Abstract
The genus Ganoderma has been little studied in arid areas worldwide. Ganoderma subincrustatum and Ganoderma weberianum strains were obtained from the Sonoran Desert, Sonora, Mexico. Ganoderma spp. synthesize triterpenoids such as ganoderic acids with antiproliferative activity because they inhibit specific targets, induce apoptosis, and increase the activity of killer cells. Mycelium and fruiting body chloroform extracts from G. subincrustatum and G. weberianum were tested on HeLa, A549, L-929, and RAW 264.7 cell lines. Extracts from the fruiting body present higher antiproliferative activity than mycelium. All extracts induced vesicle and cellular debris formation in all cell lines, being non-selective for cancerous cells. Chloroform extract from G. subincrustatum fruiting bodies presented higher activity against all cell lines. Fractions F7 and F15 from this extract exhibited an IC50 of 37.9 and 41.9 µg/mL on the A549 cell line, respectively; however, chloroform crude extract showed higher activity (IC50 of <25 µg/mL) in all cell lines. Flow cytometry assays of F7 revealed cell death by apoptosis in A549 cells. NMR suggested the presence of ganoderic acids in F7. In future research, it will be interesting to characterize these fractions (metabolites, their bioactivities, and mechanism of action).
1. Introduction
Some species of Ganoderma, mainly Ganoderma lucidum, have been used in traditional Asian medicine for more than six millennia to modulate immunological activity and to treat hypertension, arthritis, asthma, anorexia, hepatitis, cancer, and other illnesses [1,2,3]. The study of natural products is the basis for the discovery of new molecules with different structures and bioactivities. It allows improving the drug design for effective anticancer therapies [4,5]. Cancer is the most prevalent disease worldwide; according to the World Health Organization, it is the first or second highest cause of death before the age of 70 [6]. Since cancer is characterized by uncontrolled cell proliferation, angiogenesis, and apoptosis evasion, the discovery of novel molecules and extracts to induce programmed cell death is relevant. Thus, apoptosis is one of the major targets for non-surgical cancer treatment [7].
Ganoderma has a cosmopolitan distribution, comprising more than 450 species by the year 2022, many of them unstudied (www.indexfungorum.org/names/Names.asp; accessed on 04/September/2023). The most studied species is G. lucidum; however, metabolites and bioactivities have been reported for another 24 species. Some of the studied species are G. amboinense, G. applanatum, G. boninense, G. capense, G. lipsiensis, G. neojaponicum, G. orbiforme, G. pfeifferi, G. sinense, G. tropicum, G. tsugae, among others [8,9]. Isolated compounds from G. weberianum have antidiabetic and antiproliferative activity [10]. This indicates the lack of information on more than 400 species of Ganoderma genus.
Ganoderma species synthesize biologically active compounds, mainly lanostane-type triterpenoids (e.g., ganoderic acids: GA; triterpene alcohols), which exhibit antiproliferative activity against different cell lines [11,12,13]. Ganoderma triterpenoids are synthesized via the mevalonate/isoprenoid pathway from acetyl-coenzyme A [14]. Stages of conversion, construction, condensation, and post-modification are involved in its biosynthesis. The final stage, once lanosterol is formed, includes acylation, oxidation, and reduction reactions, to produce a complex group of molecules with different functional groups, like hydroxyl, carboxyl, and acetoxy [2,8]. Due to the relevance of Ganoderma triterpenoids, genes involved in this pathway have been studied [15].
For G. lucidum the expression of genes (HMGR, FPS, SQS, LS) related to the mevalonate/isoprenoid pathway increases during primordia development, which is correlated with the highest triterpenoid content in this phase of its life cycle [16]. Likewise, it has been reported that secondary mycelium contains mainly 3α-substituted compounds while the fruiting body presents mainly 3keto- or 3β-substituted compounds [17], which could be related to its bioactivity. Traditionally, Ganoderma cultivation used wood logs outdoors. Several decades ago, a modern indoor cultivation technique was developed using wood sawdust and agricultural by-products [18]. Because fruiting body production is a time-consuming activity for obtaining products from Ganoderma (months), the production of mycelia by liquid culture seems to be a promising technique; because it offers inter-batch homogeneity, less time is required, and fermentation factors can be directed toward the production of bioactive compounds [19,20].
Ganoderma weberianum is a poorly studied species with few reports, while for G. subincrustatum we did not find any work on the profile of its bioactive compounds with antiproliferative activity. Both species are distributed in Sonora, Mexico; therefore, to take advantage of our natural resources, this work aims to generate knowledge about the antiproliferative activity of chloroform extracts from the mycelium and fruiting bodies of G. subincrustatum and G. weberianum against different cell lines. Furthermore, apoptosis induction and nuclear magnetic resonance (NMR) of the bioactive fraction were addressed.
2. Materials and Methods
2.1. Fungal Strains
Ganoderma subincrustatum was collected from a live Prunus persica tree in a peach orchard at La Costa de Hermosillo, while G. weberianum was taken from a Quercus sp. tree in La Sierra de Mazatán, Ures, Sonora, México. Both species were previously recorded by López-Peña et al. [21,22]. Strains were deposited in the Fungus Collection of the Plant and Fungi Biotechnology Laboratory of the Research Center in Food and Development A.C.; G. subincrustatum was registered as BH-1 and G. weberianum as BH-21. Ganoderma strains were maintained on malt extract agar in Petri dishes.
2.2. Liquid and Solid Culture
Liquid medium composition for mycelium production comprised the following: glucose (16 g/L), peptone (2.93 g/L), corn flour (21 g/L), and soybean protein powder (7 g/L) [23]. A total of 400 mL of culture medium was inoculated with 16 agar disks 1 cm in diameter with mycelium for each strain. Liquid fermentation was maintained at 28 ± 1 °C, shaking at 120 rpm, initial pH 5.5, for 13 d in the dark. Mycelium was recovered by centrifugation and washed with distilled water. Fruiting body production was carried out using oak wood chips. The sterilized substrate was inoculated with spawn (wheat grain at 5% w/w) previously prepared. Incubation was conducted at 27 ± 1 °C in the dark. Fruiting body development conditions were maintained at 27 ± 1 °C, 90% relative humidity, light intensity of 48–58 μmol·m−2·s−1, and 500 to 1000 ppm of CO2 in air [24]. The fruiting body was harvested when the pileus fully developed or stopped growing. Mycelia and fruiting bodies were freeze-dried and stored in a dry place until use.
2.3. Extracts
Mycelia and dried fruiting bodies were ground with an electric mixer to obtain a powder, from which 1 g of sample was extracted with 95% ethanol (30 mL) by sonication for 45 min (Ultra Sonic Cleaner, Branson 2210R-MT), with a power of 210.6 W and operating frequency of 40 kHz. Biomass was removed by centrifugation and filtration; then, the extracts were concentrated under reduced pressure conditions (BÜCHI Rotavapor, RE 121). Afterwards, dried residues were suspended in distilled water and then extracted with chloroform; the chloroform layer was recovered [23], then dried at 30 °C and stored at −20 °C until bioactivity assays. The extracts obtained were the following: Gsm (G. subincrustatum mycelial extract), Gsfb (G. subincrustatum fruiting body extract), Gwm (G. weberianum mycelial extract), and Gwfb (G. weberianum fruiting body extract).
2.4. Bioguided Fractionation and Chemical Analysis
Ganoderma subincrustatum fruiting body extract (2.2 g) was fractioned by column chromatography (5.5 × 50 cm) using Sigma Aldrich silica gel 60 (200–400 mesh). The column was eluted with ethyl acetate-methanol (EtOAc-MeOH) in different proportions (v/v, 100:0, 95:5, 90:10, 85:15, 80:20, 50:50, 0:100). Three hundred thirty-three fractions (10 mL) were collected and analyzed by thin-layer chromatography (TLC). The spots on the TLC were visualized under UV light (254/366 nm) then stained with p-anisaldehyde-H2SO4-EtOH (1:1:98), followed by heating at 110 °C until positive areas appeared [25]. Twenty-six (F1 to F26) subfractions were grouped according to TLC patterns: F1 (8.2 mg), F2 (83.1 mg), F3 (13.4 mg), F4 (27.8 mg), F5 (323.8 mg), F6 (93.3 mg), F7 (114.2 mg), F8 (6.2 mg), F9 (37 mg), F10 (12.7 mg), F11 (34.3 mg), F12 (22.6 mg), F13 (61.7 mg), F14 (8.5 mg), F15 (39.2 mg), F16 (11.8 mg), F17 (44.3 mg), F18 (18.7 mg), F19 (14.3 mg), F20 (7.8 mg), F21 (9.3 mg), F22 (17.4 mg), F23 (18.2 mg), F24 (30.8 mg), F25 (55 mg), and F26 (337. 8 mg).
The chemical analysis of the F7 subfraction was performed by 1H and 13C NMR analysis (nuclear magnetic resonance). The NMR spectra were recorded at 400 MHz for 1H and 100.6 MHz for 13C in an Agilent 400 MHz NMR Magnet Fourier transform instrument. The spectra were recorded in CD3Cl (Chloromethane-D3) at 25 °C and the chemical shift was expressed in δ values (ppm); TMS (tetramethylsilane) was used as an internal reference [26].
2.5. Cell Culture
Cell lines were cultured in DMEM (Dulbecco’s Modified Eagle Medium), supplemented with 5% heat-inactivated fetal bovine serum and penicillin-streptomycin (100 U/mL) in 25 cm2 culture dishes, which were kept in an incubator isotherm (Fisher Scientific, Waltham, MA, USA) with 5% CO2, at 37 °C and 95% relative humidity. Cancer cell lines HeLa (human cervical carcinoma), A549 (human lung carcinoma), and non-cancerous cell line L-929 (subcutaneous connective tissue) were purchased from the American Type Culture Collection (ATCC, Gaithersburg, MD, USA). The RAW 264.7 (macrophages transformed by the virus Albeson leukemia) line was kindly provided by Dr. Emil R. Unanue (Department of Pathology and Immunology, Washington University in St. Louis, MO, USA).
2.6. Antiproliferative Activity
The antiproliferative activity of the chloroform extracts and chromatographic fractions was determined using the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay (Cell proliferation Kit I, Roche), according to Müller et al. [27], with some modifications. The MTT assay is based on the ability of viable cells to reduce MTT to formazan through mitochondrial dehydrogenases. Briefly, cell suspensions of 200,000 cells/mL were placed in 96 ELISA plates (Costar, Corning, NY, USA) and incubated for 24 h. Afterward, extracts were dissolved in dimethyl sulfoxide (DMSO) and redissolved in DMEM to give a final DMSO concentration of 0.25%, showing no interference with the assay. Extract evaluations (25–100 µg/mL) were performed over 48 h. After the treatment, morphological changes were observed with an inverted microscope and microphotographs were taken with the help of a digital camera. Finally, according to the manufacturer’s instructions, 10 µL of MTT solution was added to each well and incubated for 4 h in a humid atmosphere at 37 °C. A total of 100 µL of the solubilization buffer was added to the wells and incubated overnight, and absorbance measurements were performed at 570 and 655 nm in a microplate reader. The antiproliferative activity was determined as IC50 (concentration of extract or fraction that inhibits the 50% of cell proliferation) value using GraphPad Prism 5 (GraphPad Software, Inc., San Diego, CA, USA).
2.7. Flow Cytometry
The apoptotic effect of subfraction F7 on A549 cells was determined by double staining with annexin V-FITC (AN) and propidium iodide (PI) (Sigma-Aldrich, Saint Louis, MO, USA). Briefly, 2 × 105 cells/mL were placed in a 12-well cell culture plate (Costar, Corning, NY, USA) and incubated for 24 h. Cells were then treated with F7 (40 µg/mL) for 24 h. The antineoplastic doxorubicin hydrochloride (DOXO) (1 µM) was used as a positive control. After treatment, cells were collected and washed twice with PBS and then resuspended for 10 min in a staining solution containing AN (1 µg/mL) and PI (10 µg/mL). Finally, 1 × 105 cells were acquired by fluorescence-activated cell sorting (FACS) (Canto II FACS, Becton Dickinson, CA, USA) according to Torres-Moreno et al. [26]. A dot plot was introduced to identify viable cells, dead cells, cells in early apoptosis, and cells in late apoptosis. Viable cells were recognized as doubly negative for AN and PI (AN−/PI−), dead cells were negative for AN and positive for PI (AN−/PI+), cells in early apoptosis were identified as AN positive, and PI negative (AN+/PI−), whereas cells in late apoptosis were recognized as being doubly positive for AN and PI (AN+/PI+). The cells in each quadrant were expressed as a percentage of the total number of stained cells.
2.8. Statistical Analysis
A one-way analysis of variance was applied to the antiproliferative activity and apoptosis induction. The Tukey-Kramer test was performed to compare treatments with their respective controls (p ≤ 0.05). Data are presented as mean ± standard deviation.
3. Results and Discussions
3.1. Antiproliferative Activity of G. subincrustatum and G. weberianum
Fruiting body extracts showed higher bioactivity than mycelium extracts in all evaluated cell lines, although we used a complex culture media with acceptable GA production in G. lucidum [23]. Gsfb extract exhibited higher activity than Gwfb against all cell lines. The lower activity was shown in Gwm with an IC50 ≥ 100 (µg/mL) in all cell lines. The non-cancerous cell line L-929 was highly affected by Gsfb and Gwfb, which suggests that the antiproliferative activity of Gsfb and Gwfb extracts is non-selective for cancer cell lines (Table 1). This could be attributed to a higher production of bioactive compounds in fruiting bodies, as was also observed by Chen et al. [16].

Table 1.
Antiproliferative activity of Ganoderma spp. strains against several cell lines: mycelium vs. fruiting body.
Crude extracts with IC50 of ≤30 µg/mL are considered promising, according to the National Cancer Institute [28]. Several extracts, fractions, and isolated compounds from Ganoderma spp. have proved to exert antiproliferative activity against different cell lines. Zolj et al. [29] tested a commercial product from G. lucidum spores (ReishiMax GLp) with 6% triterpenes content. They obtained an antiproliferative activity at 0.4–10 mg/mL (24–120 h treatments) on the human non-small cell lung adenocarcinoma (NCI-H1793) cell line. Liu et al. [30] investigated the antiproliferative activity of ethanolic extracts from G. lucidum and G. sinensis against different tumor cell lines and obtained higher IC50 values than the present study (71–378 µg/mL). Nevertheless, the extracts tested in the present study were designed to obtain a triterpenoid-enrich fraction [23], as we partitioned the crude ethanolic extract between water and chloroform, and the chloroform layer was recovered for the first stage of antiproliferative assays.
After 48 h of treatment, Gsfb and Gwfb induced vesicle and debris formation in all cell lines, being more evident in the A549 cell line (Figure 1). Apoptosis is the main target for anticancer therapy and some of the morphological hallmarks of apoptosis induction are cell shrinkage, fragmentation, formation of apoptotic bodies, and phagocytosis by adjacent cells [31]. Crude extracts and different triterpenoids isolated from Ganoderma spp. induce morphological changes related to apoptosis on several cell lines. Jang et al. [32] evaluated an ethanol–water (25/75) extract of G. lucidum on human gastric carcinoma cells. They observed morphological changes with treatment (1.5% for 72 h), like cytoplasmatic filaments and apoptotic bodies. Methanolic extract (80%) of G. applanatun also induced morphological changes in a human colon cancer cell line (Caco-2), at 80 and 160 µg/mL [33]. Furthermore, ganoderic acids Mf and S induce morphological changes and apoptosis in HeLa cells [34]. These results were the basis for choosing Gsfb extract for further analysis, including column chromatography to obtain different fractions and antiproliferative activity of those fractions on A549 cell lines.
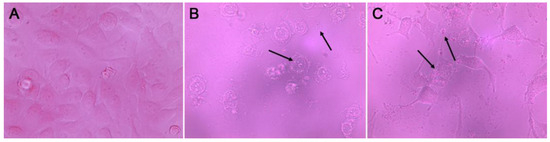
Figure 1.
A549 cell line morphology. (A) Control, (B) Ganoderma weberianum fruiting body extract (100 µg/mL), (C) Ganoderma subincrustatum fruiting body extract (100 µg/mL). 40× magnification. Arrows indicate vesicle and debris formation.
3.2. Bioactive Fractions of G. subincrustatum
A column chromatography assay yielded 333 fractions (10 mL), which were grouped into 26 fractions, according to TLC patterns. Then, the antiproliferative activity of the 26 chromatographic grouped fractions (F1–F26) was tested against the A549 cell line. F7 and F15 showed significant differences at the evaluated concentrations (50–6.25 µg/mL), compared to the control (p < 0.05) with IC50 values of 37.9 ± 6.2 and 41.9 ± 4.9 µg/mL, respectively (Figure 2). The IC50 values of both fractions were higher than those of the chloroform extract. This was probably caused by the separation of some bioactive molecules by column fractionation. Due to the significance of lung cancer worldwide, the A549 cell line has been used to assess the antiproliferative activity of several products through numerous studies. The leaf extracts of Sarcocephalus pobeguinii showed a correlation between free radical scavenging capacity and the antiproliferative activity against A549 cells (R2 = 0.99), with an IC50 of 50.46–857.25 µg/mL [28], while in another study the extracts of Populus niga buds showed an IC50 of 72.49 µg/mL [35]. Novel triazole hybrids of myrrhanone C, a natural polypodane bicyclic triterpene, the oxime-based triazole 4a showed potent activity vs. the A549 cell line with an IC50 of 6.16 µm. This compound arrested the cell cycle in the G2/M phase and induced apoptosis [36]. Novel 1,2,3-triazole hybrids of myrrhanone B, compounds 30 and 29 showed activity vs. the A549 cell line with IC50 values of 15.34 ± 0.37 and 28.12 ± 2.64 µm, respectively; compound 29 increased to 40.8% the SubG1 cells at 10 µm, which induced cell apoptosis [37]. Furthermore, titanium dioxide nanoparticles (TiO2 NPs) (5 nm) induced cytotoxicity, DNA damage, and apoptosis in the A549 cell line in a concentration range of 50 to 200 µg/mL for 48 h. Flow cytometric analysis showed a G2/M phase arrest and a higher percentage of apoptotic cells [38]. Carvacrol nanoemulsion induced apoptosis through increasing apoptotic proteins (Bax, caspase 3, caspase 9, cytochrome C), cell cycle arrest (reducing CDK2, CDK4, cyclin E, cyclin D1), and inhibiting autophagy (reducing the conversion of LC-3) into the doxorubicin-resistant A549 cell line [39]. Because of the aforementioned, we selected F7 for apoptosis analyses by flow cytometry and NMR characterization.
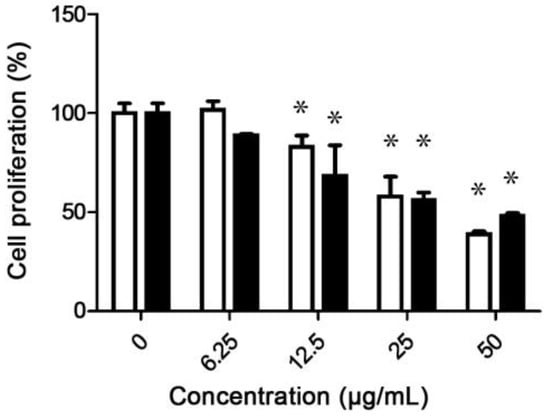
Figure 2.
Antiproliferative activity of fraction F7 (white bars) and fraction F15 (black bars) against the A549 cell line. Asterisks indicate significant differences between treatments and respective controls (p < 0.05).
3.3. Apoptosis Induction of F7
Targeting apoptosis cell death has great relevance in cancer treatment due to its uncontrolled cell proliferation. Flow cytometry allows the detection of morphological and biochemical hallmarks in cells by apoptosis. AN and PI allow classifying the cells through the measurement of the differences in the plasmatic membrane integrity and permeability, for which the cells can be classified as viable cells (FITC−/PI−), dead cells (FITC−/PI+), early apoptosis (FITC+/PI−), and late apoptosis (FITC+/PI+) [40]. F7 was able to induce cell death by apoptosis; after 24 h of treatment, total apoptosis was 5.9% in the control vs. 19.8% in the cells treated with F7 (p < 0.05), with the highest percentage in late apoptosis. However, the percentage of dead cells after F7 treatment was higher than the percentage of total apoptosis. These findings suggest that F7 induces an antiproliferative effect in A549 cells through apoptotic cell death and other mechanisms that disrupt cell membrane integrity (Figure 3).
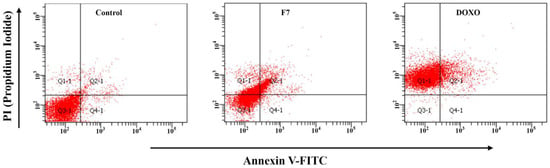
Figure 3.
Apoptosis induction of F7 (40 µg/mL for 24 h) from Ganoderma subincrustatum. Dot plots represent mean ± standard deviation from three independent experiments (n = 3). Control (DMSO 0.02%), F7 (Fraction 7), DOXO (Doxorubicin hydrochloride, 1 µM).
Different ganoderic acids and extracts from Ganoderma spp. induce apoptosis cell death. GA T from G. lucidum induces mitochondria-mediated apoptosis in 95-D cancer cells (lung) with 50% of apoptotic cells at 8 h after treatment (50 µg/mL) [41]. GA H (~99% purity) induces a high apoptosis rate at 2 mg/mL after 24 h of treatment in DU-145 human prostate cancer cells [42]. An ethanol extract (25%) of G. lucidum shows ~30% of apoptosis at high concentration (2%) in the human gastric carcinoma AGS cell line [32]. G. applanatum methanol extract (80%) induces apoptosis via p53-dependent and independent pathways in Ehrlich’s solid tumor [33].
The non-small cell lung cancer cell line is characterized by the overexpression of the epidermal growth factor receptor (EGFR) and its downstream effector pathways (PI3K/AKT/mTOR, RAS/RAF/MAPK, and STAT3) that promote several oncogenic processes such as cell proliferation, survival, differentiation, angiogenesis, invasion, and metastasis [26]. Xia et al. [43] reported that ganoderic acid DM induces apoptosis and autophagy in A549 and NCI-H460 through AKT/mTOR inhibition. The C-26 carboxylic group of ganoderic acid DM binds with α and β-tubulin in PC-3 (human prostate cancer) cells via an amidation reaction [44]. This suggests that the apoptotic effect of molecules in F7 could be associated with the activity modulation of these proteins by ganoderic acids in F7, particularly with disruption of the proper functioning of microtubules.
Likewise, if this molecule is produced by our strains, it can probably explain the non-selective bioactivity against different cell lines. Several bioactive molecules in certain extracts could result in high activity because components may exert bioactivity through different mechanisms of action. On the other hand, compared to the non-specific antineoplastic agent DOXO, F7 was more effective in inducing apoptosis in A549 cells (p < 0.05). However, the percentage of dead cells was higher in DOXO-treated cells compared to F7 (p < 0.05). This may be associated with the lower cytotoxicity of F7 relative to DOXO. The cytotoxicity of DOXO associated with its non-selectivity against cancer cells has been widely reported [45,46,47]; the clinical use of DOXO is limited by its side effects, the most dangerous is dose-dependent cumulative cardiotoxicity [48]. These findings can explain why DOXO induces greater cell death in A549 cells in relation to F7, which shows a greater apoptotic capability in relation to this reference drug. More research is needed to understand the non-selective mechanism.
Other authors have reported the ability of ganoderic acid-R (GA-R) to restore the sensitivity of multidrug-resistant (MDR) cells to DOXO via apoptosis induction. GA-R induces apoptosis in KB-A-1/Dox MDR cells in a dose-response manner from 35% to 90% after 48 h of treatment. The evaluation of the cytotoxicity showed that DOXO (0.75 µM) induces cytotoxicity less than 30% in KB-A-1/Dox cells. However, the combination of DOXO (0.75 µM) plus GA-R (5 µg/mL) significantly increased the inhibition ratio by more than 50%, which indicates that GA-R restored the sensibility of KB-A-1/Dox cells to DOXO [49]. Although GA-R shows greater apoptotic activity in KB-A-1/Dox MDR cells compared to the apoptotic effect shown by F7 in HeLa cells, these results suggest that F7 or ganoderic acids from Gsfb can be candidates to be investigated for their possible effect against MDR cancer cells.
3.4. NMR Analysis of F7 Chromatographic Fraction
Lanostane-type triterpenoids are the major bioactive constituents of the Ganoderma genus. F7 showed characteristic displacements of lanostane-type terpenoids by NMR (1H, 400 MHz; 13C, 100.6 MHz). F7 presented lanostane-type signals between 10–80, ~70, 135–150, and 195–220 ppm (Figure 4A). Lanostane-type molecules can show 0–3 signals between 195 and 220 ppm (Figure 4B). These characteristics shifts of ketone carbonyl groups can occur at C-3, C-7, C-11, C-23, and C-26 of Ganoderma lanostane-type compounds (Figure 4C) [13]. For each spectrum of F7, three signals above 195 ppm were observed, suggesting a maximum of three different lanostane-type compounds with a carbonyl group (Table 2). In fact, 164 ppm displacement can be explained if a lanostane is included. To know which specific molecules in F7 are exerting antiproliferative and apoptosis-inducing activity, it is necessary to design experiments for the isolation of its compounds. In addition, it is necessary to investigate the mechanisms of action of these.
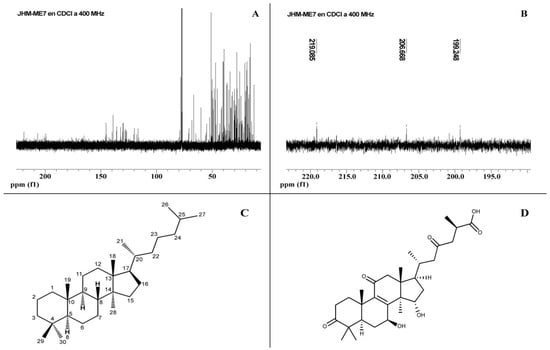
Figure 4.
(A,B) 13C-NMR of Fraction F7 from Ganoderma subincrustatum. (C) Lanostane with α and β-unsaturated carbonyl groups. (D) Ganoderic acid A, one of the most abundant triterpenoids in G. lucidum.
Table 2.
Chemical displacement of fraction F7 from Ganoderma subincrustatum with a lanostane-type skeleton.
Most of the Ganoderma highly oxygenated triterpenoids contain 27 or 30 carbon atoms, and only a few have 24. Acid triterpenoids with 27–30 carbon atoms in Ganoderma spp. have been isolated from mycelia, fruiting bodies, and spores, showing a carboxyl group at C26 [50]. Some of the GA isolated from fruiting bodies were A, B8, C1, H, K, AM1, and J (Figure 4D) from spores γ, δ, ε, ζ, η, θ, and β. Moreover, GA Mi was obtained from the mycelium of G. lucidum. The substituents can be at the C-3, 7, 11, 12, 15, 22, 23, 24, and 25 positions of the lanostane skeleton [13].
4. Conclusions
Chloroform extracts from the fruiting body of Ganoderma subincrustatum and G. weberianum were more effective in inhibiting cell proliferation compared to mycelium extracts, suggesting that the compound’s profile and its concentration vary at different development stages of Ganoderma species. The F7 chromatographic fraction showed the highest antiproliferative activity against the A549 lung cancer cell line and had the ability to induce apoptotic cell death. NMR analysis suggests that F7 is composed of ganoderic acids due to the 171.8 signal (13Ca) corresponding to a carboxyl group at C26; this could be involved in the antiproliferative and apoptotic effect of this fraction. The greater antiproliferative effect of the chloroform extracts compared to the chromatographic fractions suggests the synergistic or additive effect of the bioactive metabolites in the extracts. The strain of G. subincrustatum studied in this work should be used for further experiments to characterize the metabolite profile and its bioactivities and to fully understand its mechanisms of action.
Author Contributions
Conceptualization, D.L.-P. and M.E.; methodology, D.L.-P., H.T.-M. and M.V.-G.; validation, D.L.-P. and M.E.; formal analysis, D.L.-P., H.T.-M. and M.V.-G.; investigation, D.L.-P., A.G. and M.E.; resources, R.E.R.-Z. and M.E.; data curation, D.L.-P.; writing—original draft preparation, D.L.-P.; writing—review and editing, D.L.-P., H.T.-M., M.V.-G. and M.E.; visualization, D.L.-P., R.E.R.-Z. and M.E.; supervision, M.E. and A.G.; project administration, M.E.; funding acquisition, R.E.R.-Z. and M.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Council for Humanities, Science and Technology México (CONAHCyT A1-S-34237).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are grateful to Georgina Vargas for her technical assistance in the laboratory analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ahmad, M.F.; Wahab, S.; Ahmad, F.A.; Ashraf, S.A.; Abullais, S.S.; Saad, H.H. Ganoderma lucidum: A potential pleiotropic approach of ganoderic acid in health reinforcement and factors influencing their production. Fungal Biol. Rev. 2022, 39, 100–125. [Google Scholar] [CrossRef]
- Angulo-Sanchez, L.T.; López-Peña, D.; Torres-Moreno, H.; Gutiérrez, A.; Gaitán-Hernández, R.; Esqueda, M. Biosynthesis, gene expression, and pharmacological properties of triterpenoids of Ganoderma species (Agaricomycetes): A review. Int. J. Med. Mushrooms 2022, 24, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, J.Q.; Zhang, J.; Li, Z.M.; Liu, H.G.; Wang, Y.Z. Traditional uses, chemical components and pharmacological activities of the genus Ganoderma P. Karst.: A review. RSC Adv. 2020, 10, 42084–42097. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ng, J.M.; Wong, C.C.; Ng, E.K.W.; Yu, J. Molecular alterations of cancer cell and tumour microenvironment in metastatic gastric cancer. Oncogene 2018, 37, 4903–4920. [Google Scholar] [CrossRef]
- Lichota, A.; Gwozdzinski, K. Anticancer activity of natural compounds from plant and marine environment. Int. J. Mol. Sci. 2018, 19, 3533. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A target for anticancer therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef]
- Galappaththi, M.C.A.; Patabendige, N.M.; Premarathne, B.M.; Hapuarachchi, K.K.; Tibpromma, S.; Dai, D.Q.; Suwannarach, N.; Rapior, S.; Karunarathna, S.C. A review of Ganoderma triterpenoids and their bioactivities. Biomolecules 2022, 13, 24. [Google Scholar] [CrossRef]
- Paterson, R.R. Ganoderma—A therapeutic fungal biofactory. Phytochemistry 2006, 67, 1985–2001. [Google Scholar] [CrossRef]
- Yang, L.; Kong, D.X.; Xiao, N.; Ma, Q.Y.; Xie, Q.Y.; Guo, J.C.; Ying, D.C.; Ma, H.X.; Hua, Y.; Dai, H.F.; et al. Antidiabetic lanostane triterpenoids from the fruiting bodies of Ganoderma weberianum. Bioorg. Chem. 2022, 127, 106025. [Google Scholar] [CrossRef]
- Cao, L.; Jin, H.; Liang, Q.; Yang, H.; Li, S.; Liu, Z.; Yuan, Z. A new anti-tumor cytotoxic triterpene from Ganoderma lucidum. Nat. Prod. Res. 2021, 36, 4125–4131. [Google Scholar] [CrossRef]
- Li, X.C.; Liu, F.; Su, H.G.; Peng, C.; Zhou, Q.M.; Liu, J.; Huang, Y.J.; Guo, L.; Xiong, L. Twelve undescribed derivatives of ganoderic acid isolated from Ganoderma luteomarginatum and their cytotoxicity against three human cancer cell lines. Phytochemistry 2021, 183, 112617. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Zhang, H.; Sun, X.; Zhao, H.; Wu, L.; Zhu, D.; Yang, G.; Shao, Y.; Zhang, X.; Mao, X.; et al. A comprehensive review of the structure elucidation and biological activity of triterpenoids from Ganoderma spp. Molecules 2014, 19, 17478–17535. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Yen, G.C. Ganoderic acid and lucidenic acid (triterpenoid). Enzymes 2014, 36, 33–56. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.S.; Ji, S.L.; Ren, M.F.; He, Y.L.; Jing, X.R.; Xu, J.W. Enhanced accumulation of individual ganoderic acids in a submerged culture of Ganoderma lucidum by the overexpression of squalene synthase gene. Biochem. Eng. J. 2014, 90, 178–183. [Google Scholar] [CrossRef]
- Chen, S.; Xu, J.; Liu, C.; Zhu, Y.; Nelson, D.R.; Zhou, S.; Li, C.; Wang, L.; Guo, X.; Sun, Y.; et al. Genome sequence of the model medicinal mushroom Ganoderma lucidum. Nat. Commun. 2012, 3, 913. [Google Scholar] [CrossRef]
- Xu, J.W.; Zhao, W.; Zhong, J.J. Biotechnological production and application of ganoderic acids. Appl. Microbiol. Biotechnol. 2010, 87, 457–466. [Google Scholar] [CrossRef]
- Zhou, X.W.; Su, K.Q.; Zhang, Y.M. Applied modern biotechnology for cultivation of Ganoderma and development of their products. Appl. Microbiol. Biotechnol. 2012, 93, 941–963. [Google Scholar] [CrossRef]
- Hsu, K.D.; Cheng, K.C. From nutraceutical to clinical trial: Frontiers in Ganoderma development. Appl. Microbiol. Biotechnol. 2018, 102, 9037–9051. [Google Scholar] [CrossRef]
- Swallah, M.S.; Bondzie-Quaye, P.; Wu, Y.; Acheampong, A.; Sossah, F.L.; Elsherbiny, S.M.; Huang, Q. Therapeutic potential and nutritional significance of Ganoderma lucidum—A comprehensive review from 2010 to 2022. Food Funct. 2023, 14, 1812–1838. [Google Scholar] [CrossRef]
- López-Peña, D.; Gutiérrez, A.; Hernández-Navarro, E.; Valenzuela, R.; Esqueda, M. Diversidad y distribución de Ganoderma (Polyporales: Ganodermataceae) en Sonora, México. Bot. Sci. 2016, 94, 431–439. [Google Scholar] [CrossRef]
- López-Peña, D.; Samaniego-Rubiano, C.; Estrada-Morales, I.; Gutiérrez, A.; Gaitán-Hernández, R.; Esqueda, M. Morphological characteristics of wild and cultivated Ganoderma subincrustatum Murrill (Polyporales: Ganodermataceae) from Sonora, Mexico. Sci. Fungorum 2019, 49, e1213. [Google Scholar] [CrossRef]
- Xu, P.; Ding, Z.Y.; Qian, Z.; Zhao, C.X.; Zhang, K.C. Improved production of mycelial biomass and ganoderic acid by submerged culture of Ganoderma lucidum SB97 using complex media. Enzyme Microb. Technol. 2008, 42, 325–331. [Google Scholar] [CrossRef]
- Morales-Estrada, R.I.; Gaitán-Hernández, R.; Gutiérrez, A.; Vargas, G.; Jiménez, A.; Rascón, A.; Esqueda, M. Vineyard pruning waste improves bioconversion and chemical composition of native Ganoderma spp. (Agaricomycetes) strain from Mexico. Int. J. Med. Mushrooms 2018, 20, 775–789. [Google Scholar] [CrossRef] [PubMed]
- López-Romero, J.C.; González-Ríos, H.; Peña-Ramos, A.; Velazquez, C.; Navarro, M.; Robles-Zepeda, R.; Martínez-Benavidez, E.; Higuera-Ciapara, I.; Virués, C.; Olivares, J.L.; et al. Seasonal effect on the biological activities of Litsea glaucescens Kunth extracts. Evid. Based Complement. Altern. Med. 2018, 2018, 2738489. [Google Scholar] [CrossRef] [PubMed]
- Torres-Moreno, H.; Marcotullio, M.C.; Velázquez, C.; Ianni, F.; Garibay-Escobar, A.; Robles-Zepeda, R.E. Cucurbitacin IIb, a steroidal triterpene from Ibervillea sonorae induces antiproliferative and apoptotic effects on cervical and lung cancer cells. Steroids 2020, 157, 108597. [Google Scholar] [CrossRef]
- Müller, C.I.; Kumagai, T.; O’Kelly, J.; Seeram, N.P.; Heber, D.; Koeffler, H.P. Ganoderma lucidum causes apoptosis in leukemia, lymphoma, and multiple myeloma cells. Leuk. Res. 2006, 30, 841–848. [Google Scholar] [CrossRef]
- Mfotie, E.; Munvera, A.M.; Mkounga, P.; Nkengfack, A.E.; McGaw, L.J. Phytochemical analysis with free radical scavenging, nitric oxide inhibition and antiproliferative activity of Sarcocephalus pobeguinii extracts. BMC Complement. Altern. Med. 2017, 17, 199. [Google Scholar] [CrossRef]
- Zolj, S.; Smith, M.P.; Goines, J.C.; Ali, T.S.; Huff, M.O.; Robinson, D.L.; Lau, J.M. Antiproliferative effects of a triterpene-enriched extract from lingzhi or reishi medicinal mushroom, Ganoderma lucidum (Agaricomycetes), on human lung cancer cells. Int. J. Med. Mushrooms 2018, 20, 1173–1183. [Google Scholar] [CrossRef]
- Liu, Y.W.; Gao, J.L.; Guan, J.; Qian, Z.M.; Feng, K.; Li, S.P. Evaluation of antiproliferative activities and action mechanisms of extracts from two species of Ganoderma on tumor cell lines. J. Agric. Food Chem. 2009, 57, 3087–3093. [Google Scholar] [CrossRef]
- Saraste, A.; Pulkki, K. Morphologic and biochemical hallmarks of apoptosis. Cardiovasc. Res. 2000, 45, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Jang, K.J.; Han, M.H.; Lee, B.H.; Kim, B.W.; Kim, C.H.; Yoon, H.M.; Choi, Y.H. Induction of apoptosis by ethanol extracts of Ganoderma lucidum in human gastric carcinoma cells. J. Acupunct. Meridian Stud. 2010, 3, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Elkhateeb, W.A.; Zaghlol, G.M.; El-Garawani, I.M.; Ahmed, E.F.; Rateb, M.E.; Moneim, A.E.A. Ganoderma applanatum secondary metabolites induced apoptosis through different pathways: In vivo and in vitro anticancer studies. Biomed. Pharmacother. 2018, 101, 264–277. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.M.; Zhong, J.J. Ganoderic acid Mf and S induce mitochondria mediated apoptosis in human cervical carcinoma HeLa cells. Phytomedicine 2011, 18, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Kis, B.; Pavel, I.Z.; Haidu, D.; Ștefănuț, M.N.; Diaconeasa, Z.; Moacă, E.A.; Dehelean, C.A.; Șipos, S.; Ivan, A.; Danciu, C. Inorganic element determination of romanian Populus nigra L. buds extract and in vitro antiproliferative and pro-apoptotic evaluation on A549 human lung cancer cell line. Pharmaceutics 2021, 13, 986. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekhar, M.; Nayak, V.L.; Ramakrishna, S.; Mallavadhani, U.V. Novel triazole hybrids of myrrhanone C, a natural polypodane triterpene: Synthesis, cytotoxic activity and cell based studies. Eur. J. Med Chem. 2016, 14, 293–307. [Google Scholar] [CrossRef]
- Madasu, C.; Karri, S.; Sangaraju, R.; Sistla, R.; Uppuluri, M.V. Synthesis and biological evaluation of some novel 1,2,3-triazole hybrids of myrrhanone B isolated from Commiphora mukul gum resin: Identification of potent antiproliferative leads active against prostate cancer cells (PC-3). Eur. J. Med. Chem. 2020, 188, 111974. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, H.; Zhou, J.; Li, F.; Wang, J.; Chen, M.; Liu, Q. Cytotoxicity, DNA damage, and apoptosis induced by titanium dioxide nanoparticles in human non-small cell lung cancer A549 cells. Environ. Sci. Pollut. Res. Int. 2015, 22, 5519–5530. [Google Scholar] [CrossRef]
- Khan, I.; Bahuguna, A.; Bhardwaj, M.; Khaket, T.P.; Kang, S.C. Carvacrol nanoemulsion evokes cell cycle arrest, apoptosis induction and autophagy inhibition in doxorubicin resistant-A549 cell line. Artif. Cells Nanomed. Biotechnol. 2018, 46 (Suppl. S1), 664–675. [Google Scholar] [CrossRef]
- Rieger, A.M.; Nelson, K.L.; Konowalchuk, J.D.; Barreda, D.R. Modified annexin V/propidium iodide apoptosis assay for accurate assessment of cell death. J. Vis. Exp. 2011, 50, 2597. [Google Scholar] [CrossRef]
- Tang, W.; Liu, J.W.; Zhao, W.M.; Wei, D.Z.; Zhong, J.J. Ganoderic acid T from Ganoderma lucidum mycelia induces mitochondria mediated apoptosis in lung cancer cells. Life Sci. 2006, 80, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Li, S.; Zhuo, Y.; Chen, J.; Qin, X.; Guo, G. Anticancer effect of triterpenes from Ganoderma lucidum in human prostate cancer cells. Oncol. Lett. 2017, 14, 7467–7472. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Dai, L.; Wang, L.; Zhu, J. Ganoderic acid DM induces autophagic apoptosis in non-small cell lung cancer cells by inhibiting the PI3K/Akt/mTOR activity. Chem. Biol. Interact. 2020, 316, 108932. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shimizu, K.; Tanaka, A.; Shinobu, W.; Ohnuki, K.; Nakamura, T.; Kondo, R. Target proteins of ganoderic acid DM provides clues to various pharmacological mechanisms. Sci. Rep. 2012, 2, 905. [Google Scholar] [CrossRef]
- Zhang, Q.; Wu, L. In vitro and in vivo cardioprotective effects of curcumin against doxorubicin-induced cardiotoxicity: A systematic review. J. Oncol. 2022, 2022, 7277562. [Google Scholar] [CrossRef]
- Supasena, W.; Muangnoi, C.; Praengam, K.; Wui Wong, T.; Qiu, G.; Ye, S.; Wu, J.; Tanasupawat, S.; Rojsitthisak, P. Enhanced selective cytotoxicity of doxorubicin to breast cancer cells by methoxypolyethylene glycol conjugation via a novel beta-thiopropanamide linker. Eur. Polym. J. 2020, 141, 110056. [Google Scholar] [CrossRef]
- Osman, A.M.M.; Bayoumi, H.M.; Al-Harthi, S.E.; Damanhouri, Z.A.; ElShal, M.F. Modulation of doxorubicin cytotoxicity by resveratrol in a human breast cancer cell line. Cancer Cell. Int. 2012, 12, 47. [Google Scholar] [CrossRef]
- Torres-Moreno, H.; Marcotullio, M.C.; Velazquez, C.; Arenas-Luna, V.M.; Hernández-Gutiérrez, S.; Robles-Zepeda, R.E. Cucurbitacin IIb from Ibervillea sonorae induces apoptosis and cell cycle arrest via STAT3 inhibition. Anticancer Agents Med. Chem. 2020, 20, 1188–1196. [Google Scholar] [CrossRef]
- Ouyang, J.J.; Wang, Y.Q.; Tang, W. Ganoderic acid restores the sensitivity of multidrug resistance cancer cells to doxorubicin. Adv. Mater. Res. 2014, 834, 573–576. [Google Scholar] [CrossRef]
- Liang, C.; Tian, D.; Liu, Y.; Li, H.; Zhu, J.; Li, M.; Xin, M.; Xia, J. Review of the molecular mechanisms of Ganoderma lucidum triterpenoids: Ganoderic acids A, C2, D, F, DM, X and Y. Eur. J. Med. Chem. 2019, 174, 130–141. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).