
Kolliker’s Organ and Its Functional Role in the Development of Corti’s Organ and Auditory Systems

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Search Methods
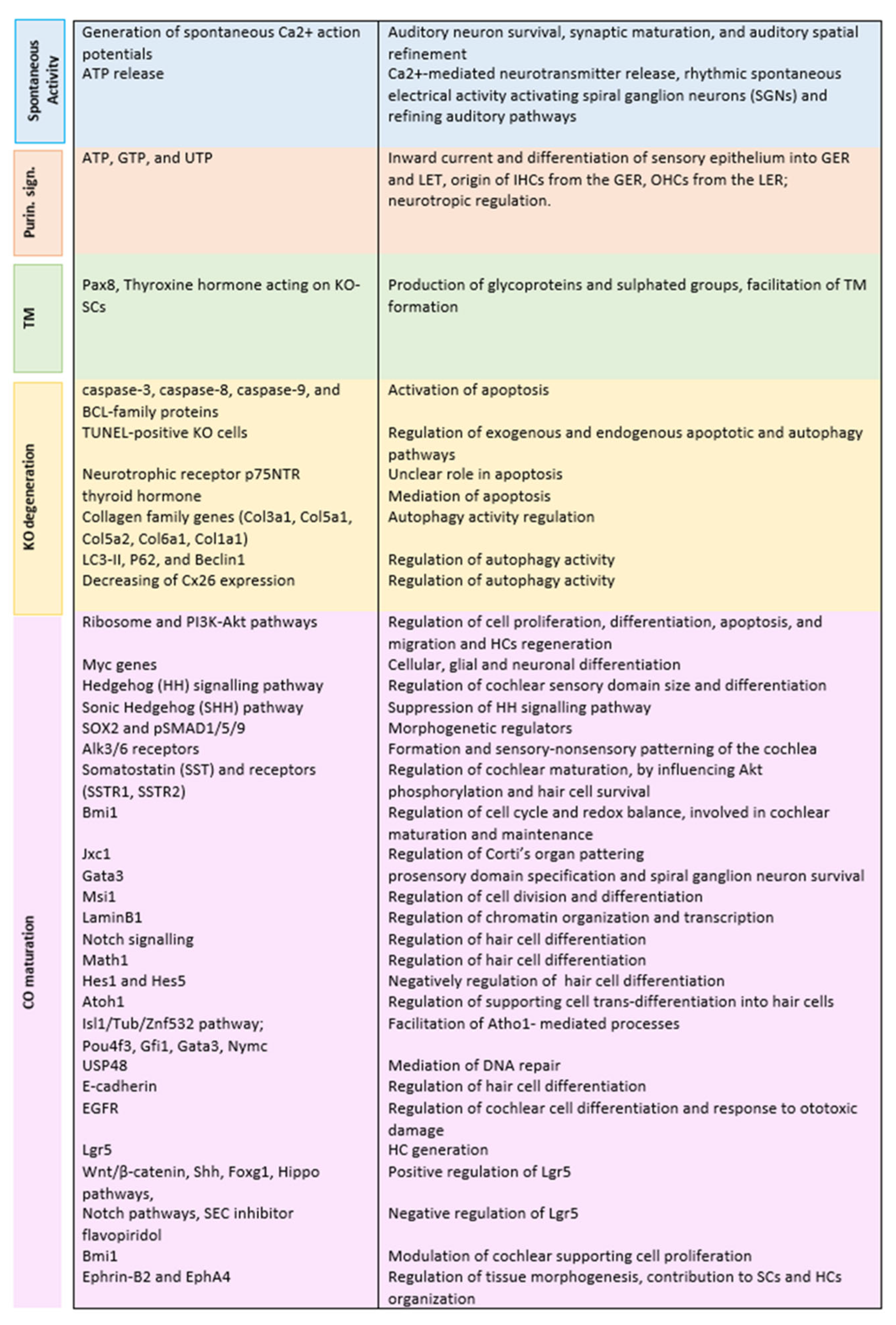
3. Results
3.1. Regulation of Spontaneous Cochlear Activity and Purinergic Signalling
3.2. Possible Mechanisms of Spontaneous Activity Generation and Mediators in Kölliker’s Organ
3.3. Role of KO in Tectorial Membrane Formation
3.4. Molecular Factors Influencing KO Degeneration
3.5. Autophagy in KO-Supporting Cells
3.6. KO and Cochlear Immune Response
3.7. Molecular Signalling Pathways in Kölliker’s Organ-Supporting Cell Degeneration and Cochlear Hearing Development
3.8. Trans-Differentiation Potential of Kölliker’s Organ-Supporting Cells into Hair Cells
3.8.1. Role of Atho1 and Hes Genes
3.8.2. Atoh1 and Transcription Factor Interplay
3.8.3. USP48 and Cochlear Development
3.8.4. EGFR and Cochlear Cell Differentiation
3.8.5. Lgr5-Positive Progenitor Cells and Wnt Signalling
3.8.6. Bmi1 and Hair Cell Survival
3.8.7. Ephrin-B2 and Cell Fate Regulation
3.8.8. Single-Cell Transcriptome Sequencing
4. Discussion
5. Future Directions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| KO | Kölliker’s organ |
| PLC | Phospholipase C |
| IP3 | Inositol triphosphate |
| SNG | Spiral ganglion neuron |
| GER | Greater epithelial ridge |
| LER | Lesser epithelial ridge |
| IHC | Inner hair cell |
| OHC | Outer hair cell |
| TM | Tectorial membrane |
References
- Geal-Dor, M.; Freeman, S.; Li, G.; Sohmer, H. Development of hearing in neonatal rats: Air and bone conducted ABR thresholds. Hear. Res. 1993, 69, 236–242. [Google Scholar] [CrossRef]
- Locher, H.; Frijns, J.H.; van Iperen, L.; de Groot, J.C.; A Huisman, M.; Lopes, S.M.C.d.S. Neurosensory development and cell fate determination in the human cochlea. Neural Dev. 2013, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Hudspeth, A.J. How hearing happens. Neuron 1997, 19, 947–950. [Google Scholar] [CrossRef] [PubMed]
- Martini, A.; Mazzarello, P.; Mira, E.; Mudry, A. Alfonso Corti: The Discovery of the Hearing Organ; Kugler Publications: Amsterdam, The Netherlands, 2024. [Google Scholar]
- Kolliker, A. Bau der Schnecke. In Handbuch der Gewebelehre des Menschen; Wilhelm Engelmann: Leipzig, Germany, 1863; Volume 4, p. 708. [Google Scholar]
- Caragli, V. Kolliker and Corti’s organs: Morphological aspects and development of the hearing organ. Hear. Balance Commun. 2025, 23. [Google Scholar]
- Tritsch, N.X.; Yi, E.; Gale, J.E.; Glowatzki, E.; Bergles, D.E. The origin of spontaneous activity in the developing auditory system. Nature 2007, 450, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Tritsch, N.X.; Bergles, D.E. Developmental regulation of spontaneous activity in the Mammalian cochlea. J. Neurosci. 2010, 30, 1539–1550. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, B.; Hu, B.; Yang, S. Cell junction proteins within the cochlea: A review of recent research. J. Otolaryngol. 2015, 10, 131–135. [Google Scholar] [CrossRef]
- Huang, L.C.; Thorne, P.R.; Vlajkovic, S.M.; Housley, G.D. Differential expression of P2Y receptors in the rat cochlea during development. Purinergic Signal. 2010, 6, 231–248. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ebeid, M.; Barnas, K.; Zhang, H.; Yaghmour, A.; Noreikaite, G.; Bjork, B.C. PRDM16 expression and function in mammalian cochlear development. Dev. Dyn. 2022, 251, 1666–1683. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lim, D.J. Functional structure of the organ of Corti: A review. Hear. Res. 1986, 22, 117–146. [Google Scholar] [CrossRef]
- Lelli, A.; Asai, Y.; Forge, A.; Holt, J.R.; Géléoc, G.S. Tonotopic gradient in the developmental acquisition of sensory transduction in outer hair cells of the mouse cochlea. J. Neurophysiol. 2009, 101, 2961–2973. [Google Scholar] [CrossRef]
- Blankenship, A.G.; Feller, M.B. Mechanisms underlying spontaneous patterned activity in developing neural circuits. Nat. Rev. Neurosci. 2010, 11, 18–29. [Google Scholar] [CrossRef]
- Wang, H.C.; Bergles, D.E. Spontaneous activity in the developing auditory system. Cell Tissue Res. 2015, 361, 65–75. [Google Scholar] [CrossRef]
- Beurg, M.; Safieddine, S.; Roux, I.; Bouleau, Y.; Petit, C.; Dulon, D. Calcium- and otoferlin-dependent exocytosis by immature outer hair cells. J. Neurosci. 2008, 28, 1798–1803. [Google Scholar] [CrossRef]
- Marcotti, W. Functional assembly of mammalian cochlear hair cells. Exp. Physiol. 2012, 97, 438–451. [Google Scholar] [CrossRef]
- Sendin, G.; Bourien, J.; Rassendren, F.; Puel, J.L.; Nouvian, R. Spatiotemporal pattern of action potential firing in developing inner hair cells of the mouse cochlea. Proc. Natl. Acad. Sci. USA 2014, 111, 1999–2004. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liang, Y.; Huang, L.; Yang, J. Differential expression of ryanodine receptor in the developing rat cochlea. Eur. J. Histochem. 2009, 53, e30. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Dayaratne, M.W.; Vlajkovic, S.M.; Lipski, J.; Thorne, P.R. Kölliker’s organ and the development of spontaneous activity in the auditory system: Implications for hearing dysfunction. Biomed. Res. Int. 2014, 2014, 367939. [Google Scholar] [CrossRef]
- Rodriguez, L.; Simeonato, E.; Scimemi, P.; Anselmi, F.; Calì, B.; Crispino, G.; Ciubotaru, C.D.; Bortolozzi, M.; Ramirez, F.G.; Majumder, P.; et al. Reduced phosphatidylinositol 4,5-bisphosphate synthesis impairs inner ear Ca2+ signaling and high-frequency hearing acquisition. Proc. Natl. Acad. Sci. USA 2012, 109, 14013–14018. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.L.; Ceriani, F.; Houston, O.; Polishchuk, R.; Polishchuk, E.; Crispino, G.; Zorzi, V.; Mammano, F.; Marcotti, W. Connexin-mediated signaling in nonsensory cells is crucial for the development of sensory inner hair cells in the mouse cochlea. J. Neurosci. 2017, 37, 258–268. [Google Scholar] [CrossRef]
- Zhao, H.B.; Yu, N.; Fleming, C.R. Gap junctional hemichannel-mediated ATP release and hearing controls in the inner ear. Proc. Natl. Acad. Sci. USA 2005, 102, 18724–18729. [Google Scholar] [CrossRef]
- Lim, D.J.; Anniko, M. Developmental morphology of the mouse inner ear. A scanning electron microscopic observation. Acta Otolaryngol. Suppl. 1985, 422, 1–69. [Google Scholar] [CrossRef] [PubMed]
- Simonneau, L.; Gallego, M.; Pujol, R. Comparative expression patterns of T-, N-, E-cadherins, beta-catenin, and polysialic acid neural cell adhesion molecule in rat cochlea during development: Implications for the nature of Kölliker’s organ. J. Comp. Neurol. 2003, 459, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Housley, G.D.; Bringmann, A.; Reichenbach, A. Purinergic signaling in special senses. Trends Neurosci. 2009, 32, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Abbracchio, M.P.; Burnstock, G.; Verkhratsky, A.; Zimmermann, H. Purinergic signalling in the nervous system: An overview. Trends Neurosci. 2009, 32, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.C.; Greenwood, D.; Thorne, P.R.; Housley, G.D. Developmental regulation of neuron-specific P2X3 receptor expression in the rat cochlea. J. Comp. Neurol. 2005, 484, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Pirvola, U.; Ylikoski, J. Neurotrophic factors during inner ear development. Curr. Top. Dev. Biol. 2003, 57, 207–223. [Google Scholar] [CrossRef] [PubMed]
- Mou, K.; Hunsberger, C.L.; Cleary, J.M.; Davis, R.L. Synergistic effects of BDNF and NT-3 on postnatal spiral ganglion neurons. J. Comp. Neurol. 1997, 386, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, D.; Jagger, D.J.; Huang, L.C.; Hoya, N.; Thorne, P.R.; Wildman, S.S.; King, B.F.; Pak, K.; Ryan, A.F.; Housley, G.D. P2X receptor signaling inhibits BDNF-mediated spiral ganglion neuron development in the neonatal rat cochlea. Development 2007, 134, 1407–1417. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, P.; Housley, G.D.; Thorne, P.R. Expression of the P2X7 receptor subunit of the adenosine 5’-triphosphate-gated ion channel in the developing and adult rat cochlea. Audiol. Neurotol. 2003, 8, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, P.; Housley, G.D.; Luo, L.; Ryan, A.F.; Thorne, P.R. Transient expression of P2X(1) receptor subunits of ATP-gated ion channels in the developing rat cochlea. Brain Res. Dev. Brain Res. 2001, 126, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.L.; Kuhn, S.; Franz, C.; Ingham, N.; Furness, D.N.; Knipper, M.; Steel, K.P.; Adelman, J.P.; Holley, M.C.; Marcotti, W. Presynaptic maturation in auditory hair cells requires a critical period of sensory-independent spiking activity. Proc. Natl. Acad. Sci. USA 2013, 110, 8720–8725. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Dale, N. Dynamic ATP signalling and neural development. J. Physiol. 2008, 586, 2429–2436. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Eckrich, T.; Blum, K.; Milenkovic, I.; Engel, J. Fast Ca2+ Transients of Inner Hair Cells Arise Coupled and Uncoupled to Ca2+ Waves of Inner Supporting Cells in the Developing Mouse Cochlea. Front. Mol. Neurosci. 2018, 11, 264. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zuo, J.; Curtis, L.M.; Yao, X.; Ten Cate, W.J.; Rarey, K.E. Expression of Na, K-ATPase alpha and beta isoforms in the neonatal rat cochlea. Acta Otolaryngol. 1995, 115, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Forge, A.; Becker, D.; Casalotti, S.; Edwards, J.; Marziano, N.; Nevill, G. Gap junctions in the inner ear: Comparison of distribution patterns in different vertebrates and assessement of connexin composition in mammals. J. Comp. Neurol. 2003, 467, 207–231. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.B. Connexin26 is responsible for anionic molecule permeability in the cochlea for intercellular signalling and metabolic communications. Eur. J. Neurosci. 2005, 21, 1859–1868. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Majumder, P.; Crispino, G.; Rodriguez, L.; Ciubotaru, C.D.; Anselmi, F.; Piazza, V.; Bortolozzi, M.; Mammano, F. ATP-mediated cell-cell signaling in the organ of Corti: The role of connexin channels. Purinergic Signal. 2010, 6, 167–187. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Anselmi, F.; Hernandez, V.H.; Crispino, G.; Seydel, A.; Ortolano, S.; Roper, S.D.; Kessaris, N.; Richardson, W.; Rickheit, G.; Filippov, M.A.; et al. ATP release through connexin hemichannels and gap junction transfer of second messengers propagate Ca2+ signals across the inner ear. Proc. Natl. Acad. Sci. USA 2008, 48, 18770–18775. [Google Scholar] [CrossRef]
- Wang, X.H.; Streeter, M.; Liu, Y.P.; Zhao, H.B. Identification and characterization of pannexin expression in the mammalian cochlea. J. Comp. Neurol. 2009, 512, 336–346, Erratum in J. Comp. Neurol. 2010, 518, 1156. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tamagawa, Y.; Kitamura, K.; Ishida, T.; Ishikawa, K.; Tanaka, H.; Tsuji, S.; Nishizawa, M. A gene for a dominant form of non-syndromic sensorineural deafness (DFNA11) maps within the region containing the DFNB2 recessive deafness gene. Hum. Mol. Genet. 1996, 5, 849–852. [Google Scholar] [CrossRef] [PubMed]
- Lippe, W.R. Rhythmic spontaneous activity in the developing avian auditory system. J. Neurosci. 1994, 14 Pt 2, 1486–1495. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gale, J.E.; Piazza, V.; Ciubotaru, C.D.; Mammano, F. A mechanism for sensing noise damage in the inner ear. Curr. Biol. 2004, 14, 526–529. [Google Scholar] [CrossRef] [PubMed]
- Vlajkovic, S.M.; Thorne, P.R.; Housley, G.D.; Muñoz, D.J.; Kendrick, I.S. Ecto-nucleotidases terminate purinergic signalling in the cochlear endolymphatic compartment. Neuroreport 1998, 9, 1559–1565. [Google Scholar] [PubMed]
- Zhu, Y.; Zhao, H.B. ATP activates P2X receptors to mediate gap junctional coupling in the cochlea. Biochem. Biophys. Res. Commun. 2012, 426, 528–532. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sobkowicz, H.M.; Loftus, J.M.; Slapnick, S.M. Tissue culture of the organ of Corti. Acta Otolaryngol. Suppl. 1993, 502, 3–36. [Google Scholar] [PubMed]
- Coppens, A.G.; Résibois, A.; Poncelet, L. Immunolocalization of calbindin D28k and calretinin in the dog cochlea during postnatal development. Hear. Res. 2000, 145, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Legrand, C.; Bréhier, A.; Clavel, M.C.; Thomasset, M.; Rabié, A. Cholecalcin (28-kDa CaBP) in the rat cochlea. Development in normal and hypothyroid animals. An immunocytochemical study. Brain Res. 1988, 466, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Buckiová, D.; Syka, J. Calbindin and S100 protein expression in the developing inner ear in mice. J. Comp. Neurol. 2009, 513, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Tauris, J.; Christensen, E.I.; Nykjaer, A.; Jacobsen, C.; Petersen, C.M.; Ovesen, T. Cubilin and megalin co-localize in the neonatal inner ear. Audiol. Neurotol. 2009, 14, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Dechesne, C.J.; Lavigne-Rebillard, M.; Brehier, A.; Thomasset, M.; Sans, A. Appearance and distribution of neuron-specific enolase and calbindin (CaBP 28 kDa) in the developing human inner ear. Brain Res. 1988, 469, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Dechesne, C.J.; Thomasset, M. Calbindin (CaBP 28 kDa) appearance and distribution during development of the mouse inner ear. Brain Res. 1988, 468, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Johnson, K.R.; Lett, J.M.; Voss, R.; Salt, A.N.; Hartsock, J.J.; Steyger, P.S.; Ohlemiller, K.K. CACHD1-deficient mice exhibit hearing and balance deficits associated with a disruption of calcium homeostasis in the inner ear. Hear. Res. 2021, 409, 108327. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liu, W.; Glueckert, R.; Schrott-Fischer, A.; Rask-Andersen, H. Human cochlear microanatomy—An electron microscopy and super-resolution structured illumination study and review. Hear. Balance Commun. 2020, 18, 256–269. [Google Scholar] [CrossRef]
- Uziel, A.; Gabrion, J.; Ohresser, M.; Legrand, C. Effects of hypothyroidism on the structural development of the organ of Corti in the rat. Acta Otolaryngol. 1981, 92, 469–480. [Google Scholar] [CrossRef]
- Remezal, M.; Gil-Loyzaga, P. Incorporation of D3H glucosamine to the adult and developing cochlear tectorial membrane of normal and hypothyroid rats. Hear. Res. 1993, 66, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Munyer, P.D.; Schulte, B.A. Developmental expression of proteoglycans in the tectorial and basilar membrane of the gerbil cochlea. Hear. Res. 1995, 85, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Richardson, G.P.; Lukashkin, A.N.; Russell, I.J. The tectorial membrane: One slice of a complex cochlear sandwich. Curr. Opin. Otolaryngol. Head Neck Surg. 2008, 16, 458–464. [Google Scholar] [CrossRef]
- Khetarpal, U.; Morton, C.C. Inner ridge cells may be the main source of tectorial membrane type II collagen: Evidence from quantitative mRNA in situ hybridization. Acta Otolaryngol. 1998, 118, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Tannenbaum, J.; Slepecky, N.B. Localization of microtubules containing posttranslationally modified tubulin in cochlear epithelial cells during development. Cell Motil. Cytoskelet. 1997, 38, 146–162. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Wang, X.; Li, Y.; Song, Q.; Yang, J. Impaired tectorial membrane and ribbon synapse maturation in the cochlea of mice with congenital hypothyroidism. Biochem. Biophys. Res. Commun. 2023, 655, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Christ, S.; Biebel, U.W.; Hoidis, S.; Friedrichsen, S.; Bauer, K.; Smolders, J.W. Hearing loss in athyroid pax8 knockout mice and effects of thyroxine substitution. Audiol. Neurootol. 2004, 9, 88–106. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Chen, J.; Yang, J. Autophagy precedes apoptosis during degeneration of the Kölliker’s organ in the development of rat cochlea. Eur. J. Histochem. 2019, 63, 3025. [Google Scholar] [CrossRef]
- Hou, S.; Chen, P.; Chen, J.; Chen, J.; Sun, L.; Chen, J.; He, B.; Li, Y.; Qin, H.; Hong, Y.; et al. Distinct expression patterns of apoptosis and autophagy-associated proteins and genes during postnatal development of spiral ganglion neurons in rat. Neural Plast. 2020, 2020, 9387560. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cai, L.; He, Y.; Yang, J. Apoptosis pattern and alterations of expression of apoptosis-related factors of supporting cells in Kölliker’s organ in vivo in early stage after birth in rats. Eur. J. Histochem. 2017, 61, 2706. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, K.; Takahashi, K.; Kitamura, K.; Momoi, T.; Yoshikawa, Y. Mitosis and apoptosis in postnatal auditory system of the C3H/He strain. Brain Res. 2001, 901, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Rüsch, A.; Ng, L.; Goodyear, R.; Oliver, D.; Lisoukov, I.; Vennström, B.; Richardson, G.; Kelley, M.W.; Forrest, D. Retardation of cochlear maturation and impaired hair cell function caused by deletion of all known thyroid hormone receptors. J. Neurosci. 2001, 21, 9792–9800. [Google Scholar] [CrossRef]
- Knipper, M.; Gestwa, L.; Ten Cate, W.J.; Lautermann, J.; Brugger, H.; Maier, H.; Zimmermann, U.; Rohbock, K.; Köpschall, I.; Wiechers, B.; et al. Distinct thyroid hormone-dependent expression of TrKB and p75NGFR in nonneuronal cells during the critical TH-dependent period of the cochlea. J. Neurobiol. 1999, 38, 338–356. [Google Scholar] [CrossRef]
- Sato, T.; Doi, K.; Taniguchi, M.; Yamashita, T.; Kubo, T.; Tohyama, M. Progressive hearing loss in mice carrying a mutation in the p75 gene. Brain Res. 2006, 1091, 224–234. [Google Scholar] [CrossRef]
- Coen, L.; Le Blay, K.; Rowe, I.; Demeneix, B.A. Caspase9 regulates apoptosis/proliferation balance during metamorphic brain remodeling in Xenopus. Proc. Natl. Acad. Sci. USA 2007, 104, 8502–8507. [Google Scholar] [CrossRef]
- Qu, X.; Zou, Z.; Sun, Q.; Luby-Phelps, K.; Cheng, P.; Hogan, R.N.; Gilpin, C.; Levine, B. Autophagy gene-dependent clearance of apoptotic cells during embryonic development. Cell 2007, 128, 931–946. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Gao, D.; Chen, J.; Hou, S.; Li, Y.; Huang, Y.; Mammano, F.; Chen, J.; Yang, J. Failure of Hearing Acquisition in Mice with Reduced Expression of Connexin 26 Correlates with the Abnormal Phasing of Apoptosis Relative to Autophagy and Defective ATP-Dependent Ca2+ Signaling in Kölliker’s Organ. Front. Cell. Neurosci. 2022, 16, 816079. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Borse, V.; Kaur, T.; Hinton, A.; Ohlemiller, K.; Warchol, M.E. Programmed Cell Death Recruits Macrophages into the Developing Mouse Cochlea. Front. Cell Dev. Biol. 2021, 9, 777836. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hayashi, Y.; Suzuki, H.; Nakajima, W.; Uehara, I.; Tanimura, A.; Himeda, T.; Koike, S.; Katsuno, T.; Kitajiri, S.I.; Koyanagi, N.; et al. Cochlear supporting cells function as macrophage-like cells and protect audiosensory receptor hair cells from pathogens. Sci. Rep. 2020, 10, 6740. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cheng, C.; Wang, Y.; Guo, L.; Lu, X.; Zhu, W.; Muhammad, W.; Zhang, L.; Lu, L.; Gao, J.; Tang, M.; et al. Age-related transcriptome changes in Sox2+ supporting cells in the mouse cochlea. Stem Cell Res. Ther. 2019, 10, 365. [Google Scholar] [CrossRef]
- Chen, Y.; Gu, Y.; Li, Y.; Li, G.-L.; Chai, R.; Li, W.; Li, H. Generation of mature and functional hair cells by co-expression of Gfi1, Pou4f3 and Atoh1 in the postnatal mouse cochlea. Cell Rep. 2021, 35, 109016. [Google Scholar] [CrossRef]
- Tang, D.; Lin, Q.; He, Y.; Chai, R.; Li, H. Inhibition of H3K9me2 reduces hair cell regeneration after hair cell loss in the zebrafish lateral line by down-regulating the Wnt and Fgf signaling pathways. Front. Mol. Neurosci. 2016, 9, 39. [Google Scholar] [CrossRef]
- Ni, W.; Zeng, S.; Li, W.; Chen, Y.; Zhang, S.; Tang, M.; Sun, S.; Chai, R.; Li, H. Wnt activation followed by Notch inhibition promotes mitotic hair cell regeneration in the postnatal mouse cochlea. Oncotarget 2016, 7, 66754–66768. [Google Scholar] [CrossRef]
- Waqas, M.; Zhang, S.; He, Z.; Tang, M.; Chai, R. Role of Wnt and Notch signaling in regulating hair cell regeneration in the cochlea. Front. Med. 2016, 10, 237–249. [Google Scholar] [CrossRef]
- He, Z.-H.; Zou, S.-Y.; Li, M.; Liao, F.-L.; Wu, X.; Sun, H.-Y.; Zhao, X.-Y.; Hu, Y.-J.; Li, D.; Xu, X.-X.; et al. The nuclear transcription factor FoxG1 affects the sensitivity of mimetic aging hair cells to inflammation by regulating autophagy pathways. Redox. Biol. 2020, 28, 101364. [Google Scholar] [CrossRef]
- He, Z.-H.; Li, M.; Fang, Q.-J.; Liao, F.-L.; Zou, S.-Y.; Wu, X.; Sun, H.-Y.; Zhao, X.-Y.; Hu, Y.-J.; Xu, X.-X.; et al. FOXG1 promotes aging inner ear hair cell survival through activation of the autophagy pathway. Autophagy 2021, 17, 4341–4362. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Dong, Y.; Qiang, R.; Zhang, Y.; Zhang, X.; Chen, Y.; Jiang, P.; Ma, X.; Wu, L.; Ai, J.; et al. Characterization of Strip1 expression in mouse cochlear hair cells. Front. Genet. 2021, 12, 625867. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Sun, X.; Zhang, L.; Jin, Y.; Chai, R.; Yang, L.; Zhang, A.; Liu, X.; Bai, X.; Li, J.; et al. Tuberous sclerosis complex-mediated mTORC1 overactivation promotes age-related hearing loss. J. Clin. Investig. 2018, 128, 4938–4955. [Google Scholar] [CrossRef]
- Fu, X.; Li, P.; Zhang, L.; Song, Y.; An, Y.; Zhang, A.; Liu, W.; Ye, C.; Zhang, Y.; Yue, R.; et al. Activation of Rictor/mTORC2 signaling acts as a pivotal strategy to protect against sensorineural hearing loss. Proc. Natl. Acad. Sci. USA 2022, 119, e2107357119. [Google Scholar] [CrossRef] [PubMed]
- Chai, R.; Kuo, B.; Wang, T.; Liaw, E.J.; Xia, A.; Jan, T.A.; Liu, Z.; Taketo, M.M.; Oghalai, J.S.; Nusse, R.; et al. Wnt signaling induces proliferation of sensory precursors in the postnatal mouse cochlea. Proc. Natl. Acad. Sci. USA 2012, 109, 8167–8172. [Google Scholar] [CrossRef]
- Liu, L.; Chen, Y.; Qi, J.; Zhang, Y.; He, Y.; Ni, W.; Li, W.; Zhang, S.; Sun, S.; Taketo, M.M.; et al. Wnt activation protects against neomycin-induced hair cell damage in the mouse cochlea. Cell Death Dis. 2016, 7, e2136. [Google Scholar] [CrossRef]
- Pelletier, J.; Thomas, G.; Volarevic, S. Ribosome biogenesis in cancer: New players and therapeutic avenues. Nat. Rev. Cancer 2018, 18, 51–63. [Google Scholar] [CrossRef]
- Jia, X.; Wen, Z.; Sun, Q.; Zhao, X.; Yang, H.; Shi, X.; Xin, T. Apatinib suppresses the proliferation and apoptosis of gastric cancer cells via the PI3K/Akt signaling pathway. J. BUON 2019, 24, 1985–1991. [Google Scholar]
- Bu, C.; Xu, L.; Han, Y.; Wang, M.; Wang, X.; Liu, W.; Chai, R.; Wang, H. c-Myb protects cochlear hair cells from cisplatin-induced damage via the PI3K/Akt signaling pathway. Cell Death Discov. 2022, 8, 78. [Google Scholar] [CrossRef]
- Mullen, L.M.; Pak, K.K.; Chavez, E.; Kondo, K.; Brand, Y.; Ryan, A.F. Ras/p38 and PI3K/Akt but not Mek/Erk signaling mediate BDNF-induced neurite formation on neonatal cochlear spiral ganglion explants. Brain Res. 2012, 1430, 25–34. [Google Scholar] [CrossRef]
- Xia, L.; Zhu, W.; Wang, Y.; He, S.; Chai, R. Regulation of neural stem cell proliferation and differentiation by graphene-based biomaterials. Neural Plast. 2019, 2019, 3608386. [Google Scholar] [CrossRef]
- Romand, R.; Hirning-Folz, U.; Ehret, G. N-myc expression in the embryonic cochlea of the mouse. Hear. Res. 1994, 72, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Frutos, E.; López-Hernández, I.; Vendrell, V.; Neves, J.; Gallozzi, M.; Gutsche, K.; Quintana, L.; Sharpe, J.; Knoepfler, P.S.; Eisenman, R.N.; et al. N-myc controls proliferation, morphogenesis, and patterning of the inner ear. J. Neurosci. 2011, 31, 7178–7189. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Driver, E.C.; Pryor, S.P.; Hill, P.; Turner, J.; Rüther, U.; Biesecker, L.G.; Griffith, A.J.; Kelley, M.W. Hedgehog signaling regulates sensory cell formation and auditory function in mice and humans. J. Neurosci. 2008, 28, 7350–7358. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.J.; Munnamalai, V.; Umulis, D.M. Early precision of radial patterning of the mouse cochlea is achieved by a linear BMP signaling gradient and is further refined by SOX2. bioRxiv 2022. [Google Scholar] [CrossRef]
- Ohyama, T.; Basch, M.L.; Mishina, Y.; Lyons, K.M.; Segil, N.; Groves, A.K. BMP signaling is necessary for patterning the sensory and nonsensory regions of the developing mammalian cochlea. J. Neurosci. 2010, 30, 15044–15051. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bodmer, D.; Brand, Y.; Radojevic, V. Somatostatin receptor types 1 and 2 in the developing mammalian cochlea. Dev. Neurosci. 2012, 34, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, Y.; Li, W.; Ni, W.; Chai, R.; Chen, Y. Spatiotemporal expression of Bmi1 in the developing mouse cochlea. Int. J. Clin. Exp. Med. 2017, 10, 556–562. [Google Scholar]
- Chen, Z.; Montcouquiol, M.; Calderon, R.; Jenkins, N.A.; Copeland, N.G.; Kelley, M.W.; Noben-Trauth, K. Jxc1/Sobp, encoding a nuclear zinc finger protein, is critical for cochlear growth, cell fate, and patterning of the organ of corti. J. Neurosci. 2008, 28, 6633–6641. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Luo, X.J.; Deng, M.; Xie, X.; Huang, L.; Wang, H.; Jiang, L.; Liang, G.; Hu, F.; Tieu, R.; Chen, R.; et al. GATA3 controls the specification of prosensory domain and neuronal survival in the mouse cochlea. Hum. Mol. Genet. 2013, 22, 3609–3623. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sakaguchi, H.; Yaoi, T.; Suzuki, T.; Okano, H.; Hisa, Y.; Fushiki, S. Spatiotemporal patterns of Musashi1 expression during inner ear development. Neuroreport 2004, 15, 997–1001. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Chen, J.; Chu, H. Differential Expression of LaminB1 in the Developing Rat Cochlea. J. Int. Adv. Otol. 2019, 15, 106–111. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zine, A.; Van De Water, T.R.; de Ribaupierre, F. Notch signaling regulates the pattern of auditory hair cell differentiation in mammals. Development 2000, 127, 3373–3383. [Google Scholar] [CrossRef] [PubMed]
- Chai, R.; Xia, A.; Wang, T.; Jan, T.A.; Hayashi, T.; Bermingham-McDonogh, O.; Cheng, A.G.-L. Dynamic expression of Lgr5, a Wnt target gene, in the developing and mature mouse cochlea. J. Assoc. Res. Otolaryngol. 2011, 12, 455–469. [Google Scholar] [CrossRef]
- Bramhall, N.F.; Shi, F.; Arnold, K.; Hochedlinger, K.; Edge, A.S. Lgr5-positive supporting cells generate new hair cells in the postnatal cochlea. Stem Cell Rep. 2014, 2, 311–322. [Google Scholar] [CrossRef]
- Zheng, J.L.; Gao, W.Q. Overexpression of Math1 induces robust production of extra hair cells in postnatal rat inner ears. Nat. Neurosci. 2000, 3, 580–586. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhai, S.Q.; Shou, J.; Song, W.; Sun, J.H.; Guo, W.; Zheng, G.L.; Hu, Y.Y.; Gao, W.Q. Isolation, growth and differentiation of hair cell progenitors from the newborn rat cochlear greater epithelial ridge. J. Neurosci. Methods 2007, 164, 271–279. [Google Scholar] [CrossRef]
- Bermingham, N.A.; Hassan, B.A.; Price, S.D.; Vollrath, M.A.; Ben-Arie, N.; Eatock, R.A.; Bellen, H.J.; Lysakowski, A.; Zoghbi, H.Y. Math1: An essential gene for the generation of inner ear hair cells. Science 1999, 284, 1837–1841. [Google Scholar] [CrossRef]
- Zine, A.; Aubert, A.; Qiu, J.; Therianos, S.; Guillemot, F.; Kageyama, R.; de Ribaupierre, F. Hes1 and Hes5 activities are required for the normal development of the hair cells in the mammalian inner ear. J. Neurosci. 2001, 21, 4712–4720. [Google Scholar] [CrossRef]
- McGovern, M.M.; Zhou, L.; Randle, M.R.; Cox, B.C. Spontaneous hair cell regeneration is prevented by increased notch signaling in supporting cells. Front. Cell. Neurosci. 2018, 12, 120. [Google Scholar] [CrossRef]
- Liu, Z.; Dearman, J.A.; Cox, B.C.; Walters, B.J.; Zhang, L.; Ayrault, O.; Zindy, F.; Gan, L.; Roussel, M.F.; Zuo, J. Age-dependent in vivo conversion of mouse cochlear pillar and Deiters’ cells to immature hair cells by Atoh1 ectopic expression. J. Neurosci. 2012, 32, 6600–6610. [Google Scholar] [CrossRef]
- Richardson, R.T.; Atkinson, P.J. Atoh1 gene therapy in the cochlea for hair cell regeneration. Expert Opin. Biol. Ther. 2015, 15, 417–430. [Google Scholar] [CrossRef]
- Walters, B.J.; Coak, E.; Dearman, J.; Bailey, G.; Yamashita, T.; Kuo, B.; Zuo, J. In vivo interplay between p27Kip1, GATA3, ATOH1 and POU4F3 converts non-sensory cells to hair cells in adult mice. Cell Rep. 2017, 19, 307–320. [Google Scholar] [CrossRef]
- Collado, M.S.; Thiede, B.R.; Baker, W.; Askew, C.; Igbani, L.M.; Corwin, J.T. The postnatal accumulation of junctional E-cadherin is inversely correlated with the capacity for supporting cells to convert directly into sensory hair cells in mammalian balance organs. J. Neurosci. 2011, 31, 11855–11866. [Google Scholar] [CrossRef]
- Bassani, S.; van Beelen, E.; Rossel, M.; Voisin, N.; Morgan, A.; Arribat, Y.; Chatron, N.; Chrast, J.; Cocca, M.; Delprat, B.; et al. Variants in USP48 encoding ubiquitin hydrolase are associated with autosomal dominant non-syndromic hereditary hearing loss. Hum. Mol. Genet. 2021, 30, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Zine, A.; de Ribaupierre, F. Tissue-specific levels and cellular distribution of epidermal growth factor receptors within control and neomycin-damaged neonatal rat Organ of Corti. J. Neurobiol. 1999, 38, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Zine, A.; Nyffeler, M.; de Ribaupierre, F. Spatial expression patterns of epidermal growth factor receptor gene transcripts in the postnatal mammalian cochlea. Hear. Res. 2000, 141, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Korrapati, S.; Roux, I.; Glowatzki, E.; Doetzlhofer, A. Notch signaling limits supporting cell plasticity in the hair cell-damaged early postnatal murine cochlea. PLoS ONE 2013, 8, e73276. [Google Scholar] [CrossRef]
- Mizutari, K.; Fujioka, M.; Hosoya, M.; Bramhall, N.; Okano, H.J.; Okano, H.; Edge, A.S. Notch inhibition induces cochlear hair cell regeneration and recovery of hearing after acoustic trauma. Neuron 2013, 77, 58–69. [Google Scholar] [CrossRef]
- Gregorieff, A.; Liu, Y.; Inanlou, M.R.; Khomchuk, Y.; Wrana, J.L. Yap-dependent reprogramming of Lgr5+ stem cells drives intestinal regeneration and cancer. Nature 2015, 526, 715–718. [Google Scholar] [CrossRef]
- Chen, Y.; Lu, X.; Guo, L.; Ni, W.; Zhang, Y.; Zhao, L.; Wu, L.; Sun, S.; Zhang, S.; Tang, M.; et al. Hedgehog signaling promotes the proliferation and subsequent hair cell formation of progenitor cells in the neonatal mouse cochlea. Front. Mol. Neurosci. 2017, 10, 426. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Guo, L.; Lu, L.; Xu, X.; Zhang, S.; Gao, J.; Waqas, M.; Zhu, C.; Chen, Y.; Zhang, X.; et al. Characterization of the transcriptomes of Lgr5+ hair cell progenitors and Lgr5- supporting cells in the mouse cochlea. Front. Mol. Neurosci. 2017, 10, 122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, Y.; Dong, Y.; Guo, L.; Zhang, Z.; Shao, B.; Qi, J.; Zhou, H.; Zhu, W.; Yan, X.; et al. Knockdown of Foxg1 in supporting cells increases the trans-differentiation of supporting cells into hair cells in the neonatal mouse cochlea. Cell. Mol. Life Sci. 2020, 77, 1401–1419. [Google Scholar] [CrossRef]
- Wu, J.; Li, W.; Lin, C.; Chen, Y.; Cheng, C.; Sun, S.; Tang, M.; Chai, R.; Li, H. Co-regulation of the Notch and Wnt signaling pathways promotes supporting cell proliferation and hair cell regeneration in mouse utricles. Sci. Rep. 2016, 6, 29418. [Google Scholar] [CrossRef]
- Lu, X.; Sun, S.; Qi, J.; Li, W.; Liu, L.; Zhang, Y.; Chen, Y.; Zhang, S.; Wang, L.; Miao, D.; et al. Bmi1 regulates the proliferation of cochlear supporting cells via the canonical Wnt signaling pathway. Mol. Neurobiol. 2017, 54, 1326–1339. [Google Scholar] [CrossRef]
- Fang, Q.; Zhang, Y.; Chen, X.; Li, H.; Cheng, L.; Zhu, W.; Zhang, Z.; Tang, M.; Liu, W.; Wang, H.; et al. Threedimensional graphene enhances neural stem cell proliferation through metabolic regulation. Front. Bioeng. Biotechnol. 2019, 7, 436. [Google Scholar] [CrossRef]
- Chen, Y.; Li, L.; Ni, W.; Zhang, Y.; Sun, S.; Miao, D.; Chai, R.; Li, H. Bmi1 regulates auditory hair cell survival by maintaining redox balance. Cell Death Dis. 2015, 6, e1605. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, L.M.; Liu, H. Comparison of ephrin-A ligand EphA receptor distribution in the developing inner ear. Anat. Rec. 1999, 254, 127–134. [Google Scholar] [CrossRef]
- Miko, I.J.; Henkemeyer, M.; Cramer, K.S. Auditory brainstem responses are impaired in EphA4 and ephrin-B2 deficient mice. Hear. Res. 2008, 235, 39–46. [Google Scholar] [CrossRef]
- Defourny, J.; Lallemend, F.; Malgrange, B. Structure and development of cochlear afferent innervation in mammals. Am. J. Physiol. Cell Physiol. 2011, 301, C750–C761. [Google Scholar] [CrossRef]
- Millimaki, B.B.; Sweet, E.M.; Riley, B.B. Sox2 is required for maintenance and regeneration, but not initial development, of hair cells in the zebrafish inner ear. Dev. Biol. 2010, 338, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.C.; Amoyel, M.; Qiu, X.; Jiang, Y.J.; Xu, Q.; Wilkinson, D.G. Notch activation regulates the segregation and differentiation of rhombomere boundary cells in the zebrafish hindbrain. Dev. Cell 2004, 6, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Tossell, K.; Kiecker, C.; Wizenmann, A.; Lang, E.; Irving, C. Notch signalling stabilises boundary formation at the midbrain-hindbrain organiser. Development 2011, 138, 3745–3757. [Google Scholar] [CrossRef] [PubMed]
- Kolla, L.; Kelly, M.C.; Mann, Z.F.; Anaya-Rocha, A.; Ellis, K.; Lemons, A.; Palermo, A.T.; So, K.S.; Mays, J.C.; Orvis, J.; et al. Characterization of the development of the mouse cochlear epithelium at the single cell level. Nat. Commun. 2020, 11, 2389. [Google Scholar] [CrossRef]
- Chen, J.; Gao, D.; Chen, J.; Hou, S.; He, B.; Li, Y.; Li, S.; Zhang, F.; Sun, X.; Mammano, F.; et al. SingleCell RNA sequencing analysis reveals greater epithelial ridge cells degeneration during postnatal development of cochlea in rats. Front. Cell. Dev. Biol. 2021, 9, 719491. [Google Scholar] [CrossRef]
- Zine, A.; Romand, R. Development of the auditory receptors of the rat: A SEM study. Brain Res. 1996, 721, 49–58. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Criteria | |
|---|---|
| Inclusion | -Studies analysing Kölliker’s organ -Human or mammalian subjects |
| Exclusion | -Non-English papers |
| Database | Query |
|---|---|
| Pubmed | “Kolliker’s organ” [All Fields] OR “Kolliker organ” [All Fields] |
| Scopus | TITLE-ABS-KEY(“Kolliker’s organ” OR “Kolliker organ”) |
| Embase | “Kolliker’s organ” OR “Kolliker organ” |
| CINAHL | “Kolliker’s organ” OR “Kolliker organ” |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caragli, V.; Di Pasquale Fiasca, V.M.; Genovese, E.; Martini, A. Kolliker’s Organ and Its Functional Role in the Development of Corti’s Organ and Auditory Systems. Audiol. Res. 2025, 15, 75. https://doi.org/10.3390/audiolres15040075
Caragli V, Di Pasquale Fiasca VM, Genovese E, Martini A. Kolliker’s Organ and Its Functional Role in the Development of Corti’s Organ and Auditory Systems. Audiology Research. 2025; 15(4):75. https://doi.org/10.3390/audiolres15040075
Chicago/Turabian StyleCaragli, Valeria, Valerio M. Di Pasquale Fiasca, Elisabetta Genovese, and Alessandro Martini. 2025. "Kolliker’s Organ and Its Functional Role in the Development of Corti’s Organ and Auditory Systems" Audiology Research 15, no. 4: 75. https://doi.org/10.3390/audiolres15040075
APA StyleCaragli, V., Di Pasquale Fiasca, V. M., Genovese, E., & Martini, A. (2025). Kolliker’s Organ and Its Functional Role in the Development of Corti’s Organ and Auditory Systems. Audiology Research, 15(4), 75. https://doi.org/10.3390/audiolres15040075