengupta, K.; Banerjee, K. Standing biomass and carbon storage of above-ground structures in dominant mangrove trees in the Sundarbans. For. Ecol. Manag. 2011, 261, 1325–1335. [Google Scholar] [CrossRef]
- Canadell, J.G.; Pitelka, L.F.; Ingram, J.S.I. The effects of elevated [CO2] on plant-soil carbon below-ground: A summary and synthesis. Plant Soil 1995, 187, 391–400. [Google Scholar] [CrossRef]
- Hall, G.M.J.; Wiser, S.K.; Allen, R.B.; Beets, P.N.; Goulding, C.J. Strategies to estimate national forest carbon stocks from inventory data: The 1990 New Zealand baseline. Glob. Chang. Biol. 2001, 7, 389–403. [Google Scholar] [CrossRef]
- Joshi, H.G.; Ghose, M. Community structure, species diversity, and aboveground biomass of the Sundarbans mangrove swamps. Trop. Ecol. 2014, 55, 283–303. [Google Scholar]
- Banerjee, K. Decadal change in the surface water salinity profile of Indian Sundarbans: A Potential Indicator of Climate Change. J. Mar. Sci. Res. Dev. 2013, 2, 3. [Google Scholar] [CrossRef]
- Suzuki, E.; Tagawa, H. Biomass of a mangrove forest and a sedge marsh on Shigaki Island, South Japan. Jpn. J. Ecol. 1983, 33, 231–234. [Google Scholar]
- Woodroffe, C.D. Studies of a mangrove basin, Tuff Crater, New Zealand. I: Mangrove biomass and production of detritus, Estuarine. Coast. Shelf Sci. 1985, 20, 265–280. [Google Scholar] [CrossRef]
- Doyen, A. La mangrove a usage multiple de I’estuarine Saloum (Senegal). In Selected Papers of the Dakar Symposium on Acid Sulphate Soils; Publication no. 44; Dost, H., Ed.; International Institute for Land Reclamation and Improvement: Wageningen, The Netherlands, 1986; pp. 176–201. [Google Scholar]
- Imbert, D.; Rollet, B. Phytomass eaerienneet production primairedans la mangrove du Grand Cul-de-Sac Maria (Guadeloupe, Antilles francaises). Bull. D’écol. 1989, 20, 27–39. [Google Scholar]
- Golley, F.; Odum, H.T.; Wilson, R. The structure and metabolism of a Puerto Rican red mangrove forest in May. Ecology 1962, 43, 9–19. [Google Scholar] [CrossRef]
- Christensen, B. Biomass and productivity of Rhizophora apiculata B1 in a mangrove in southern Thailand. Aquat. Bot. 1978, 4, 43–52. [Google Scholar] [CrossRef]
- Lugo, A.E.; Snedaker, S.C. The ecology of mangroves. Annu. Rev. Ecol. Evol. Syst. 1974, 5, 39–64. [Google Scholar] [CrossRef]
- Kathiresan, K.; Gomathi, V.; Anburaj, R.; Saravanakumar, K.; Asmathunisha, N.; Sahu, S.K.; Shanmugaarasu, V.; Anandhan, S. Carbon sequestration potential of Rhizophora mucronata and A. marina as influenced by age, season, growth and sediment characteristics in southeast coast of India. J. Coast Conserv. 2013, 17, 397. [Google Scholar] [CrossRef]
- Komiyama, A.; Moriya, H.; Prawiroatmodjo, S.; Toma, T.; Ogino, K. Forest primary productivity. In Biological System of Mangrove; Ogino, K., Chihara, M., Eds.; Ehime University: Matsuyama, Japan, 1988; pp. 97–117. [Google Scholar]
- Putz, F.; Chan, H.T. Tree growth, dynamics, and productivity in a mature mangrove forest in Malaysia. For. Ecol. Manag. 1986, 17, 211–230. [Google Scholar] [CrossRef]
- Amarasinghe, M.D.; Balasubramaniam, S. Net primary productivity of two mangrove forests stands on the northwest coast of Sri Lanka. Hydrobiologia 1992, 247, 37–47. [Google Scholar] [CrossRef]
- Mall, L.P.; Singh, V.P.; Garge, A. Study of biomass, litter fall, litter decomposition and soil respiration in monogenetic mangrove and mixed mangrove forests of Andaman Islands. Trop. Ecol. 1991, 32, 144–152. [Google Scholar]
- Camacho, L.D.; Gevana, D.T.; Carandang, A.P.; Camacho, S.C.; Combalicer, E.A.; Rebugio, L.L.; Youn, Y.C. Tree biomass and carbon stock of a community managed mangrove forest in Bohol, Philippines. For. Sci. Technol. 2011, 7, 161–167. [Google Scholar] [CrossRef]
- Ren, H.; Chen, H.; Li, Z.A.; Han, W. Biomass accumulation and carbon storage of four different aged Sonneratia apetala plantations in Southern China. Plant Soil 2010, 327, 279–291. [Google Scholar] [CrossRef]
- Plaza, C.; Courtier-Murias, D.; Fernandez, J.M.; Polo, A.; Simpson, A.J. Physical, chemical and biochemical mechanisms of soil organic matter stabilization under conservation tillage systems: A central role for microbes and microbial by-products in C sequestration. Soil. Biol. Biochem. 2013, 57, 124–134. [Google Scholar] [CrossRef]
- Justrow, J.; Miller, R. Soil Processes and the Carbon Cycle; Rattan Lal, J.M.K., Ronald, F.F., Bobby, S., Eds.; CRC Press: Boca Raton, FL, USA, 1997; pp. 207–223. [Google Scholar]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar]
- Kuzyakov, Y.; Friedel, J.K.; Stahr, K. Review of mechanisms and quantification of priming effects. Soil Biol. Biochem. 2000, 32, 1485–1498. [Google Scholar] [CrossRef]
- Conde, E.; Cardenas, M.; Ponce-Mendoza, A.; Luna-Guido, M.L.; Cruz-Mondragón, C.; Dendooven, L. The impacts of inorganic nitrogen application on mineralization of 14C labelled maize and glucose, and on priming effect in saline alkaline soil. Soil Biol. Biochem. 2005, 37, 681–691. [Google Scholar] [CrossRef]
- Olafsson, J.; Olafsdittir, S.R.; Benoit-Cattin, A.; Takahashi, T. The Irminger sea and the Iceland sea time series measurements of sea water carbon and nutrient chemistry 1983–2008. Earth Syst. Sci. Data 2010, 2, 99–104. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stations | Coordinates | Site Description | |
|---|---|---|---|
| Latitude (N) | Longitude (E) | ||
| (a) | |||
| Stn.1 | 20°44′21.37″ | 86°52′00.00″ | It is a protected mixed natural mangrove forest dominated by H. fomes, E. agallocha, and A. officinalis. The vegetation has a mean stand density of 60 trees per 100 m2, mean DBH is 0.50 m, and mean height of 12 m. The area is a tourist site that receives a freshwater discharge from rivers like Bramhani, Baitarani, and the distributaries of Bhitarkanika river and different creeks, channels, etc. Eighty percent of crocodile nesting occurs here and it has huge reptile diversity like snakes, turtles, monitor lizards, etc. Owing to dense mangrove vegetation, the sediment is often black in color. |
| Stn.2 | 20°42′56.85″ | 86°51′48.40″ | This site is similar to Dangmal in all conditions (climatic, vegetation, and river-fed) but it is more pristine and undisturbed w.r.t mangrove habitat. This site is famous for crocodile nesting and bird watching sites, Baga–gahana. The biomass of the A. officinalis is very high in comparison to all other study sites. The vegetation has a mean stand density of 80 trees per 100 m2, mean DBH is 0.60 m, and mean height of 12 m. The soil is also very rich in organic matter derived from mangrove litter. |
| Stn.3 | 20°38′38.81″ | 86°52′20.29″ | This site is an anthropogenically stressed area in comparison to other stations. The area is mostly surrounded by villages and agriculture and aquaculture are the main occupations of the residents. Other activities include ecotourism, transportation, fishing, and household pollution. This is the gateway for BWLS. The distributions of mangrove species are unequal and are found in patches due to cutting and plantation. The dominant species include P. paludosa, E. agallocha and R. mucronata. The vegetation has a mean stand density of 70 trees per 100 m2, mean DBH is 0.40 m, and mean height of 8 m. This site also comes under subtropical and humid climatic zones. The area receives a lot of anthropogenic wastes from adjoining villages, agriculture, and aquaculture discharges. |
| Stn.4 | 20°41′08.12″ | 86°59′16.03″ | The site is near the sea mouth. One side is open to the Bay of Bengal and the other side is on the bank of river Bausagada. Both the ends of this river are open to the sea, hence it is always tide fed and maintains higher salinity than other study stations. This site also faces anthropogenic pressures like cutting of forest trees, fishing, ecotourism, and other anthropogenic effects. Plastic pollution is very high on this site, mostly on the oceanic coast. The area was dominated by E. agallocha, A. marina, L. racemosa and C. decandra. The vegetation has a mean stand density of 180 trees per 100 m2, mean DBH is 0.30 m, and mean height 6 m. Owing to the position of the station, the station is very dynamic with fresh water on one side and marine water on the other. Organic matter load is comparatively higher. |
| Stn.5 | 20°42′16.91″ | 86°01′56.34″ | The site is called Ekakula, which means one mouth is open to the rivers Bausagada and Patsala. Both sides are open to the Bay of Bengal, hence it always maintains the highest salinity. The site is dominated by high salt-tolerant species like A. marina, E. agallocha, A. corniculatum, A. rotundifolia, A. alba, R. mucronata, and S. alba. The vegetation has a mean stand density of 120 trees per 100 m2, the mean DBH is 0.15 m, and the average height of the tree is below 7 m but the density of the species is more. The soil is usually loose, sandy in character, but due to the high density of mangroves has rich litterfall. |
| (b) | |||
| Stn.1 | 20°25′50.05″ | 86°43′50.21″ | It is a Proposed Forest (PF) block with an area of 369.75 ha and surrounded by the Gobari river in the south and Chataka in the east, Kandarapatia PRF block in the north, and Gobari river with Jambu village in the west. The waterway plays an important role by inundating the forest block diversity. The dominant species are Avicennia marina Ceriops decandra, Excocecaria agallocha, Acanthus ilicifolius and Avicennia officinalis. Aegiceras corniculatum, Avicennia alba, and Rhizophora mucronata are also found in small numbers. Xylocarpus granatum is rare in this site. Dalbergia spinosa, Sonneratia apetala, Tamarix troupii, Aegialitis rotundifolia and Phoenix paludosa are found in this forest block. One of the non-mangrove species Casuarina species forest patches is also found in this region. The vegetation has a mean stand density of 55 trees per 100 m2, mean DBH is 0.60 m, and mean height 10 m. The forest block is degraded by agricultural runoff, anthropogenic activity, shrimp culture, and grazing. |
| Stn.2 | 20°22′48.89″ | 86°45′09.31″ | This forest block is the highest area among the selected study sites which comprises about 1394.744 ha. This study site comes under Proposed Reserved Forest (PRF) block. Distributaries of the Gobari river play a crucial role in mangrove growth and regeneration. Kharnasi River separates Kansaridia and Bhitara Kharnasi forest block in the west and one of the creek formations called Kalpana Jore, in the northern part nearer to the Bay of Bengal called as the Chataka and extending to the east. In the southern part, Hetamundia forest and Kajalapatia village are present. |
| Excoecaria agallocha is the dominant species on this site. Rhizophora mucronata, Ceriops decandra, and Brownlowia tersa are mostly dominant in the area. Xylocarpus granatum, Rhizophora apiculata, Avicennia officinalis, Avicennia marina and Dalbergia spinosa are moderately distributed. Apart from that other species present are Agiceras corniculatum, Acanthus ilicifolius, Sonneratia apetala, Avicennia alba, Kandelia candel, Bruguiera cylindrical and Bruguiera gymnorrhiza. The vegetation has a mean stand density of 50 trees per 100 m2, mean DBH is 0.35 m, and mean height of 7 m. The forest block exhibits rich species diversity but it is in a degraded state due to anthropogenic activity and overexploitation. | |||
| Stn.3 | 20°22′03.197″ | 86°43′13.18″ | Kharnasi forest block is classified as Bhitara Kharnasi Reserved Forest (RF-A) and Bahara Kharinasi Reserved Forest (RF-B). Our study site is the Bhitara Kharnasi (RF-A), which has a total area of 577.072 ha. It is surrounded by the distributaries of Kharnasi river and Gobari river which meet the Bay of Bengal. Kharnasi River separates the Bhitara Kharnasi and Bahara Kharnasi in the western part. In the north, the name of the water body is called Chataka which connects with the Bay of Bengal. Kansaridia and Sanatubi forest blocks are present in the east and south respectively. Distributaries play an important role by flushing fresh water inside the region and making it a dense forest. |
| Excocecaria agallocha is a dominant species in this region followed by Avicennia officinalis, Heritiera fomes, Brownlowia tersa, Ceriops decandra, Rhizophora mucronata, Avicennia marina and Xylocarpus granatum are small in number along with Sonneratia apetala, Avicennia alba, Bruguiera gymnorrhiza and Phoenix paludosa. The vegetation has a mean stand density of 80 trees per 100 m2, mean DBH is 0.45 m, and mean height of 9 m. Anthropogenic activities, shrimp culture, and polluted agricultural flushing by the waterways are the main reasons for the diminishing of mangrove growth. | |||
| Stn.4 | 20°27′53.61″ | 86°41′15.41″ | This forest block comes under the Reserve Forest (RF) block with an area of about 137.784 ha. This forest patch is a mixture of natural and plantation forests. The plantation program was done by MSSRF with the help of local villagers and the Forest department of Odisha. The forest block is surrounded by the Gobari river in the south, the Chataka and distributaries of the Jagjore river in the east and north respectively, and Kantilo village with Luna nai which falls in the Gobari river in the western part of the forest block. To get a sufficient tidal inundation, canals have been excavated in the Kantilo forest block. |
| The dominant species at this site is Rhizophora mucronata. Avicennia marina, Avicennia officinalis, Excocecaria agallocha are also found in numbers. Xylocarpus granatum is rare and Ceriops decandra, Agiceras corniculatum, Acanthus ilicifolius, Sonneratia apetala, Avicennia alba, Kandelia candel, Tamarix troupii, Phoenix paludosa, Bruguiera parviflora and Sonneratia alba are also found in the region. The vegetation has a mean stand density of 150 trees per 100 m2, mean DBH is 0.30 m, and mean height of 8 m. | |||
| The main causes of mangrove destruction are anthropogenic activities, agricultural runoff, grazing, shrimp culture and plastics, polythene, etc., inside the forest area. | |||
| Stn.5 | 20°26′52.77″ | 86°43′32.83″ | This forest block comes under the Proposed Reserve Forest (PRF), which has represented an area of about 105.668 ha. It is surrounded by Jambu forest block in the south, the Bay of Bengal in the east (Chataka), the Jagajhore river at Jagajhore in the north, and Kandarapatia village with a non-mangrove land patch in the west. The forest block is inundated during high tide which is favorable for the distribution of species composition. |
| Excoecaria agallocha is dominant in the study area followed by Avicennia marina and Avicennia officinalis along with Ceriops decandra. A very small number of Rhizophora mucronata is found near the saline embankment of the site. Xylocarpus granatum is rare in this region. Other species found are Aegiceras corniculatum, Avicennia alba, Bruguiera cylindrica, Tamarix troupii, Aegialitis rotundifolia, Phoenix paludosa and Pongamia pinnata. The vegetation has a mean stand density of 70 trees per 100 m2, a mean DBH is 0.30 m, and a mean height of 7 m. Runoff from agricultural land, polluted water from shrimp culture, grazing, and anthropogenic activities are the main cause for the destruction of this dense mangrove patch. | |||
| Species | Biomass Contribution by Different Floral Components towards AGB (in %) | Carbon Contribution by Different Floral Components towards AGC (in %) | ||||||
|---|---|---|---|---|---|---|---|---|
| Stem | Branch | Leaf | Stilt Roots | Stem | Branch | Leaf | Stilt Roots | |
| A. marina | 63.01 ± 19.16 | 34.45 ± 17.92 | 2.54 ± 1.38 | – | 28.98 ± 8.81 | 15.16 ± 7.88 | 1.07 ± 0.58 | – |
| (46 ± 0.4) * | (44 ± 0.3) * | (42 ± 0.4) * | ||||||
| A. officinalis | 77.58 ± 12.39 | 20.81 ± 11.50 | 1.61 ± 0.90 | – | 41.12 ± 6.57 | 9.78 ± 5.40 | 0.80 ± 0.45 | – |
| (53 ± 0.3) * | (47 ± 0.2) * | (50 ± 0.8) * | ||||||
| E. agallocha | 57.81 ± 21.69 | 39.97 ± 20.69 | 2.22 ± 1.25 | – | 26.59 ± 9.98 | 17.99 ± 9.31 | 0.98 ± 0.55 | – |
| (46 ± 0.4) * | (45 ± 0.2) * | (44 ± 0.3) * | ||||||
| R. mucronata | 42.51 ± 21.22 | 19.69 ± 8.48 | 3.65 ± 1.73 | 34.14 ± 16.10 | 23.81 ± 11.88 | 10.63 ± 4.58 | 1.79 ± 0.85 | 18.78 ± 8.85 |
| (56 ± 0.4) * | (54 ± 0.2) * | (49 ± 0.4) * | (55 ± 0.6) * | |||||
| X. granatum | 71.22 ± 13.92 | 21.28 ± 10.30 | 7.50 ± 3.70 | 36.98 ± 7.23 | 10.26 ± 4.96 | 3.39 ± 1.67 | ||
| (51.93 ± 0.4) * | (48.2 ± 0.2) * | (45.2 ± 0.4) * | ||||||
| Species | Biomass Contribution by Different Floral Components towards AGB (in %) | Carbon Contribution by Different Floral Components towards AGC (in %) | ||||||
|---|---|---|---|---|---|---|---|---|
| Stem | Branch | Leaf | Stilt Roots | Stem | Branch | Leaf | Stilt Roots | |
| A. marina | 79.03 ± 7.43 | 20.07 ± 7.10 | 0.90 ± 0.47 | – | 34.62 ± 9.31 | 8.39 ± 6.38 | 0.37 ± 0.18 | – |
| (43.8 ± 0.5) * | (41.8 ± 0.2) * | (41.4 ± 0.3) * | ||||||
| A. officinalis | 89.53 ± 4.30 | 9.91 ± 3.67 | 1.28 ± 0.68 | – | 40.92 ± 5.17 | 3.89 ± 2.49 | 0.61 ± 0.25 | – |
| (45.7 ± 0.3) * | (39.3 ± 0.2) * | (47.3 ± 0.6) * | ||||||
| E. agallocha | 56.89 ± 19.43 | 37.37 ± 16.96 | 5.75 ± 2.64 | – | 26.74 ± 8.48 | 12.33 ± 5.31 | 2.71 ± 0.95 | – |
| (47 ± 0.5) * | (33 ± 0.3) * | (47.2 ± 0.4) * | ||||||
| R. mucronata | 44.90 ± 4.41 | 22.54 ± 3.10 | 3.04 ± 0.42 | 29.52 ± 1.11 | 23.35 ± 10.28 | 7.84 ± 3.28 | 1.23 ± 0.66 | 14.02 ± 5.84 |
| (52 ± 0.2) * | (34.8 ± 0.4) * | (40.3 ± 0.3) * | (47.5 ± 0.5) * | |||||
| X. granatum | 71.93 ± 13.88 | 20.43 ± 10.19 | 7.65 ± 3.76 | 36.68 ± 6.48 | 9.34 ± 4.41 | 3.37 ± 1.32 | – | |
| Stations | AGC (tha–1) | SOC (tha–1) | TC (tha–1) | CO2 Equivalent (t) |
|---|---|---|---|---|
| Stn.1 (B) | 62.43 ± 16.96 | 6.68 ± 0.47 | 69.11 ± 17.43 | 253.63 ± 63.97 |
| (M) | 83.39 ± 15.74 | 3.61 ± 0.30 | 87.00 ± 16.04 | 319.29 ± 58.87 |
| Stn.2 (B) | 314.76 ± 110.34 | 7.71 ± 0.45 | 322.47 ± 110.79 | 1183.46 ± 406.60 |
| (M) | 65.85 ± 16.90 | 7.55 ± 0.56 | 73.40 ± 17.46 | 269.38 ± 64.08 |
| Stn.3 (B) | 111.59 ± 21.97 | 5.83 ± 0.24 | 117.42 ± 22.21 | 430.93 ± 81.51 |
| (M) | 70.02 ± 20.77 | 7.71 ± 0.45 | 77.73 ± 21.22 | 285.27 ± 77.88 |
| Stn.4 (B) | 133.96 ± 22.12 | 3.52 ± 0.12 | 137.48 ± 22.24 | 504.55 ± 81.62 |
| (M) | 176.19 ± 42.13 | 5.36 ± 0.29 | 181.55 ± 42.42 | 666.29 ± 155.68 |
| Stn.5 (B) | 57.78 ± 12.27 | 3.55 ± 0.11 | 61.33 ± 12.38 | 225.08 ± 45.43 |
| (M) | 45.96 ± 6.52 | 5.39 ± 0.25 | 51.35 ± 6.77 | 188.45 ± 24.85 |
| Mean ± SD (B) | 136.10 ± 36.73 | 5.46 ± 1.68 | 141.56 ± 38.41 | 519.53 ± 140.96 |
| (M) | 93.22 ± 21.56 | 5.92 ± 1.54 | 99.14 ± 23.07 | 345.75 ± 84.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banerjee, K.; Mitra, A.; Villasante, S. Carbon Cycling in Mangrove Ecosystem of Western Bay of Bengal (India). Sustainability 2021, 13, 6740. https://doi.org/10.3390/su13126740
Banerjee K, Mitra A, Villasante S. Carbon Cycling in Mangrove Ecosystem of Western Bay of Bengal (India). Sustainability. 2021; 13(12):6740. https://doi.org/10.3390/su13126740
Chicago/Turabian StyleBanerjee, Kakoli, Abhijit Mitra, and Sebastián Villasante. 2021. "Carbon Cycling in Mangrove Ecosystem of Western Bay of Bengal (India)" Sustainability 13, no. 12: 6740. https://doi.org/10.3390/su13126740
APA StyleBanerjee, K., Mitra, A., & Villasante, S. (2021). Carbon Cycling in Mangrove Ecosystem of Western Bay of Bengal (India). Sustainability, 13(12), 6740. https://doi.org/10.3390/su13126740