
Genetic Diversity of Plasmodium vivax Surface Ookinete Protein Pvs25 and Host Genes in Individuals Living along the Thai–Myanmar Border and Their Relationships with Parasite Density


Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Study Area and Sample Processing
2.3. Polymorphism Host Genotyping
2.4. Statistical Analysis
2.5. Sequencing and Bioinformatic Analysis of Pvs25 Sequences
2.6. Construction of the Global Database of Pvs25
3. Results
3.1. Study Population and Demographic Data
3.2. Genotype and Allele Distribution of the Host Genes
3.3. Associations between Host Genotype Variants and Parasite Density
3.4. Genetic Diversity of Pvs25
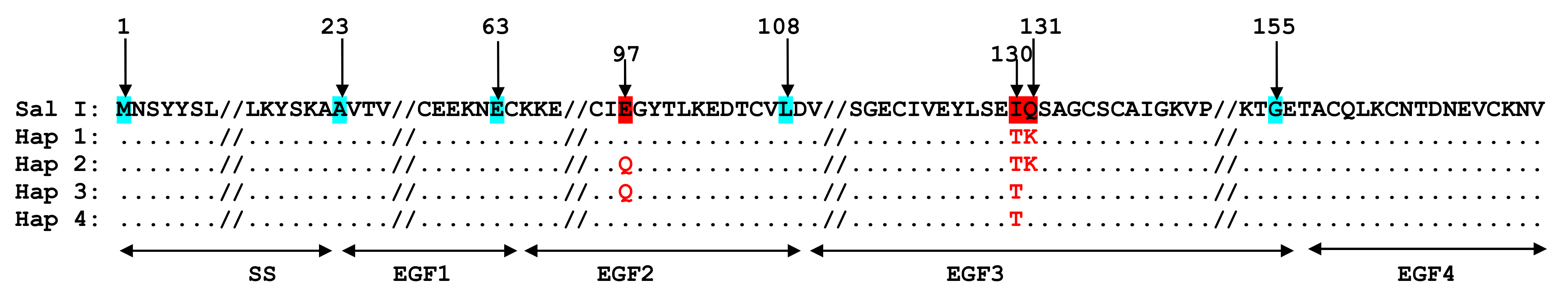
3.4.1. Polymorphism of Pvs25
3.4.2. Pvs25 Haplotypes
3.4.3. Nucleotide Diversity and Natural Selection of the Pvs25 Gene
3.4.4. Comparison of Amino Acid Polymorphisms in Thai Pvs25 and the Global Pvs25
4. Discussion
4.1. Genetic Diversity of Host Genes
4.1.1. TLRs and Adaptor Proteins
4.1.2. Parasite-Binding Genes
4.2. Genetic Diversity of Pv
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Omar, A.H.; Yasunami, M.; Yamazaki, A.; Shibata, H.; Ofori, M.F.; Akanmori, B.D.; Shuaibu, M.N.; Kikuchi, M.; Hirayama, K. Toll-like receptor 9 (TLR9) polymorphism associated with symptomatic malaria: A cohort study. Malar. J. 2012, 11, 168. [Google Scholar] [CrossRef] [PubMed]
- Jalei, A.A.; Chaijaroenkul, W.; Na-Bangchang, K. Plasmodium falciparum drug resistance gene status in the Horn of Africa: A systematic review. Afr. J. Pharm. Pharmacol. 2018, 12, 361–373. [Google Scholar]
- Payne, R.O.; Silk, S.E.; Elias, S.C.; Milne, K.H.; Rawlinson, T.A.; Llewellyn, D.; Shakri, A.R.; Jin, J.; Labbé, G.M.; Edwards, N.J. Human vaccination against Plasmodium vivax Duffy-binding protein induces strain-transcending antibodies. JCI Insight 2017, 2, e93683. [Google Scholar] [CrossRef]
- Wu, Y.; Ellis, R.D.; Shaffer, D.; Fontes, E.; Malkin, E.M.; Mahanty, S.; Fay, M.P.; Narum, D.; Rausch, K.; Miles, A.P. Phase 1 trial of malaria transmission blocking vaccine candidates Pfs25 and Pvs25 formulated with montanide ISA 51. PLoS ONE 2008, 3, e2636. [Google Scholar] [CrossRef] [PubMed]
- Dhangadamajhi, G.; Kar, S.K.; Ranjit, M. The survival strategies of malaria parasite in the red blood cell and host cell polymorphisms. Malar. Res. Treat. 2010, 2010, 973094. [Google Scholar] [CrossRef]
- Ramirez, A.D.R.; de Jesus, M.C.S.; Rossit, J.; Reis, N.F.; Santos-Filho, M.C.; Sudré, A.P.; de Oliveira-Ferreira, J.; de Souza Baptista, A.R.; Storti-Melo, L.M.; Machado, R.L.D. Association of toll-like receptors in malaria susceptibility and immunopathogenesis: A meta-analysis. Heliyon 2022, 8, e09318. [Google Scholar] [CrossRef]
- Antonelli, L.R.; Junqueira, C.; Vinetz, J.M.; Golenbock, D.T.; Ferreira, M.U.; Gazzinelli, R.T. The immunology of Plasmodium vivax malaria. Immunol. Rev. 2020, 293, 163–189. [Google Scholar] [CrossRef]
- Sharma, S.; DeOliveira, R.B.; Kalantari, P.; Parroche, P.; Goutagny, N.; Jiang, Z.; Chan, J.; Bartholomeu, D.C.; Lauw, F.; Hall, J.P. Innate immune recognition of an AT-rich stem-loop DNA motif in the Plasmodium falciparum genome. Immunity 2011, 35, 194–207. [Google Scholar] [CrossRef]
- Junqueira, C.; Barbosa, C.R.; Costa, P.A.; Teixeira-Carvalho, A.; Castro, G.; Sen Santara, S.; Barbosa, R.P.; Dotiwala, F.; Pereira, D.B.; Antonelli, L.R. Cytotoxic CD8+ T cells recognize and kill Plasmodium vivax–infected reticulocytes. Nat. Med. 2018, 24, 1330–1336. [Google Scholar] [CrossRef]
- Bach, F.A.; Sandoval, D.M.; Mazurczyk, M.; Themistocleous, Y.; Rawlinson, T.A.; Harding, A.C.; Kemp, A.; Silk, S.E.; Barrett, J.R.; Edwards, N.J. A systematic analysis of the human immune response to Plasmodium vivax. J. Clin. Investig. 2023, 133, e152463. [Google Scholar] [CrossRef]
- Kariuki, S.N.; Williams, T.N. Human genetics and malaria resistance. Hum. Genet. 2020, 139, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Leoratti, F.M.; Farias, L.; Alves, F.P.; Suarez-Mútis, M.C.; Coura, J.R.; Kalil, J.; Camargo, E.P.; Moraes, S.L.; Ramasawmy, R. Variants in the Toll-like Receptor Signaling Pathway and Clinical Outcomes of Malaria. J. Infect. Dis. 2008, 198, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Capelluto, D.G. Tollip: A multitasking protein in innate immunity and protein trafficking. Microbes Infect. 2012, 14, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Brasil, L.W.; Barbosa, L.R.; de Araujo, F.J.; da Costa, A.G.; da Silva, L.; Pinheiro, S.K.; de Almeida, A.C.; Kuhn, A.; Vitor-Silva, S.; de Melo, G.C. TOLLIP gene variant is associated with Plasmodium vivax malaria in the Brazilian Amazon. Malar. J. 2017, 16, 116. [Google Scholar] [CrossRef]
- Costa, A.G.; Ramasawmy, R.; Ibiapina, H.N.S.; Sampaio, V.S.; Xábregas, L.A.; Brasil, L.W.; Tarragô, A.M.; Almeida, A.C.G.; Kuehn, A.; Vitor-Silva, S. Association of TLR variants with susceptibility to Plasmodium vivax malaria and parasitemia in the Amazon region of Brazil. PLoS ONE 2017, 12, e0183840. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, L.O.; Fernandes, F.; Monteiro, E.F.; Curado, I.; Holcman, M.M.; Wunderlich, G.; Santos, S.E.; Soler, J.M.; Kirchgatter, K. Influence of polymorphisms in toll-like receptors (TLRs) on malaria susceptibility in low-endemic area of the Atlantic Forest, São Paulo, Brazil. Acta Trop 2018, 182, 309–316. [Google Scholar] [CrossRef] [PubMed]
- De Mendonça, V.R.; Goncalves, M.S.; Barral-Netto, M. The host genetic diversity in malaria infection. J. Trop. Med. 2012, 2012, 940616. [Google Scholar] [CrossRef]
- Kaur, H.; Sehgal, R.; Kumar, A.; Sehgal, A.; Bharti, P.K.; Bansal, D.; Mohapatra, P.K.; Mahanta, J.; Sultan, A.A. Exploration of genetic diversity of Plasmodium vivax circumsporozoite protein (Pvcsp) and Plasmodium vivax sexual stage antigen (Pvs25) among North Indian isolates. Malar. J. 2019, 18, 308. [Google Scholar] [CrossRef]
- Sattabongkot, J.; Tsuboi, T.; Hisaeda, H.; Tachibana, M.; Suwanabun, N.; Rungruang, T.; Cao, Y.-M.; Stowers, A.W.; Sirichaisinthop, J.; Coleman, R.E. Blocking of transmission to mosquitoes by antibody to Plasmodium vivax malaria vaccine candidates Pvs25 and Pvs28 despite antigenic polymorphism in field isolates. Am. J. Trop. Med. Hyg. 2003, 69, 536–541. [Google Scholar] [CrossRef]
- Lê, H.G.; Kang, J.-M.; Jun, H.; Lee, J.; Moe, M.; Thái, T.L.; Lin, K.; Myint, M.K.; Yoo, W.G.; Sohn, W.-M. Genetic diversity and natural selection of transmission-blocking vaccine candidate antigens Pvs25 and Pvs28 in Plasmodium vivax Myanmar isolates. Acta Trop. 2019, 198, 105104. [Google Scholar] [CrossRef]
- Wang, S.; Tian, P.; Li, S.; Liu, H.; Guo, X.; Huang, F. Genetic diversity of transmission-blocking vaccine candidate antigens Pvs25 and Pvs28 in Plasmodium vivax isolates from China. BMC Infect. Dis. 2022, 22, 944. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Zheng, L.; Zhu, X.; Wang, G.; Pan, Y.; Li, Y.; Yang, Y.; Lin, Y.; Cui, L.; Cao, Y. Genetic diversity of transmission-blocking vaccine candidates Pvs25 and Pvs28 in Plasmodium vivax isolates from Yunnan Province, China. Parasit. Vectors 2011, 4, 224. [Google Scholar] [CrossRef]
- Han, E.T.; Lee, W.J.; Sattabongkot, J.; Jang, J.W.; Nam, M.H.; An, S.S.A.; Suh, I.; Lim, C.S. Sequence polymorphisms of Plasmodium vivax ookinete surface proteins (Pvs25 and Pvs28) from clinical isolates in Korea. Trop. Med. Int. Health 2010, 15, 1072–1076. [Google Scholar] [CrossRef]
- Kang, J.-M.; Ju, H.-L.; Moon, S.-U.; Cho, P.-Y.; Bahk, Y.-Y.; Sohn, W.-M.; Park, Y.-K.; Cha, S.H.; Kim, T.-S.; Na, B.-K. Limited sequence polymorphisms of four transmission-blocking vaccine candidate antigens in Plasmodium vivax Korean isolates. Malar. J. 2013, 12, 144. [Google Scholar] [CrossRef] [PubMed]
- Prajapati, S.K.; Joshi, H.; Dua, V.K. Antigenic repertoire of Plasmodium vivax transmission-blocking vaccine candidates from the Indian subcontinent. Malar. J. 2011, 10, 111. [Google Scholar] [CrossRef] [PubMed]
- Zakeri, S.; Razavi, S.; Djadid, N.D. Genetic diversity of transmission blocking vaccine candidate (Pvs25 and Pvs28) antigen in Plasmodium vivax clinical isolates from Iran. Acta Trop. 2009, 109, 176–180. [Google Scholar] [CrossRef]
- Tsuboi, T.; Kaneko, O.; Cao, Y.-M.; Tachibana, M.; Yakushijin, Y.; Nagao, T.; Kanbara, H.; Torii, M. A rapid genotyping method for the vivax malaria transmission-blocking vaccine candidates, Pvs25 and Pvs28. Parasitol. Int. 2004, 53, 211–216. [Google Scholar] [CrossRef] [PubMed]
- González-Cerón, L.; Alvarado-Delgado, A.; Martínez-Barnetche, J.; Rodríguez, M.H.; Ovilla-Muñoz, M.; Pérez, F.; Hernandez-Avila, J.E.; Sandoval, M.A.; del Carmen Rodríguez, M.; Villarreal-Treviño, C. Sequence variation of ookinete surface proteins Pvs25 and Pvs28 of Plasmodium vivax isolates from Southern Mexico and their association to local anophelines infectivity. Infect. Genet. Evol. 2010, 10, 645–654. [Google Scholar] [CrossRef]
- Gowda, D.C.; Wu, X. Parasite recognition and signaling mechanisms in innate immune responses to malaria. Front. Immunol. 2018, 9, 3006. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, M.E.; Schofield, L. Toll-like receptors and malaria–sensing and susceptibility. J. Trop. Dis. Public. Health 2013, 2, 1–7. [Google Scholar] [CrossRef]
- Vogel, S.N.; Fitzgerald, K.A.; Fenton, M.J. TLRs: Differential adapter utilization by toll-like receptors mediates TLR-specific patterns of gene expression. Mol. Interv. 2003, 3, 466. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Holowka, T.; Song, Y.; Zierow, S.; Leng, L.; Chen, Y.; Xiong, H.; Griffith, J.; Nouraie, M.; Thuma, P.E. A Plasmodium-encoded cytokine suppresses T-cell immunity during malaria. Proc. Natl. Acad. Sci. USA 2012, 109, E2117–E2126. [Google Scholar] [CrossRef] [PubMed]
- Sirisabhabhorn, K.; Chaijaroenkul, W.; Na-Bangchang, K. Genetic diversity of human host genes involved in immune response and the binding of malaria parasite in patients residing along the Thai-Myanmar border. Trop. Med. Infect. Dis. 2021, 6, 174. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.G.; Ramasawmy, R.; Val, F.F.A.; Ibiapina, H.N.S.; Oliveira, A.C.; Tarragô, A.M.; Garcia, N.P.; Heckmann, M.I.O.; Monteiro, W.M.; Malheiro, A. Polymorphisms in TLRs influence circulating cytokines production in Plasmodium vivax malaria: TLR polymorphisms influence cytokine productions in malaria-vivax. Cytokine 2018, 110, 374–380. [Google Scholar] [CrossRef]
- Mockenhaupt, F.P.; Hamann, L.; von Gaertner, C.; Bedu-Addo, G.; von Kleinsorgen, C.; Schumann, R.R.; Bienzle, U. Common polymorphisms of toll-like receptors 4 and 9 are associated with the clinical manifestation of malaria during pregnancy. J. Trop. Dis. Public. Health 2006, 194, 184–188. [Google Scholar] [CrossRef]
- Zakeri, S.; Pirahmadi, S.; Mehrizi, A.A.; Djadid, N.D. Genetic variation of TLR-4, TLR-9 and TIRAP genes in Iranian malaria patients. Malar. J. 2011, 10, 77. [Google Scholar] [CrossRef]
- Lazarus, R.; Klimecki, W.T.; Raby, B.A.; Vercelli, D.; Palmer, L.J.; Kwiatkowski, D.J.; Silverman, E.K.; Martinez, F.; Weiss, S.T. Single-nucleotide polymorphisms in the Toll-like receptor 9 gene (TLR9): Frequencies, pairwise linkage disequilibrium, and haplotypes in three US ethnic groups and exploratory case–control disease association studies. Genomics 2003, 81, 85–91. [Google Scholar] [CrossRef]
- Kar, A.; Panigrahi, S.; Tripathy, S.; Mohapatra, M.K.; Tayung, K.; Dhangadamajhi, G. Influence of common variants of TLR4 and TLR9 on clinical outcomes of Plasmodium falciparum malaria in Odisha, India. Infect. Genet. Evol. 2015, 36, 356–362. [Google Scholar] [CrossRef]
- Rani, A.; Nawaz, S.K.; Irfan, S.; Arshad, M.; Bashir, R.; Shaheen, N. Role of MyD88-adaptor-like gene polymorphism rs8177374 in modulation of malaria severity in the Pakistani population. Braz. J. Infect. Dis. 2017, 21, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Khor, C.C.; Chapman, S.J.; Vannberg, F.O.; Dunne, A.; Murphy, C.; Ling, E.Y.; Frodsham, A.J.; Walley, A.J.; Kyrieleis, O.; Khan, A. A Mal functional variant is associated with protection against invasive pneumococcal disease, bacteremia, malaria and tuberculosis. Nat. Genet. 2007, 39, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Araujo, F.J.d.; Silva, L.D.O.d.; Mesquita, T.G.; Pinheiro, S.K.; Vital, W.d.S.; Chrusciak-Talhari, A.; Guerra, J.A.d.O.; Talhari, S.; Ramasawmy, R. Polymorphisms in the TOLLIP gene influence susceptibility to cutaneous leishmaniasis caused by Leishmania guyanensis in the Amazonas State of Brazil. PLoS Negl. Trop. Dis. 2015, 9, e0003875. [Google Scholar] [CrossRef] [PubMed]
- Shah, J.A.; Vary, J.C.; Chau, T.T.; Bang, N.D.; Yen, N.T.; Farrar, J.J.; Dunstan, S.J.; Hawn, T.R. Human TOLLIP regulates TLR2 and TLR4 signaling and its polymorphisms are associated with susceptibility to tuberculosis. J. Immunol. 2012, 189, 1737–1746. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Buelna, M.; Fafutis-Morris, M.; Tovar-Cuevas, A.J.; Alvarado-Navarro, A.; Valle, Y.; Padilla-Gutierrez, J.R.; Muñoz-Valle, J.F.; Figuera-Villanueva, L.E. Role of toll-interacting protein gene polymorphisms in leprosy Mexican patients. Biomed. Res. Int. 2013, 2013, 459169. [Google Scholar] [CrossRef] [PubMed]
- Fairhurst, R.M.; Wellems, T.E. Modulation of malaria virulence by determinants of Plasmodium falciparum erythrocyte membrane protein-1 display. Curr. Opin. Hematol. 2006, 13, 124–130. [Google Scholar] [CrossRef]
- Carvalho, B.O.; Lopes, S.C.P.; Nogueira, P.A.; Orlandi, P.P.; Bargieri, D.Y.; Blanco, Y.C.; Mamoni, R.; Leite, J.A.; Rodrigues, M.M.; Soares, I.S.; et al. On the Cytoadhesion of Plasmodium vivax–Infected Erythrocytes. J. Infect. Dis. 2010, 202, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Baluni, M.; Fatima, T.; Zia, A.; Reddy, D.H.; Dhole, T.N. Association of ICAM-1 (K469E) and MCP-1-2518 A > G polymorphism with risk of Japanese encephalitis in North Indian population. Cytokine 2018, 111, 420–427. [Google Scholar] [CrossRef]
- King, C.L.; Adams, J.H.; Xianli, J.; Grimberg, B.T.; McHenry, A.M.; Greenberg, L.J.; Siddiqui, A.; Howes, R.E.; da Silva-Nunes, M.; Ferreira, M.U. Fya/Fyb antigen polymorphism in human erythrocyte Duffy antigen affects susceptibility to Plasmodium vivax malaria. Proc. Natl. Acad. Sci. USA 2011, 108, 20113–20118. [Google Scholar] [CrossRef]
- Talha, A.A.; Pirahmadi, S.; Mehrizi, A.A.; Djadid, N.D.; Nour, B.Y.; Zakeri, S. Molecular genetic analysis of Plasmodium vivax isolates from Eastern and Central Sudan using pvcsp and pvmsp-3α genes as molecular markers. Infect. Genet. Evol. 2015, 32, 12–22. [Google Scholar] [CrossRef]
- Suphakhonchuwong, N.; Chaijaroenkul, W.; Rungsihirunrat, K.; Na-Bangchang, K.; Kuesap, J. Evaluation of Plasmodium vivax isolates in Thailand using polymorphic markers Plasmodium merozoite surface protein (PvMSP) 1 and PvMSP3. Parasitol. Res. 2018, 117, 3965–3978. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, T.; Kaslow, D.C.; Gozar, M.M.G.; Tachibana, M.; Cao, Y.-M.; Torii, M. Sequence polymorphism in two novel Plasmodium vivax ookinete surface proteins, Pvs25 and Pvs28, that are malaria transmission-blocking vaccine candidates. Mol. Med. 1998, 4, 772–782. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.K.; Singh, K.; Su, H.-P.; Klein, M.M.; Stowers, A.W.; Saul, A.J.; Long, C.A.; Garboczi, D.N. The essential mosquito-stage P25 and P28 proteins from Plasmodium form tile-like triangular prisms. Nat. Struct. Mol. Biol. 2006, 13, 90–91. [Google Scholar] [CrossRef] [PubMed]
- González-Cerón, L.; Rodríguez, M.H.; Nettel-Cruz, J.A.; Hernández-Ávila, J.E.; Malo-García, I.R.; Santillán-Valenzuela, F.; Villarreal-Treviño, C. Plasmodium vivax CSP-Pvs25 variants from southern Mexico produce distinct patterns of infectivity for Anopheles albimanus versus An. pseudopunctipennis, in each case independent of geographical origin. Parasit. Vect 2019, 12, 86–92. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Gene Type | Gene | No. Successfully Amplified | SNP | Genotypes, n (%) | HWE, p-Value | ||||
|---|---|---|---|---|---|---|---|---|---|
| Homozygous Wild Type | Heterozygous Genotype | Homozygous Mutant | Total Alleles, n (%) | ||||||
| Immune-related genes | TIRAP | 488 | rs8177374 | CC: 448 (88.9) | CT: 36 (7.1) | TT: 20 (4.0) | C: 932 (92.5) | T: 76 (7.5) | <0.001 |
| TLR6 | 475 | rs5743810 | TT: 440 (92.6) | CT: 30 (6.3) | CC: 5 (1) | T: 910 (95.8) | C: 40 (4.2) | <0.001 | |
| TLR9 | 487 | rs187084 | TT: 180 (36.9) | CT: 212 (43.4) | CC: 96 (19.7) | T: 572 (58.7) | C: 402 (41.3) | 0.024 | |
| 485 | rs5743836 | TT: 14 (2.9) | CT: 0 (0.0) | CC: 471(97.1) | T: 28 (2.9) | C: 942 (97.1) | <0.001 | ||
| TOLLIP | 499 | rs3750920 | CC: 180 (36.1) | CT: 243 (48.7) | TT: 76 (15.2) | T: 603 (60.4) | C: 395 (39.6) | 0.685 | |
| 504 | rs5743899 | AA: 210 (41.7) | AG: 250 (49.6) | GG: 44 (8.7) | A: 670 (66.5) | G: 338 (33.5) | 0.011 | ||
| Cytoadherence genes | ICAM-1 | 482 | rs5498 | AA: 288 (59.8) | AG: 165 (34.2) | GG: 29 (6.0) | A: 741(76.9) | G: 223 (23.1) | 0.411 |
| DARC | 486 | - | FYA/FYA: 396 (81.5) | FYA/FYB: 17 (3.5) | FYB/FYB: 73 (15.0) | FYA: 809 (83.2) | FYB: 163 (16.8) | <0.001 | |
| Polymorphism | Genotype | Log10 Parasite Density | p-Value (p-corrected) |
|---|---|---|---|
| TIRAP (S180L) | CC | 4.28 (2.17–5.42) | 0.303 |
| CT | 4.15 (3.34–5.26) | ||
| TT | 4.15 (3.59–5.26) | ||
| TLR6 (rs5743810) | TT | 3.95 (3.65–4.65) | 0.626 |
| CT | 4.34 (3.61–5.26) | ||
| CC | 4.19 (2.71–5.42) | ||
| TLR9 (rs5743836) | TT | 4.14 (3.48–4.91) | 0.501 |
| CC | 4.22 (2.71–5.42) | ||
| TLR9 (rs187084) | TT | 4.34 (3.31–5.42) | 0.007 (0.014) 1 0.007 (0.043) 2 |
| CT | 4.15 (3.15–5.26) | ||
| CC | 4.17 (3.15–4.77) | ||
| TOLLIP (rs3750920) | CC | 4.17 (3.31–5.26) | 0.039 (0.033) 3 |
| CT | 4.28 (3.44–5.42) | ||
| TT | 4.15 (2.71–4.66) | ||
| TOLLIP (rs5743899) | AA | 4.26 (3.09–5.42) | 0.182 |
| AG | 4.27 (2.71–5.31) | ||
| GG | 4.11 (3.34–5.08) | ||
| ICAM-1 (rs5498) | AA | 4.15 (3.15–5.42) | 0.012 (0.020) 4 |
| AG | 4.34 (3.44–5.31) | ||
| GG | 4.10 (2.71–4.68) | ||
| DARC | FYA/FYA | 4.28 (2.71–5.42) | 0.592 |
| FYB/FYB | 4.03 (3.57–4.88) | ||
| FYAB | 4.15 (3.46–5.08) |
| Position | AA Changes | Total n (%) | First Period | Second Period |
|---|---|---|---|---|
| N (%) | N (%) | |||
| E97Q 1 | E | 207 (90.8%) | 113 (49.6%) | 94 (42.5%) |
| Q | 21 (9.2%) | 18 (7.9%) | 3 (1.3%) | |
| I130T | T | 228 (100%) | 131 (57.5%) | 97 (42.5%) |
| Q131K | Q | 212 (93%) | 123 (54.0%) | 89 (39%) |
| K | 16 (7%) | 8 (3.5%) | 8 (3.5%) |
| K | S | No. Mutations | H | Hd ± SD | π ± SD | Tajima’s D (p-Value) | |
|---|---|---|---|---|---|---|---|
| EGF1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| EGF2 | 0.161 | 1 | 1 | 2 | 0.161 ± 0.031 | 0.0012 ± 0.0002 | −0.034 (>0.1) |
| EGF3 | 0.123 | 1 | 1 | 2 | 0.123 ± 0.029 | 0.0009 ± 0.0002 | −0.256 (>0.1) |
| EGF4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Full length | 0.284 | 2 | 2 | 4 | 0.226 ± 0.036 | 0.0006 ± 0.0001 | −0.1964 (>0.1) |
| N | SS | EGF1 | EGF2 | EGF4 | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Geographic origin | 2 * | 27 | 35 | 38 | 40 | 45 | 55 | 61 | 63 | 79 | 86 | 87 | 97 | 101 | 106 | 123 | 130 | 131 | 132 | 135 | 136 | 146 | 153 | 156 | 158 | 159 | 161 | 162 | 178 | 180 | |
| Sal-I strain | N | D | L | M | N | M | E | K | E | C | A | Q | E | L | C | I | I | Q | S | C | S | E | K | E | A | C | L | K | K | Q | |
| India | 100 | - | - | - | - | - | - | - | - | - | C/R | - | - | E/Q | - | - | - | T | Q/K | - | - | - | - | - | - | - | - | - | - | - | - |
| Iran | 4 | - | - | - | - | - | - | - | - | - | - | - | Q/K | E/Q | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - |
| China | 354 | - | D/N | L/M | - | - | - | - | - | - | - | - | Q/L | E/Q | - | - | - | T | Q/K | - | - | - | - | - | - | - | - | - | - | - | - |
| Thailand | 3 | - | - | - | - | - | - | - | - | - | - | - | - | E/Q | - | - | - | T | Q/K | - | - | - | - | - | - | - | - | - | - | - | - |
| Myanmar | 62 | - | - | - | - | N/S | - | E/G | - | - | - | - | - | E/Q | - | - | - | T | Q/K | - | - | S/P | E/G | - | - | A/V | - | - | - | - | - |
| South Korea | 100 | N/D | - | L/P | - | - | M/T | K/C | E/K | - | A/P | - | E/Q | L/S | C/R | I/F | T | - | - | C/G | - | - | K/R | E/D | - | C/R | L/W | K/R | K/R | Q/E | |
| North Korea | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Bangladesh | 4 | - | - | - | - | - | - | - | - | - | - | - | - | E/K | - | - | - | T | Q/K | - | - | - | - | - | - | - | - | - | - | - | - |
| Indonesia | 1 | - | - | - | - | - | - | - | - | - | - | - | - | Q | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Mexico | 64 | - | - | - | - | - | - | - | - | - | - | - | Q/K | - | - | - | - | I/T | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Colombia | 1 | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Venzuela | 16 | - | - | - | M/T | - | - | - | - | - | - | - | Q/K | - | - | - | - | - | - | - | C/R | S/R | - | - | - | - | - | - | - | - | - |
| Brazil | 1 | - | - | - | - | - | - | - | - | - | - | - | Q/K | - | - | - | - | I/T | - | - | - | - | - | - | - | - | - | - | - | - | - |
| PNG | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | T | K | R | - | - | - | - | - | - | - | - | - | - | - |
| Mauratinea | 1 | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Present Study | 228 | - | - | - | - | - | - | - | - | - | - | - | - | E/Q | - | - | - | T | Q/K | - | - | - | - | - | - | - | - | - | - | - | - |
| Overall = 16 countries | |||||||||||||||||||||||||||||||
| N of substitutioins | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 7 | 9 | 1 | 1 | 1 | 14 | 7 | 1 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| % Frequency | 6.3 | 6.3 | 12.5 | 6.3 | 6.3 | 6.3 | 6.3 | 6.3 | 6.3 | 6.3 | 6.3 | 43.8 | ### | 6.3 | 6.3 | 6.3 | 87.5 | 43.8 | 6.3 | 12.5 | 12.5 | 6.3 | 6.3 | 6.3 | 6.3 | 6.3 | 6.3 | 6.3 | 6.3 | 6.3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jalei, A.A.; Chaijaroenkul, W.; Na-Bangchang, K. Genetic Diversity of Plasmodium vivax Surface Ookinete Protein Pvs25 and Host Genes in Individuals Living along the Thai–Myanmar Border and Their Relationships with Parasite Density. Microbiol. Res. 2024, 15, 693-707. https://doi.org/10.3390/microbiolres15020045
Jalei AA, Chaijaroenkul W, Na-Bangchang K. Genetic Diversity of Plasmodium vivax Surface Ookinete Protein Pvs25 and Host Genes in Individuals Living along the Thai–Myanmar Border and Their Relationships with Parasite Density. Microbiology Research. 2024; 15(2):693-707. https://doi.org/10.3390/microbiolres15020045
Chicago/Turabian StyleJalei, Abdifatah Abdullahi, Wanna Chaijaroenkul, and Kesara Na-Bangchang. 2024. "Genetic Diversity of Plasmodium vivax Surface Ookinete Protein Pvs25 and Host Genes in Individuals Living along the Thai–Myanmar Border and Their Relationships with Parasite Density" Microbiology Research 15, no. 2: 693-707. https://doi.org/10.3390/microbiolres15020045
APA StyleJalei, A. A., Chaijaroenkul, W., & Na-Bangchang, K. (2024). Genetic Diversity of Plasmodium vivax Surface Ookinete Protein Pvs25 and Host Genes in Individuals Living along the Thai–Myanmar Border and Their Relationships with Parasite Density. Microbiology Research, 15(2), 693-707. https://doi.org/10.3390/microbiolres15020045