Abstract
Biofilms are a spontaneously formed slimy matrix of extracellular polymeric substances (EPS) enveloping miniature bacterial colonies, which aid in pathogen colonization, shielding the bacteria from antibiotics, as well as imparting them resistance towards the same. Biofilms employ a robust communication mechanism called quorum sensing that serves to keep their population density constant. What is most significant about biofilms is that they contribute to the development of bacterial virulence by providing protection to pathogenic species, allowing them to colonize the host, and also inhibiting the activities of antimicrobials on them. They grow on animate surfaces (such as on teeth and intestinal mucosa, etc.) and inanimate objects (like catheters, contact lenses, pacemakers, endotracheal devices, intrauterine devices, and stents, etc.) alike. It has been reported that as much as 80% of human infections involve biofilms. Serious implications of biofilms include the necessity of greater concentrations of antibiotics to treat common human infections, even contributing to antimicrobial resistance (AMR), since bacteria embedded within biofilms are protected from the action of potential antibiotics. This review explores various contemporary strategies for controlling biofilms, focusing on their modes of action, mechanisms of drug resistance, and innovative approaches to find a solution in this regard. This review interestingly targets the extracellular polymeric matrix as a highly effective strategy to counteract the potential harm of biofilms since it plays a critical role in biofilm formation and significantly contributes to antimicrobial resistance.
1. Introduction
Marshall and co-workers were the first to propose the concept and formation of biofilms [1]. According to Fletcher and Casterton, “Biofilm is a crucial phenomenon of development in the life span of microbes that gives free-living bacterial cells special features, benefits, and a higher level of organization during colonization” [2]. According to Flemming and Wuertz, biofilms are collections of microorganisms with distinct sessile cells that multiply to form tiny clusters, microcolonies, and subsequently larger quantities [3]. A direct communication between the film underneath the biofilm and the solid support occurs in a multi-layered heterogeneous microbial mat. These biofilms play very useful roles in certain biotechnological applications, such as drinking water screening, wastewater degradation, and biofuel generation [4]. However, biofilms are known to have a detrimental impact on the quality of drinking water as they cause biofouling, corrosion, and degradation. Microscopic organisms that create biofilms among higher eukaryotes, including humans, have been reported [5]. The most common examples of biofilms formed in eukaryotic ecosystems include those associated with human dental plaque, skin, and gut. The growing use of medical equipment has created luxurious new habitats for bacteria to produce biofilms.
Typically, microbial biofilms become coated with some high molecular weight extracellular polymeric substances (EPS), which empower the biofilm bacteria to endure harsh growth conditions [6]. These EPS provide structural strength, help the biofilm adhere to surfaces, capture nutrients, and provide protection from antimicrobial treatments as much as from host immune responses. Proteins, lipids, polysaccharides, and extracellular DNA (eDNA) comprise the EPS, which play a crucial role in the pathogenesis of various microbial diseases [7]. A schematic overview of the structure of a biofilm is shown in Figure 1. Additionally, it has been revealed that microbial cells in biofilms are resistant to an array of conditions, including UV light, metal toxins, acid exposure, desiccation, and pH variations [8]. In addition to these physical and chemical tolerances, EPS also provide immunological resistance to a number of harmful bacteria residing inside biofilms by preventing their phagocytosis by polymorphonuclear cells such as neutrophils. Furthermore, antimicrobials administered to the patient to handle these pathogens are not able to access them well due to the EPS layer covering them [7]. The extracellular DNA chelates human antimicrobial peptides (AMPs) in the EPS, which further reduces their antibacterial action [9]. Many studies have been conducted to determine how biofilms are produced and subsequent preventive solutions to address this issue, particularly resistance to antibiotics imparted by them [10]. Additionally, the EPS permit shuffling of genomes by horizontal gene transfer (HGT), facilitate cell-to-cell communication by quorum sensing, and maintain the community of biofilm cells snugly close together [11].
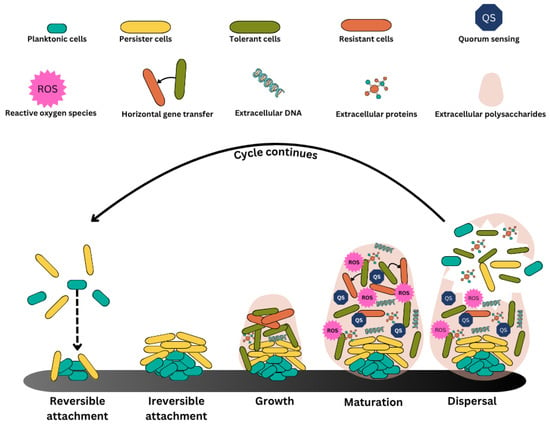
Figure 1.
Phases of biofilm development and overview of mature biofilm structure. Biofilm formation is the process by which planktonic cells develop into a complex, multicellular population. At the beginning of this process, planktonic cells bind to a substrate reversibly, which leads to irreversible attachment. As the biofilm grows, cells coordinate using quorum sensing, forming a complex structure that is reinforced by the deposition of the extracellular matrix. When the cells proliferate and adjust to their new environment, the biofilm reaches maturity. Planktonic cells are eventually released from the adult biofilm upon reaching a specific cell density, or quorum, at which point they disperse to form new biofilms and initiate the cycle again.
In this review, we present the information collected from PubMed, Scopus, Google, and Google Scholar using the phrases like “Biofilm”, “Biofilm + Antibiotic tolerance”, “Antibiofilm agents”, “NPs + Biofilms”, “Chemical agents + Biofilms”, Biological agents + Biofilms”, “Physical agents + Biofilms”, “Biofilms of Antibiotic Resistant bacteria”, and “Components of Biofilms”.
2. Formation of Biofilms
On any surface, biofilms develop in three stages. Cell adherence to a surface is the first step, leading to cell assembly to create microcolonies and biofilm growth into a mature structure (Figure 1). Weak van der Waals forces first bring planktonic (free-floating) cells into contact with a surface, thereby causing their irreversible attachment to the same. Once established, the cells begin to secrete extracellular polymeric substances (EPS) and proteins called adhesins. Bacterial deposition is influenced by sedimentation, Brownian motion, and hydrodynamic forces [12]. Biofilm-associated protein (BAP), SasG, protein A, fibronectin-binding proteins, OmpA, and several other surface-associated proteins participate in biofilm formation, particularly in the initial phases of attachment [13]. Cell-to-cell contact via quorum sensing (QS) stimulates alterations in gene expression, and these connected cells proliferate and create microcolonies while producing more EPS. Bacterial communities grow in an increasingly ordered manner with established metabolic cooperation as the biofilm ages, forming intricate three-dimensional structures with water pathways for nutrient movement. The complex architecture, dense bacterial populations, thick EPS matrix, established nutritional gradients, and improved resistance to antimicrobials are characteristics of a mature biofilm. Eventually, the biofilm proceeds into the dispersal phase, characterized by sloughing off larger portions, releasing single cells [14]. However, when comparing the efficacy of EPS-targeting strategies to quorum sensing (QS) inhibition and physical clearance techniques, a distinct variation in efficiency profiles emerges. EPS-targeting methods, such as enzymatic degradation or nanoparticle-mediated disruption, are particularly effective in weakening the physical integrity of mature biofilms and enhancing antibiotic penetration [15]. For example, silver nanoparticles have been shown to reduce MRSA and MRSE biofilm formation by over 95% through direct EPS disruption [16], thereby enhancing the delivery and action of antimicrobial agents. In contrast, QS inhibitors primarily function during the early stages of biofilm formation by interfering with cell-to-cell communication and gene expression responsible for virulence and EPS production [17]. Usually, these are important factors in preventing biofilm development; their effects on established biofilms are limited and indirect. While physical methods like photoinactivation using blue light can achieve rapid biofilm reduction within minutes, they do not rely on specific bacterial pathways, making them effective against both drug-sensitive and drug-resistant strains [18]. Thus, EPS-targeting approaches may offer superior efficacy for mature biofilms, particularly when used in combination with antibiotics, whereas QS inhibitors are more preventive, and physical methods provide rapid, nonspecific clearance. A combination of these strategies may provide the most effective results in complex, multi-species, and clinically persistent biofilm environments [18].
3. Quorum Sensing (QS)
Biofilms employ QS, a cell-to-cell communication mechanism, to control their population density. This complex regulatory mechanism prevents the density of biofilm cells from becoming too high, making use of signaling molecules called autoinducers. The producers of autoinducers are none other than the bacterial cells, and their quantity rises with increasing cell density. The extent of coordination is the critical threshold level of autoinducers reached at a specific microbial cell concentration [19]. During this time, several target genes are either stimulated or suppressed by autoinducer receptor binding. Through synchronizing virulence features and attaining optimal biofilm formation, this QS system control enables bacteria to react in a coordinated manner to assist the whole microbial community [20]. The bacterial population is able to adapt to various situations because of this concerted functioning, which causes the biofilm to operate much like a multicellular organism. QS not only maintains the bacterial population size but also plays a role in regulating food availability for the bacteria, promoting antibiotic resistance, and aiding spread mutations across the biofilm colony that are advantageous to the microbe [21]. Furthermore, by modulating motility genes, EPS generation, and cell surface characteristics, QS plays essential roles in biofilm development and maturation. The system also helps bacterial populations coordinate their defensive mechanisms against competitor microbes and host immunological responses by producing specific metabolites and virulence factors. Because different bacterial species can employ different QS molecules, mixed-species biofilms can facilitate both species-specific communication and cross-species interactions.
4. Biofilm Antibiotic Tolerance (BAT)
As a thumb rule, bacteria in biofilms are really more resistant to antimicrobial intervention than their counterparts in planktonic habitats [22]. Although the mechanisms of antibiotic resistance of planktonic bacteria are usually well established (Figure 2), how this antibiotic resistance is brought about in biofilms is not well understood. However, the antimicrobial tolerance contributed by biofilms seems to operate differently. Interestingly, antimicrobial sensitivity is quickly recovered as soon as the biofilm-residing cells are separated from the main population, although earlier in the biofilm mode, the same cells exhibited antibiotic resistance [23]. Therefore, biofilm antibiotic tolerance (BAT) is expected to incorporate alternate paths to bacterial antimicrobial resistance [24]. These observed features suggest that biofilm-associated tolerance may involve supplementary or modified resistance strategies.
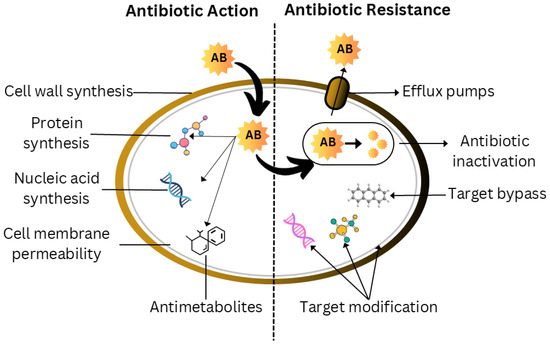
Figure 2.
Intricate relationships between antibiotic activity and bacterial resistance. Antibiotics primarily function by preventing the synthesis of proteins, nucleic acids, and cell walls; enhancing the permeability of cell membranes; and generating antimetabolites. However, in order to counteract these effects, bacteria have evolved a number of resistance mechanisms. These strategies include changing the target to prevent antibiotic binding, inactivating medications through modification or degradation, avoiding the target by other metabolic pathways, and actively removing antibiotics from the cell using efflux pumps. Clarifying these complex relationships is essential to the development of effective antibiotic treatments and strategies to combat the growing issue of antibiotic resistance.
BAT is the term used to describe the capability of the bacterial pathogens integrated in the form of biofilms to withstand antimicrobial treatment by making use of their pre-existing gene complement. Innate BAT (as a consequence of biofilm formation) and induced BAT (arising from the antimicrobial treatment response) are the two types of BAT witnessed. Restricted antibiotic penetration attributed to the prevalence of EPS [25], and reduced growth rate due to deprivation of oxygen and nutrients in internal areas of biofilms, thereby reduces antibiotic activity as antibiotics primarily impact actively dividing cells [26], and persistent cells that exhibit high levels of resistance to antibiotics while at rest [27] have all been identified as major innate factors that directly influence BAT. Furthermore, biofilms exhibit substantial horizontal gene transfer, which enables the antibiotic resistance genes to proliferate quickly across the population. Next, the three-dimensional structure of biofilms creates chemical gradients and microenvironments that can neutralize or inhibit antimicrobial agents. The biofilm matrix also contains extracellular DNA (eDNA), which can bind and sequester antimicrobial compounds, further contributing to resistance. Stress responses within biofilms can trigger increased expression of efflux pumps and other defense mechanisms. Additionally, following antimicrobial treatment, the existence of metabolically dormant “persisted cells” serves as a reservoir for biofilm regrowth.
5. Antibiofilm Agents
Antibiofilm agents can be categorized into three main types: physical agents like ultrasound waves and mechanical elimination that directly disrupt biofilm structure; chemical agents, including enzymes, acids, and antimicrobial compounds that break down or inhibit biofilm components; and biological agents such as bacteriophages that kill bacteria and probiotics which give competition to pathogenic bacteria that make the biofilms.
5.1. Physical Agents
5.1.1. Nanomaterials
Techniques based on nanotechnology are believed to have promise in the battle against infections linked to drug-resistant biofilms in medical devices and biomaterials. The use of nanoparticle (NP)-covered structures as biofilm inhibitory agents has been described in a modest number of research reports [28]. Although their sizes are comparable to those of biomolecules, nanomaterials have a far greater surface-to-volume ratio than biomolecules, which enhances their chemical and bioactivities. NPs are also tiny enough to pass through cell walls and biofilm coatings, causing permanent cellular damage.
New and exciting options for effective biofilm management and treatment have emerged due to recent breakthroughs in nanotechnology. Zinc, titanium, silver, gold, and copper have been found to have antibacterial and antibiofilm qualities, making them the right substitutes for antibiotics without appreciably raising the possibility of resistance. It was clearly demonstrated that metal-based NPs exhibit significantly better antibacterial qualities as compared to their microsized counterparts [29]. Silver nanoparticles have received a lot of attention since they have an oligodynamic impact with a wide range of antibacterial action, and they are especially good at fighting microbe colonization in biomedical illnesses. By interfering with and harming bacterial membranes, enzyme activity, electron transport chains, and cell growth through interaction of silver ions with sulfhydryl groups occurring in the bacterial walls, silver nanoparticles (Ag NPs) create an antibacterial impact (Figure 3). Ag NPs were also demonstrated to break biofilm structures by disrupting intermolecular interactions.
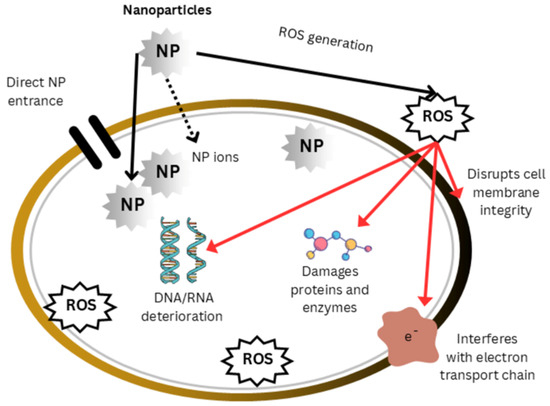
Figure 3.
Mode of action of nanoparticles. Nanoparticles affect the body through direct interactions with cellular components as well as indirect processes that involve the production of reactive oxygen species (ROS). The direct interactions result in the destruction of proteins and enzymes, the breakdown of nucleic acids, the disruption of electron transport systems (ETSs), and the loss of membrane integrity. These disruptions endanger energy production, cellular homeostasis, and genomic integrity, ultimately resulting in nanoparticle toxicity and cellular dysfunction.
After exposure to Ag NPs for 24 h, biofilm formation by clinically retrieved MRSA and methicillin-resistant S. epidermidis (MRSE) was less than 95%, while biofilm production by P. aeruginosa and S. epidermidis was also inhibited [30]. Bacterial adherence was reduced by silver-impregnated hydroxyapatite and silver–titania matrices. They prevented the formation of biofilms by both Gram-negative and Gram-positive bacteria, and TiO2 served as a superior matrix that inhibited the agglomeration of silver and permitted the continuous release of silver ions [31]. The antibacterial qualities of metal oxide nanoparticles were also investigated; ZnO NPs were shown to have superior antibacterial qualities and low toxicities in mammals, in addition to being more effective than other metal oxide NPs in preventing the growth of common pathogens and the creation of biofilms [32]. ZnO NPs have been demonstrated to cure skin wound infections and prevent the development of biofilms when used in conjunction with beta-chitin dressings in rat models. Furthermore, bacteriostatic and bacteria-resistant capabilities of nanotextured ZnO have been demonstrated to be superior to titania nanophase [33]. However, Proteus and P. aeruginosa have been shown to be resistant to zinc [34].
Compared to other metal oxides, nanosized TiO2 is also a safer and non-toxic antibacterial substance. Often serving as a photocatalyst, it is used in a number of environmentally beneficial processes, including antifogging and self-cleaning. Cu NPs have strong antibacterial activity against a range of microorganisms [35]. However, compared to Ag or Zn NPs, they appear to have fewer antibacterial qualities, requiring higher concentrations to provide the desired antimicrobial effects. At these levels, CuO NPs may be harmful to mammalian cells [36]. Although CuO NPs have exceptional antibacterial capabilities, their antibiofilm capabilities are limited by a narrow antibacterial window. For instance, in an NP-covered tooth prototype, CuO and ZnO NPs showed a potent biofilm inhibitory effect [37].
S. epidermis infestation on orthopedic implants was eradicated with iron NPs of 8 nm [38]. Additionally, antibiotic-conjugated magnetic iron nanoparticles performed better than unconjugated ferromagnetic NPs against E. faecalis in planktonic as well as biofilm forms [39]. Gold NPs have minimal to no antibacterial action on their own [40]. In spite of this, gold nanoparticles (NPs) linked to medications, active ingredients, or biomolecules have strong antibacterial and antibiofilm capabilities against a variety of illnesses, including MDR strains [41]. Gold NPs have been coupled with targeted molecules to provide particular antibiofilm actions since they are harmless to cells.
5.1.2. Photoinactivation
One novel method of treating biofilms is photoinactivation, which uses light to either directly knock down the bacteria by activating intracellular porphyrins that create ROS or to awaken an inert photosensitive dye that eventually emits dangerous reactive oxygen species [42]. The ability of porphyrins, both synthetic variants and indigenous compounds such as protoporphyrin IX, to generate singlet oxygen and other ROS upon activation by light has sparked interest in them as potent antimicrobial photosensitizers. Since these tetrapyrrolic chemicals preferentially concentrate in microbial cells and biofilms, they are particularly helpful for targeted antimicrobial treatment [43]. Blue light has shown an antimicrobial effect because of its cheapness and non-toxicity. Although UV radiation has previously been reported to be antibacterial, its mutagenic effects make it unsuitable for use in humans. Because the pathways of antibiotic resistance do not really alter the efficiency of UV inactivation, it has the benefit of being equally effective against both drug-resistant and sensitive microorganisms.
Blue light, particularly, has been examined for its efficacy against various bacteria and fungi [42]. Escherichia coli, Acinetobacter baumannii, Clostridium difficile (spores and vegetative cells), Klebsiella pneumoniae, Mycobacterium spp., Pseudomonas aeruginosa, Staphylococcus epidermidis, Streptococcus pyogenes, and Staphylococcus aureus, including methicillin-resistant Staphylococcus aureus (MRSA) strains, have all shown sensitivity in vitro [42]. Blue light, which has a wavelength of 405 nm, has been shown to be efficient against a variety of biofilms and to accomplish amazing destruction in 15 to 60 min [44].
To date, there has been no evidence of bacteria being resistant to photoinactivation therapy, although the synthesis of pigments by bacteria has been suggested as a resistance mechanism. In certain bacterial species, the production of pigments is associated with virulence. S. aureus is famous for generating triterpenoid carotenoids, which give colonies their golden appearance. The relation between pigment formation and species survival under light has already been discovered. Pigments have antioxidant properties that may guard cells against irradiation [45]. Additionally, the effectiveness of photoinactivation can be enhanced by combining it with photosensitizing agents or other antimicrobial treatments, creating a synergistic effect. The penetration depth of blue light through biofilms and tissue is an important consideration for clinical applications, as it can affect treatment efficacy in deeper-seated infections. This process is frequently brought on by environmental factors like stress or nutrient depletion, and the dispersed cells can then colonize newer surfaces, forming more and more biofilms. Cells coordinate using quorum sensing, forming a complex structure that is reinforced by the deposition of extracellular matrix. When the cells proliferate and adjust to their new environment, the biofilm reaches maturity. Planktonic cells are eventually released from the adult biofilm upon reaching a specific cell density, or quorum. From that point, they disperse to form new biofilms and initiate the cycle again.
5.1.3. Organic Nanoparticles
Emitting antibiotics, antimicrobials, or bacteriostatins, or modifying the polymeric nanoparticle surfaces and polymer-based articles with alkyl pyrimidines or quaternary ammonium compounds to enable killing just by contact, are methods used to obtain antibacterial properties. By causing cellular membrane rupture through mutual exchange of ions between live pathogenic bacteria and charged surfaces of polymers, the antibacterial polycationic groups may cause cell damage. By hydrophobic complexation, EPS polysaccharides interact with sulfate molecules of functionalized polystyrene NPs, disrupting biofilm formation [46]. Antibiofilm action against E. coli was discovered in a nanoporous matrix material made of sodium dodecyl sulfate. Phosphatidylcholine nanohybrid-coated conjugated poly (lactic-co-glycolic acid) NPs showed enhanced antibiofilm action against E. coli [47], and with a silicone-functionalized PDMS surface (known as brush design) were very successful in eliminating E. coli, S. aureus, and Candida albicans bacterial/fungal biofilms without causing cytotoxic effects in mammals [48].
5.1.4. Metal–Polymer Nanocomposites
Organic polymers have poor mechanical qualities for device applications. Nevertheless, layer-by-layer plasma polymerization, spin coating, or dip coating can be used to deposit them on metallic surfaces [49]. By controlling the release of Ga NPs, which Zn NPs were less effective at doing, Ga and Zn NPs combined into a polyether urethane hybrid PEG scaffold reduced P. aeruginosa infection in mice [50], and silicone TiO2 NP metal–polymer composites reduced S. aureus adhesion by 93% as compared to silicon that was not treated [51]. These composites also promise to prevent implant-associated infections due to their controlled release properties and ability to maintain long-term antimicrobial activity.
Dendrimers
These refer to 3-D structures that may contain both hydrophilic and hydrophobic substances in the vacuum regions of their heavily branching architectures. Without the use of extra antibiotics, the antimicrobial activity of the produced low molecular weight peptide dendrimers towards S. aureus and E. coli was shown [52]. Other research has shown that P. aeruginosa adhesion was disrupted and biofilm development was avoided when fucose-specific lectins (LecB) bonded to fucose peptide dendrimer ligands [53].
Cyclodextrins
Cyclodextrins (CDs) are glucopyranose-based cyclic chemical molecules that are used to liquefy hydrophobic substances in an aqueous phase. Membrane functionalization of CDs with polyethylene/polypropylene loaded with miconazole suppressed the growth of C. albicans biofilms by 96% in vitro. Furthermore, gold surface functionalized CD grafted anidulafungin and thymol were found to have fungicidal efficacy against C. albicans biofilms [54]. Additionally, CD–agar hydrogels loaded with ciprofloxacin showed broad antibacterial activity against S. aureus, S. epidermidis, P. aeruginosa, and E. coli in addition to controlled drug release at higher drug loading and retention [55].
5.1.5. Lipid-Based NPs and Microemulsions
Liposomes are being used for a variety of medicinal purposes, especially biofilm-related medicines, due to their resemblance to biological cell membranes. In cultured cells and on bacterial isolates of E. coli, P. aeruginosa, A. lwoffii, B. bronchiseptica, K. pneumoniae, and A. baumannii, different drug-loaded liposomes reduced lipase, protease, and chitinase, hindered biofilm formation, and interfered with quorum sensing [56]. Biofilm-forming bacteria have already been eradicated using a solid lipid nanoparticle (SLN) formulation comprising antimicrobial drugs. While one SLN formulation including eugenol demonstrated antifungal efficacy against oral candidiasis in a rodent model, another SLN preparation comprising PVA hydrogenated castor oil packed with tilmicosin was utilized to cure the infection in a mouse model of S. aureus-induced mastitis [47]. Microemulsions had a strong antibiofilm effect on C. albicans and P. aeruginosa by rupturing cytoplasmic cell walls, coagulating cytoplasm, and changing intracellular metabolism [57].
5.1.6. Nanofibers
A promising tactic against infections linked to biofilms is the use of antimicrobial systems based on nanofibers. Copper particles [35], iron chelators like DHBA [58,59], antimicrobial peptides like nisin [58], and antibiotics like ciprofloxacin [60] have all been shown to be effective in preventing the formation of biofilms by major pathogens like Pseudomonas aeruginosa and Staphylococcus aureus. These electrospun polymer nanofibers also provide for prolonged drug release and improved penetration capabilities for wound dressing applications.
5.1.7. Responsive Smart Nanoparticles
Energy-absorbing nanoparticles and external energy have been used in combination to effectively treat antimicrobial infections. Pathogenic cells suffer irreversible damage when metal nanoparticles (NPs) or polymer-based systems are activated by external energy sources such as visible light, temperature, near-infrared radiation (NIR), or high-frequency alternating magnetic fields (AMF) [47]. Gold, iron oxide, and graphene nanoparticles are examples of photothermal agents that have been used to absorb and transform NIR light into thermal energy. Gram-positive, Gram-negative, and mixed bacteria were thermally inactivated by subjecting gold or graphene nanoparticles to near-infrared radiation [61]. Temperatures of NP–bacterial suspensions were reported to be higher than the bacteria’s physiological limitations [47]. Recent developments have also shown promise in using pH-responsive nanoparticles that specifically target the acidic microenvironment of biofilms, allowing for targeted drug delivery. Additionally, magnetic field-responsive nanoparticles have shown potential in biofilm disruption through localized hyperthermia and enhanced drug penetration.
Nanoscale carriers were also utilized in photodynamic therapy (PDT), which uses light and photosensitizers to kill infections [62]. When photosensitizer–NP complexes are exposed to light, they create cytotoxic reactive oxygen species (ROS), which cause planktonic and biofilm bacteria to lyse. The efficacy of combining photosensitizers on NPs to eliminate specific pathogens or biofilms was investigated [63]. MRSA, Candida albicans, and multispecies bacterial biofilms were strongly inactivated by NPs complexed with porphyrin, methylene blue, or rose Bengal [64,65]. PDT has potential applications in treating wound infections, but practical usage is currently limited due to photosensitizers’ physicochemical properties, dosages provided, light dosimetry, and medicine release control.
While these nanotechnology-based approaches show great promise, several key factors need consideration for their effective implementation. The size and shape of nanoparticles significantly affect their antibiofilm efficacy; generally, smaller particles (1–100 nm) show better penetration into biofilms [29]. In contrast, specific shapes (like triangular silver nanoparticles) may exhibit enhanced antimicrobial activity compared to their spherical counterparts. Surface modification of nanoparticles through functionalization with specific ligands or polymers is crucial for improving their targeting ability, biocompatibility, and reducing potential toxicity to human cells [66]. However, translating these technologies from laboratory to clinical applications faces challenges, including scale-up issues, cost considerations, and regulatory requirements. Furthermore, the environmental impact of nanoparticle accumulation and its long-term effects on ecosystems must be carefully evaluated before widespread implementation. These considerations highlight the need for continued research into safer, more sustainable nanoparticle-based antibiofilm strategies.
5.1.8. Mechanisms of Antimicrobial and Antibiofilm Nanoparticles
The distinct physicochemical characteristics of nanoparticles (NPs) and their interactions with microbial cells are responsible for their antibacterial and antibiofilm capabilities. Key factors influencing their efficacy include the size of the nanoparticles, structure, surface charge, and composition. These properties determine how NPs interact with bacterial cells and biofilms, leading to various mechanisms of action. (a) Cell Membrane Modifications: NPs can adhere to the bacterial cell membrane, causing physical damage such as pore formation, membrane disruption, or leakage of cellular contents. The surface charge of NPs, which makes it easier for them to interact with the negatively charged bacterial membrane, frequently has an impact on this. (b) Failure of Respiratory Activity: NPs can interfere with the electron transport chain in bacterial cells, disrupting ATP production and leading to energy depletion. (c) Fatty Acid Peroxidation: Interaction of NPs with bacterial membranes can induce lipid peroxidation, compromising membrane integrity and function. (d) Reactive Oxygen Species (ROS) Production: The production of ROS by NPs, especially metal and metal oxide nanoparticles, can result in oxidative stress and harm cellular constituents such as proteins, lipids, and DNA. (e) DNA Unraveling: NPs can penetrate bacterial cells and interact with DNA, causing strand breaks, mutations, or inhibition of replication and transcription processes. (f) Conversion of Protein Thiols into Nitroso Derivatives: NPs can modify protein thiol groups into their nitroso derivatives, altering protein function and disrupting essential cellular processes. (g) Metabolic Pathway Disruptions: NPs may interfere with enzymatic activities and metabolic pathways, inhibiting vital cellular functions. In addition to their antibacterial effects, NPs exhibit antibiofilm activity by penetrating biofilms’ extracellular polymeric substances (EPS) matrix, disrupting biofilm structure, and inhibiting bacterial adhesion and colonization. Combining these mechanisms makes NPs a promising tool for combating antimicrobial resistance and biofilm-associated infections.
5.2. Chemical Agents
5.2.1. Phytochemicals
Natural chemicals with strong antibiofilm capabilities include alkaloids, essential oils, lectins, phenolics, polyacetylenes, polypeptides, and terpenoids (Table 1). Among the different types of phenolics, tannins exhibit antibiofilm action [67]. These substances affect biofilms by six distinct mechanisms: binding to proteins, interacting with eukaryotic DNA, causing substrate deprivation, disrupting membranes, binding to cell walls and adhesin complexes, and blocking viral fusion [68].

Table 1.
Phytochemicals having antibiofilm activities.
Phytochemicals are important in preventing bacterial adhesions and silencing genes involved in biofilm formation by inhibiting QS inducers such as AHL, autoinducers, and autoinducer type 2 from disrupting QS [80]. Interfering with forces is essential to promote biofilm structure, and restricting the availability of nutrients can stymie the initial growth of biofilms. Garlic extract has been demonstrated to suppress QS molecules in Pseudomonas and Vibrio spp. biofilms [81] and Emodin inhibit QS through proteolysis of linked transcription factors [82].
Reports suggest Psidium guajava L. (ethanolic and acetone extract), Eugenia spp. Extracts, and norbgugaine (an alkaloid) have an anti-adhesive effect [83]. Phytocompounds that hinder cell surface attachment are particularly polyphenols like 7-epiclusianone, casbane, and tannic acid. By targeting adhesin proteins, breaking up pre-formed biofilms, and preventing the production of EPS, the crude extract of A. philippense L. prevents biofilm development in its early stages [84].
Pyridones influence the expression of the CsgA curli component and prevent curli biogenesis. Phloretin, ginkgolic acid, and phytocompounds from Malaysian plants control the curli and pilli genes [85]. Even if recent research shows that an integrated approach has always been better than a customized approach for diseases linked to biofilms, phyto-compounds with less adverse effects could be a better therapeutic option.
5.2.2. Biosurfactants
By changing cellular adhesion capabilities through decreased hydrophobicity of the cell surface, membrane rupture, and blockage of the electron transport chain, biosurfactants (BS) stop the formation of biofilms and reduce cellular energy needs [86]. It was investigated how BS from L. plantarum and P. acidilactici affected the expression of genes linked to biofilms and QS signaling molecules in S. aureus. They discovered that by controlling the expression of genes linked to biofilms, such as dltB, icaA, cidA, and others, BS prevents S. aureus from producing biofilms. The BS from L. plantarum significantly reduced the cidA gene. P. acidilactici biosurfactant (sarA) downregulates staphylococcal accessory regulatory genes, autoinducer-2 (AI-2) signaling molecules, and accessory gene regulator (agrA) [87]. Table 2 outlines the biosurfactants that display antimicrobial activity.
Table 2.
Biosurfactants having antimicrobial activities.
Both Gram-positive and Gram-negative bacteria’s ability to produce biofilms can be inhibited by rhamnolipids and sophorolipids [95]. Cell-associated biosurfactant from L. acidophilus inhibits the formation of P. vulgaris and S. aureus biofilms on polydimethylsiloxane (PDMS)-based implants, according to certain studies [96].
5.2.3. Quaternary Ammonium Compounds
A hydrophilic quaternary ammonium group and a hydrophobic alkyl chain combine to form the amphiphilic structure of quaternary ammonium compounds (QACs). They are broad-spectrum bactericidal agents. Even though they are known as AMP mimics, their structures are greatly simpler. They work by rupturing the bacterial plasma membrane, which releases metabolites and causes cell lysis [97]. Researchers created mono-, bis-, and trisQACs that could destroy S. aureus and E. faecalis biofilms. Tris-QAC-10 was able to completely eliminate S. aureus biofilms that had established at 50 M and effectively eliminate E. faecalis biofilms that were already present at 25 M [98]. Despite the considerable eukaryotic cell toxicity of the published QACs, a class of multiQACs was reported that was both much less toxic than earlier QACs and had exceptional biofilm eradication efficacy (total destruction of S. aureus biofilms at 25 µm) [99]. Researchers have investigated the eradication activity of dicationic porphyrins QACs XF-70 and XF-73 against S. aureus biofilms as well as their antibacterial activities. They found that at 2.6 µm, they could eliminate biofilms [100].
QACs’ biofilm elimination characteristics make them a viable technique for treating biofilm-related illnesses, but their intrinsic toxicity is still an obstacle that must be solved. Otherwise, their clinical usage may be limited to primarily topical therapies. Furthermore, QACs’ biofilm eradication agents (BEA) action appears to be considerably more favorable in managing Gram-positive bacteria than Gram-negative pathogens, similar to AMPs. However, it is significantly more practical to artificially modify their fundamental structures to increase Gram-negative activity since they are smaller and simpler than AMPs.
5.2.4. Phenazines and Quinolines
Many bacteria, such as P. aeruginosa, Pantoea agglomerans, and Streptomyces spp. produce phenazines, which are redox-active secondary metabolites. The most well-known example is pyocyanin, a dibenzo-annulated pyrazine that is produced by P. aeruginosa [101]. Although they tend to be more sensitive, both Gram-positive and Gram-negative bacteria are susceptible to phenazines and their derivatives. Interest in using phenazines as BEAs was prompted by the finding that pyocyanin allowed P. aeruginosa biofilm infestations to outnumber S. aureus biofilm invasions in the lungs of individuals with cystic fibrosis [102].
Compounds based on phenazine have demonstrated a strong ability to remove S. aureus biofilms [103]. The most potent of these substances was bromophenazine-8, which completely destroyed biofilms at concentrations between 62.5 and 100 M. Halogenated phenazine-14 had the best biofilm eradication activity against all three diseases (S. aureus, S. epidermidis, and E. faecium), with complete removal taking place at concentrations ranging from 0.2 to 12 M [104]. Since halogenated phenazines do not harm mammalian cells, these substances or their derivatives may be appealing treatment choices for diseases linked to Gram-positive biofilms [104]. Halogenated quinolones have been shown to possess outstanding biofilm elimination activity, particularly against Gram-positive pathogens.
Quinolines are heterocyclic aromatic chemicals that resemble phenazines in structure and are commonly connected with antimalarial medications. Because of the resemblance between these two chemicals, quinolines are being studied as BEAs. Biofilm inhibitory activity towards S. aureus and S. epidermidis was found in a range of halogenated quinolines. However, biofilm eradication activity was lacking and needed to be improved. S. epidermidis biofilms were entirely destroyed by halogenated quinoline-3 at only 3.0 M, whereas E. faecium biofilms were totally eradicated by halogenated quinoline-4 at only 1.0 M [105].
It has been shown that quinolines and halogenated phenazines kill biofilms. However, it seems that only Gram-positive pathogens like S. aureus, S. epidermidis, and E. fecium are affected by them. Despite these compounds’ outstanding efficiency and lower cytotoxicity, no in vivo studies have been undertaken. Nonetheless, these chemicals are among the most intriguing BEAs ever discovered.
5.2.5. Antibiotics
Response of biofilms to antibiotics may be studied under three heads: mechanism of action, spectrum of action, and chemical structure. Both Gram-positive and Gram-negative bacteria can be treated with bactericidal or bacteriostatic antibiotics [106]. They are categorized as macrolides, lactams, aminoglycosides, glycopeptides, tetracyclines, and quinolones according to their molecular makeup.
By binding penicillin-binding proteins (PBPs), β-lactam limits the development of bacterial cell walls. PBPs are proteins that join the peptide units in the peptidoglycan sheet. Cell lysis occurs after β-lactams are linked to PBPs [107]. Monobactams, cephalosporins, carbapenems, carbacephems, and penicillins are among the lactam antibiotics. In the late 1960s, penicillin-resistant microorganisms were first reported. These bacteria produced the enzymes β-lactamases, which could break down β-lactam antibiotics. The introduction of carbapenem, a new family of β-lactams, alleviated this problem since carbapenem is not susceptible to β-lactamases. Carbapenems have the greatest breadth of action among all known lactams [108].
Glycopeptides also affect bacterial cell wall production, as well as blocking PBPs and inhibiting peptidoglycan synthesis [109]. Macrolides, oxazolidinones, and aminoglycosides inhibit bacterial growth by targeting protein synthesis in the cell’s ribosome. The suppression of mRNA extension during translation is caused by macrolide binding to the 50s ribosomal subunit [109]. Like macrolides, oxazolidinones bind to the 50s subunit and inhibit the development of a 70s initiation complex, limiting the synthesis of proteins [110]. The 50s class of blockers is made up of these two categories. By binding to the ribosomal 30s subunit, aminoglycosides and tetracycline stop aminoacyl-tRNA from getting to the ribosome, which lowers protein synthesis. While aminoglycosides have a broad bactericidal mode of action, tetracycline and macrolides are often bacteriostatic [109].
The production of nucleic acids is required for cell viability. By stopping DNA helicases from relaxing the double-helical helix of DNA, which is essential for reproduction and repair, quinolones inhibit the growth of bacteria. Additionally, quinolones impair the activity of DNA gyrase II and IV in bacteria, which negatively affects RNA polymerase and prevents the synthesis of RNA [109]. Para-aminobenzoic acid (PABA), a precursor for folic acid synthesis in bacterial cells, is structurally mimicked by sulfonamides. The latter inhibits the development of bacteria by preventing cell division, which is required to create nucleic acids [111]. Unfortunately, contemporary antibiotics are ineffective against these bacteria, making it more challenging to treat illnesses caused by these germs.
New enzymatic targets are being investigated for biofilm management in addition to conventional antibiotic methods. With several inhibitors demonstrating antibacterial action against uropathogenic E. coli, polyphosphate kinase 1 (PPK1) has become an attractive target [112]. PPK1 is a desirable target for the development of novel antimicrobial methods since it is essential for the bacterial stress response, pathogenicity, and biofilm formation.
5.2.6. Nanoparticles of Natural Products for the Eradication of Biofilms
Nanotechnology offers a transformative strategy in combating biofilm-associated infections, particularly when associated with the natural product-based compounds. The use of NPs synthesized from or conjugated with natural products combines the inherent antimicrobial properties of plant-derived phytochemicals [113]. Several natural products, such as phenolics, flavonoids, essential oils, terpenoids, and alkaloids, are recognized for their potent antibiofilm activity [114]. When formulated as nanoparticles, these agents demonstrate significantly improved performance against biofilms due to increased stability, targeted delivery, and the ability to break the extracellular polymeric substances (EPS) matrix. For instance, tannic acid, emodin, curcumin, and phytol compounds exhibit enhanced antibiofilm effects when incorporated into nanostructures, as they can disrupt quorum sensing, inhibit EPS production, and impair microbial adhesion [115].
Phytochemical-based nanoparticles can also overcome solubility and degradation issues often associated with crude plant extracts. Nanocarriers such as liposomes, solid lipid nanoparticles (SLNs), polymeric nanoparticles, and nanoemulsions have been used to encapsulate these natural molecules, improving their efficacy against both Gram-positive and Gram-negative bacteria. For example, lipid-based NPs loaded with eugenol or curcumin have been shown to inhibit biofilm formation and reduce virulence factor expression in Staphylococcus aureus and Pseudomonas aeruginosa [116]. Similarly, SLNs containing essential oils or phenolic acids have demonstrated robust antibiofilm activity by disrupting microbial membranes and interfering with biofilm architecture.
5.3. Biological Agents
5.3.1. Antimicrobial Peptides
AMPs are antimicrobials with broad-spectrum activity commonly used to treat microbial biofilms. Antibiotics known as AMPs work by targeting the bacterial cell membrane, which reduces the likelihood of bacterial resistance [117]. AMPs are found in animals (including humans), plants, and microorganisms. They work by electrostatically engaging with phospholipids in the membrane and then inserting themselves into the membrane to kill bacteria. Antimicrobial peptides and their effects from various sources are listed in Table 3. When AMPs and antimicrobial medications are combined, several molecular processes involved in the production of biofilms are blocked [118]. The primary drawback of using AMPs to treat diseases based on biofilms is their extreme vulnerability to bacterial protease degradation.
Table 3.
Antimicrobial peptides and their effects.
Biofilm formation by P. aeruginosa was shown to be inhibited by the membrane-disturbing activity of esculentin-1a, also known as Esc (1-21), an AMP derived from frog skin, and its D-amino acid-containing diastereomer, Esc (1-21)-1c. This Esc (1-21)-1c has previously been shown to have a potential effect against cystic fibrosis patients’ persistent lung Pseudomonas infections [127]. D-amino acids at Leu14 and Ser17 in the AMP esculentin-1a sequence improve AMP stability [127], decrease the swimming, swarming, and twitching motility of P. aeruginosa, and ultimately prevent the development of biofilms. The peptide works in three ways to prevent P. aeruginosa biofilm development. By downregulating the fleN gene, which controls the pace of flagella in P. aeruginosa, it first prevents flagella-mediated swimming. Secondly, at very low doses, like 1/8 MIC, it slows P. aeruginosa twitching motility and reduces the mRNA expression of type IV pili synthesis genes, both of which are critical for microcolony formation and colonization as a part of biofilm development. Lastly, it prevents the synthesis of the lasB gene, which codes for the virulence factor elastase LasB, and the lasI gene, which codes for the AHL synthase QS molecule [127].
5.3.2. Antimicrobial Lipids
Fatty acids and monoglycerides are examples of single-chain lipid amphiphiles, which are antimicrobial in action. Since Koch et al. originally documented the antibacterial properties of soap in the 1800s and later on reported that fatty acids may inhibit the growth of B. anthracis, the antimicrobial properties of these substances were made clear [128]. The bactericidal qualities of monoglycerides and fatty acids have since then been the subject of research. Antimicrobial modus operandi of lipids includes cell lysis, heightened permeability of membranes, disruption of the electron transport chain, and inhibition of bacterial enzymes [129].
Marshall and co-workers were among the first to look at the ability of heat and glycerol monolaurate to eradicate foodborne pathogen L. monocytogenes biofilms. Glycerol monolaurate (182 µm) and heat (65 °C) were shown to entirely eradicate 7-day-old adherent cells (biofilms) following just 5 min of contact time. In a subsequent investigation, Schlievert and Peterson demonstrated that glycerol monolaurate alone completely destroyed S. aureus and H. influenzae biofilms at a concentration of 1822 µm [130]. Lopes et al. developed a glycerol monolaurate nanocapsule that reduced P. aeruginosa biofilm development by up to 78% when administered at a concentration of 228 µm [131]. These studies demonstrate the partial removal of biofilms by glycerol monolaurate. The active concentration required for biofilm eradication remains significant, particularly for certain diseases, compared to other BEAs.
The antibiofilm activity of two fatty acids, docosahexaenoic acid and eicosatetraenoic acid, against Porphyromonas gingivalis and Fusobacterium nucleatum biofilms was examined. They discovered that at 100 µm, they were eliminating a notable percentage of live cells of P. gingivalis (61% and 47%, respectively) [132]. In a follow-up study, the same group tested the efficacy of the same two fatty acids against Streptococcus mutans biofilms. They significantly damaged the outer membrane of cells that resided in the biofilm (58.8% and 62.5%), reducing the thickness of the biofilm by 19% and 42%, respectively [133]. Crucially, a number of studies have shown that human cells are not harmed by dosages of docosahexaenoic acid up to 100 M and eicosapentaenoic acid up to 200 M [134]. The potential of antimicrobial lipids as BEAs is unquestionably exciting because of their low toxicity and superior antibiofilm and biofilm removal capabilities. Since the majority of human diets contain both monoglycerides and fatty acids, it is important to identify and look into the possibility of regular exposure to these substances and the resultant resistance.
5.3.3. Bacteriophages
Bacteriophages, a diverse group of viruses known for killing bacteria, have been conventionally used as antibacterial substances for years. Although the majority of this use was not controlled and was documented in anecdotal reports, phages have been employed therapeutically to treat infections of E. coli, S. aureus, K. pneumoniae, S. Typhimurium, and P. aeruginosa. A vast range of phage concentrations, phage varieties, and administration methods were documented [135]. Phage therapy has reemerged as an option to kill the infecting bacteria, considering the increase in AMR as multidrug resistance mechanisms that are becoming common among the latter. Bacteriophages are very specific and work by taking over the host machinery for their multiplication soon after infection. Phage recognizes and binds to target cells with the help of its tail fibers, which are highly specific, followed by the release of phage nucleic acid into bacterial cells either mechanically or enzymatically. Subsequently, viral nucleic acid is used to replicate itself using host enzymes. Virions are released by cell bursting when they attain a burst size.
Though the use of phages in therapeutics provides an edge, there is a need for a quick and accurate diagnosis to use appropriate phages against pathogens. The possibility of genetically editing phages to change their features is an intriguing promise for phage technology. For instance, shifts in host specificity and the addition of enzymes that accelerate extracellular matrix constituent breakdown have improved biofilm eradication efficiency [136].
Being particular for strains of bacteria has an advantage, and employing phages as antibiofilms is a unique notion with proof to back it up. The phage’s ability to attack biofilms has long been recognized, making it a great potential tool in curing biofilm-associated infections. Phages can infiltrate biofilms and diminish the viable count by moving through the bacterial population. Phage has also shown promise in avoiding Staphylococci or Pseudomonas infection of catheters [137]. Infections in wounds are potential therapeutic areas for phage use since the administration is simple and therapy progression can be tracked easily.
Few studies specifically address or evaluate cytotoxicity, despite the fact that phages are often recognized as generally regarded as safe (GRAS) owing to their widespread usage and absence of obvious harm from the available clinical data.
5.3.4. Collaborative Therapy Strategy
Recent research in antibiofilms focused on the effectiveness of collaborative or combination therapies, particularly those that provide strength to phage–antibiotic synergy (PAS). This approach utilizes the complementary mechanisms of bacteriophages and antibiotics to enhance biofilm eradication. One of the major advantages of PAS is that bacteriophages can selectively disrupt the biofilm matrix and lyse bacterial cells, which improves the penetration and efficacy of antibiotics, especially against resistant strains embedded within biofilms [138]. A notable example of this approach involves engineered bacteriophages carrying β-lactamase inhibitors, which have been shown to significantly enhance the activity of β-lactam antibiotics such as meropenem [139]. In a previous study, phages engineered to deliver β-lactamase inhibitors increased antibiotic efficacy by up to 100-fold in biofilm environments formed by multidrug-resistant bacteria [140]. This demonstrates how genetically modified phages can not only lyse bacteria but also modulate resistance mechanisms at the infection site. Furthermore, PAS has shown effectiveness in both planktonic and biofilm-associated infections in in vitro and in vivo models [141]. The dual mode of action can delay or reduce the development of antibiotic resistance and enhance treatment specificity by targeting resistant subpopulations, including persisted cells. Besides β-lactamase inhibitors, other strategies include combining phages with aminoglycosides, fluoroquinolones, or even antimicrobial peptides, depending on the pathogen type and resistance profile. Some studies have shown that sub-MIC concentrations of antibiotics can enhance phage adsorption and replication, further potentiating bacterial clearance [142].
Advanced monitoring tools that enable real-time evaluation of therapy efficacy are increasingly supporting the success of collaborative treatments. Researchers can now track the kinetics of biofilm development during therapeutic interventions thanks to novel fluorescent probes that see c-di-GMP in live bacterial cells [143]. Our comprehension of how combination medicines impact bacterial communication networks and biofilm architecture is improved by these real-time monitoring capabilities.
5.3.5. CRISPR
A relatively novel method for altering the genomes of prokaryotes and eukaryotes is the CRISPR-Cas9 system (clustered regularly interspaced short palindromic repeats and CRISPR-associated protein 9) [144]. Zuberi et al. recently employed CRISPR technology to control biofilm-associated infections by knocking down the luxS gene and the fimbriae-associated gene (fimH) [145,146]. Hegde et al. employed CRISPR/Cas9 to delete the ompA (outer membrane protein A) gene in the dominant mosquito microbiome, Cedecea neteri [147]. The ompA mutant had a lower capacity to build biofilms, had a lower infection rate in adults, and was also reported to be less frequent in adults [147]. As a result, CRISPR is another viable biofilm management method.
6. Challenges and Prospects
Intricate and dynamic interactions among the area, the incoming pathogenic bacterium, and the extracellular polymeric substances trigger biofilm production. Biofilms’ adhesive strength and viscoelastic properties make removing them from surfaces difficult, and resident bacteria acquire antimicrobial resistance. The fact that antimicrobials themselves will remain as biofilm leftovers (containing damaged cells), enabling microbial reuse and stimulating the colonization of additional bacteria, is the fundamental thing that determines that biofilms are challenging to eliminate. Following a significant dispersion incident, the discharge of planktonic cells can overwhelm the host’s immune system for such dispersing mediators with little antimicrobial effect. Further, scattered cells have already been demonstrated to become more pathogenic than biofilms and ordinary planktonic cells. This emphasizes the need to combine treatment approaches that tackle the EPS matrix and the bacteria that reside there. A drug’s ability to penetrate existing biofilms should be evaluated in addition to the hazards mentioned above, since this characteristic may result in de novo antimicrobial resistance by exposing bacteria to sub-lethal antibiotic dosages and influencing potential cytotoxicity. It may be advantageous to target the pathogenic microenvironment for aggressive biofilm dispersal in response to environmental factors such as hypoxia, acidic pH, and metabolites produced by the pathogen. We can break down the matrix and destroy the existing bacteria in this way, removing the infectious niche with accuracy and minimum damage to nearby tissues. With the expanding study scope about using dispersion agents to remove pharmaceutical biofilms, in vivo investigations and clinical testing are severely constrained. It is very difficult to extrapolate these findings to more complicated, multi-species biofilms in in vivo environments because the majority of the study was conducted in vitro on monospecies biofilms.
When using proteases and other enzymes that cause destruction, host toxicity should likely be considered, as well as inhibitory relationships within the host environment, such as proteolytic degradation or relatively mild inhibition of therapeutic drugs. Many of these issues make dispersion agents extremely challenging to adopt in practice. Notwithstanding these restrictions, biofilm dispersion research remains a thriving and interesting subject. Dispersal chemicals can either drive bacteria to actively tear down their biofilms or attack EPS on a molecular level. One potential new area that may hold the key to future biofilm eradication is multi-targeted or combinatorial therapy. Better clinical treatment of biofilm-related diseases and the resolution of problems like antibiotic resistance and soon-to-be-available medications may result from an understanding of the properties of biofilm dispersion agents.
7. Conclusions
Biofilms cause a formidable challenge in both clinical and environmental concerns due to their ability to confer enhanced antimicrobial resistance and persistence to pathogenic microorganisms. The EPS matrix plays a critical role in shielding biofilm-embedded cells from antibiotics and host immune responses, while quorum sensing, horizontal gene transfer, and the presence of dormant persisted cells further make it difficult to eradicate. Conventional antibiotics often fail to penetrate biofilms effectively, necessitating the development of specialized antibiofilm strategies. This review highlights a wide array of current and emerging approaches, including physical, chemical, and biological agents targeting various stages and components of biofilm formation and maturation.
Moreover, the integration of collaborative therapeutic strategies such as phage antibiotic synergy (PAS) represents a significant advancement in the fight against biofilm-associated infections. By simultaneously providing the targeted lytic activity of bacteriophages and the broad-spectrum efficacy of antibiotics, PAS offers a robust, comprehensive approach to overcoming the structural and metabolic defenses of mature biofilms. The ability of engineered phages to deliver resistance-modulating agents, such as β-lactamase inhibitors, further enhances antibiotic efficacy, with some studies demonstrating up to a 100-fold increase in effectiveness. As research in this area progresses, PAS exemplifies the evolving paradigm of personalized and precision medicine in antimicrobial therapy, and its inclusion in the antibiofilm arsenal may be key to addressing the escalating challenge of multidrug-resistant biofilm-related infections.
Author Contributions
B.S., M.D., V.K., A.A. and J.J.A. contributed to the study’s conception and design. The first draft of the manuscript was written by B.S., M.D., V.K., A.A. and D.N.C., J.J.A. edited the manuscript extensively and wrote the final draft. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are included in the text; however, the reference raw data of this article will be made available by the authors, without undue reservation, to any qualified researcher.
Acknowledgments
The authors are thankful to Delhi Technological University (DTU), Swami Shraddhanand College (SSNC), and NIFTEM for providing the resources and support necessary to complete this paper. The access to research facilities at DTU and the academic environment at SSNC proved invaluable in this research and writing process. We are grateful for the opportunity to have benefited from such esteemed institutions.
Conflicts of Interest
Ahire JJ was employed by Dr. Reddy’s Laboratories Limited. Dr. Reddy’s Laboratories had no direct and indirect role in the design/analysis/writing/publication of this research article. Other authors have no conflicts of interest to declare.
References
- Marshall, K.C.; Stout, R.; Mitchell, R. Mechanism of the Initial Events in the Sorption of Marine Bacteria to Surfaces. J. Gen. Microbiol. 1971, 68, 337–348. [Google Scholar] [CrossRef]
- Geesey, G.G.; Mutch, R.; Costerton, J.W.; Green, R.B. Sessile bacteria: An important component of the microbial population in small mountain streams 1. Limnol. Oceanogr. 1978, 23, 1214–1223. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wuertz, S. Bacteria and archaea on Earth and their abundance in biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- de Vos, W.M. Microbial biofilms and the human intestinal microbiome. NPJ Biofilms Microbiomes 2015, 1, 15005. [Google Scholar] [CrossRef] [PubMed]
- Mahto, K.U.; Das, S. Bacterial biofilm and extracellular polymeric substances in the moving bed biofilm reactor for wastewater treatment: A review. Bioresour. Technol. 2022, 345, 126476. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Datta, S.; Narayanan, K.B.; Rajnish, K.N. Bacterial exo-polysaccharides in biofilms: Role in antimicrobial resistance and treatments. J. Genet. Eng. Biotechnol. 2021, 19, 140. [Google Scholar] [CrossRef] [PubMed]
- Sonawane, J.M.; Rai, A.K.; Sharma, M.; Tripathi, M.; Prasad, R. Microbial biofilms: Recent advances and progress in environmental bioremediation. Sci. Total. Environ. 2022, 824, 153843. [Google Scholar] [CrossRef]
- Campoccia, D.; Montanaro, L.; Arciola, C.R. Extracellular DNA (eDNA). A Major Ubiquitous Element of the Bacterial Biofilm Architecture. Int. J. Mol. Sci. 2021, 22, 9100. [Google Scholar] [CrossRef]
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef]
- Vandana; Das, S. Genetic regulation, biosynthesis and applications of extracellular polysaccharides of the biofilm matrix of bacteria. Carbohydr. Polym. 2022, 291, 119536. [Google Scholar] [CrossRef]
- Santore, M.M. Interplay of physico-chemical and mechanical bacteria-surface interactions with transport processes controls early biofilm growth: A review. Adv. Colloid Interface Sci. 2022, 304, 102665. [Google Scholar] [CrossRef] [PubMed]
- Biswas, R.; Jangra, B.; Ashok, G.; Ravichandiran, V.; Mohan, U. Current Strategies for Combating Biofilm-Forming Pathogens in Clinical Healthcare-Associated Infections. Indian J. Microbiol. 2024, 64, 781–796. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Tang, X.; Dong, W.; Sun, N.; Yuan, W. A Review of Biofilm Formation of Staphylococcus aureus and Its Regulation Mechanism. Antibiotics 2022, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Asma, S.T.; Imre, K.; Morar, A.; Herman, V.; Acaroz, U.; Mukhtar, H.; Arslan-Acaroz, D.; Shah, S.R.A.; Gerlach, R. An Overview of Biofilm Formation–Combating Strategies and Mechanisms of Action of Antibiofilm Agents. Life 2022, 12, 1110. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-H. Nanoparticle-Based Therapies for Wound Biofilm Infection: Opportunities and Challenges. IEEE Trans. NanoBiosci. 2016, 15, 294–304. [Google Scholar] [CrossRef]
- Singh, S.; Bhatia, S. Quorum Sensing Inhibitors: Curbing Pathogenic Infections through Inhibition of Bacterial Communication. Iran J. Pharm Res. 2021, 20, 486–514. [Google Scholar] [CrossRef]
- Wu, X.; Wang, H.; Xiong, J.; Yang, G.-X.; Hu, J.-F.; Zhu, Q.; Chen, Z. Staphylococcus aureus biofilm: Formulation, regulatory, and emerging natural products-derived therapeutics. Biofilm 2024, 7, 100175. [Google Scholar] [CrossRef] [PubMed]
- Erkihun, M.; Asmare, Z.; Endalamew, K.; Getie, B.; Kiros, T.; Berhan, A. Medical Scope of Biofilm and Quorum Sensing during Biofilm Formation: Systematic Review. Bacteria 2024, 3, 118–135. [Google Scholar] [CrossRef]
- Mahto, K.U.; Kumari, S.; Das, S. Unraveling the complex regulatory networks in biofilm formation in bacteria and relevance of biofilms in environmental remediation. Crit. Rev. Biochem. Mol. Biol. 2022, 57, 305–332. [Google Scholar] [CrossRef]
- D’aquila, P.; De Rose, E.; Sena, G.; Scorza, A.; Cretella, B.; Passarino, G.; Bellizzi, D. Quorum Quenching Approaches against Bacterial-Biofilm-Induced Antibiotic Resistance. Antibiotics 2024, 13, 619. [Google Scholar] [CrossRef] [PubMed]
- Shree, P.; Singh, C.K.; Sodhi, K.K.; Surya, J.N.; Singh, D.K. Biofilms: Understanding the structure and contribution towards bacterial resistance in antibiotics. Med. Microecol. 2023, 16, 10084. [Google Scholar] [CrossRef]
- Dutt, Y.; Dhiman, R.; Singh, T.; Vibhuti, A.; Gupta, A.; Pandey, R.P.; Raj, V.S.; Chang, C.-M.; Priyadarshini, A. The Association between Biofilm Formation and Antimicrobial Resistance with Possible Ingenious Bio-Remedial Approaches. Antibiotics 2022, 11, 930. [Google Scholar] [CrossRef]
- Verderosa, A.D.; Totsika, M.; Fairfull-Smith, K.E. Bacterial Biofilm Eradication Agents: A Current Review. Front. Chem. 2019, 7, 495483. [Google Scholar] [CrossRef]
- Davenport, E.K.; Call, D.R.; Beyenal, H. Differential Protection from Tobramycin by Extracellular Polymeric Substances from Acinetobacter baumannii and Staphylococcus aureus Biofilms. Antimicrob. Agents Chemother. 2014, 58, 4755–4761. [Google Scholar] [CrossRef] [PubMed]
- Uruén, C.; Chopo-Escuin, G.; Tommassen, J.; Mainar-Jaime, R.C.; Arenas, J. Biofilms as Promoters of Bacterial Antibiotic Resistance and Tolerance. Antibiotics 2020, 10, 3. [Google Scholar] [CrossRef]
- Wood, T.K.; Knabel, S.J.; Kwan, B.W. Bacterial Persister Cell Formation and Dormancy. Appl. Environ. Microbiol. 2013, 79, 7116–7121. [Google Scholar] [CrossRef]
- Lahiri, D.; Nag, M.; Sheikh, H.I.; Sarkar, T.; Edinur, H.A.; Pati, S.; Ray, R.R. Microbiologically-Synthesized Nanoparticles and Their Role in Silencing the Biofilm Signaling Cascade. Front. Microbiol. 2021, 12, 636588. [Google Scholar] [CrossRef]
- Raha, S. Ahmaruzzaman ZnO nanostructured materials and their potential applications: Progress, challenges and perspectives. Nanoscale Adv. 2022, 4, 1868–1925. [Google Scholar] [CrossRef]
- Moradi, F.; Ghaedi, A.; Fooladfar, Z.; Bazrgar, A. Recent advance on nanoparticles or nanomaterials with anti-multidrug resistant bacteria and anti-bacterial biofilm properties: A systematic review. Heliyon 2023, 9, e22105. [Google Scholar] [CrossRef]
- Naik, K.; Kowshik, M. Anti-biofilm efficacy of low temperature processed AgCl–TiO2 nanocomposite coating. Mater. Sci. Eng. C 2014, 34, 62–68. [Google Scholar] [CrossRef]
- Lee, J.-H.; Kim, Y.-G.; Cho, M.H.; Lee, J. ZnO nanoparticles inhibit Pseudomonas aeruginosa biofilm formation and virulence factor production. Microbiol. Res. 2014, 169, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Sathiyaseelan, A.; Naveen, K.V.; Zhang, X.; Han, K.; Wang, M.-H. Research progress on chitosan-zinc oxide nanocomposites fabrication, characterization, biomedical and environmental applications. Coord. Chem. Rev. 2023, 496, 215398. [Google Scholar] [CrossRef]
- Webster, T.J.; Geilich, B.M. Reduced adhesion of Staphylococcus aureus to ZnO/PVC nanocomposites. Int. J. Nanomed. 2013, 8, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Ahire, J.J.; Hattingh, M.; Neveling, D.P.; Dicks, L.M.T.; Omri, A. Copper-Containing Anti-Biofilm Nanofiber Scaffolds as a Wound Dressing Material. PLoS ONE 2016, 11, e0152755. [Google Scholar] [CrossRef] [PubMed]
- Naz, S.; Gul, A.; Zia, M.; Mukhtiar, A. Toxicity of copper oxide nanoparticles: A review study. IET Nanobiotechnol. 2020, 14, 1–13. [Google Scholar] [CrossRef]
- Ahmed, M.E.; Faiq, N.H.; Almutairi, H.H.; Alam, M.W. Biosynthesized ZnO-CuO Nanocomposite for Biofilm Formation of Proteus mirabilis upon LuxS Gene Expression. Inorganics 2025, 13, 65. [Google Scholar] [CrossRef]
- Mohanta, Y.K.; Chakrabartty, I.; Mishra, A.K.; Chopra, H.; Mahanta, S.; Avula, S.K.; Patowary, K.; Ahmed, R.; Mishra, B.; Mohanta, T.K.; et al. Nanotechnology in combating biofilm: A smart and promising therapeutic strategy. Front. Microbiol. 2023, 13, 1028086. [Google Scholar] [CrossRef]
- Chifiriuc, M.C.; Grumezescu, A.M.; Andronescu, E.; Ficai, A.; Cotar, A.I.; Grumezescu, V.; Bezirtzoglou, E.; Lazar, V.; Radulescu, R. Water dispersible magnetite nanoparticles influence the efficacy of antibiotics against planktonic and biofilm embedded Enterococcus faecalis cells. Anaerobe 2013, 22, 14–19. [Google Scholar] [CrossRef]
- Yu, Q.; Li, J.; Zhang, Y.; Wang, Y.; Liu, L.; Li, M. Inhibition of gold nanoparticles (AuNPs) on pathogenic biofilm formation and invasion to host cells. Sci. Rep. 2016, 6, 26667. [Google Scholar] [CrossRef]
- Mu, H.; Tang, J.; Liu, Q.; Sun, C.; Wang, T.; Duan, J. Potent Antibacterial Nanoparticles against Biofilm and Intracellular Bacteria. Sci. Rep. 2016, 6, srep18877. [Google Scholar] [CrossRef] [PubMed]
- Enwemeka, C.S.; Baker, T.L.; Bumah, V.V. The role of UV and blue light in photo-eradication of microorganisms. J. Photochem. Photobiol. 2021, 8, 100064. [Google Scholar] [CrossRef]
- Amos-Tautua, B.M.; Songca, S.P.; Oluwafemi, O.S. Application of Porphyrins in Antibacterial Photodynamic Therapy. Molecules 2019, 24, 2456. [Google Scholar] [CrossRef]
- Ferrer-Espada, R.; Liu, X.; Goh, X.S.; Dai, T. Antimicrobial Blue Light Inactivation of Polymicrobial Biofilms. Front. Microbiol. 2019, 10, 721. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Naskar, A.; Acharya, K.; Mandal, S. Antibiofilm and antivirulence efficacy of Pleurotus platypus methanolic extract against Staphylococcus aureus through ROS generation and cell membrane disruption. Microb. Pathog. 2024, 196, 106992. [Google Scholar] [CrossRef]
- Fulaz, S.; Vitale, S.; Quinn, L.; Casey, E. Nanoparticle–Biofilm Interactions: The Role of the EPS Matrix. Trends Microbiol. 2019, 27, 915–926. [Google Scholar] [CrossRef]
- Ramasamy, M.; Lee, J. Recent Nanotechnology Approaches for Prevention and Treatment of Biofilm-Associated Infections on Medical Devices. BioMed Res. Int. 2016, 2016, 1–17. [Google Scholar] [CrossRef]
- Haktaniyan, M.; Bradley, M. Polymers showing intrinsic antimicrobial activity. Chem. Soc. Rev. 2022, 51, 8584–8611. [Google Scholar] [CrossRef]
- Hora, J.; Hall, C.; Evans, D.; Charrault, E. Inorganic Thin Film Deposition and Application on Organic Polymer Substrates. Adv. Eng. Mater. 2017, 20, 1700868. [Google Scholar] [CrossRef]
- Depan, D.; Misra, R. On the determining role of network structure titania in silicone against bacterial colonization: Mechanism and disruption of biofilm. Mater. Sci. Eng. C 2014, 34, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, N.P.S.; Yazdanpanah, A.; Mozafari, M. Polymer–metal nanocomposites with antimicrobial activity. In Biocidal Polymers; De Gruyter: Berlin, Germany, 2019; pp. 83–106. [Google Scholar]
- Paul, S.; Verma, S.; Chen, Y.-C. Peptide Dendrimer-Based Antibacterial Agents: Synthesis and Applications. ACS Infect. Dis. 2024, 10, 1034–1055. [Google Scholar] [CrossRef]
- Alfei, S.; Caviglia, D. Prevention and Eradication of Biofilm by Dendrimers: A Possibility Still Little Explored. Pharmaceutics 2022, 14, 2016. [Google Scholar] [CrossRef]
- Mirza, M.A.; Aggarwal, G.; Bharti, S.; Zakir, F. Antifungal Biofilm Strategies: A Less Explored Area in Wound Management. Curr. Pharm. Biotechnol. 2022, 23, 1497–1513. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Fernandez, B.; Lopez-Viota, M.; Concheiro, A.; Alvarez-Lorenzo, C. Synergistic performance of cyclodextrin–agar hydrogels for ciprofloxacin delivery and antimicrobial effect. Carbohydr. Polym. 2011, 85, 765–774. [Google Scholar] [CrossRef]
- Alhariri, M.; Omri, A. Efficacy of Liposomal Bismuth-Ethanedithiol-Loaded Tobramycin after Intratracheal Administration in Rats with Pulmonary Pseudomonas aeruginosa Infection. Antimicrob. Agents Chemother. 2013, 57, 569–578. [Google Scholar] [CrossRef]
- Al-Adham, I.S.; Ashour, H.; Al-Kaissi, E.; Khalil, E.; Kierans, M.; Collier, P.J. Studies on the kinetics of killing and the proposed mechanism of action of microemulsions against fungi. Int. J. Pharm. 2013, 454, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Ahire, J.J.; Dicks, L.M.T. Nisin Incorporated With 2,3-Dihydroxybenzoic Acid in Nanofibers Inhibits Biofilm Formation by a Methicillin-Resistant Strain of Staphylococcus aureus. Probiotics Antimicrob. Proteins 2015, 7, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Ahire, J.J.; Dicks, L.M.T. 2,3-Dihydroxybenzoic Acid-Containing Nanofiber Wound Dressings Inhibit Biofilm Formation by Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2014, 58, 2098–2104. [Google Scholar] [CrossRef]
- Ahire, J.J.; Neveling, D.P.; Hattingh, M.; Dicks, L.M.T.; Coenye, T. Ciprofloxacin-Eluting Nanofibers Inhibits Biofilm Formation by Pseudomonas aeruginosa and a Methicillin-Resistant Staphylococcus aureus. PLoS ONE 2015, 10, e0123648. [Google Scholar] [CrossRef]
- An, X.; Erramilli, S.; Reinhard, B.M. Plasmonic nano-antimicrobials: Properties, mechanisms and applications in microbe inactivation and sensing. Nanoscale 2021, 13, 3374–3411. [Google Scholar] [CrossRef]
- Escudero, A.; Carrillo-Carrión, C.; Castillejos, M.C.; Romero-Ben, E.; Rosales-Barrios, C.; Khiar, N. Photodynamic therapy: Photosensitizers and nanostructures. Mater. Chem. Front. 2021, 5, 3788–3812. [Google Scholar] [CrossRef]
- Ribeiro, M.; Gomes, I.; Saavedra, M.; Simões, M. Photodynamic therapy and combinatory treatments for the control of biofilm-associated infections. Lett. Appl. Microbiol. 2022, 75, 548–564. [Google Scholar] [CrossRef]
- Khan, A.; Azam, A.; Alam, F. Gold nanoparticles enhance methylene blue– induced photodynamic therapy: A novel therapeutic approach to inhibit Candida albicans biofilm. Int. J. Nanomed. 2012, 7, 3245–3257. [Google Scholar] [CrossRef]
- Shrestha, A.; Kishen, A. Antibiofilm Efficacy of Photosensitizer-functionalized Bioactive Nanoparticles on Multispecies Biofilm. J. Endod. 2014, 40, 1604–1610. [Google Scholar] [CrossRef]
- Ahmad, F.; Salem-Bekhit, M.M.; Khan, F.; Alshehri, S.; Khan, A.; Ghoneim, M.M.; Wu, H.-F.; Taha, E.I.; Elbagory, I. Unique Properties of Surface-Functionalized Nanoparticles for Bio-Application: Functionalization Mechanisms and Importance in Application. Nanomaterials 2022, 12, 1333. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, X.; Zhen, L.; Ares, J.N.; Vackier, T.; Lange, H.; Crestini, C.; Steenackers, H.P. Effect of chemical modifications of tannins on their antimicrobial and antibiofilm effect against Gram-negative and Gram-positive bacteria. Front. Microbiol. 2023, 13, 987164. [Google Scholar] [CrossRef] [PubMed]
- Elkhalifa, M.E.; Ashraf, M.; Ahmed, A.; Usman, A.; Hamdoon, A.A.; A Elawad, M.; Almalki, M.G.; Mosa, O.F.; Niyazov, L.N.; Ayaz, M. Polyphenols and Their Nanoformulations as Potential Antibiofilm Agents Against Multidrug-Resistant Pathogens. Futur. Microbiol. 2024, 19, 255–279. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, T.H.; van Gennip, M.; Phipps, R.K.; Shanmugham, M.S.; Christensen, L.D.; Alhede, M.; Skindersoe, M.E.; Rasmussen, T.B.; Friedrich, K.; Uthe, F.; et al. Ajoene, a Sulfur-Rich Molecule from Garlic, Inhibits Genes Controlled by Quorum Sensing. Antimicrob. Agents Chemother. 2012, 56, 2314–2325. [Google Scholar] [CrossRef]
- Reiter, J.; Levina, N.; Van der Linden, M.; Gruhlke, M.; Martin, C.; Slusarenko, A.J. Diallylthiosulfinate (Allicin), a Volatile Antimicrobial from Garlic (Allium sativum), Kills Human Lung Pathogenic Bacteria, Including MDR Strains, as a Vapor. Molecules 2017, 22, 1711. [Google Scholar] [CrossRef]
- Yan, X.; Gu, S.; Shi, Y.; Cui, X.; Wen, S.; Ge, J. The effect of emodin on Staphylococcus aureus strains in planktonic form and biofilm formation in vitro. Arch. Microbiol. 2017, 199, 1267–1275. [Google Scholar] [CrossRef]
- Adeosun, I.J.; Baloyi, I.T.; Cosa, S. Anti-Biofilm and Associated Anti-Virulence Activities of Selected Phytochemical Compounds against Klebsiella pneumoniae. Plants 2022, 11, 1429. [Google Scholar] [CrossRef]
- Vikram, A.; Jesudhasan, P.R.; Pillai, S.D.; Patil, B.S. Isolimonic acid interferes with Escherichia coli O157:H7 biofilm and TTSS in QseBC and QseA dependent fashion. BMC Microbiol. 2012, 12, 261. [Google Scholar] [CrossRef]
- Vikram, A.; Jesudhasan, P.R.; Jayaprakasha, G.K.; Pillai, S.D.; Patil, B.S. Citrus limonoids interfere with Vibrio harveyi cell–cell signalling and biofilm formation by modulating the response regulator LuxO. Microbiology 2011, 157, 99–110. [Google Scholar] [CrossRef]
- Raorane, C.J.; Lee, J.-H.; Kim, Y.-G.; Rajasekharan, S.K.; García-Contreras, R.; Lee, J. Antibiofilm and Antivirulence Efficacies of Flavonoids and Curcumin Against Acinetobacter baumannii. Front. Microbiol. 2019, 10, 990. [Google Scholar] [CrossRef]
- Shehabeldine, A.M.; Ashour, R.M.; Okba, M.M.; Saber, F.R. Callistemon citrinus bioactive metabolites as new inhibitors of methicillin-resistant Staphylococcus aureus biofilm formation. J. Ethnopharmacol. 2020, 254, 112669. [Google Scholar] [CrossRef]
- Samoilova, Z.; Tyulenev, A.; Muzyka, N.; Smirnova, G.; Oktyabrsky, O. Tannic and gallic acids alter redox-parameters of the medium and modulate biofilm formation. AIMS Microbiol. 2019, 5, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Gartika, M.; Pramesti, H.T.; Kurnia, D.; Satari, M.H. A terpenoid isolated from sarang semut (Myrmecodia pendans) bulb and its potential for the inhibition and eradication of Streptococcus mutans biofilm. BMC Complement. Altern. Med. 2018, 18, 151. [Google Scholar] [CrossRef] [PubMed]
- Vashistt, J. Study on Natural Antimicrobial Compounds Against Bacterial Isolates. Ph.D. Thesis, Jaypee University, Solan, India, 2022. [Google Scholar]
- Ćirić, A.; Petrović, J.; Glamočlija, J.; Smiljković, M.; Nikolić, M.; Stojković, D.; Soković, M.D. Natural products as biofilm formation antagonists and regulators of quorum sensing functions: A comprehensive review update and future trends. S. Afr. J. Bot. 2019, 120, 65–80. [Google Scholar] [CrossRef]
- Lu, L.; Hu, W.; Tian, Z.; Yuan, D.; Yi, G.; Zhou, Y.; Cheng, Q.; Zhu, J.; Li, M. Developing natural products as potential anti-biofilm agents. Chin. Med. 2019, 14, 11. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-B.; Wang, S.; Wang, C.; Huang, Q.-Y.; Bai, J.-W.; Chen, J.-Q.; Chen, X.-Y.; Li, Y.-H. Emodin affects biofilm formation and expression of virulence factors in Streptococcus suis ATCC700794. Arch. Microbiol. 2015, 197, 1173–1180. [Google Scholar] [CrossRef]
- Mishra, R.; Panda, A.K.; De Mandal, S.; Shakeel, M.; Bisht, S.S.; Khan, J. Natural Anti-biofilm Agents: Strategies to Control Biofilm-Forming Pathogens. Front. Microbiol. 2020, 11, 566325. [Google Scholar] [CrossRef]
- Adnan, M.; Patel, M.; Deshpande, S.; Alreshidi, M.; Siddiqui, A.J.; Reddy, M.N.; Emira, N.; De Feo, V. Effect of Adiantum philippense Extract on Biofilm Formation, Adhesion With Its Antibacterial Activities Against Foodborne Pathogens, and Characterization of Bioactive Metabolites: An in vitro-in silico Approach. Front. Microbiol. 2020, 11, 823. [Google Scholar] [CrossRef]
- Yahya, M.F.Z.R. Anti-biofilm Potential and Mode of Action of Malaysian Plant Species: A Review. Sci. Lett. 2020, 14, 34. [Google Scholar] [CrossRef]
- Satputea, S.K.; Banpurkar, A.G.; Banat, I.M.; Sangshetti, J.N.; Patil, R.H.; Gade, W.N. Multiple Roles of Biosurfactants in Biofilms. Curr. Pharm. Des. 2016, 22, 1429–1448. [Google Scholar] [CrossRef]
- Yan, X.; Gu, S.; Cui, X.; Shi, Y.; Wen, S.; Chen, H.; Ge, J. Antimicrobial, anti-adhesive and anti-biofilm potential of biosurfactants isolated from Pediococcus acidilactici and Lactobacillus plantarum against Staphylococcus aureus CMCC26003. Microb. Pathog. 2019, 127, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Ohadi, M.; Forootanfar, H.; Dehghannoudeh, G.; Eslaminejad, T.; Ameri, A.; Shakibaie, M.; Adeli-Sardou, M. Antimicrobial, anti-biofilm, and anti-proliferative activities of lipopeptide biosurfactant produced by Acinetobacter junii B6. Microb. Pathog. 2020, 138, 103806. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aziz, M.M.; Al-Omar, M.S.; Mohammed, H.A.; Emam, T.M. In Vitro and Ex Vivo Antibiofilm Activity of a Lipopeptide Biosurfactant Produced by the Entomopathogenic Beauveria bassiana Strain against Microsporum canis. Microorganisms 2020, 8, 232. [Google Scholar] [CrossRef]
- Janek, T.; Drzymała, K.; Dobrowolski, A. In vitro efficacy of the lipopeptide biosurfactant surfactin-C15 and its complexes with divalent counterions to inhibit Candida albicans biofilm and hyphal formation. Biofouling 2020, 36, 210–221. [Google Scholar] [CrossRef]
- Karlapudi, A.P.; Venkateswarulu, T.C.; Srirama, K.; Kota, R.K.; Mikkili, I.; Kodali, V.P. Evaluation of anti-cancer, anti-microbial and anti-biofilm potential of biosurfactant extracted from an Acinetobacter M6 strain. J. King Saud Univ.—Sci. 2020, 32, 223–227. [Google Scholar] [CrossRef]
- Abdollahi, S.; Tofighi, Z.; Babaee, T.; Shamsi, M.; Rahimzadeh, G.; Rezvanifar, H.; Saeidi, E.; Amiri, M.M.; Ashtiani, Y.S.; Samadi, N. Evaluation of Anti-oxidant and Anti-biofilm Activities of Biogenic Surfactants Derived from Bacillus amyloliquefaciens and Pseudomonas aeruginosa. IJ Pharm. Res. 2018, 19, 115–126. [Google Scholar]
- Elshikh, M.; Funston, S.; Chebbi, A.; Ahmed, S.; Marchant, R.; Banat, I.M. Rhamnolipids from non-pathogenic Burkholderia thailandensis E264: Physicochemical characterization, antimicrobial and antibiofilm efficacy against oral hygiene related pathogens. New Biotechnol. 2017, 36, 26–36. [Google Scholar] [CrossRef]
- Sacco, L.P.; Castellane, T.C.L.; Polachini, T.C.; Lemos, E.G.d.M.; Alves, L.M.C. Exopolysaccharides produced by Pandoraea shows emulsifying and anti-biofilm activities. J. Polym. Res. 2019, 26, 91. [Google Scholar] [CrossRef]
- Sharahi, J.Y.; Azimi, T.; Shariati, A.; Safari, H.; Tehrani, M.K.; Hashemi, A. Advanced strategies for combating bacterial biofilms. J. Cell. Physiol. 2019, 234, 14689–14708. [Google Scholar] [CrossRef]
- Satpute, S.K.; Mone, N.S.; Das, P.; Banat, I.M.; Banpurkar, A.G. Inhibition of pathogenic bacterial biofilms on PDMS based implants by L. acidophilus derived biosurfactant. BMC Microbiol. 2019, 19, 39. [Google Scholar] [CrossRef]
- Saverina, E.A.; Frolov, N.A.; Kamanina, O.A.; Arlyapov, V.A.; Vereshchagin, A.N.; Ananikov, V.P. From Antibacterial to Antibiofilm Targeting: An Emerging Paradigm Shift in the Development of Quaternary Ammonium Compounds (QACs). ACS Infect. Dis. 2023, 9, 394–422. [Google Scholar] [CrossRef]
- Jennings, M.C.; Ator, L.E.; Paniak, T.J.; Minbiole, K.P.C.; Wuest, W.M. Biofilm-Eradicating Properties of Quaternary Ammonium Amphiphiles: Simple Mimics of Antimicrobial Peptides. ChemBioChem 2014, 15, 2211–2215. [Google Scholar] [CrossRef] [PubMed]
- Forman, M.E.; Jennings, M.C.; Wuest, W.M.; Minbiole, K.P.C. Building a Better Quaternary Ammonium Compound (QAC): Branched Tetracationic Antiseptic Amphiphiles. ChemMedChem 2016, 11, 1401–1405. [Google Scholar] [CrossRef]
- Nadar, S.; Khan, T.; Patching, S.G.; Omri, A. Development of Antibiofilm Therapeutics Strategies to Overcome Antimicrobial Drug Resistance. Microorganisms 2022, 10, 303. [Google Scholar] [CrossRef] [PubMed]
- Meirelles, L.A.; Newman, D.K. Both toxic and beneficial effects of pyocyanin contribute to the lifecycle of Pseudomonas aeruginosa. Mol. Microbiol. 2018, 110, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, L.E.P.; Okegbe, C.; Price-Whelan, A.; Sakhtah, H.; Hunter, R.C.; Newman, D.K. Bacterial Community Morphogenesis Is Intimately Linked to the Intracellular Redox State. J. Bacteriol. 2013, 195, 1371–1380. [Google Scholar] [CrossRef]
- Garrison, A.T.; Bai, F.; Abouelhassan, Y.; Paciaroni, N.G.; Jin, S.; Iii, R.W.H. Bromophenazine derivatives with potent inhibition, dispersion and eradication activities against Staphylococcus aureus biofilms. RSC Adv. 2015, 5, 1120–1124. [Google Scholar] [CrossRef]
- Garrison, A.T.; Abouelhassan, Y.; Kallifidas, D.; Bai, F.; Ukhanova, M.; Mai, V.; Jin, S.; Luesch, H.; Huigens, R.W. Halogenated Phenazines that Potently Eradicate Biofilms, MRSA Persister Cells in Non-Biofilm Cultures, and Mycobacterium tuberculosis. Angew. Chem. Int. Ed. Engl. 2015, 54, 14819–14823. [Google Scholar] [CrossRef]
- Basak, A.; Abouelhassan, Y.; Norwood, V.M.; Bai, F.; Nguyen, M.T.; Jin, S.; Huigens, R.W. Synthetically Tuning the 2-Position of Halogenated Quinolines: Optimizing Antibacterial and Biofilm Eradication Activities via Alkylation and Reductive Amination Pathways. Chem.—Eur. J. 2016, 22, 9181–9189. [Google Scholar] [CrossRef]
- Zhang, F.; Cheng, W. The Mechanism of Bacterial Resistance and Potential Bacteriostatic Strategies. Antibiotics 2022, 11, 1215. [Google Scholar] [CrossRef] [PubMed]
- Nikolaidis, I.; Favini-Stabile, S.; Dessen, A. Resistance to antibi—Otics targeted to the bacterial cell wall. Protein Sci. 2014, 23, 243–259. [Google Scholar] [CrossRef]
- Husna, A.; Rahman, M.; Badruzzaman, A.T.M.; Sikder, M.H.; Islam, M.R.; Rahman, T.; Alam, J.; Ashour, H.M. Extended-Spectrum β-Lactamases (ESBL): Challenges and Opportunities. Biomedicines 2023, 11, 2937. [Google Scholar] [CrossRef]
- Lee, V.C. The Antibiotic Resistance Crisis Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Tsai, K.; Stojković, V.; Lee, D.J.; Young, I.D.; Szal, T.; Klepacki, D.; Vázquez-Laslop, N.; Mankin, A.S.; Fraser, J.S.; Fujimori, D.G. Structural basis for context-specific inhibition of translation by oxazolidinone antibiotics. Nat. Struct. Mol. Biol. 2022, 29, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.; Zou, W.; Zhong, Y. Sulfonamides repress cell division in the root apical meristem by inhibiting folates synthesis. J. Hazard. Mater. Adv. 2022, 5, 100045. [Google Scholar] [CrossRef]
- Peng, L.; Zeng, L.; Jin, H.; Yang, L.; Xiao, Y.; Lan, Z.; Yu, Z.; Ouyang, S.; Zhang, L.; Sun, N. Discovery and antibacterial study of potential PPK1 inhibitors against uropathogenic E. coli. J. Enzym. Inhib. Med. Chem. 2020, 35, 1224–1232. [Google Scholar] [CrossRef]
- Díaz-Puertas, R.; Álvarez-Martínez, F.J.; Falco, A.; Barrajón-Catalán, E.; Mallavia, R. Phytochemical-Based Nanomaterials against Antibiotic-Resistant Bacteria: An Updated Review. Polymers 2023, 15, 1392. [Google Scholar] [CrossRef]
- Nwafor, I.R.; Alhassan, Y.; Udoh, J.I.; Odanibeh, D.; Oyaniyi, J.; Efoli-Bam, V.K.; Azubuike, E.O.; Ojobor, J.-F.C.; Nwokafor, C.V. Plant-derived Bioactive Compounds and Their Mechanistic Roles in Combating Microbial Biofilms. Microbiol. Res. J. Int. 2024, 34, 74–85. [Google Scholar] [CrossRef]
- Asfour, H.Z. Anti-quorum sensing natural compounds. J. Microsc. Ultrastruct. 2018, 6, 1–10. [Google Scholar] [CrossRef]
- Karaca, A.C.; Rezaei, A.; Qamar, M.; Assadpour, E.; Esatbeyoglu, T.; Jafari, S.M. Lipid-based nanodelivery systems of curcumin: Recent advances, approaches, and applications. Food Chem. 2024, 463, 141193. [Google Scholar] [CrossRef]
- Hirt, H.; Hall, J.W.; Larson, E.; Gorr, S.-U.; Kreth, J. A D-enantiomer of the antimicrobial peptide GL13K evades antimicrobial resistance in the Gram positive bacteria Enterococcus faecalis and Streptococcus gordonii. PLoS ONE 2018, 13, e0194900. [Google Scholar] [CrossRef] [PubMed]
- Shahrour, H.; Ferrer-Espada, R.; Dandache, I.; Bárcena-Varela, S.; Sánchez-Gómez, S.; Chokr, A.; Martinez-de-Tejada, G. AMPs as Anti-biofilm Agents for Human Therapy and Prophylaxis. In Antimicrobial Peptides: Basics for Clinical Application; Springer: Berlin/Heidelberg, Germany, 2019; pp. 257–279. [Google Scholar]
- Yuan, Y.; Zai, Y.; Xi, X.; Ma, C.; Wang, L.; Zhou, M.; Shaw, C.; Chen, T. A novel membrane-disruptive antimicrobial peptide from frog skin secretion against cystic fibrosis isolates and evaluation of anti-MRSA effect using Galleria mellonella model. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2019, 1863, 849–856. [Google Scholar] [CrossRef]
- Mohammadi, M.; Taheri, B.; Momenzadeh, N.; Salarinia, R.; Nabipour, I.; Farshadzadeh, Z.; Bargahi, A. Identification and Characterization of Novel Antimicrobial Peptide from Hippocampus comes by In Silico and Experimental Studies. Mar. Biotechnol. 2018, 20, 718–728. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xi, X.; Ma, C.; Chen, X.; Zhou, M.; Burrows, J.F.; Chen, T.; Wang, L. A Novel Dermaseptin Isolated from the Skin Secretion of Phyllomedusa tarsius and Its Cationicity-Enhanced Analogue Exhibiting Effective Antimicrobial and Anti-Proliferative Activities. Biomolecules 2019, 9, 628. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, L.; Wu, Y.; Wang, L.; Ma, C.; Xi, X.; Bininda-Emonds, O.R.; Shaw, C.; Chen, T.; Zhou, M. Evaluation of the bioactivity of a mastoparan peptide from wasp venom and of its analogues designed through targeted engineering. Int. J. Biol. Sci. 2018, 14, 599–607. [Google Scholar] [CrossRef]
- Khozani, R.S.; Shahbazzadeh, D.; Harzandi, N.; Feizabadi, M.M.; Bagheri, K.P. Kinetics Study of Antimicrobial Peptide, Melittin, in Simultaneous Biofilm Degradation and Eradication of Potent Biofilm Producing MDR Pseudomonas aeruginosa Isolates. Int. J. Pept. Res. Ther. 2019, 25, 329–338. [Google Scholar] [CrossRef]
- de Barros, E.; Gonçalves, R.M.; Cardoso, M.H.; Santos, N.C.; Franco, O.L.; Cândido, E.S. Snake Venom Cathelicidins as Natural Antimicrobial Peptides. Front. Pharmacol. 2019, 10, 1415. [Google Scholar] [CrossRef]
- Basso, V.; Tran, D.Q.; Schaal, J.B.; Tran, P.; Eriguchi, Y.; Ngole, D.; Cabebe, A.E.; Park, A.Y.; Beringer, P.M.; Ouellette, A.J.; et al. Rhesus Theta Defensin 1 Promotes Long Term Survival in Systemic Candidiasis by Host Directed Mechanisms. Sci. Rep. 2019, 9, 16905. [Google Scholar] [CrossRef]
- von Borowski, R.G.; Chat, S.; Schneider, R.; Nonin-Lecomte, S.; Bouaziz, S.; Giudice, E.; Zimmer, A.R.; Gnoatto, S.C.; Macedo, A.J.; Gillet, R. First-in-class matrix anti-assembly peptide prevents staphylococcal biofilm in vitro and in vivo. bioRxiv. 2020. [Google Scholar] [CrossRef]
- Casciaro, B.; Cappiello, F.; Loffredo, M.R.; Ghirga, F.; Mangoni, M.L. The Potential of Frog Skin Peptides for Anti-Infective Therapies: The Case of Esculentin-1a(1-21)NH2. Curr. Med. Chem. 2020, 27, 1405–1419. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Santhakumari, S.; Poonguzhali, P.; Geetha, M.; Dyavaiah, M.; Xiangmin, L. Bacterial Biofilm Inhibition: A Focused Review on Recent Therapeutic Strategies for Combating the Biofilm Mediated Infections. Front. Microbiol. 2021, 12, 676458. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jackman, J.A.; Valle-González, E.R.; Cho, N.-J. Antibacterial Free Fatty Acids and Monoglycerides: Biological Activities, Experimental Testing, and Therapeutic Applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef] [PubMed]
- Schlievert, P.M.; Peterson, M.L.; Kaufmann, G.F. Glycerol Monolaurate Antibacterial Activity in Broth and Biofilm Cultures. PLoS ONE 2012, 7, e40350. [Google Scholar] [CrossRef]
- Lopes, L.Q.S.; Vaucher, R.d.A.; Giongo, J.L.; Gündel, A.; Santos, R.C.V. Characterisation and anti-biofilm activity of glycerol monolaurate nanocapsules against Pseudomonas aeruginosa. Microb. Pathog. 2019, 130, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Zhou, Z.; Dong, J.; Zhang, J.; Xia, Y.; Shu, R. Antibacterial and antibiofilm activities of docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) against periodontopathic bacteria. Microb. Pathog. 2016, 99, 196–203. [Google Scholar] [CrossRef]
- Sun, M.; Dong, J.; Xia, Y.; Shu, R. Antibacterial activities of docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) against planktonic and biofilm growing Streptococcus mutans. Microb. Pathog. 2017, 107, 212–218. [Google Scholar] [CrossRef]
- Jameel, F.; Agarwal, P.; Ahmad, R.; Siddiqui, S.; Serajuddin, M. Synergistic anticancer effects of omega-3 fatty acids (EPA/DHA) and anticancer drug Doxorubicin against human lung adenocarcinoma. Food Biosci. 2024, 61, 104710. [Google Scholar] [CrossRef]
- Bai, J.; Kim, Y.-T.; Ryu, S.; Lee, J.-H. Biocontrol and Rapid Detection of Food-Borne Pathogens Using Bacteriophages and Endolysins. Front. Microbiol. 2016, 7, 474. [Google Scholar] [CrossRef]
- Palafox-Rivera, P.; Tapia-Rodriguez, M.R.; Lopez-Romero, J.C.; Lugo-Flores, M.A.; Quintero-Cabello, K.P.; Silva-Espinoza, B.A.; Cruz-Valenzuela, M.R.; Nazzaro, F.; Ayala-Zavala, J.F. Exploring the potential of hydrolytic enzymes combined with antibacterial agents to disrupt pathogenic biofilms and disinfect released cells. Biofouling 2025, 41, 131–143. [Google Scholar] [CrossRef]
- Alipour-Khezri, E.; Skurnik, M.; Zarrini, G. Pseudomonas aeruginosa Bacteriophages and Their Clinical Applications. Viruses 2024, 16, 1051. [Google Scholar] [CrossRef]
- Ferriol-González, C.; Domingo-Calap, P. Phages for Biofilm Removal. Antibiotics 2020, 9, 268. [Google Scholar] [CrossRef]
- Vrancianu, C.O.; Gheorghe, I.; Dobre, E.-G.; Barbu, I.C.; Cristian, R.E.; Popa, M.; Lee, S.H.; Limban, C.; Vlad, I.M.; Chifiriuc, M.C. Emerging Strategies to Combat β-Lactamase Producing ESKAPE Pathogens. Int. J. Mol. Sci. 2020, 21, 8527. [Google Scholar] [CrossRef]
- Davies, D.T.; Everett, M. Designing Inhibitors of β-Lactamase Enzymes to Overcome Carbapenem Resistance in Gram-Negative Bacteria. Accounts Chem. Res. 2021, 54, 2055–2064. [Google Scholar] [CrossRef] [PubMed]
- Pirlar, R.F.; Halili, N.; Travnik, T.; Trampuz, A.; Karbysheva, S. In vitro activity of cefiderocol against Gram-negative aerobic bacilli in planktonic and biofilm form–alone and in combination with bacteriophages. Sci. Rep. 2025, 15, 17105. [Google Scholar] [CrossRef]
- El-Din, E.M.R.S.; Zaki, B.M.; El-Aziz, A.M.A.; Ali, Y.M. Bacteriophage-Antibiotic Synergy Enhances Therapeutic Efficacy Against Multidrug-Resistant Klebsiella pneumoniae Infections. J. Appl. Microbiol. 2025, 136, lxaf131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-H.; Huang, X.-H.; Wong, W.-L.; Luo, J.-R.; Guo, X.-C.; Liu, W.; Hou, J.; She, M.-T.; Jiang, W.-H.; Sun, N.; et al. A red fluorescent small-molecule for visualization of higher-order cyclic dimeric guanosine monophosphate (c-di-GMP) structure in live bacterial cells and real-time monitoring of biofilm formation on biotic and abiotic surfaces. Sens. Actuators B Chem. 2022, 376, 132992. [Google Scholar] [CrossRef]
- Gou, Y.; Liu, W.; Wang, J.J.; Tan, L.; Hong, B.; Guo, L.; Liu, H.; Pan, Y.; Zhao, Y. CRISPR-Cas9 knockout of qseB induced asynchrony between motility and biofilm formation in Escherichia coli. Can. J. Microbiol. 2019, 65, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Zuberi, A.; Ahmad, N.; Khan, A.U. CRISPRi Induced Suppression of Fimbriae Gene (fimH) of a Uropathogenic Escherichia coli: An Approach to Inhibit Microbial Biofilms. Front. Immunol. 2017, 8, 1552. [Google Scholar] [CrossRef] [PubMed]
- Zuberi, A.; Misba, L.; Khan, A.U. CRISPR Interference (CRISPRi) Inhibition of luxS Gene Expression in E. coli: An Approach to Inhibit Biofilm. Front. Cell. Infect. Microbiol. 2017, 7, 214. [Google Scholar] [CrossRef] [PubMed]
- Hegde, S.; Nilyanimit, P.; Kozlova, E.; Anderson, E.R.; Narra, H.P.; Sahni, S.K.; Heinz, E.; Hughes, G.L.; Bonizzoni, M. CRISPR/Cas9-mediated gene deletion of the ompA gene in symbiotic Cedecea neteri impairs biofilm formation and reduces gut colonization of Aedes aegypti mosquitoes. PLOS Neglected Trop. Dis. 2019, 13, e0007883. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).