
Genomic Insights into Plant Growth-Promoting Traits of Lysinibacillus fusiformis and Bacillus cereus from Rice Fields in Panama

 , , , , ,
, , , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Source and Information of the Microbial Isolates
2.2. Whole-Genome Sequencing (WGS) by Illumina MiSeq and Oxford Nanopore MinION
2.3. Whole-Genome Assembly
2.4. Genome-Based Taxonomic and Phylogenetic Analysis
2.5. Genome Annotation and Identification of Genes Associated with Plant Growth-Promoting Traits
2.6. Identification and Analysis of Biosynthetic Gene Clusters (BGCs)
3. Results and Discussion
3.1. Genomic Assembling Metrics and Annotation of the Isolates
3.2. Taxonomic Annotation
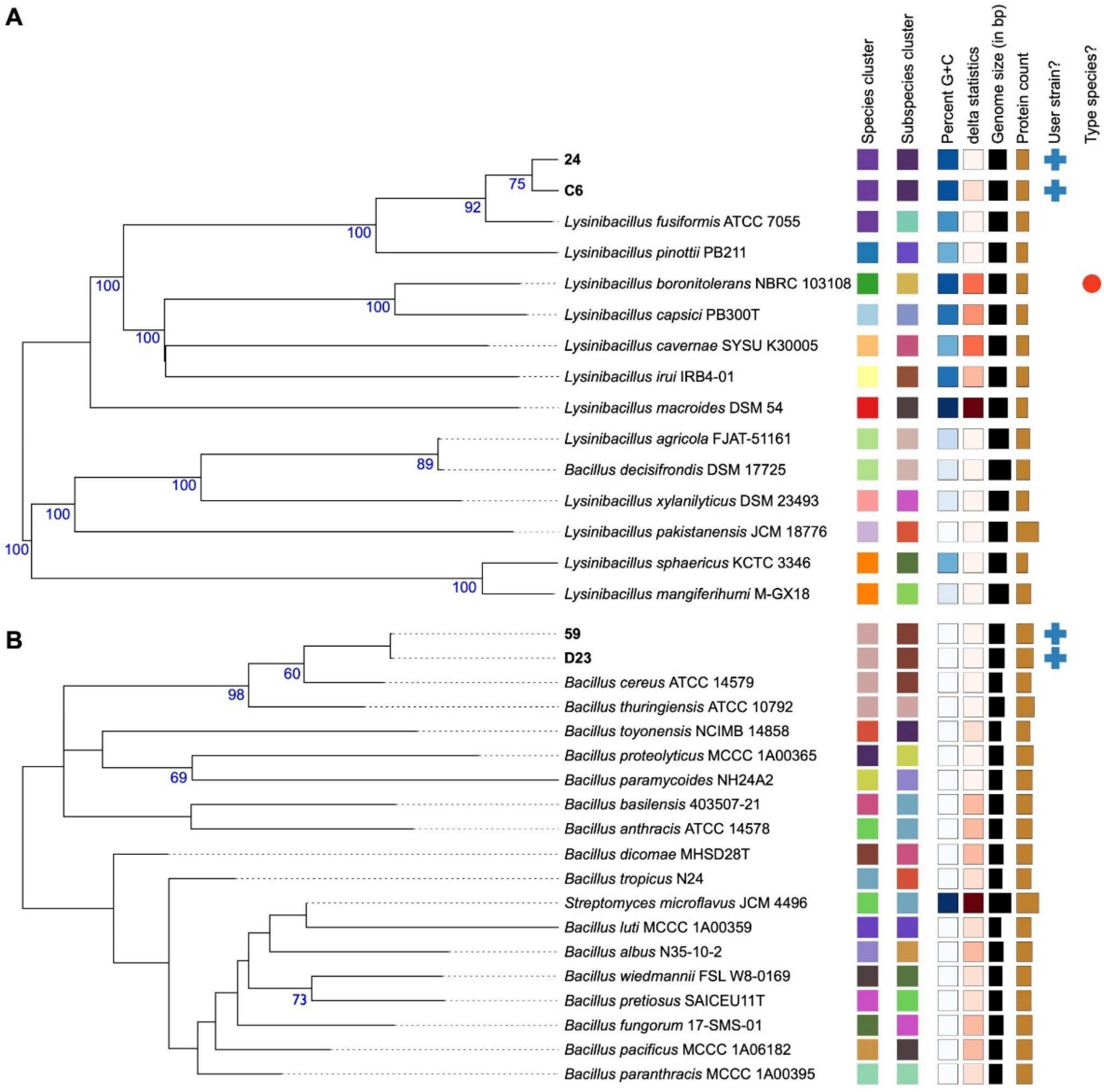
3.3. Phylogenomic Classification, ANI, and dDDH Analysis
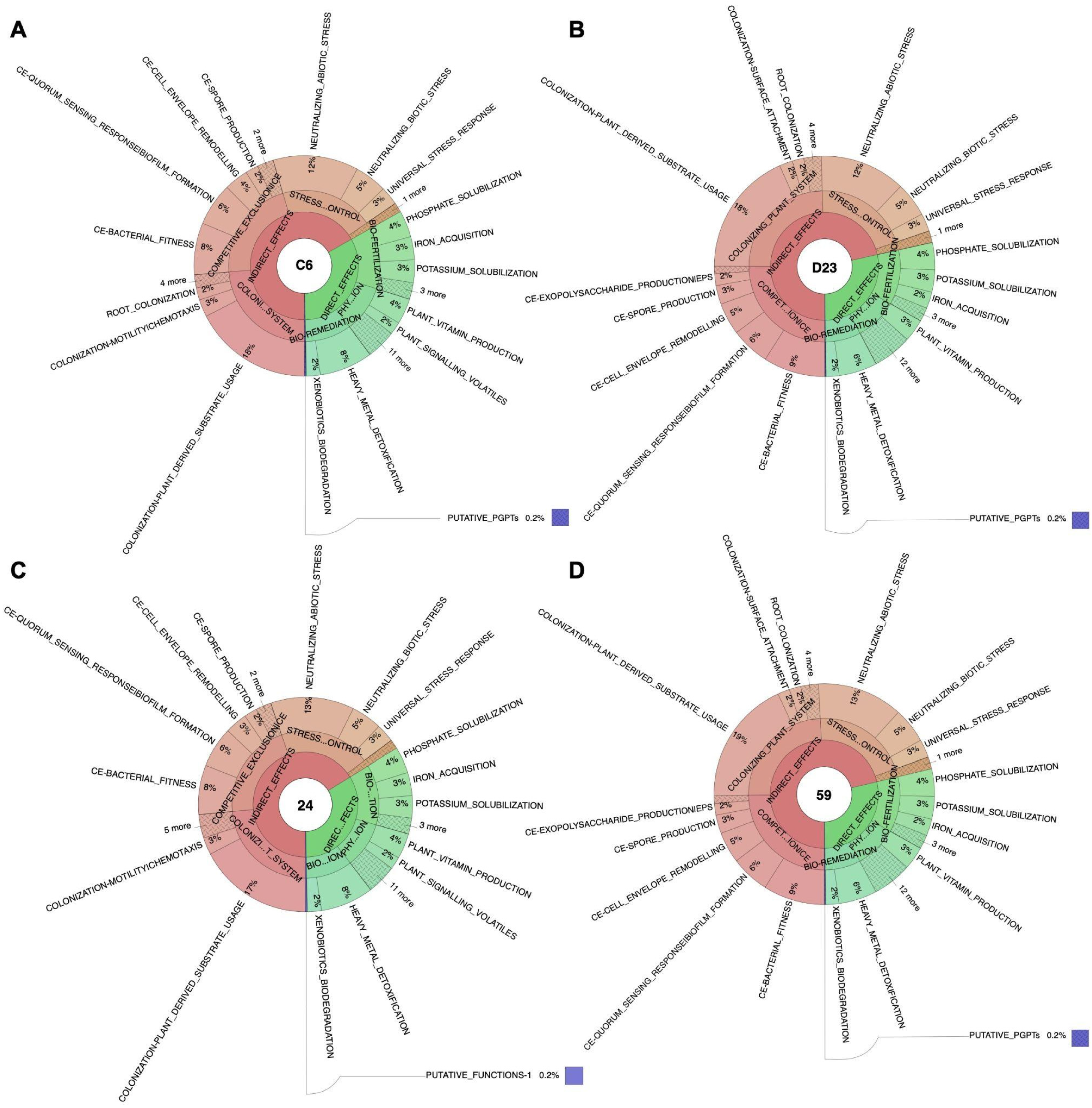
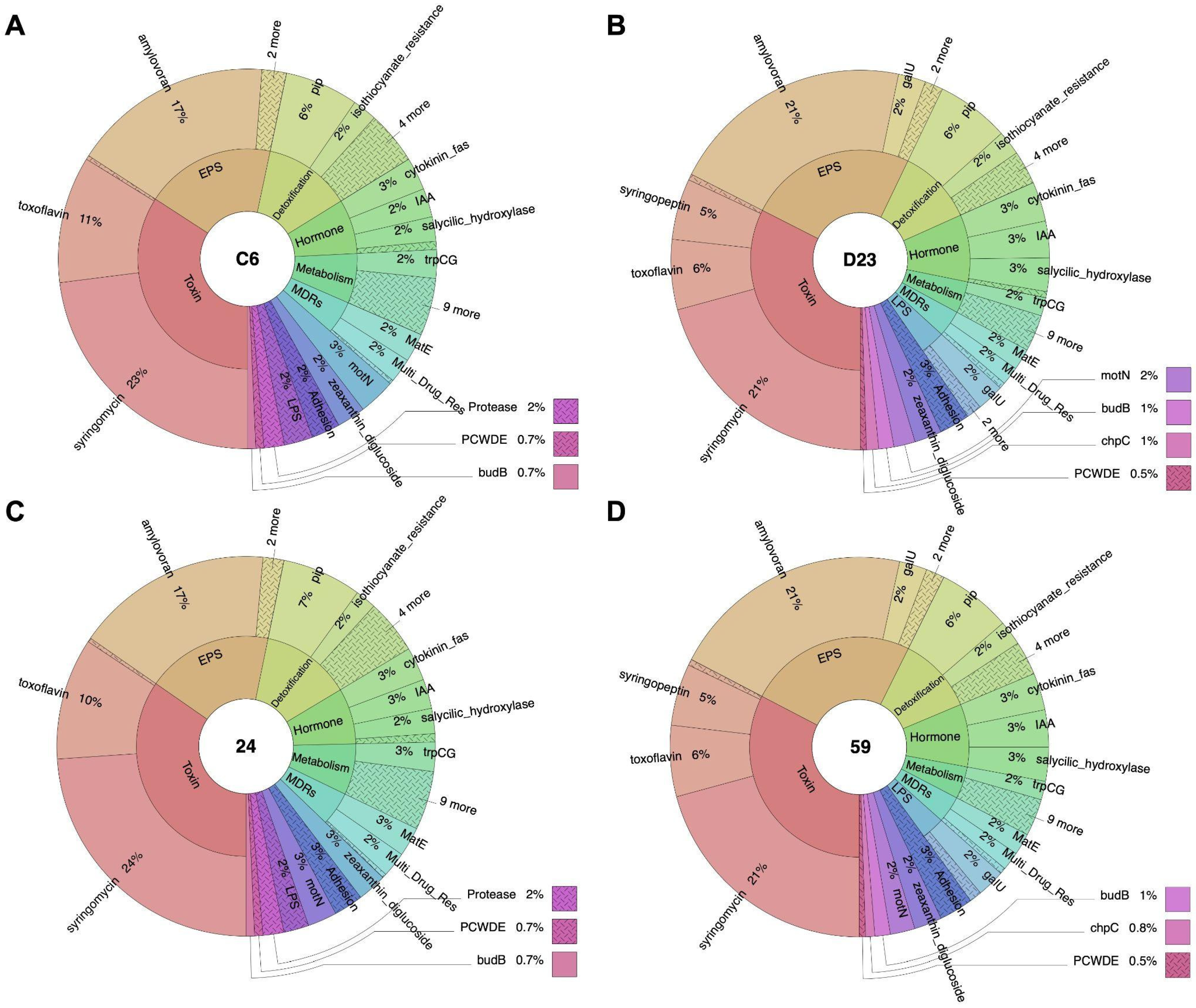
3.4. Identification of Genetic Factors Involved in Plant–Bacteria Interactions
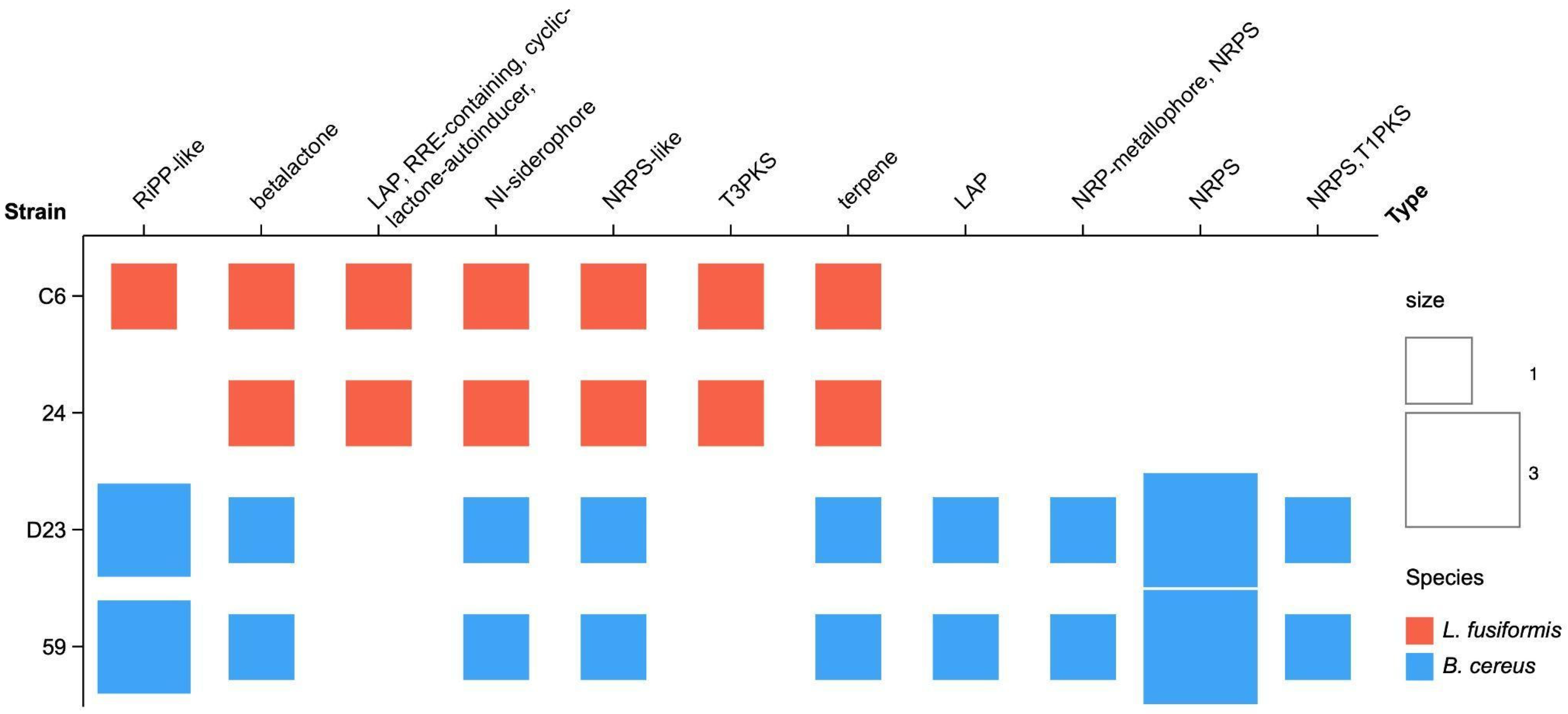
3.5. Genome Mining for BGCs Encoding Bioactive Compounds
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, W.; Zhao, X. Understanding Global Rice Trade Flows: Network Evolution and Implications. Foods 2023, 12, 3298. [Google Scholar] [CrossRef] [PubMed]
- Muthayya, S.; Sugimoto, J.D.; Montgomery, S.; Maberly, G.F. An Overview of Global Rice Production, Supply, Trade, and Consumption. Ann. N. Y. Acad. Sci. 2014, 1324, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, J.; He, Y.; Liu, F.; Feng, X.; Zhang, J. Assessment of the Vigor of Rice Seeds by near-Infrared Hyperspectral Imaging Combined with Transfer Learning. RSC Adv. 2020, 10, 44149–44158. [Google Scholar] [CrossRef]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield Trends Are Insufficient to Double Global Crop Production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef]
- Parry, M.L. Climate Change and World Agriculture; Routledge: London, UK, 2019; ISBN 9781000692778. [Google Scholar]
- Kumari, A.; Lakshmi, G.A.; Krishna, G.K.; Patni, B.; Prakash, S.; Bhattacharyya, M.; Singh, S.K.; Verma, K.K. Climate Change and Its Impact on Crops: A Comprehensive Investigation for Sustainable Agriculture. Agronomy 2022, 12, 3008. [Google Scholar] [CrossRef]
- Shrestha, J.; Kandel, M.; Subedi, S.; Shah, K.K. Role of Nutrients in Rice (Oryza sativa L.): A Review. Agrica 2020, 9, 53. [Google Scholar] [CrossRef]
- Lightfoot, D.A. Developing Crop Varieties with Improved Nutrient-Use Efficiency. In Engineering Nitrogen Utilization in Crop Plants; Shrawat, A., Zayed, A., Lightfoot, D.A., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–11. ISBN 9783319929583. [Google Scholar]
- Thapa, S.; Bhandari, A.; Ghimire, R.; Xue, Q.; Kidwaro, F.; Ghatrehsamani, S.; Maharjan, B.; Goodwin, M. Managing Micronutrients for Improving Soil Fertility, Health, and Soybean Yield. Sustain. Sci. Pract. Policy 2021, 13, 11766. [Google Scholar] [CrossRef]
- Davis, A.G.; Huggins, D.R.; Reganold, J.P. Linking Soil Health and Ecological Resilience to Achieve Agricultural Sustainability. Front. Ecol. Environ. 2023, 21, 131–139. [Google Scholar] [CrossRef]
- Gomiero, T. Soil Degradation, Land Scarcity and Food Security: Reviewing a Complex Challenge. Sustainability 2016, 8, 281. [Google Scholar] [CrossRef]
- Penuelas, J.; Coello, F.; Sardans, J. A Better Use of Fertilizers Is Needed for Global Food Security and Environmental Sustainability. Agric. Food Secur. 2023, 12, 1–9. [Google Scholar] [CrossRef]
- Kremer, R.J. Impacts of Genetically Engineered Crops on the Soil Microbiome, Biological Processes, and Ecosystem Services. In GMOs: Implications for Biodiversity Conservation and Ecological Processes; Chaurasia, A., Hawksworth, D.L., Pessoa de Miranda, M., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 129–147. ISBN 9783030531836. [Google Scholar]
- Tripathi, S.; Srivastava, P.; Devi, R.S.; Bhadouria, R. Chapter 2-Influence of Synthetic Fertilizers and Pesticides on Soil Health and Soil Microbiology. In Agrochemicals Detection, Treatment and Remediation; Prasad, M.N.V., Ed.; Butterworth-Heinemann: Oxford, UK, 2020; pp. 25–54. ISBN 9780081030172. [Google Scholar]
- Poria, V.; Dębiec-Andrzejewska, K.; Fiodor, A.; Lyzohub, M.; Ajijah, N.; Singh, S.; Pranaw, K. Plant Growth-Promoting Bacteria (PGPB) Integrated Phytotechnology: A Sustainable Approach for Remediation of Marginal Lands. Front. Plant Sci. 2022, 13, 999866. [Google Scholar] [CrossRef] [PubMed]
- Tchuisseu Tchakounté, G.V.; Berger, B.; Patz, S.; Fankem, H.; Ruppel, S. Community Structure and Plant Growth-Promoting Potential of Cultivable Bacteria Isolated from Cameroon Soil. Microbiol. Res. 2018, 214, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Abedinzadeh, M.; Etesami, H.; Alikhani, H.A. Characterization of Rhizosphere and Endophytic Bacteria from Roots of Maize (Zea mays L.) Plant Irrigated with Wastewater with Biotechnological Potential in Agriculture. Biotechnol. Rep. Amst. 2019, 21, e00305. [Google Scholar] [CrossRef] [PubMed]
- Zambrano-Mendoza, J.L.; Sangoquiza-Caiza, C.A.; Campaña-Cruz, D.F.; Yánez-Guzmán, C.F. Use of Biofertilizers in Agricultural Production. In Technology in Agriculture; IntechOpen: London, UK, 2021. [Google Scholar]
- Mandic-Mulec, I.; Stefanic, P.; van Elsas, J.D. Ecology of Bacillaceae. Microbiol. Spectr. 2015, 3, TBS-0017-2013. [Google Scholar] [CrossRef]
- Naureen, Z.; Rehman, N.U.; Hussain, H.; Hussain, J.; Gilani, S.A.; Al Housni, S.K.; Mabood, F.; Khan, A.L.; Farooq, S.; Abbas, G.; et al. Exploring the Potentials of Lysinibacillus Sphaericus ZA9 for Plant Growth Promotion and Biocontrol Activities against Phytopathogenic Fungi. Front. Microbiol. 2017, 8, 1477. [Google Scholar] [CrossRef]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef]
- Malik, K.A.; Bilal, R.; Mehnaz, S.; Rasul, G.; Mirza, M.S.; Ali, S. Association of Nitrogen-Fixing, Plant-Growth-Promoting Rhizobacteria (PGPR) with Kallar Grass and Rice. In Opportunities for Biological Nitrogen Fixation in Rice and Other Non-Legumes: Papers presented at the Second Working Group Meeting of the Frontier Project on Nitrogen Fixation in Rice held at the National Institute for Biotechnology and Genetic Engineering (NIBGE), Faisalabad, Pakistan, 13–15 October 1996; Ladha, J.K., de Bruijn, F.J., Malik, K.A., Eds.; Springer: Dordrecht, The Netherlands, 1997; pp. 37–44. ISBN 9789401171137. [Google Scholar]
- Ramakrishna, W.; Yadav, R.; Li, K. Plant Growth Promoting Bacteria in Agriculture: Two Sides of a Coin. Appl. Soil Ecol. 2019, 138, 10–18. [Google Scholar] [CrossRef]
- Ruiu, L. Plant-Growth-Promoting Bacteria (PGPB) against Insects and Other Agricultural Pests. Agronomy 2020, 10, 861. [Google Scholar] [CrossRef]
- Abdelkrim, S.; Abid, G.; Chaieb, O.; Taamalli, W.; Mannai, K.; Louati, F.; Jebara, M.; Jebara, S.H. Plant Growth Promoting Rhizobacteria Modulates the Antioxidant Defense and the Expression of Stress Responsive Genes Providing Pb Accumulation and Tolerance of Grass Pea. Res. Sq. 2022, 30, 10789–10802. [Google Scholar] [CrossRef]
- Tsotetsi, T.; Nephali, L.; Malebe, M.; Tugizimana, F. Bacillus for Plant Growth Promotion and Stress Resilience: What Have We Learned? Plants 2022, 11, 2482. [Google Scholar] [CrossRef]
- Admassie, M.; Wold-Hawariat, Y.; Alemu, T. Characterization and Diversity of Plant Growth Promoting Endophytic and Rhizosphere Bacteria Isolated from theRhizosphere and Tissues of Pepper and Their Effect on Plant Growth Promotion and Disease Suppression of Phytophthora Capsici. Res. Sq. 2021, 1–19. [Google Scholar] [CrossRef]
- Tiwari, S.; Prasad, V.; Chauhan, P.S.; Lata, C. Bacillus Amyloliquefaciens Confers Tolerance to Various Abiotic Stresses and Modulates Plant Response to Phytohormones through Osmoprotection and Gene Expression Regulation in Rice. Front. Plant Sci. 2017, 8, 1510. [Google Scholar] [CrossRef] [PubMed]
- Maitra, S.; Brestic, M.; Bhadra, P.; Shankar, T.; Praharaj, S.; Palai, J.B.; Shah, M.M.R.; Barek, V.; Ondrisik, P.; Skalický, M.; et al. Bioinoculants-Natural Biological Resources for Sustainable Plant Production. Microorganisms 2021, 10, 51. [Google Scholar] [CrossRef] [PubMed]
- del Orozco-Mosqueda, M.C.; Flores, A.; Rojas-Sánchez, B.; Urtis-Flores, C.A.; Morales-Cedeño, L.R.; Valencia-Marin, M.F.; Chávez-Avila, S.; Rojas-Solis, D.; Santoyo, G. Plant Growth-Promoting Bacteria as Bioinoculants: Attributes and Challenges for Sustainable Crop Improvement. Agronomy 2021, 11, 1167. [Google Scholar] [CrossRef]
- Fisberg, R.M.; Fontanelli, M.M.; Kowalskys, I.; Gómez, G.; Rigotti, A.; Cortés, L.Y.; García, M.Y.; Pareja, R.G.; Herrera-Cuenca, M.; Fisberg, M.; et al. Total and Whole Grain Intake in Latin America: Findings from the Multicenter Cross-Sectional Latin American Study of Health and Nutrition (ELANS). Eur. J. Nutr. 2022, 61, 489–501. [Google Scholar] [CrossRef]
- Singh, V.; Zhou, S.; Ganie, Z.; Valverde, B.; Avila, L.; Marchesan, E.; Merotto, A.; Zorrilla, G.; Burgos, N.; Norsworthy, J.; et al. Rice Production in the Americas. In Rice Production Worldwide; Chauhan, B.S., Jabran, K., Mahajan, G., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 137–168. ISBN 9783319475165. [Google Scholar]
- Di Benedetto, N.A.; Corbo, M.R.; Campaniello, D.; Cataldi, M.P.; Bevilacqua, A.; Sinigaglia, M.; Flagella, Z. The Role of Plant Growth Promoting Bacteria in Improving Nitrogen Use Efficiency for Sustainable Crop Production: A Focus on Wheat. AIMS Microbiol. 2017, 3, 413–434. [Google Scholar] [CrossRef]
- Pathania, P.; Rajta, A.; Singh, P.C.; Bhatia, R. Role of Plant Growth-Promoting Bacteria in Sustainable Agriculture. Biocatal. Agric. Biotechnol. 2020, 30, 101842. [Google Scholar] [CrossRef]
- Armada, E.; Leite, M.F.A.; Medina, A.; Azcón, R.; Kuramae, E.E. Native Bacteria Promote Plant Growth under Drought Stress Condition without Impacting the Rhizomicrobiome. FEMS Microbiol. Ecol. 2018, 94, fiy092. [Google Scholar] [CrossRef]
- Aviles, C.F.A.; Acosta, C.B.C.; de los Santos Villalobos, S.; Santoyo, G.; Cota, F.I.P. Characterization of Native Plant Growth-Promoting Bacteria (PGPB) and Their Effect on the Development of Maize (Zea mays L.). Biotecnia 2022, 24, 15–22. [Google Scholar] [CrossRef]
- Özdal, M.; Gür Özdal, Ö.; Sezen, A.; Algur, Ö.F. Biosynthesis of Indole-3-Acetic Acid by Bacillus Cereus Immobilized Cells. Cumhur. Sci. J. 2016, 37, 212. [Google Scholar] [CrossRef]
- Milagres, A.M.; Machuca, A.; Napoleão, D. Detection of Siderophore Production from Several Fungi and Bacteria by a Modification of Chrome Azurol S (CAS) Agar Plate Assay. J. Microbiol. Methods 1999, 37, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.-P. A Simple Double-Layered Chrome Azurol S Agar (SD-CASA) Plate Assay to Optimize the Production of Siderophores by a Potential Biocontrol Agent Bacillus. Afr. J. Microbiol. Res. 2011, 5, 4321–4327. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 12 March 2023).
- De Coster, W.; D’Hert, S.; Schultz, D.T.; Cruts, M.; Van Broeckhoven, C. NanoPack: Visualizing and processing long-read sequencing data. Bioinformatics 2018, 34, 2666–2669. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Wick, R.; Menzel, P. Filtlong: Quality Filtering Tool for Long Reads. 2019. Available online: https://github.com/rrwick/Filtlong (accessed on 10 February 2023).
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the Quality of Microbial Genomes Recovered from Isolates, Single Cells, and Metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef]
- Chaumeil, P.-A.; Mussig, A.J.; Hugenholtz, P.; Parks, D.H. GTDB-Tk: A Toolkit to Classify Genomes with the Genome Taxonomy Database. Bioinformatics 2019, 36, 1925–1927. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. TYGS Is an Automated High-Throughput Platform for State-of-the-Art Genome-Based Taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome Sequence-Based Species Delimitation with Confidence Intervals and Improved Distance Functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef]
- Auch, A.F.; von Jan, M.; Klenk, H.-P.; Göker, M. Digital DNA-DNA Hybridization for Microbial Species Delineation by Means of Genome-to-Genome Sequence Comparison. Stand. Genom. Sci. 2010, 2, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Ouk Kim, Y.; Park, S.-C.; Chun, J. OrthoANI: An Improved Algorithm and Software for Calculating Average Nucleotide Identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Schwengers, O.; Jelonek, L.; Dieckmann, M.A.; Beyvers, S.; Blom, J.; Goesmann, A. Bakta: Rapid and Standardized Annotation of Bacterial Genomes via Alignment-Free Sequence Identification. Microb. Genom. 2021, 7, 000685. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI Prokaryotic Genome Annotation Pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Martínez-García, P.M.; López-Solanilla, E.; Ramos, C.; Rodríguez-Palenzuela, P. Prediction of Bacterial Associations with Plants Using a Supervised Machine-Learning Approach. Environ. Microbiol. 2016, 18, 4847–4861. [Google Scholar] [CrossRef]
- Patz, S.; Gautam, A.; Becker, M.; Ruppel, S.; Rodríguez-Palenzuela, P.; Huson, D.H. PLaBAse: A Comprehensive Web Resource for Analyzing the Plant Growth-Promoting Potential of Plant-Associated Bacteria. bioRxiv 2021, 2021, 12.13.472471. [Google Scholar]
- Medema, M.H.; Blin, K.; Cimermancic, P.; de Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. antiSMASH: Rapid Identification, Annotation and Analysis of Secondary Metabolite Biosynthesis Gene Clusters in Bacterial and Fungal Genome Sequences. Nucleic Acids Res. 2011, 39, W339–W346. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M.; Fetter, A.; Terlouw, B.R.; Metcalf, W.W.; Helfrich, E.J.N.; et al. antiSMASH 7.0: New and Improved Predictions for Detection, Regulation, Chemical Structures and Visualisation. Nucleic Acids Res. 2023, 51, W46–W50. [Google Scholar] [CrossRef]
- Mauri, M.; Elli, T.; Caviglia, G.; Uboldi, G.; Azzi, M. RAWGraphs: A Visualisation Platform to Create Open Outputs. In Proceedings of the 12th Biannual Conference on Italian SIGCHI Chapter, Cagliari, Italy, 18–20 September 2017; Association for Computing Machinery: New York, NY, USA, 2017; pp. 1–5. [Google Scholar]
- Zeng, Q.; Xie, J.; Li, Y.; Gao, T.; Xu, C.; Wang, Q. Comparative Genomic and Functional Analyses of Four Sequenced Bacillus Cereus Genomes Reveal Conservation of Genes Relevant to Plant-Growth-Promoting Traits. Sci. Rep. 2018, 8, 17009. [Google Scholar] [CrossRef]
- Pudova, D.S.; Lutfullin, M.T.; Shagimardanova, E.I.; Hadieva, G.F.; Shigapova, L.; Toymentseva, A.A.; Kabanov, D.A.; Mardanova, A.M.; Vologin, S.G.; Sharipova, M.R. Draft Genome Sequence Data of Lysinibacillus Fusiformis Strain GM, Isolated from Potato Phyllosphere as a Potential Probiotic. Data Brief 2018, 21, 2504–2509. [Google Scholar] [CrossRef]
- Smith, B.R.; Unckless, R.L. Draft Genome Sequence of Lysinibacillus Fusiformis Strain Juneja, a Laboratory-Derived Pathogen of Drosophila Melanogaster. Genome Announc. 2018, 6, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Passera, A.; Rossato, M.; Oliver, J.S.; Battelli, G.; Shahzad, G.-I.-R.; Cosentino, E.; Sage, J.M.; Toffolatti, S.L.; Lopatriello, G.; Davis, J.R.; et al. Characterization of Lysinibacillus Fusiformis Strain S4C11: In Vitro, in Planta, and in Silico Analyses Reveal a Plant-Beneficial Microbe. Microbiol. Res. 2021, 244, 126665. [Google Scholar] [CrossRef] [PubMed]
- Šmarda, P.; Bureš, P.; Horová, L.; Leitch, I.J.; Mucina, L.; Pacini, E.; Tichý, L.; Grulich, V.; Rotreklová, O. Ecological and Evolutionary Significance of Genomic GC Content Diversity in Monocots. Proc. Natl. Acad. Sci. USA 2014, 111, E4096–E4102. [Google Scholar] [CrossRef] [PubMed]
- Ranea, J.A.G.; Buchan, D.W.A.; Thornton, J.M.; Orengo, C.A. Evolution of Protein Superfamilies and Bacterial Genome Size. J. Mol. Biol. 2004, 336, 871–887. [Google Scholar] [CrossRef]
- Mijakovic, I.; Grangeasse, C.; Stülke, J. Regulatory Potential of Post-Translational Modifications in Bacteria; Frontiers Media SA: Lausanne, Switzerland, 2015; ISBN 9782889196104. [Google Scholar]
- Martínez-Cano, D.J.; Reyes-Prieto, M.; Martínez-Romero, E.; Partida-Martínez, L.P.; Latorre, A.; Moya, A.; Delaye, L. Evolution of Small Prokaryotic Genomes. Front. Microbiol. 2014, 5, 742. [Google Scholar] [CrossRef]
- Ondov, B.D.; Treangen, T.J.; Melsted, P.; Mallonee, A.B.; Bergman, N.H.; Koren, S.; Phillippy, A.M. Mash: Fast Genome and Metagenome Distance Estimation Using MinHash. Genome Biol. 2016, 17, 132. [Google Scholar] [CrossRef]
- Vebø, H.C.; Solheim, M.; Snipen, L.; Nes, I.F.; Brede, D.A. Comparative Genomic Analysis of Pathogenic and Probiotic Enterococcus Faecalis Isolates, and Their Transcriptional Responses to Growth in Human Urine. PLoS ONE 2010, 5, e12489. [Google Scholar] [CrossRef]
- Solanki, S.; Devi, D. Analysis of Virulence Genes of Staphylococcus Aureus, Streptococcus. E. coli Isolated from Bovine Subclinical Mastitis. Indian J. Anim. Res. 2022, 1–7. [Google Scholar] [CrossRef]
- Thompson, C.C.; Chimetto, L.; Edwards, R.A.; Swings, J.; Stackebrandt, E.; Thompson, F.L. Microbial Genomic Taxonomy. BMC Genom. 2013, 14, 913. [Google Scholar] [CrossRef]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.-W.; De Meyer, S.; et al. Proposed Minimal Standards for the Use of Genome Data for the Taxonomy of Prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef]
- Van Rossum, T.; Ferretti, P.; Maistrenko, O.M.; Bork, P. Diversity within Species: Interpreting Strains in Microbiomes. Nat. Rev. Microbiol. 2020, 18, 491–506. [Google Scholar] [CrossRef] [PubMed]
- Gat, D.; Mazar, Y.; Cytryn, E.; Rudich, Y. Origin-Dependent Variations in the Atmospheric Microbiome Community in Eastern Mediterranean Dust Storms. Environ. Sci. Technol. 2017, 51, 6709–6718. [Google Scholar] [CrossRef] [PubMed]
- Deng, A.; Wang, T.; Wang, J.; Li, L.; Wang, X.; Liu, L.; Wen, T. Adaptive Mechanisms of Bacillus to near Space Extreme Environments. Sci. Total Environ. 2023, 886, 163952. [Google Scholar] [CrossRef] [PubMed]
- Sebastian, A.M.; Umesh, M.; Priyanka, K.; Preethi, K. Isolation of Plant Growth-Promoting Bacillus Cereus from Soil and Its Use as a Microbial Inoculant. Arab. J. Sci. Eng. 2021, 46, 151–161. [Google Scholar] [CrossRef]
- Kulkova, I.; Dobrzyński, J.; Kowalczyk, P.; Bełżecki, G.; Kramkowski, K. Plant Growth Promotion Using Bacillus Cereus. Int. J. Mol. Sci. 2023, 24, 9759. [Google Scholar] [CrossRef]
- Dotaniya, M.L.; Meena, V.D.; Basak, B.B.; Meena, R.S. Potassium Uptake by Crops as Well as Microorganisms. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Meena, V.S., Maurya, B.R., Verma, J.P., Meena, R.S., Eds.; Springer: New Delhi, India, 2016; pp. 267–280. ISBN 9788132227762. [Google Scholar]
- Jaiswal, D.K.; Verma, J.P.; Prakash, S.; Meena, V.S.; Meena, R.S. Potassium as an Important Plant Nutrient in Sustainable Agriculture: A State of the Art. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Meena, V.S., Maurya, B.R., Verma, J.P., Meena, R.S., Eds.; Springer: New Delhi, India, 2016; pp. 21–29. ISBN 9788132227762. [Google Scholar]
- Jha, Y.; Subramanian, R.B. Regulation of Plant Physiology and Antioxidant Enzymes for Alleviating Salinity Stress by Potassium-Mobilizing Bacteria. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Meena, V.S., Maurya, B.R., Verma, J.P., Meena, R.S., Eds.; Springer: New Delhi, India, 2016; pp. 149–162. ISBN 9788132227762. [Google Scholar]
- Wu, S.C.; Cao, Z.H.; Li, Z.G.; Cheung, K.C.; Wong, M.H. Effects of Biofertilizer Containing N-Fixer, P and K Solubilizers and AM Fungi on Maize Growth: A Greenhouse Trial. Geoderma 2005, 125, 155–166. [Google Scholar] [CrossRef]
- Aguirre-Monroy, A.M.; Santana-Martínez, J.C.; Dussán, J. Lysinibacillus Sphaericus as a Nutrient Enhancer during Fire-Impacted Soil Replantation. Appl. Environ. Soil Sci. 2019, 2019, 3075153. [Google Scholar] [CrossRef]
- Kumar Sahu, P.; Shivaprakash, M.K.; Karaba, N.; Mallesha, B.C.; Subbarayappa, C.T.; Brahmaprakash, G.P. Effect of Bacterial Endophytes Lysinibacillus Sp. on Plant Growth and Fruit Yield of Tomato (Solanum Lycopersicum). Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 3399–3408. [Google Scholar] [CrossRef]
- Martínez, S.A.; Dussán, J. Lysinibacillus Sphaericus Plant Growth Promoter Bacteria and Lead Phytoremediation Enhancer with Canavalia Ensiformis. Environ. Prog. Sustain. Energy 2018, 37, 276–282. [Google Scholar] [CrossRef]
- Sodiq, A.H.; Setiawati, M.R.; Santosa, D.A.; Widayat, D. The Effect of Bio-Fertilizer Applications Bacillus Cereus and Lysinibacillus Sp On Paprika Plants (Capsicum annuum L.) On Plant Nutrient Content and Cultivation Media. IOP Conf. Ser. Earth Environ. Sci. 2022, 978, 012032. [Google Scholar] [CrossRef]
- Gibbons, S.M.; Gilbert, J.A. Microbial Diversity--Exploration of Natural Ecosystems and Microbiomes. Curr. Opin. Genet. Dev. 2015, 35, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Bertola, M.; Mattarozzi, M.; Sanangelantoni, A.M.; Careri, M.; Visioli, G. PGPB Colonizing Three-Year Biochar-Amended Soil: Towards Biochar-Mediated Biofertilization. J. Soil Sci. Plant Nutr. 2019, 19, 841–850. [Google Scholar] [CrossRef]
- Koza, N.A.; Adedayo, A.A.; Babalola, O.O.; Kappo, A.P. Microorganisms in Plant Growth and Development: Roles in Abiotic Stress Tolerance and Secondary Metabolites Secretion. Microorganisms 2022, 10, 1528. [Google Scholar] [CrossRef]
- Dhawi, F.; Hess, A. Poor-Soil Rhizosphere Enriched with Different Microbial Activities Influence the Availability of Base Elements. Open J. Ecol. 2017, 7, 495–502. [Google Scholar] [CrossRef]
- Caniato, M.M.; de Catarino, A.M.; Sousa, T.F.; da Silva, G.F.; Bichara, K.P.d.A.; da Cruz, J.C.; de Assis, L.A.G.; Hanada, R.E. Diversity of Bacterial Strains in Biochar-Enhanced Amazon Soil and Their Potential for Growth Promotion and Biological Disease Control in Tomato. Acta Amazon. 2020, 50, 278–288. [Google Scholar] [CrossRef]
- Rafique, M.; Sultan, T.; Ortas, I.; Chaudhary, H.J. Enhancement of Maize Plant Growth with Inoculation of Phosphate-Solubilizing Bacteria and Biochar Amendment in Soil. Soil Sci. Plant Nutr. 2017, 63, 460–469. [Google Scholar] [CrossRef]
- Chen, W.; Luo, Y. The Effect of Salt-Dissolving Growth-Promoting Rhizobacteria on the Growth of Pepper. Front. Sustain. Dev. 2023, 3, 62–67. [Google Scholar] [CrossRef]
- Hilário, S.; Gonçalves, M.F.M.; Matos, I.; Rangel, L.F.; Sousa, J.A.; Santos, M.J.; Ayra-Pardo, C. Comparative Genomics Reveals Insights into the Potential of Lysinibacillus Irui as a Plant Growth Promoter. Appl. Microbiol. Biotechnol. 2024, 108, 370. [Google Scholar] [CrossRef]
- Ahsan, N.; Shimizu, M. Lysinibacillus Species: Their Potential as Effective Bioremediation, Biostimulant, and Biocontrol Agents. Rev. Agric. Sci. 2021, 9, 103–116. [Google Scholar] [CrossRef]
- Majeed, R.E. Isolation and Diagnosis of Lysinibacillus Fusiformis Obtained from Soil and Its Use as Biofertilizer in Wheat Crop. SABRAO J. Breed. Genet. 2024, 56, 1207–1218. [Google Scholar] [CrossRef]
- Singh, B.; Kaur, J.; Singh, K. Bioremediation of Malathion in Soil by Mixed Bacillus Culture. Adv. Biosci. Biotechnol. 2013, 4, 674–678. [Google Scholar] [CrossRef]
- Singh, B.; Kaur, J.; Singh, K. Biodegradation of Malathion by Brevibacillus Sp. Strain KB2 and Bacillus Cereus Strain PU. World J. Microbiol. Biotechnol. 2012, 28, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Kaur, J.; Singh, K. Transformation of Malathion by Lysinibacillus Sp. Isolated from Soil. Biotechnol. Lett. 2012, 34, 863–867. [Google Scholar] [CrossRef] [PubMed]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of Action of Microbial Biological Control Agents Against Plant Diseases: Relevance Beyond Efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef]
- Madriz-Ordeñana, K.; Pazarlar, S.; Jørgensen, H.J.L.; Nielsen, T.K.; Zhang, Y.; Nielsen, K.L.; Hansen, L.H.; Thordal-Christensen, H. The Bacillus Cereus Strain EC9 Primes the Plant Immune System for Superior Biocontrol of Fusarium Oxysporum. Plants 2022, 11, 687. [Google Scholar] [CrossRef]
- Mishra, A.K.; Baek, K.-H. Salicylic Acid Biosynthesis and Metabolism: A Divergent Pathway for Plants and Bacteria. Biomolecules 2021, 11, 705. [Google Scholar] [CrossRef]
- Mayer, E.; Dörr de Quadros, P.; Fulthorpe, R. Plantibacter Flavus, Curtobacterium Herbarum, Paenibacillus Taichungensis, and Rhizobium Selenitireducens Endophytes Provide Host-Specific Growth Promotion of Arabidopsis Thaliana, Basil, Lettuce, and Bok Choy Plants. Appl. Environ. Microbiol. 2019, 85, e00383-19. [Google Scholar] [CrossRef]
- Zhang, D.; Xu, H.; Gao, J.; Portieles, R.; Du, L.; Gao, X.; Borroto Nordelo, C.; Borrás-Hidalgo, O. Endophytic Bacillus Altitudinis Strain Uses Different Novelty Molecular Pathways to Enhance Plant Growth. Front. Microbiol. 2021, 12, 692313. [Google Scholar] [CrossRef]
- Heydarian, Z.; Yu, M.; Gruber, M.; Glick, B.R.; Zhou, R.; Hegedus, D.D. Inoculation of Soil with Plant Growth Promoting Bacteria Producing 1-Aminocyclopropane-1-Carboxylate Deaminase or Expression of the Corresponding acdS Gene in Transgenic Plants Increases Salinity Tolerance in Camelina Sativa. Front. Microbiol. 2016, 7, 1966. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Hernandez, A.G.; Glick, B.R.; Rossi, M.J. The Extreme Plant-Growth-Promoting Properties of Pantoea Phytobeneficialis MSR2 Revealed by Functional and Genomic Analysis. Environ. Microbiol. 2020, 22, 1341–1355. [Google Scholar] [CrossRef]
- Leite, M.; Pereira, A.; Souza, A.; Andrade, P.; Barbosa, M.; Andreote, F.; Freire, F.; Sobral, J. Potentially Diazotrophic Endophytic Bacteria Associated to Sugarcane Are Effective in Plant Growth-Promotion. J. Exp. Agric. Int. 2018, 21, 1–15. [Google Scholar] [CrossRef]
- Thomas, J.; Kim, H.R.; Rahmatallah, Y.; Wiggins, G.; Yang, Q.; Singh, R.; Glazko, G.; Mukherjee, A. RNA-Seq Reveals Differentially Expressed Genes in Rice (Oryza sativa) Roots during Interactions with Plant-Growth Promoting Bacteria, Azospirillum Brasilense. PLoS ONE 2019, 14, e0217309. [Google Scholar] [CrossRef] [PubMed]
- Wessler, S.; Schneider, G.; Backert, S. Bacterial Serine Protease HtrA as a Promising New Target for Antimicrobial Therapy? Cell Commun. Signal. 2017, 15, 4. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.H.; Bolken, T.C.; Jones, K.F.; Zeller, G.O.; Hruby, D.E. Conserved DegP Protease in Gram-Positive Bacteria Is Essential for Thermal and Oxidative Tolerance and Full Virulence in Streptococcus Pyogenes. Infect. Immun. 2001, 69, 5538–5545. [Google Scholar] [CrossRef]
- Chao, Y.; Bergenfelz, C.; Sun, R.; Han, X.; Achour, A.; Hakansson, A.P. The Serine Protease HtrA Plays a Key Role in Heat-Induced Dispersal of Pneumococcal Biofilms. Sci. Rep. 2020, 10, 22455. [Google Scholar] [CrossRef]
- Ren, H.; Liu, J. AsnB Is Involved in Natural Resistance of Mycobacterium Smegmatis to Multiple Drugs. Antimicrob. Agents Chemother. 2006, 50, 250–255. [Google Scholar] [CrossRef]
- Qian, G.; Liu, C.; Wu, G.; Yin, F.; Zhao, Y.; Zhou, Y.; Zhang, Y.; Song, Z.; Fan, J.; Hu, B.; et al. AsnB, Regulated by Diffusible Signal Factor and Global Regulator Clp, Is Involved in Aspartate Metabolism, Resistance to Oxidative Stress and Virulence in Xanthomonas Oryzae Pv. Oryzicola. Mol. Plant Pathol. 2013, 14, 145–157. [Google Scholar] [CrossRef]
- Rogers, E.W.; Dalisay, D.S.; Molinski, T.F. (+)-Zwittermicin A: Assignment of Its Complete Configuration by Total Synthesis of the Enantiomer and Implication of D-Serine in Its Biosynthesis. Angew. Chem. Int. Ed Engl. 2008, 47, 8086–8089. [Google Scholar] [CrossRef]
- Broderick, N.A.; Goodman, R.M.; Handelsman, J.; Raffa, K.F. Effect of Host Diet and Insect Source on Synergy of Gypsy Moth (Lepidoptera: Lymantriidae) Mortality to Bacillus Thuringiensis Subsp. Kurstaki by Zwittermicin A. Environ. Entomol. 2003, 32, 387–391. [Google Scholar] [CrossRef]
- Yin, Q.-J.; Ying, T.-T.; Zhou, Z.-Y.; Hu, G.-A.; Yang, C.-L.; Hua, Y.; Wang, H.; Wei, B. Species-Specificity of the Secondary Biosynthetic Potential in Bacillus. Front. Microbiol. 2023, 14, 1271418. [Google Scholar] [CrossRef]
- Jähne, J.; Le Thi, T.T.; Blumenscheit, C.; Schneider, A.; Pham, T.L.; Le Thi, P.T.; Blom, J.; Vater, J.; Schweder, T.; Lasch, P.; et al. Novel Plant-Associated Brevibacillus and Lysinibacillus Genomospecies Harbor a Rich Biosynthetic Potential of Antimicrobial Compounds. Microorganisms 2023, 11, 168. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Wang, C.; Song, X.; Ding, X.; Wu, L.; Wu, H.; Gao, X.; Borriss, R. Bacillus Velezensis FZB42 in 2018: The Gram-Positive Model Strain for Plant Growth Promotion and Biocontrol. Front. Microbiol. 2018, 9, 2491. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, I.D.A.; Bach, E.; da Silva Moreira, F.; Müller, A.R.; Rangel, C.P.; Wilhelm, C.M.; Barth, A.L.; Passaglia, L.M.P. Antifungal Potential against Sclerotinia Sclerotiorum (Lib.) de Bary and Plant Growth Promoting Abilities of Bacillus Isolates from Canola (Brassica napus L.) Roots. Microbiol. Res. 2021, 248, 126754. [Google Scholar] [CrossRef]
- van Santen, J.A.; Poynton, E.F.; Iskakova, D.; McMann, E.; Alsup, T.A.; Clark, T.N.; Fergusson, C.H.; Fewer, D.P.; Hughes, A.H.; McCadden, C.A.; et al. The Natural Products Atlas 2.0: A Database of Microbially-Derived Natural Products. Nucleic Acids Res. 2022, 50, D1317–D1323. [Google Scholar] [CrossRef]
- Devi, S.; Sharma, S.; Tiwari, A.; Bhatt, A.K.; Singh, N.K.; Singh, M.; Kaushalendra; Kumar, A. Screening for Multifarious Plant Growth Promoting and Biocontrol Attributes in Bacillus Strains Isolated from Indo Gangetic Soil for Enhancing Growth of Rice Crops. Microorganisms 2023, 11, 1085. [Google Scholar] [CrossRef]
- Bhattacharyya, C.; Banerjee, S.; Acharya, U.; Mitra, A.; Mallick, I.; Haldar, A.; Haldar, S.; Ghosh, A.; Ghosh, A. Evaluation of Plant Growth Promotion Properties and Induction of Antioxidative Defense Mechanism by Tea Rhizobacteria of Darjeeling, India. Sci. Rep. 2020, 10, 15536. [Google Scholar] [CrossRef]
- Shabanamol, S.; Divya, K.; George, T.K.; Rishad, K.S.; Sreekumar, T.S.; Jisha, M.S. Characterization and in Planta Nitrogen Fixation of Plant Growth Promoting Endophytic Diazotrophic Lysinibacillus Sphaericus Isolated from Rice (Oryza sativa). Physiol. Mol. Plant Pathol. 2018, 102, 46–54. [Google Scholar] [CrossRef]
- Ali, Q.; Ayaz, M.; Mu, G.; Hussain, A.; Yuanyuan, Q.; Yu, C.; Xu, Y.; Manghwar, H.; Gu, Q.; Wu, H.; et al. Revealing Plant Growth-Promoting Mechanisms of Bacillus Strains in Elevating Rice Growth and Its Interaction with Salt Stress. Front. Plant Sci. 2022, 13, 994902. [Google Scholar] [CrossRef]
- Lee, S.; Kim, J.-A.; Song, J.; Choe, S.; Jang, G.; Kim, Y. Plant Growth-Promoting Rhizobacterium Bacillus Megaterium Modulates the Expression of Antioxidant-Related and Drought-Responsive Genes to Protect Rice (Oryza sativa L.) from Drought. Front. Microbiol. 2024, 15, 1430546. [Google Scholar] [CrossRef]
- Logan, N.A. Bacillus and Relatives in Foodborne Illness. J. Appl. Microbiol. 2012, 112, 417–429. [Google Scholar] [CrossRef]
- Adeleke, B.; Ayangbenro, A.; Babalola, O. Genomic Analysis of Endophytic Bacillus Cereus T4S and Its Plant Growth-Promoting Traits. Plants 2021, 10, 1776. [Google Scholar] [CrossRef] [PubMed]
- Kotiranta, A.; Lounatmaa, K.; Haapasalo, M. Epidemiology and Pathogenesis of Bacillus Cereus Infections. Microbes Infect. 2000, 2, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Märtlbauer, E.; Dietrich, R.; Luo, H.; Ding, S.; Zhu, K. Multifaceted Toxin Profile, an Approach toward a Better Understanding of probiotic Bacillus Cereus. Crit. Rev. Toxicol. 2019, 49, 342–356. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | C6 | D23 | 24 | 59 |
|---|---|---|---|---|
| Total size (bp) | 4,758,247 | 6,199,443 | 4,682,368 | 6,188,954 |
| G+C ratio (%) | 37.62 | 34.84 | 37.65 | 34.87 |
| No. of contigs | 11 | 20 | 5 | 19 |
| Completeness (%) | 99.34 | 99.17 | 99.34 | 99.01 |
| Contamination (%) | 0.33 | 1.98 | 1.32 | 0.62 |
| Predicted genes | 4812 | 6296 | 4680 | 6290 |
| CDSs (total) | 4662 | 6143 | 4532 | 6139 |
| CDSs (with protein) | 4614 | 5820 | 4489 | 5750 |
| tRNA genes | 108 | 109 | 106 | 104 |
| rRNA genes | 37 | 39 | 37 | 42 |
| Isolate ID | Species | ANI | Type Strain | dDDH (%) [CI] |
|---|---|---|---|---|
| C6 | Lysinibacillus fusiformis | 97.30 | Lysinibacillus fusiformis ATCC 7055 | 76.6 [73.6–79.3] |
| D23 | Bacillus cereus | 97.71 | Bacillus cereus ATCC 14579 | 79.3 [76.4–82.0] |
| 24 | Lysinibacillus fusiformis | 97.33 | Lysinibacillus fusiformis ATCC 7055 | 76.5 [73.5–79.3] |
| 59 | Bacillus cereus | 97.75 | Bacillus cereus ATCC 14579 | 79.3 [76.3–81.9] |
| Strain | Region/Contig | Type | Location | Most Similar Known Cluster | Biosynthetic Class | Similarity (%) |
|---|---|---|---|---|---|---|
| Lysinibacillus fusiformis strain C6 | 1.1 | Terpene | 230,739–251,560 | — | — | — |
| 1.2 | NI-siderophore | 964,677–996,223 | petrobactin | Other | 33 | |
| 1.3 | LAP, RRE-containing, cyclic-lactone-autoinducer, thiopeptide | 1,909,518–1,937,160 | — | — | — | |
| 1.4 | NRPS-like | 2,710,419–2,753,586 | kijanimicin | Polyketide | 4 | |
| 1.5 | T3PKS | 2,891,113–2,932,195 | bacillibactin/bacillibactin E/bacillibactin F | NRP | 30 | |
| 1.6 | RiPP-like | 3,016,181–3,026,375 | — | — | — | |
| 1.7 | betalactone | 3,667,608–3,691,832 | fengycin | NRP | 46 | |
| Lysinibacillus fusiformis strain 24 | 1.1 | NI-siderophore | 361,784–393,330 | petrobactin | Other | 33 |
| 1.2 | Terpene | 1,049,941–1,070,762 | — | — | — | |
| 1.3 | betalactone | 2,259,332–2,283,556 | fengycin | NRP | 46 | |
| 1.4 | T3PKS | 3,019,730–3,060,812 | bacillibactin/bacillibactin E/bacillibactin F | NRP | 30 | |
| 1.5 | NRPS-like | 3,198,178–3,241,345 | kijanimicin | Polyketide | 4 | |
| 1.6 | LAP, RRE-containing, cyclic-lactone-autoinducer, thiopeptide | 4,038,807–4,066,450 | — | — | — | |
| Bacillus cereus strain D23 | 1.1 | NI-siderophore | 659,342–691,049 | petrobactin | Other | 100 |
| 1.2 | NRP-metallophore, NRPS | 974,014–1,025,762 | bacillibactin | NRP | 85 | |
| 1.3 | NRPS | 1,115,705–1,181,616 | — | — | — | |
| 1.4 | betalactone | 1,207,713–1,232,951 | fengycin | NRP | 40 | |
| 1.5 | RiPP-like | 1,285,369–1,295,698 | — | — | — | |
| 1.6 | RiPP-like | 1,392,772–1,403,038 | — | — | — | |
| 1.7 | NRPS | 1,419,276–1,466,292 | — | — | — | |
| 1.8 | Terpene | 2,252,959–2,274,812 | molybdenum cofactor | Other | 17 | |
| 2.1 | NRPS-like | 114,857–158,438 | — | — | — | |
| 2.2 | LAP | 957,913–977,828 | — | — | — | |
| 3.1 | NRPS, T1PKS | 7476–20,292 | zwittermicin A | NRP+Polyketide | 100 | |
| 3.2 | NRPS | 349,886–405,034 | — | — | — | |
| Bacillus cereus strain 59 | 1.1 | LAP | 75,694–99,200 | — | — | — |
| 1.2 | NI-siderophore | 755,354–787,061 | petrobactin | Other | 100 | |
| 1.3 | NRP-metallophore, NRPS | 1,069,984–1,121,732 | bacillibactin | NRP | 85 | |
| 1.4 | NRPS | 1,211,677–1,277,588 | bogorol A | NRP | 11 | |
| 1.5 | betalactone | 1,303,719–1,328,957 | fengycin | NRP | 40 | |
| 1.6 | RiPP-like | 1,381,111–1,391,440 | — | — | — | |
| 1.7 | RiPP-like | 1,488,512–1,498,778 | — | — | — | |
| 1.8 | NRPS | 1,515,017–1,562,033 | — | — | — | |
| 1.9 | Terpene | 2,348,738–2,370,591 | molybdenum cofactor | Other | 17 | |
| 1.10 | NRPS-like | 4,638,196–4,681,777 | — | — | — | |
| 2.1 | NRPS | 77,842–129,420 | puwainaphycin F/minutissamide A/minutissamide B/minutissamide C/minutissamide D | NRP | 30 | |
| 2.2 | NRPS, T1PKS | 367,039–458,465 | zwittermicin A | NRP+Polyketide | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguilar, C.; Herrera, R.; Causadías, J.L.; Bernal, B.; Chavarria, O.; González, C.; Gondola, J.; Moreno, A.; Martínez, A.A. Genomic Insights into Plant Growth-Promoting Traits of Lysinibacillus fusiformis and Bacillus cereus from Rice Fields in Panama. Microbiol. Res. 2025, 16, 95. https://doi.org/10.3390/microbiolres16050095
Aguilar C, Herrera R, Causadías JL, Bernal B, Chavarria O, González C, Gondola J, Moreno A, Martínez AA. Genomic Insights into Plant Growth-Promoting Traits of Lysinibacillus fusiformis and Bacillus cereus from Rice Fields in Panama. Microbiology Research. 2025; 16(5):95. https://doi.org/10.3390/microbiolres16050095
Chicago/Turabian StyleAguilar, Celestino, Rito Herrera, José L. Causadías, Betzaida Bernal, Oris Chavarria, Claudia González, Jessica Gondola, Ambar Moreno, and Alexander A. Martínez. 2025. "Genomic Insights into Plant Growth-Promoting Traits of Lysinibacillus fusiformis and Bacillus cereus from Rice Fields in Panama" Microbiology Research 16, no. 5: 95. https://doi.org/10.3390/microbiolres16050095
APA StyleAguilar, C., Herrera, R., Causadías, J. L., Bernal, B., Chavarria, O., González, C., Gondola, J., Moreno, A., & Martínez, A. A. (2025). Genomic Insights into Plant Growth-Promoting Traits of Lysinibacillus fusiformis and Bacillus cereus from Rice Fields in Panama. Microbiology Research, 16(5), 95. https://doi.org/10.3390/microbiolres16050095