Abstract
Bacillus thuringiensis (Bt) is a rod-shaped, Gram-positive soil bacterium that belongs to the phylum Firmicutes and the genus Bacillus. It is a spore-forming bacterium. During sporulation, it produces a wide range of crystalline proteins that are toxic to different orders of insects. Sporulation, structure assembly, and germination are essential stages in the cell cycle of B. thuringiensis. The majority of studies on these issues have focused on the model organism Bacillus subtilis, followed by Bacillus cereus and Bacillus anthracis. The machinery for sporulation and germination extrapolated to B. thuringiensis. However, in the light of recent findings concerning the role of the sporulation proteins (SPoVS), the germination receptors (Gr), and the cortical enzymes in Bt, the theory strengthened that conservation in sporulation, structure assembly, and germination programs drive the survival and success of B. thuringiensis in the environment and the insect host. In the present minireview, the latter pinpointed and reviewed.
1. Introduction
The phylum Firmicutes (now referred to by a new, name, Bacillota) includes known spore-forming bacteria of the genera Bacillus and Clostridium. The genus Bacillus includes to Bacillus cereus, Bacillus subtilis, Bacillus anthracis, Bacillus megaterium, and Bacillus thuringiensis. The majority of these are soil bacilli and have relevance at the level of the food industry, pathogenesis, biological weapons, and in biotechnology (nanotechnology, therapeutics) [1,2,3,4,5,6,7]. Meanwhile, the members of the genus Clostridium, such as Clostridium perfringes, Clostridium botulinicum, and Clostridium tetanus mainly have a role in food spoilage, food-borne disease, intoxication, gas gangrene, pseudomembranous colitis, botulism, human pathogenesis (toxin production), and in the biotechnological industry (chemical products) [8,9,10,11,12,13,14,15,16].
A feature shared between the genera Bacillus and Clostridium is the sporulation, structure assembly, and germination for survival and DNA protection [17,18]. The manner how they carry out these biological events at the molecular level is the subject of the present review, addressing general knowledge of the soil bacterium Bacillus thuringiensis and insight into the molecular programs that make this bacterium more than a successful insect pathogen in the environment and in the host [3,4,6].
Sporulation in the phylum Firmicutes plays a fundamental role as a cytological and morphological process during life cycle. Genes and proteins constitute players in spore formation and germination [10,17,19,20,21,22,23,24]. Currently, the high-throughput technologies, integrated as the omic technologies, should allow for deep insight into the unveiling of the complex machinery of the sporulation and germination of spore-forming bacteria [25,26].
The genes and the proteins for each stage are conserved among species of Bacillus. However, in Clostridium spp., there are some differences due to the environmental conditions vs. the soil rhizosphere [1,6,8,17,27,28,29,30]. Indeed, a recent study using 108 genomes of different genera of the phylum Firmicutes found that the majority of the bacilli share a core of at least 60 genes deriving from the 500 genes that participate in sporulation [24], while many spore coat proteins (small acid-soluble proteins) and germination proteins are not present in Clostridia and in some Bacilli. These shed light on the conservation, but also on the evolution and diversity, in both sporulation and germination in the phylum Firmicutes, thus implying a lifestyle shared between the genera Bacillus and Clostridium [2,24].
Spores are the forms most preserved, resistant, and propagated to the utmost degree in any geographical latitude [11,25,31]. Spores are highly resistant to extreme conditions, including physical conditions (such as temperature, pH, radiation), chemical conditions (salinity), or even biological conditions (selection pressure) [32]. Indeed, bacteria have survived under the conditions of an ancient primitive earth thanks to their ability to sporulate [11,24,31,33].
However, how do the spores permit the microorganism to survive and persist for long time periods? A cue is the structural architecture of the spore. Recent electron cryotomography (ECT) permits three-dimensional (3D) study reconstruction of the Gram-negative and Gram-positive bacterial cell walls. This analysis, in conjunction with biochemical and genetic evidence, supports the hypothesis that sporulation could be the ancient biological evolution process that gave rise to the second membrane in diderm cells (Gram-negative bacteria). The interconversion of the thin and thick peptidoglycan layer facilitated this process.
The second membrane in diderm bacteria is richer in lipopolysaccharides (LPS) and outer proteins. In other words, the chemical composition of the outer and inner membranes of the spore play a role in resistance and protection under harsh conditions. The dynamic of sporulating regulatory proteins, the morphogenetic coat, and other proteins are involved in the early, middle, and late stages in sporulation or in spore biogenesis [34]. On referring to Bacillus thuringiensis (Bt) and its remarkable soil life, there are thousands of studies regarding its mechanism of action and its biotechnological application as a bioinsecticide. However, Bt has a spectrum of action due greatly to the battery of proteins produced (ICPs) at the onset of sporulation. Recent works have revealed by combining proteomics and metabolomics that there is metabolic regulation mechanism of sporulation and ICPs synthesis. Specifically, these metabolic pathways are involved in synthesis, energy storage, carbon supply, and nutrients (amino acids, sugars), and these are under close regulation (transcriptional and translational) during sporulation and crystal synthesis [35,36]. Nonetheless, there are many questions to answer in terms of the molecular evolution and conservation of the core set of molecular components, the master germinating receptors (GR), the master SpoVA, hydrolytic enzymes, the master dipicolinic acid, import and export, gene–operon organization, biochemical composition, PG, the structural organization of the lipids of the layers between the spores, the germinating spore, and the vegetative cells [37,38]. This knowledge can impact many applications that range from immune stimulants, such as probiotics [39], to nanotechnology as the ideal and model vehicle for drug delivery and vaccines, and in evolutionary biology, developmental biology, and ecology [6,30,40]. No less relevant is that Bacillus thuringiensis is successful either outside or inside of its host (insects). Some authors suggest that there is a battle between Bt and the insects [6]. Others propose that there is a co-evolution [41] of bacteria–insects, referring in particular to the hundreds of Cry toxins that are produced concomitantly with sporulation [6,42,43], enabling these toxins with binding properties to insect midgut receptors. In nature, in the soil’s rhizosphere, there is a fruitful crosstalk among the bacterial community that involves competence for nutrients and for survival, in which the spore plays a primordial role.
2. The Soil Spore-Forming Bacterium Bacillus thuringiensis
The identity of Bacillus thuringiensis relies on a set of pore-forming proteins, known as Cry and Cyt toxins, to kill insect larvae. Therefore, Bt is considered an insect pathogen [41,42,44,45,46]. B. thuringiensis belongs to the genus Bacillus, a rod-shaped Gram-positive soil bacterium that contains genomic DNA and extrachromosomal DNA (plasmids). Interestingly, many plasmids encode the delta-endotoxins or Cry proteins, a strategy of B. thuringiensis to survive in the harsh environment of the soil’s rhizosphere and for insect and mammalian targeting [41,42,43,47]. Commitment in the life cycle of B. thuringiensis consists of a series of morphological and cytological changes that end with spore formation and crystal production. This series includes gene expression and biochemical and genetic programs [40,48]. Remarkably in B. thuringiensis, there is an arsenal (around six plasmids) of extrachromosomal DNA (pHT77 plasmids) encoding the delta-endotoxins or Cry proteins), the pAW63 plasmid harboring the RAP-PHR system [49,50,51,52,53,54,55], and also encoding another regulon system [56].
The Plasmid-Encoded Bt Crystalline Proteins
The insecticidal delta-endotoxins of Bacillus thuringiensis or Cry (Crystalline) proteins have been the subject of intense research during the last three or four decades [6,42,43]. These crystals comprise an array of immature protoxins with a molecular weight of 130 kDa, encoded in large plasmids [49,50,51]. To be active in the insect larvae host, protoxins are first solubilized and then processed in the C-terminal region favored by the enzymatic action, yielding a toxin with a molecular weight of 60–70 kDa [57,58]. The 3D structure of several Cry toxins was elucidated by X-ray resolution crystallography [59], including Cry1Ac [60], Cry2Aa [61], Cry3Aa [57], Cry3Ba [62], Cry4Aa [63], and Cry4Ba [64]. Moreover, based on sequence identity, it has been determined that the majority of the Cry toxins share three-domain structures with five highly conserved blocks in domain I [57,58]. Domain I is formed by a bundle of seven alpha-helices, with one central helix surrounded by the six other alpha helices [57]. The secondary structure of the alpha helices of domain I resemble bacterial pore-forming proteins, such as bacterial colicin I. Furthermore, the length of the helices of Cry domain I are sufficient to transverse cellular membranes [57,58]. Domain II is the most hypervariable region of the Cry toxins. The secondary topology is three anti-parallel β-sheets packed together in a β-prism with pseudo-three-fold symmetry [43,65,66,67]. Insect binding specificity is determined through the interaction of the loops of domain II and the receptors immersed in the insect’s midgut. Domain III forms a β-sandwich [57]. The latter is an arrangement of two anti-parallel β-sheets packed in a “jelly roll” topology. Specifically, in the case of Cry1Aa and Cry1Ac, a loop extension in Cry1Ac creates an N-acetygalactosamine (GalNAc) binding pocket implicated in receptor binding and further toxin proteolysis [68,69,70,71]. The action of the Cry toxins depends on the presence of a set of insect midgut receptors. This molecular crosstalk between insect receptors and the delta-endotoxins of B. thuringiensis strengthened a co-evolution theory for survival and success [3,6,41,72,73,74,75,76,77,78].
3. The General Sporulation Mechanism in the Genus Bacillus
The challenge and the goal objective of sporulation in the genus Bacillus and in other spore-forming bacteria such as Clostridium is DNA protection and survival [17,79,80]. The latter process is accomplished by the Firmicutes phylum, despite pressure selection, evolution, and diversity in the set of molecular components comprising the program and that crosstalk [24,79,81,82,83,84,85]
One of these systems is the Rap-Phr quorum sensing system, which regulates different bacterial processes, remarkably the commitment to sporulation in the Bacillus species [86,87,88,89,90,91,92,93]. How do Rap proteins act in sporulation? Rap proteins act as quorum sensors, forming a response regulator with a TPR (tetratricopeptide repeat) domain, a hydrophobic pocket able to bind the signaling peptide, thus inducing a conformational change and modulating regulator activities [91,94,95]. Therefore, RAP proteins act on phosphatases, an intermediary component of the sporulation phosphorelay system in Spo0F. Rap63 exhibited moderate activity during sporulation and is inhibited by the Phr63 peptide [48,96]. In Bacillus subtilis (frequently used as a model of the genus Bacillus), the starting sporulation programs is characterized by the phosphorylation of the master regulator Spo0A [24,34,97,98,99,100,101,102,103]. Across the genus Bacillus, the sporulation process is regulated by a cascade of sigma factors as follows: sigma F (σF); sigma E (σE): sigma G (σG), and sigma K (σK). Sigma factor K (σ(K) is a sigma factor conserved among the Bacillus genera, except in the genus Clostridium [24,34,48,103,104].
The sporulation program conserved among the members of the genus Bacillus comprises the following seven cytological and morphological changes [81,82,105] (Figure 1A): Stage 0 to Stage I, Axial filamentation; Stage II, Polar septum formation; Stage III, Forespore engulfment (ơF, ơE); Stage IV to Stage V, Cortex and coat assembly, and Stage VI to Stage VII, Spore maturation and mother-cell lysis. The morphological and cytological changes impaired in spoIID, spoIIM, and spoIIP mutants [85,106,107,108,109,110,111] and in the spoIIB–spoVG double mutant. However, the deletion of the spoVS gene, controlled by ơH, permitted to the spoIIB–spoVG double-mutant, to complete engulfment [100,101,112,113]. SpoVA proteins are involved in the uptake and release of nutrients from the core during the uptake of Ca2+ dipicolinic acid. The lytic enzymes SleB and CwlJ, found in bacilli, hydrolyze the spore cortex [48]. The spore is formed by an assembly process that involves a four-layer coat. The coating proteins described for Bacillus subtilis include cot, cot B, saf A, cot H, cot O, cot E, ger E, and cot E ger E [34] (Figure 1A). Assembly starts from the external outermost amorphous (crust) layer, followed by the rodlet, the honeycomb, the fibrous, and the nanodot particle layers, and finally, the undercoat/basement layer. Interestingly, under the exosporium of B. thuringiensis [114,115,116,117,118], a hexagonal honeycomb is exposed.
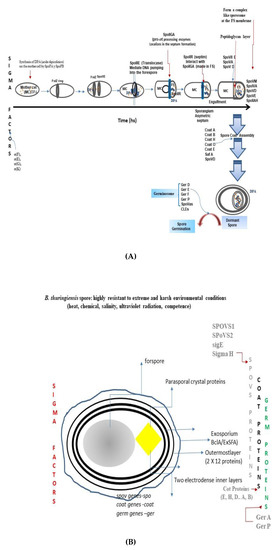
Figure 1.
(A) Sporulation, structure assembly, and germination in the genus Bacillus. Gram-positive spore-forming bacteria, Bacillus and Clostridium, follow similar morphological and cytological processes. There are some differences among members of the genus Clostridium. Sporulation program in the genus Bacillus is conserved. The spore allows us to fight against the selection pressure in the different niches and ecosystems. Therefore, the sporulation process is essential for resistance, survival, and success, and even to co-exist forever. Briefly, the sporulation process is a mechanism by which a set of sigma factors spov genes that encode the specific SPoVS proteins are involved in the regulation of the expression of the genes and proteins that accomplish each of the steps. A principal step in the sporulation of vegetative cells starts with the formation of septa (FTzS ring), followed by asymmetric division of the mother cell and the forespore, leading to the release of the forespore. (B) The components of the spore of B. thuringiensis are outlined, revealing the presence of the bipyramidal crystal (ICP) synthesized concomitantly with the sporulation. During the spore and structure assembly, there is the expression of several cot genes. These genes and their products play a role in the assemblage of the external and internal layers, similar to that of the bacteriophage T4. Moreover, some members of the genera Bacillus possess an exosporium, -an outer layer missing in B. subtilis- that confers protection and a direct connection with the environment. The expression and production of the insecticidal crystal proteins (ICP) (in yellow) are under the regulation of the sigma factors, and together with sporulation, both are under metabolic regulated mechanism at transcriptional and translated level. Some of the SpOV proteins also participate in crystal production. Thus, the spore of Bt is well-armored as an evolutive advantage for survival and success [54,55,114,115,116,117,118,119,120,121].
The lattice constant of the honeycomb structures was approximately nine nanometers (nm) for both B. cereus and B. thuringiensis spores, visualized using atomic force microscopy (AFM) by [117]. It was also possible to visualize the species-specific spore assembly and nanometer-scale structure of the spore’s surfaces. Ensamblage of the fibrous layer involves the Cot H- and Cot E-dependent proteins and the cot E-specific protein [110]; this is similar to the assembly of the spore-coating proteins, in that it mimics a non-mineral two-dimensional (2D) crystallization seeding pattern that begins to assemble the coating proteins from the inner to the outer layers in a similar manner as has been described for the bacteriophage lambda [122]. This assembly process is well characterized in B. subtilis (Figure 1A).
In bacterial-cell division, the structural and cytokinetic functions require the formation of the septum, which involves the assembly of a complex of proteins. Similar to B. subtilis sporulation, in B. thuringiensis sporulation, the sporulation-specific proteins Spo0A and SpoIIE play a role in gene regulation and in the determination of the structural properties of the specialized sporulation septum. Spore germination, nutrients, and mRNA number abundance participate, possibly providing ribonucleotides [123,124]. In B. thuringiensis, the mRNA number is 10–50-times higher than in other species of Bacillus and Clostridium [119]. How is the distribution found of mRNA in the spore compartments? A low abundance of mRNA is present in the mother cell and a high abundance of mRNA in the forespore (Figure 1A).
The transcription of these mRNA is under the control of the sigma factors F or G, and this can be similar among species of Bacillus. A minority of mRNA in the spores of these species is present at more than the molecule-per-spore, averaging only 6% of all individual mRNA identified in these spores. Thus, 94% of mRNA participates in the generation of proteins that will affect the germination of the whole spore [94,125]. The close relatives of B. subtilis, B. cereus, Bacillus anthracis, and Bacillus thuringiensis Al Hakam, as well as the spores of Bacillus megaterium and Clostridium difficile, lack several nucleotide biosynthetic enzymes, which are synthesized only at defined times in spore outgrowth [1,126,127,128].
The 60 most abundant mRNA in all five Bacillus species transcribed in the developing spore were found only in dormant species. Sigma E/K-dependent transcripts in spores might arise from weak–dependent transcription in the forespore of some of these genes [129]. A possibility could lie in the connection between the mother cell and the forespore, termed a feeding tube in the cytoplasm [124,130,131,132,133], which serves the mother cell and transfers small molecules, such as ATP and amino acids, into the developing spore. mRNA or mRNA fragments also move from the mother cell into the forespore via this feeding tube [134]. The precise time in sporulation at which the feeding tube closes occurs late in forespore development. Developing spores cannot make ribonucleotides, amino acids, or ATP, in that at least several TCA cycle enzymes are absent [1,126,134].
In referring to the structural assembly of the multilayered spore of the genus Bacillus, microscopy technology advancements permitted us to approach the spore structure assembly [135]. The structure assembly of the spore coat is accompanied by the synthesis of proteins that contribute to the multilayered structure. These proteins exert a strong influence on the core protection of the endospore, the maintenance of spore-core dehydration and dormancy, and survival in the environment, distribution, and conferring germination [127,136,137,138,139]. The cortex is synthesized within the intermembrane space surrounding the forespore after the engulfment stage during sporulation [123,124,140]. The proteins for cortex synthesis are produced in both the forespore and the mother-cell compartments. Peptidoglycan, lipids, and proteins (GerPA, GerP) (cortex lytic enzymes) form part of the outer coat, the inner coat, and the cortex, playing a structural and biochemical function. For example, in B. cereus, it has been shown that six GerP proteins share proximity with cortex–lytic enzymes in the inner coat [135].
3.1. Sporulation Program in B. thuringiensis
Recent work on B. thuringiensis sporulation has revealed the participation of several transcriptional factors. These factors determine the fate of vegetative cells in terms of sporulation, crystal formation, and germination [53,54,55,141,142,143]. As mentioned previously, sporulation in B. thuringiensis is highly conserved among the members of the genus Bacillus (B. subtlis, B. cereus). Indeed, knowledge of the sporulation program has been extrapolated from the model organism of the genus Bacillus, B. subtilis, and B. cereus [135]. The initiation of B. thuringiensis sporulation is controlled by the gradual increase in the activity of the master regulator Spo0A. This regulator is phosphorylated by the multicomponent phosphorelay system [55,121]. This system is usually affected by two negative-regulatory mechanisms: The Rap phosphatases involved in the first mechanism, active, and regulated by peptides derived from Phr proteins and followed by an export–import maturation process [55]. The second negative mechanism involves the Spo0E-P -B, -E, and -H) family of proteins, Spo0A-specific phosphatases, and small proteins (ranging between 50 and 90 amino acids), which share the conserved motif SQELD [55,121].
The transcription factor CpcR positively regulates a spo0E family gene, and variations in this gene expression modulate the production of spores in B. thuringiensis. Thus, there is a correlation between cpcr gene expression and sporulation. Indeed, one question to be analyzed is whether CpcR interacts with the network of the sporulation program to control cell differentiation [54,55,121]. Moreover, Spo0A activity impairs the presence of the cpcR in the LM1212 strain (double spore-forming population and crystal production). Furthermore, a putative phosphatase of the Spo0E family dephosphorylated and phosphorylated Spo0A-Spo0A-P [54,55,121]. This molecular strategy employed by Bt to control sporulation as a cell differentiation process aimed to ensure its survival in the environment and the host [55] (Figure 1A,B). Moreover, the crosstalk between the set of sigma factors (controlling spoVa genes) and the SPoVA proteins during the sporulation of Bacillus thuringiensis is concomitant with parasporal crystal formation [119]. Several studies determine which of these talk to which. In particular, mutants in the set of genes involved have demonstrated that the sigma factors (ơF, ơE) play a role in forespore engulfment (stage 3). Mutants in the gene spots controlled by sigma factor (ơH) are affected in terms of the efficiency of sporulation and polar septum formation (stage 2) [112,113,144,145,146].
The Sporulation Mechanism in B. thuringiensis, a Multistep Process
(1) Initiation of sporulation by the Rap-Phr system; (2) Commitment to sporulation regulated by the phosphorylation state of the major response regulator Spo0A [53,54,56]; (3) The different signals, such as nutritional deprivation, recognized by sporulation kinases [147]; (4) These kinases phosphorylate Spo0F, which is used as a substrate by the phosphotransferase Spo0B to phosphorylate Spo0A [88]; (5) Response regulator aspartate phosphatases (Rap) inhibit this signal transduction pathway by dephosphorylating the Spo0F-P response regulator [90]; (6) Rap protein activity inhibited by its related Phr peptide, and, (7) The mature Phr peptide of five, six, or seven amino acids [92,120,145,148] translated into a premature form that needs to be secreted, processed, and re-imported by oligopeptide permeases in order to be active [36,92,93,147].
Differences in the molecular mechanism of the cell’s fate affect the distribution of the SPoVA proteins in sporulating bacteria (Figure 1B). Recent studies employing double mutants and genomic studies reported that sporulating proteins are distributed according to their function and to the host environment [83,84]. The differences in the numbers of homologous genes lead to differences in the role of the spoVS genes in the B. cereus group and in B. subtilis. The spoVS gene found in B. thuringiensis, that is, two homologous genes spoVS1, and spoVS2, are both conserved in B. cereus [119]. Their expression is dependent on sigma factor H (ơH). Mutants in B. thuringiensis HD (ΔspoVS1) exhibited effects on morphological changes during sporulation [121,145], which include delay in sporulation efficiency, the formation of polar septa, and spore release. However, the mutants also failed to complete engulfment in some cells and demonstrated disporic septa [119]. Septal thinning and membrane migration play a role in forespore engulfment. The mutants also induced the decreased production of the parasporal crystal Cry1Ac [119]. Several molecular components are involved. For example, a sporulation-specific cwlC gene that encodes an N-acetyl muramyl-alanine amidase characterized in the B. thuringiensis subsp. israelensis (Bti) strain Bt-59. CwlC was the only cell-wall hydrolase in Bti found to contain both MurNAc-LAA and Amidase 02C domains [149]. Moreover, and according to transcriptional analyses, cwlC was expressed at the late sporulation stage and was controlled by SigK. In addition, two other cell-wall hydrolase genes were identified with high expression levels, e.g., the cwlB and cwlE genes controlled by SigK. In contrast, another hydrolase encoded by the cwlF gene is not under the control of the SigK factor; however, this gene is plasmid-encoded [149]. Another molecular component is SpoIIID, a small, sequence-specific DNA-binding protein that can direct the transcription of many genes, and it affects spore formation in B. subtilis and B. thuringiensis. The SpoIIID mutant strain exhibited no mother-cell lysis in the Schaeffer sporulation medium (SSM), but did in Luria-Bertani (LB) medium. The deletion of spoIIID decreased crystal protein production in HD73. Furthermore, SpoIIID positively regulated the sigK gene, while sigma factor K (σK) negatively regulated the expression of sigE [150].
On the other hand, during parasporal crystal formation, the genes cry1A (Bravo et al., 1996) [145], cry4A [151], cry8E [152], and cry11A are controlled by ơE or ơK [147,153]. The sporulating-specific transcription factors, regulating toxins such as Spo9A, can positively regulate cry1Ac [146,154]. Thanks to the progressive advances in genomics, it has been possible to determine the differences in the functions of sporulation-related genes and non-sporulating genes in the genus Bacillus, including B. cereus, and B. subtilis [35,36,154]. B. thuringiensis possesses the PlcR regulon linked to endotoxin production [80]. Bt Cry toxins are considered nonpathogenic in humans, while Cyt (cytolytic) proteins cause damage to mammalian cells [155]. Cry toxins represent nearly 20–30% of the cell’s dry weight. During sporulation, crystal proteins are localized on the spore surface, decreasing spore resistance, but increasing the insecticidal properties of the crystals [156]. The expression of the protoxin gene is controlled by sporulation-dependent promoters. All of the protoxins in B. thuringiensis subsp. israelensis and other genes or unknown functions encoded in a large plasmid (128 kb) [143,157]. Transcription depends on the mother cell at the middle stages of sporulation, mostly transcribed from the promoter cry4A-P1 and under the control of sigma 35, which is highly homologous and functions similarly in B. subtilis [153]. Cry (crystal), and Cyt (cytolytic) toxins are synthesized during sporulation and assembled into one or more crystalline parasporal bodies [158,159,160]. Several studies on B. thuringiensis subsp. kurstaki (HD1) has shown that the three Cry1A proteins co-crystallize to form a single bipyramidal crystal [161]. Even more so, during sporulation, the Cry2A inclusion assembles with and is partially embedded in the short axis of the bipyramidal crystal [162]. Remarkably, in mosquitocidal isolates of B thuringiensis, parasporal- body structures exhibit greater complexity [163,164,165]. Recent works have described Cry and Cyt inclusions as being bound together by a peripheral multilaminate fibrous matrix of unknown composition [110,117,160]. Therefore, there is a coordination of the synthesis and assembly of several protoxins and their association with the parasporal-body fibrous matrix during the synthesis of other proteins of the plasmid, suggesting that the latter are in the parasporal body [115,116,118,158,159,160]. Moreover, the genes ssrSA and ssrSB in the same operon are co-transcribed as a precursor, processed by specific ribonucleases to form mature 6S-1 and 6S-2 RNA. These non-coding RNA play a role in sporulation and parasporal crystal formation because the deletion of these genes inhibited the growth of B. thuringiensis in the stationary phase [86,96,97,98,99,102]. In the soil, Bt Cry toxins on the spore surface are protected by the exosporium. In the midgut, they are exposed and available for binding to the insect receptors [41,43,47,119]. Furthermore, studies on the Bacillus anthracis group (B. cereus and B. thuringiensis) [114] have shown that BclA, EsSY, and ExSFA are protein of the basal layer of the exosporium (the outermost external layer), serving as protection, host targeting, and dissemination) (Figure 1B). The underlying molecular mechanisms through which Cry and Cyt proteins are targeted to the parasporal-body matrix or the manner in which the structural integrity of this matrix is maintained is an issue that remains to be defined.
3.2. Routing the Regulatory Metabolism at Transcriptional and Translational Level for Sporulation and Insecticidal Crystal Protein Synthesis (ICPs) Production
To the myriad associated proteins produced for sporulation, high expression level of genes is required for insecticidal crystal proteins synthesis (ICPs). Moreover, sporulation and ICPs are energy-consuming, and require higher expression levels of genes to perform these biological processes. At this point, in the CT-43 strain chromosome, the metabolism of amino acid, carbon, and energy resources are in operons under systematic and regulated coordination at a transcriptional and translational level for routing the metabolism for sporulation and ICPs synthesis [35,36]. Thus, genes organized in operons and genes were either induced or up-regulated in response to amino acid starvation during sporulation. Thus, for the ICPs synthesis, a provision or supply of amino acids are necessary and a prerequisite for crystal protein production. More than 300 genes are involved in the aminoacid metabolism (KEGG base data). Previous isotopic work reported that 80% of amino acids for ICP synthesis came from protein turnover [166]. In B. thuringiensis, the amino acids are encoded in genes that conform to different operons, carrying thus complete biosynthetic pathways for the most common amino acids (n = 20). Interestingly, radioisotopic experiments have shown that amino acid supply for sporulation and ICP synthesis comes from protein recycling. A set of proteases, ATP proteases, regulatory proteins are transcriptionally up-regulated (Figure 2) and translational up or downregulated, which allow protein recycling to meet amino acid requirements during sporulation: a set of proteases (proteases, peptidases, and some ATP-dependent proteases) with high expression levels rapidly degrade many abnormal polypeptides. Other proteases exerting proteolysis functions are induced or up-regulated at the transcriptional level YabG (sporulation-specific proteases), CH1854 (intracellular serine protease), and CH3928 (serine protease), NprB (bacillolysin), CalY (camelysin), thermitase (thermostable serine protease), and Vpr (a high-molecular-mass minor extracellular protease) [167,168].
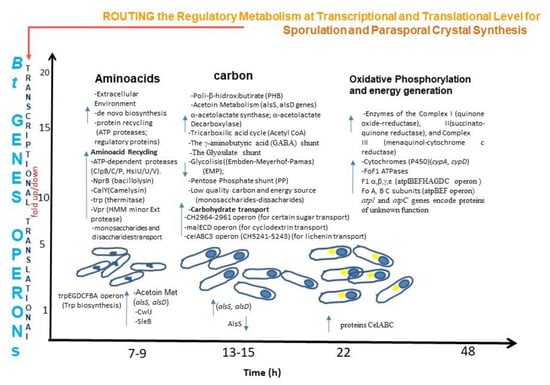
Figure 2.
A myriad of genes is associated with the different biological processes in bacteria. Sporulation and crystal synthesis require routing the regulatory metabolism (amino acid biosynthesis, amino acid transport, carbon supply, carbohydrate transport, and energy generation) at transcriptional (fold-up/down) and translational levels (amount of protein). How the different nutrient sources are routed and regulated for cell growth requires high expression levels of genes. At 7 to 8 h, many genes encoding proteases are induced, and they have functions such as transport and carbohydrate conversion for energy supply. While at 13 and 22 h, some other proteins start to increase and decrease other proteins required for the subsequent step in sporulation and crystal formation. The routing of the regulatory metabolism is quite complex. It is an up and down fold regulation as well as an increase-decrease production of proteins that successfully each cell growth accomplish this task efficiently. It seems that B. thuringiensis has evolved strategies to save energy and to profit from external nutrient sources [35,36].
Regulatory proteins control protein quality and regulate many biological processes [169,170]. Moreover, these proteins provide a large number of amino acids. In this way, protein recycling would be the source of amino acids during sporulation (Figure 2). Studies in the CT-43 strains using RNA-seq and bioinformatics have revealed that during nutrient starvation, genes and operons are induced and upregulated [171]. However, when the nutrient is rich, cells store intracellular (PHB) and extracellular (Acetoin) carbon molecules recycled under nutrient-depletion conditions. Indeed, some low-quality carbons as monosaccharides and disaccharides remained unused during exponential growth and fully utilized during sporulation and ICP synthesis.
Carbon and energy requirements Bacillus have evolved strategies to store and supply carbon and energy to carry sporulation and ICPs. Among these strategies for carbon supply are production and PHB reuse. PHB is produced as an intracellular carbon and energy storage substance in bacteria [172,173], and found a linear correlation between the final ICP and PHB concentration [172]. In the experiments performed by Wang et al., 2013 [36] with the CT-43 strain, they found that the intracellular PHB level started to increase rapidly, reaching a maximum level at 17 h, followed by a rapid decrease. PHB granules were visible by phase contrast microscope in some sporulating cells even at 15 h. Moreover, studies using RNA-seq [36] showed that most PHB synthesis-associated genes were highly expressed at 7 and 9 h and drastically reduced expression levels at 13 h. Meanwhile, PHB degradation-associated genes such as pcaD (phaZ), scoT, and phbA1 increased at 13 h. At the translational level, PhbB protein decreased production at 13 h, whereas PhbA, PhbB, and PhaC dramatically down-regulated at 22 h. Conversely, the proteins associated with the PNB degradation pathway, the protein PcaD (PhaZ), increased at 13 h. Of relevance is that PhaP and PhaQ involved in the assembly and disassembly of PHB granules maintained high-level expression at both transcriptional and translational (Figure 2). These studies indicate that the PHB regulatory metabolism (synthesis and degradation) at these levels play a role as an important carbon source in sporulation and parasporal crystal formation. Another potential alternative for carbon sources is the Pentose Phosphate (PP) shunt. All the participating enzymes of this pathway were identified by iTRAQ [36], except Zwf, glucose-6-phosphate 1-dehydrogenase) but remained almost unchanged during sporulation, strengthening the suggestion that the PP pathway is not involved at all in providing the reducing power (NADPH) and metabolic intermediates involved in many biosynthetic processes. In the PP pathway, there are three alternatives or routes that could be involved during sporulation and ICP synthesis. The predominant route to arrive at the nodal point is gluconate-6p is: (1) glucose converted into glucose-6p; (2) glucose 6p is converted into Glucono-1, 5-lactone-6p by Zwf; and (3) the intermediate further transformed into gluconate-6p by 6-phosphogluconolactonase (CH3298). An alternative route is for glucose to be converted into gluconate by Gdh (glucose 1-dehydrogenase), and gluconate catalyzed into gluconate-6p by GntK (gluconokinase). The key limiting enzymes, Zwf and Gdh in both routes were not detected during the exponential growth phase (7 h) at both the transcriptional and translational levels [36]. However, the gene zwf was slightly induced at 9 h and then up-regulated at 13 h, while the gene gdh was initially induced at 13 h at the transcriptional level. This data would imply a very significant regulatory mechanism of the PP pathway that could have a role as alternatives strategies during biological processes as the ones mentioned above. Interestingly, when CT-43 cells were grown in a GYS medium containing yeast extract, they followed a third route on the gnt operon that participates in gluconate metabolism. This gnt operon (CH2189–2191) is composed of gntP (gluconate permease), gntK, and gntZ/gndA (6-phosphogluconate dehydrogenase), and it lacks the transcriptional regulator, which expression is comparable to the negative regulator gntR described in B. subtilis [174]. The data of RNA-seq obtained by Wang et al., 2013 showed that the gnt operon expression level reached a maximum at seven hours and then gradually decreased. Furthermore, for bacteria, direct uptake of substances from the extracellular environment might be the most rapid and metabolically economical pathway. Therefore, extracellular gluconate is likely transported into the cells directly by GntP and converted into gluconate-6p by GntK, with gluconate-6p further catalyzed into ribulose-5p by GntZ/GndA, leading, to the repression of the genes zwf (glucose-6-phosphate 1-dehydrogenase) and gdh (glucose 1-dehydrogenase). Therefore, glucose does not enter the PP pathway when the extracellular environment contains gluconate, which might explain why 100% glucose catabolism was through the EMP pathway. The contribution of the PP pathway was still 5% in a glucose-glutamate-salts medium [175]. The low-quality carbon sources such as monosaccharides, disaccharides -glucosamine from chitin degradation and deacetylation, and glucose form lichenin cleavage [176,177] participate in energy metabolism and amino acid biosynthesis (particularly the branched-chain amino acids)(BCAAs) during sporulation. Furthermore, glucose and other monosaccharides (low-quality carbon sources) provided by the GYS medium could be exhausted during the exponential growth phase. They can enter the EMP pathway during sporulation to produce a large amount of pyruvate, which is also used for the high-level synthesis of dipicolinic acid (approx 25% of sporal core dry weight), a molecular component that plays a role in spore germination and resistance [178] (Figure 2). Fatty acids, b-oxidation, and the C2 and C4 compounds, the PHB (p-hydroxy-b-butyric acid) assembly and disassembly (PHB depolymerase) [179], (7) the intermediate of the TCA cycle (tricarboxylic acid), as acetyl-CoA utilized and remarkably upregulated during sporulation and ICP synthesis.
The TCA Cycle—Of relevance is that any mutant defective in the first three enzymes of the TCA cycle fails to express early sporulation genes suggesting that the activities of these enzymes are critical for sporulation [180,181]. On the contrary, a-ketoglutarate dehydrogenase, which catalyzes the fourth step of the TCA cycle, is not essential [182]. Remarkably, during sporulation, a considerable amount of acetyl-CoA is generated by pyruvate dehydrogenation, fatty acid b-oxidation, and the reuse of acetoin and PHB. It seems that acetyl-CoA would mainly flow into the TCA cycle to yield energy. The data reported by Wang et al., 2013 [36] along with other studies, have speculated that the TCA cycle is significantly modified or supplemented during sporulation via:
The glyoxylate shunt bypasses a portion of the TCA cycle to convert isocitrate to malate [183]. At transcriptional level, two glyoxylate shunt-specific genes; aceA (isocitrate lyase) and aceB (malate synthase) were up-regulated at 13 h. While at translational level, AceB production increased at 13 h and 22 h, respectively. This result implies that the glyoxylate shunt became more active during sporulation (Figure 2).
The γ-aminobutyric acid (GABA) shunt is an additional routing for the TCA cycle and is correlated with spore and parasporal crystal formation in B-thuringiensis [182,184]. GABA synthesis is through glutamate decarboxylation catalyzed by glutamate decarboxylase. Remarkably the sole glutamate decarboxylase GadB (CH2716) identified in CT-43 was not expressed at any phase at either the mRNA or protein level. Indeed GABA production was relatively weak in Bacillus strains [185]. However, the mRNA of GABA-specific permease gabP increased at 13 h, in agreement with an observation that gabP activated during nitrogen-limited growth [185]. Moreover, the GABA degradation-associated enzymes GabD (succinate-semialdehyde dehydrogenase) and GabT (4-aminobutyrate aminotransferase) were transcriptionally and translational up-regulated at 13 h, respectively. These results suggest GABA metabolism became more active during sporulation and that the utilized GABA might mainly come from the extracellular environment. Furthermore, the GABA shunt and the methyl citrate cycle are interconnected through a common node, the succinate, leading, thus, to accurate PHB reuse. Indeed, SucC and Suc D proteins were slightly decreased at 13 h, possibly implying that a significant amount of succinate is converted into succinyl-CoA during sporulation. Data from B cereus and with CT-43 strains indicated a significant increase in the levels of enzymes and cytochromes involved in energy production via the electron transport system during the transition from vegetative cells to spores [186,187]. Thus, at the transcriptional level, two cytochrome P450 genes [186], cypA and cypC, were markedly up-regulated at 13 h, and a dNADPH-cytochrome P450 reductase gene cypD induced at 13 h. (Figure 2).
Oxidative phosphorylation and energy generation through the FoF1-ATPase (ATP synthase) complex which catalyzes ATP synthesis from ADP and Pi, driven by the proton gradient generated by the respiratory chain, and organized in the operon atplBEFHAGDC [187,188,189]. The atpHAGDC encodes the γ, α, β, and ε subunits of the F1 portion, while the atpBEF operon encodes the A, C, and B subunits of the Fo portion. The atpl encodes a protein with an unknown function. The genes of these operons are regulated at the transcriptional and translational levels [36]. Thus, the atpC gene was down-regulated by more than 20-fold, and the others were decreased by about 2-5 fold during sporulation. Besides, increased expression of the enzymes that form part of Complex I, II, and III (Figure 2). In contrast, at the translational level, the g, a, b, and e subunits of the F1 portion and the C and B subunits of the Fo portion were all maintained at similar levels at 13 h and increased by more than 1.8 fold at 22 h. However, the subunit of the Fo portion failed to be quantified. These data highlight the high energy requirements of spore and parasporal crystal formation. Indeed, as outlined in Figure 2, there is a subtle regulation of the metabolism (amino acids, carbon, and energy) at the transcriptional, translational level that allows Bt under nutrient depletion, starvation can perform biological processes such as sporulation, ICP synthesis, and especially those involved in the dipicolinic acid, a molecular component of the spore core [35,36] (Figure 2).
4. The General Spore Germination Program in the Genus Bacillus
In spore germination, molecular and morphogenetic changes are carried out as crosstalk among signals, germinant nutrients, and spore components in the committed endospore in order to awaken or break dormancy [190]. For an endospore, the fate and the decision to germinate encodes in the dormant spore. The program of spore germination refers to the multistep mechanism through which spores return to life, an awakening process that enables them to reenter into metabolic activity [190,191,192,193,194]. The knowledge of the components and signals in spore germination derives from studies of the model organism Bacillus subtilis [11,111,195,196]. However, there are current efforts to update and focus on other members of the genus Bacillus, especially those that constitute a problem in pathogenesis, health, agriculture, and in the food industry as well [39,194,197].
Which are the signals that initiate the process of awakening the dormant spore? What is known is that external signals (germinants, small molecules) that sense germination-specific proteins (GR)-like receptors localize in the outer and the inner coat of the spore [127,197,198,199,200,201,202]. The phenotypical characteristic of the spore-germination stage is considered a weak stage or spot in the life cycle of Bacilli species. During this stage, the spores become susceptible to physical, chemical, and environmental conditions, starting from the inner membrane coat of the spore, to render these accessible to nutrients of low molecular weight, ions, nutrients, Dodecylamine, and water, in order to flow through the core cortex [195,198,203,204,205]. Specifically, in vivo spore germination in B.thuringiensis is carried out and favored under the alkaline and midgut-larval environment [140,141,142]. In vitro germination is activated by germinants (Dodecylamine, amino acids) in the culture medium and temperature growth at 25–27 °C in the medium [143,148,150,198]. The physical factors, such as temperature and pH, can aid the proteins of the coat’s inner membrane (IM) in undergoing the conformational changes that leave them in the appropriate conformation for interactions. This set of proteins comprises the so-called germinosome, including the GR proteins (particularly GERD), SpoVA proteins. GerP proteins favored the nutrient’s access to the inner membrane [202,206,207,208,209,210,211,212]. SpoVA spore components are present during sporulation and germination events [193]. Moreover, to awaken the dormant endospore is to favor inner membrane (IM) structure permeability through the expression of the proteins (SpoVA) that form the channels (seven in B.subtilis spores) [203,213] for the movement, passage, and release of monovalent cations, including H+, K+, and Na+ CaDPA (sodium calcium dipicolinic acid) [203,210,214,215,216,217,218]. These are proteins that take part in the release of DPA through channels encoded in one or more operons in all spore-forming bacilli [203,214,215,216,219,220]. In the particular case of B. subtilis, it is a heptacistronic operon. The mutations in the ger P locus cause a reduction in the permeability of the spore coat to germinant molecules [221]. In general, at least three proteins are present: SpoVAC; SpVAD, and SpoVAEb [193,222,223,224]. In B.thuringiensis, these proteins are organized as a biscistronic operon [225]. Once CaDPA release is complete in stage I, it triggers entry into stage II. In this latter stage, the expression and function of the molecular enzymatic machinery represented by the lytic transglycosylases (CLE, CwlJ, and SleB) [203,226,227], inactive in the dormant spore and recognize the muramic-δ-lactam modification present in the cortex. Furthermore, for in vitro spore germination, enzymatic treatment with the lysozyme removes the outer membrane and the spore-coat proteins. The concentration or amount of this enzyme is optimized to avoid damage to the incipient germinant spore. The enzymatic treatment renders the ions and nutrients permeable to the external and inner membranes, especially if they are low-molecular-weight germinant inducers. The cortex–lytic enzymes (CLE, CWlj, and SleB) should be present for the necessary cleavages and to permit the exchange of components such as dipicolinic acid (DPA). CLE can degrade the large peptidoglycan cortex layer, favoring the entry of ions and water, leading to spore-core swelling. Thus, the spore becomes a growing cell with restored metabolic activity [198,203,204,228,229] (Figure 3).
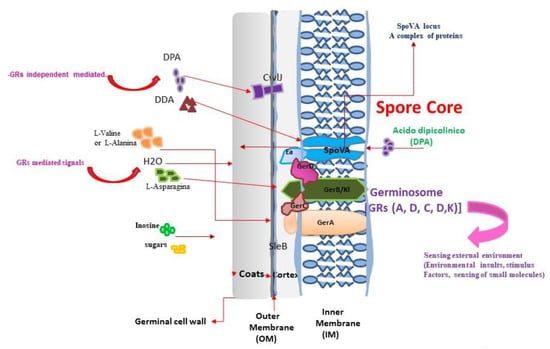
Figure 3.
Scheme of the dynamic interaction and cooperation between germinant receptors (GR), and the SpoVA proteins during the spore-germination in the genus Bacillus. For the dormant spore to wake up, start: 1, Sensing of the nutrients (aminoacids, sugars) or external stimuli from the environment or the host by the germinant receptor (GR), clustered in like germinosome (A, D, C, D, K) Schem) in the IM or other molecular components (CWLJ) in the outer membrane; 2. Interaction and cooperation between GRs, and a cooperation with SpoVA proteins; 3,Transmission downstream of the external signals, 4, Activation of the release of dipicolinic acid (DPA), and 5, Initiation of the germination process [37].
The general mechanism of spore germination can be outlined as follows:
(1) Germinant sensing; (2) Commitment to germinate; (3) Release of spore depot of dipicolinic acid (DPA); (4) Hydrolysis of peptidoglycan cortex spores; (5) Spore-core swelling and water uptake; (6) Cell-wall peptidoglycan remodeling, and (7) Restoration of core protein and inner-spore membrane–lipid mobility. This mechanism resembles a detailed general program for spore germination that is well characterized in B. subtilis, B. anthracis, and B. cereus. In B. thuringiensis, the differences in the molecular components that are involved in these processes are yet to be defined. One of the unsolved questions is related to the import and export of DPA and how the nutrients are sensed in the commitment spore [194]. Furthermore, the evolution, conservation, and diversity in the machinery of germination programs among the members of the genera Bacillus and Clostridium have recently approximated by genome sequencing [193,230,231]. It would appear that DPA is the master component whose movement determines the sporulation or germination of a vegetative cell. Indeed, in a recent study using B. subtilis, it was suggested that the subunits of the SpoVa proteins that form channels and cytoplasmic plugs play a role in the efficiency of DPA import (sporulation) and export (germination) [37,38,135]. DPA transport into the spores involves cycles of unplugging and then replugging the C-Eb membrane channel. Nutrient detection during germination triggers DPA release by unplugging the C-Eb [37,38,135] (Figure 3).
Detailed program for spore germination in which all of the molecular components are outlined: (1) Activation. Nutrient germinant plus spores (minutes to hours). Lag phase, and (2) Commitment (a major change in IM permeability and structure). GERP proteins allow the access of nutrients into the inner membrane, low-molecular-weight, i.e., Dodecylamine, ions (H+, Na+, and K+). Channel formation by the multiple spore-specific SpoVA) (n = 7) in Bacillus subtilis (Setlow and Christie., 2020; [127,198,230,232]; (3) Release of pyridine-2, 6-dicarboxylic acid (dipicolinic acid [DPA]) chelated at 1:1 with divalent cations, predominantly Calcium (Ca2+DPA) through the IM channels; (4) Enzymatic lysozyme-mediated cleavage of the cortex, favoring permeability into small molecules in the inner coat, triggering spore germination; (5) Stage I. All of the Ca2+ DPA is released by the CLE cortex degradation, and this event leads to passage into stage II [208], and 6. Stage II cortex degradation is complete. The germ-cell wall and the core take up water and expand. This marks the initiation of germination, giving rise to growing cells and to the activation of metabolic activity [195,224].
How is this program conducted? A strategy for success in the awakening of the spores is to produce effective dynamic interaction between the sensor and the nutrients. The termination “ome” refers to a set of clustered elements that performs an integrated function. Therefore, in spore germination, there is recent evidence of the local clustering of receptors in a germinosome in the IM of B. subtilis [233,234], and B. cereus [37,219]. Moreover, in some points, B.thuringiensis form foci as a complex of germinant receptors (GR) known as germinosomes [119,227,235,236,237]. These include the SpoVA proteins, involved in the uptake of Ca2+-dipicolinic acid into the forespore during sporulation, which are also responsible for its release during germination. Lytic enzymes SleB and CwlJ, found in bacilli and in some clostridia, this enzymes hydrolyze the spore cortex; other clostridia use SleC for this purpose. The diversity in the machinery of germination programs conformed by proteins. SpoVA, enzymatic, and the GR between the genera Bacillus and Clostridium have been approached by means of genome sequencing [193]. To understand how the germinated receptors (GR) integrate and transmit the downstream signal to awaken the dormant spores, one alternative is that receptors cluster in the germinosome to allow interaction with each other and with SPoVA proteins at the inner membrane (IM). Thus, germinants reach the GR after passing the outer membrane and interact specifically with GR, which respond to this and transmit to the SPoVA proteins [238] (Figure 3). Despite this, some germinants can interact directly with SPoVA independently of the GR, such as Dodecylamine (DDA) (Figure 3). In the cooperative interactions between the GR and the nutrients, the subunit secondary structure of the GR plays a role. For example, it is known that GR are commonly formed of A, B, and C subunits, encoded by tricistronic ger operons, as is the case for B. subtilis. In the spores of B. subtilis, GerA is the major GR and it has the following three subunits: GerAA; GerAB, and GerAC. The L-alanine activation of GerA requires all three subunits. Prediction studies have revealed that the secondary structure of GerAB is an alpha-helical transmembrane protein that can form water channels. Furthermore, molecular simulation studies have revealed that L-alanine can bind transiently to specific sites on GerAB [234]. In addition, the B. licheniformis genome contains the gerA family operons gerA, ynd, and gerK, in contrast to the ABC (D) organization, which characterizes the gerA operons of many Bacillus species. Indeed, B. licheniformis genomes contain a pentacistronic ynd operon, the yndD, yndE3, yndE2, yndF1, and yndE1 genes, encoding A, B, B, C, and B GR subunits, respectively (subscripts indicate paralogs). In contrast to the B subunits of Ynd, the B subunit of GerA was essential for amino-acid-induced germination [220,235,239,240]. Therefore, in addition to the operon organization of the GR receptors, dynamic structural studies have provided novel insights into the role of individual GR subunits involved in the cooperative interaction among GR, in triggering spore germination [220,235,239,240] (Figure 3). As outlined in Figure 1A,B, dormant spores can awake in several ways: (1) by external stimuli, external environmental signals; (2) in the host by nutrients; (3) by germinants receptors (GR) present in the inner membrane or in the outer membrane; (4) by other components such as CwLJ, or directly signal through SPoVA (independently of GR). In addition, the cooperative interactions of the GR (A, B, C, D), or in some cases, Ynd and GerK components which drive signalization with SPoVAC. This activation can initiate germination by release of DPA through the SPoVA channel, which is a central locus of a set of proteins, key in the import and export of DPA (Figure 3).
The Molecular Components of the Germination of B. thuringiensis
The molecular components of the germination of B. thuringiensis that are known are: (1) The enzyme Alanine racemase (AlrA), encoded by the gene homologous to the major component of the exosporium of B. cereus spores, alrA, which plays a crucial role in moderating the germination rate of B. thuringiensis spores [235]. Converted D-alanine into L-alanine, and this in turn stimulated spore germination in B. thuringiensis. This gene, cloned from B.thuringiensis subsp. kurstaki [235], is transcribed only in the sporulating cells. (2) The CLE and spore cortex–lytic enzymes are essential for germination in bacilli. A homolog of the cwlJ gene involved in spore germination was isolated from B.thuringiensis. The deduced product of this gene exhibits striking sequence similarity to the lytic enzyme, CwlJ of B. subtilis. Another open reading frame (ORF), 27 bp downstream of cwlJ, which deduced the product, shows high similarity to the YwdL of B. subtilis. Reverse transcription-polymerase-chain reaction (RT-PCR analysis indicated that cwlJ and ywdL formed a bicistronic operon in B. thuringiensis [225]. In addition, a gene-encoding spore, the cortex–lytic enzyme, designated sleB, was cloned from B. thuringiensis. The disruption of sleB did not affect the vegetative growth of B. thuringiensis. However, the fall in optical density (OD) (600 nm) in the mutant spores was much slower than in the wild-type strain during the spore germination induced by L-alanine [236] (3). The spore germination of Bacillus can start by sensing germinants such as L-alanine and by binding to specific receptors. The GerA receptor responds to L-alanine in B. subtilis. A homologous gerA operon of B. subtilis was isolated from B. thuringiensis subsp. kurstaki. Disruption of the gerA operon led to blockage of the L-alanine-initiated germination pathway and revealed a delayed inosine-induced germination response. The germination rate of the gerA complementary- strain spore deriving from the introduction of the gerA operon into the disruption mutant was even faster than that of the wild-type-strain spore [237]. Moreover, one polysaccharide deacetylase gene was cloned from B. thuringiensis and was designated pdaA. Disruption of pdaA did not affect vegetative growth and sporulation, but did affect spore germination [236]. (4) gerM is a very conservative gene of 4.5 kilobase (kb). A DNA fragment cloned from the partial DNA library of B. thuringiensis subsp. kurstaki 1.175. Sequence analysis showed that the fragment contains one complete ORF that encodes a 349-amino- acid (aa) protein, which has high homology with the GerM protein from B. subtilis. (5) The function of the gerM gene in Bt spores was analyzed utilizing gene disruption. The resulting germ mutant grows, sporulates and germinates slower than wild-type Bt spores in the presence of L-alanine or inosine. This result suggested that gerM expression is required for the normal germination initiated by L-alanine or inosine in B. thuringiensis [119,236,237] (Figure 3).
5. Implication of the Knowledge of Sporulation Structural Assembly and Germination in the Soil Bacterium B. thuringiensis
The stable and resistant nature of spores and the possibility of germinating and growing in a gut environment render them suitable for treatment in the form of probiotics and as vehicles for vaccine and drug delivery. Spore treatments have shown great promise in animal studies. However, human trials require going further. Nonetheless, spores might open the door to safe, effective, and easy-to-administer therapeutics [39,85,241]. It is pivotal to elucidate and understand the life cycle of spore-forming bacteria, especially those bacilli that threaten agriculture, the food industry, and health care [18,240,242]. Bt has become a promising and potential new avenue of alternatives against the biological control of insects and the application of biotechnology biomedicine. Furthermore, Bacillus thuringiensis is viewed as a biofactory for the production of proteins, but also of other products for bioremediation and for improvement as bioinsecticides. Moreover, arasporal delta endotoxins are highly specific against different orders of insects. However, Cyt proteins can exert a cytopathic effect on mammalian cells, specifically those changed by some types of cancer. This double sword of Bt marked the biotechnological success of B. thuringiensis; due to the versatility of Bt, great interest has emerged during the last two decades [3,4,42,119]. Work in this area ranges from basic research (mechanism of toxicity in insects) to applied science (the genetic engineering of economic crops with cry genes), the assembly of proteins for crystal formation (structural biology), and nanotechnology (drug-vehicle delivery or vehicles of subunit vaccines) [243,244,245].Furthermore, B. thuringiensis can produce floating biofilms with a ring and a pellicle [246]. During sporulation, the spores remaining in the biofilm ring are of great utility for the food industry, because they confer spore resistance on washing and cleaning procedures. The spores can restart a new biofilm when food production has resumed [103]. A recent work [247] reveals improvement in the outdoor use of Cry34Aa toxins (effective against Diptera) as encapsulated proteins in a polyhedral microcrystal toxic to Bombyx mori cypovirus. This system provides stability and long-term protein activity under hostile environmental and physical conditions and UV irradiation [247]. Bt cry genes have characteristics to enhance resistance to insect pests in genetically modified crops, the application of cry genes in plant genetic engineering, and the effect of the Bt against different species of invertebrates (nematodes, ticks, mites). New strains have been identified of Bt that produce Cyt2BA against Bradysia difformis (pests in mushroom production) [4,248]. Another examples of this is the B. thuringiensis strains GBA46 and NMTD81, and the strain FZB42, Of toxic against the phytopathogen Aphelenchoides besseyi and cause severe damage to various crops of plants and vegetables [37,71,239]. Of relevance is that the progress and the development of novel and improved biological technologies for the bioremediation of heavy metals and other pollutants [249,250], B. thuringiensis is a model for hijacking the process of the synthesis of crystal proteins in order to generate diverse proteins with applications in biotechnology and medicine (nanoparticle delivery system) [251]. B. thuringiensis is a potential and suitable cell factory for different novel, valuable bioproducts [249,250,251].
6. Conclusions and Remarks
The majority of studies regarding B. thuringiensis centered on elucidating the mechanism of action of the Cry proteins. Bt Cry toxins as bioinsecticides. However, in light of the emergence of several worthwhile studies on the Bacillus model B. subtilis, followed closely by that of B. cereus, the molecular biology of B. thuringiensis (Bt) sporulation, structural assembly, and germination has approximated it. As reviewed herein, the molecular machinery involved in the morphological and cytological processes is conserved in the genus Bacillus, which belongs to the phylum Firmicutes. Furthermore and remarkably, the metabolic regulation at transcriptional and translational levels of the sporulation and crystal proteins formation. Despite several differential spatio-temporal differences in gene expression and in sigma transcriptional and protein patterns, this morphological complex differentiation process reveals a common cell fate, preservation, and survival under nutrient limitation and harsh environmental conditions. A striking feature of the infectious life cycle of B. thuringiensis resides in the concomitant expression and production of the crystalline-delta endotoxins (Cry proteins), a strategy conferred on Bt for survival and success in the environment (biological control) and the host. Furthermore, the molecular co-evolution and molecular crosstalk in metabolism are maintained through the evolution between Cry endotoxins and the set of receptors immersed in the insect midgut, thus conferring an evolutive advantage for survival and success.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
This review paper did not require ethical approval.
Data Availability Statement
Not applicable.
Acknowledgments
In debt to the financial support of PERFIL-PRODEP PROGRAM SEP 2022-2025, and SNI CONACYT 2023-2027.
Conflicts of Interest
The author declares no conflict of interest.
References
- Liu, S.; Moayeri, M.; Leppla, S.H. Anthrax lethal and edema toxins in anthrax pathogenesis. Trends Microbiol. 2014, 22, 317–325. [Google Scholar] [CrossRef]
- Kokilaramani, S.; AlSalhi, M.S.; Devanesan, S.; Narenkuma, J.; Rajasekar, A.; Govarthanan, M. Bacillus megaterium-induced biocorrosion on mild steel and the effect of Artemisia pallens methanolic extract as a natural corrosion inhibitor. Arch. Microbiol. 2020, 202, 2311–2321. [Google Scholar] [CrossRef]
- Melo, A.L.D.A.; Soccol, V.T.; Soccol, C.R. Bacillus thuringiensis: Mechanism of action, resistance, and new applications: A review. Crit. Rev. Biotechnol. 2016, 36, 317–326. [Google Scholar] [CrossRef]
- Wang, C.; Li, W.; Kessenich, C.R.; Petrick, J.S.; Rydel, T.J.; Sturman, E.J.; Lee, T.C.; Glenn, K.C.; Edrington, T.C. Safety of the Bacillus thuringiensis-derived Cry1A.105 protein: Evidence that domain exchange preserves mode of action and safety. Regul.Toxicol. Pharmacol. 2018, 99, 50–60. [Google Scholar] [CrossRef]
- Malovichko, Y.V.; Nizhnikov, A.A. Repertoire of the Bacillus thuringiensis virulence factors unrelated to major classes of protein toxins and its role in specificity of host-pathogen interactions. Antonets KS. Toxins 2019, 11, 347. [Google Scholar] [CrossRef]
- Bach, E.; Rangel, C.P.; Ribeiro, I.D.A.; Passaglia, L.M.P. Pangenome analyses of Bacillus pumilus, Bacillus safensis, and Priestia megaterium exploring the plant-associated features of bacilli strains isolated from canola. Mol. Genet. Genom. 2022, 297, 1063–1079. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, Z.; Luo, X.; Zhang, X.; Chou, S.H.; Wang, J.; He, J. Which is stronger? A continuing battle between Cry toxins and insects. Front. Microbiol. 2021, 12, 665101. [Google Scholar] [CrossRef] [PubMed]
- Stenfois Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef]
- Logan, N.A. Bacillus and relatives in foodborne illness. J. Appl. Microbiol. 2012, 112, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Dalla-Vecchia, E.; Visser, M.; Stams, A.J.; Bernier-Latmani, R. Investigation of sporulation in the Desulfotomaculum genus: A genomic comparison with the genera Bacillus and Clostridium. Environ. Microbiol. Rep. 2014, 6, 756–766. [Google Scholar] [CrossRef]
- Talukdar, P.K.; Sarker, M.R. Characterization of putative sporulation and germination genes in Clostridium perfringens Food-Poisoning Strain SM101. Microorganisms 2022, 10, 1481. [Google Scholar] [CrossRef]
- Chakraborty, A.; Jayne ERattray, S.; Matthews, D.S.; Li, C.; Barker, B.; Jørgensen, B.B.; Hubert, C.R.J. Metabolic responses of thermophilic endospores to sudden heat-induced perturbation in marine sediment samples. Front. Microbiol. 2022, 13, 958417. [Google Scholar] [CrossRef] [PubMed]
- Burke, K.E.; Lamont, J.T. Clostridium difficile infections: A worldwide disease. Gut Liver 2014, 8, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, B.K.; McBride, S.H.M. Clostridioides difficile. Trends Microbiol. 2018, 26, 1049–1050. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, R.; Song, J.A. Terbium chloride influences Clostridium difficile spore germination. Anaerobe 2019, 58, 80–88. [Google Scholar] [CrossRef]
- Diallo, M.; Kengen, S.W.M.; López-Contreras, A.M. Sporulation in solventogenic and acetogenic clostridia. Appl. Microbiol. Biotechnol. 2021, 105, 3533–3557. [Google Scholar] [CrossRef]
- Setlow, P.; Johnson, E.A. Spores and their significance. In Food Microbiology, Fundamentals and Frontiers, 4th ed.; Doyle, M.P., Buchanan, R., Eds.; ASM Press: Washington, WA, USA, 2012; pp. 45–79. [Google Scholar]
- Bertuso, P.C.; Marangon, C.A.; Nitschke, M. Susceptibility of Vegetative Cells and Endospores of Bacillus cereus to Rhamnolipid Biosurfactants and Their Potential Application in Dairy. Microorganisms 2022, 10, 1860. [Google Scholar] [CrossRef]
- Auger, S.; Ramarao, N.; Faille, C.; Fouet, A.; Aymerich, S.; Gohar, M. Biofilm formation and cell surface properties among pathogenic and nonpathogenic strains of the Bacillus cereus group. Appl. Environ. Microbiol. 2009, 75, 6616–6618. [Google Scholar] [CrossRef]
- Vary, P.S.; Biedendieck, R.; Fuerch, T.; Meinhardt, F.; Rohde, M.; Deckwer, D.W.; Jahn, D. Bacillus megaterium--from simple soil bacterium to industrial protein production host. Appl. Microbiol. Biotechnol. 2007, 76, 957–967. [Google Scholar] [CrossRef]
- Mallozzi, M.; Viswanathan, V.K.; Vedantam, G. Spore-forming bacilli and clostridia in human disease. Future Microbiol. 2010, 5, 1109–1123. [Google Scholar] [CrossRef]
- Chi, Y.; Wang, D.; Jiang, M.; Chu Sh Wang, B.; Zhi, Y.; Zhou, P.; Zhang, D. Microencapsulation of Bacillus megaterium NCT-2 and its effect on remediation of secondary salinization soil. J. Microencapsul. 2020, 37, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhu, G.; Korza, G.; Sun, X.; Setlow, P.; Li, J. Engineering Bacillus subtilis as a versatile and stable platform for production of nanobodies. Appl. Environ. Microbiol. 2020, 86, e02938-19. [Google Scholar] [CrossRef] [PubMed]
- Galperin, M.Y.; Yutin, N.; Wolf, Y.I.; Vera-Álvarez, R.; Koonin, E.V. Conservation and evolution of the sporulation gene set in diverse members of the Firmicutes. J. Bacteriol. 2022, 204, e0007922. [Google Scholar] [CrossRef]
- Hutchison, E.A.; Miller, D.A.; Angert, E.R. Sporulation in bacteria: Beyond the standard model. Microbiol. Spectr. 2014, 2, 1–15. [Google Scholar] [CrossRef]
- Bate, A.R.; Bonneau, R.; Eichenberger, P. Bacillus subtilis Systems Biology: Applications of- Omics Techniques to the Study of Endospore Formation. Microbiol. Spectr. 2014, 2, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kolek, J.; Diallo, M.; Vasylkivska, M.; Branska, B.; Sedlar, K.; López-Contreras, A.M.; Patakova, P. Comparison of expression of key sporulation, solventogenic and acetogenic genes in C. beijerinckii NRRL B-598 and its mutant strain overexpressing spo0A. Appl Microbiol Biotechnol. 2017, 101, 8279–8291. [Google Scholar] [CrossRef]
- Al-Hinai, M.A.; Jones, S.W.; Papoutsakis, E.T. σK of Clostridium acetobutylicum is the first known sporulation-specific sigma factor with two developmentally separated roles, one early and one late in sporulation. J. Bacteriol. 2014, 196, 287–299. [Google Scholar] [CrossRef]
- Al-Hinai, M.A.; Jones, S.W.; Papoutsakis, E.T. The Clostridium sporulation programs: Diversity and preservation of endospore differentiation. Microbiol. Mol. Biol. Rev. 2015, 79, 19–37. [Google Scholar] [CrossRef]
- Zhou, Y.; Choi, Y.L.; Sun, M.; Yu, Z. Novel roles of Bacillus thuringiensis to control plant diseases. Appl. Microbiol. Biotechnol. 2008, 80, 563–572. [Google Scholar] [CrossRef]
- Tocheva, E.I.; Ortega, D.R.; Jensen, G.J. Sporulation, bacterial cell envelopes and the origin of life. Nat. Rev. Microbiol. 2016, 14, 535–542. [Google Scholar] [CrossRef]
- Wolska, K.I.; Grudniak, A.M.; Kraczkiewicz-Dowjat, A. Genetic and physiological regulation of bacterial endospore development. Pol. J. Microbiol. 2007, 56, 11–17. [Google Scholar] [PubMed]
- Gupta, R.S. Origin of diderm (Gram-negative) bacteria: Antibiotic selection pressure rather than endosymbiosis likely led to the evolution of bacterial cells with two membranes. Antonie Van Leeuwenhoek 2011, 100, 171–182. [Google Scholar] [CrossRef]
- Tu, Z.; Dekker, H.L.; Roseboom, W.; Swarge, B.N.; Setlow, P.; Brul, S.; Kramer, G. High resolution analysis of proteome dynamics during Bacillus subtilis sporulation. Int. J. Mol. Sci. 2021, 22, 9345. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Mei, H.; Qian, H.; Tang, Q.; Liu, X.; Yu, Z.; He, J. Expression profile and regulation of spore and parasporal crystal formation-associated genes in Bacillus thuringiensis. J. Proteome Res. 2013, 12, 5487–5501. [Google Scholar] [CrossRef]
- Wang, J.; Mei, H.; Zheng, C.; Qian, H.; Cui, C.; Fu, Y.; Su, J.; Liu, Z.; Yu, Z.; He, J. The metabolic regulation of sporulation and parasporal crystal formation in Bacillus thuringiensis revealed by transcriptomics and proteomics. Mol. Cell. Proteomics. 2013, 12, 1363–1376. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Vischer, N.O.E.; Wekking, D.; Boggian, A.; Setlow, P.; Brul, S. Visualization of SpoVAEa protein dynamics in dormant spores of Bacillus cereus and dynamic changes in their germinosomes and SpoVAEa during germination. Microbiol. Spectr. 2022, 10, e0066622. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Barajas-Ornelas, R.C.; Damon, J.; Ramírez, G.F.H.; Alon, A.; Brock, K.P.; Marks, D.S.; Kruse, A.C.; Rudner, D. The SpoVA membrane complex is required for dipicolinic acid import during sporulation and export during germination. Genes. Dev. 2022, 36, 634–646. [Google Scholar] [CrossRef]
- Koopman, N.; Remijas, L.; Seppen, J.; Setlow, P.; Brul, S. Mechanisms and applications of bacterial sporulation and germination in the intestine. Int. J. Mol. Sci. 2022, 23, 3405. [Google Scholar] [CrossRef]
- Ultee, E.; Ramijan, K.; Dame, R.T.; Briegel, A.; Claessen, D. Stress-induced adaptive morphogenesis in bacteria. Adv. Microb. Physiol. 2019, 74, 97–141. [Google Scholar] [CrossRef]
- De Maagd, R.A.; Bravo, A.; Berry, N.; Crickmore, N.; Schnepf, H.E. Structure, diversity, and evolution of proteins toxins from spore-forming entomopathogenic bacteria. Annu. Rev. Genet. 2003, 37, 409–433. [Google Scholar] [CrossRef]
- Bel, Y.; Ferré, J.; Hernández-Martínez, P. Bacillus thuringiensis toxins: Functional characterization and mechanism of action. Toxins 2020, 12, 785. [Google Scholar] [CrossRef] [PubMed]
- Jurat-Fuentes, J.L.; Crickmore, N. Specificity determinants for Cry insecticidal proteins: Insights from their mode of action. J. Invertebr. Pathol. 2017, 142, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Aronson, A.I.; Shai, Y. Why Bacillus thuringiensis insecticidal toxins are so effective: Unique features of their mode of action. FEMS Microbiol. Lett. 2001, 195, 1–8. [Google Scholar] [CrossRef]
- Bravo, A.; Gill, S.; Soberón, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Likitvivatanavong, S.; Gill, S.S.; Soberón, M. Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochem. Mol. Biol. 2011, 41, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Pardo-López, L.; Soberón, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: Mode of action, insect resistance and consequences for crop protection. FEMS Microbiol Rev. 2012, 37, 3–22. [Google Scholar] [CrossRef]
- Fimlaid, K.A.; Shen, A. Diverse mechanisms regulate sporulation sigma factor activity in the Firmicutes. Curr. Opin. Microbiol. 2015, 24, 88–95. [Google Scholar] [CrossRef]
- Wilcks, A.; Jayaswal, N.; Lereclus, D.; Andrup, L. Characterization of plasmid pAW63, a second self-transmissible plasmid in Bacillus thuringiensis subsp. kurstaki HD73. Microbiology 1998, 144, 1263–1270. [Google Scholar] [CrossRef] [PubMed]
- Wilcks, A.; Smidt, L.; Økstad, O.A.; Kolstø, A.B.; Mahillon, J.; Andrup, L. Replication mechanism and sequence analysis of the replicon of pAW63, a conjugative plasmid from Bacillus thuringiensis. J. Bacteriol. 1999, 181, 3193–3200. [Google Scholar] [CrossRef] [PubMed]
- Van der Auwera, G.; Andrup, L.; Mahillon, J. Conjugative plasmid pAW63 brings new insights into the genesis of the Bacillus anthracis virulence plasmid pXO2 and of the Bacillus thuringiensis plasmid pBT9727. BMC Genom. 2005, 6, 103–114. [Google Scholar] [CrossRef]
- Beuls, E.; Modrie, P.; Deserranno, C.; Mahillon, J. High-salt stress conditions increase the pAW63 transfer frequency in Bacillus thuringiensis. Appl. Environ. Microbiol. 2012, 78, 7128–7131. [Google Scholar] [CrossRef] [PubMed]
- Fazion, F.; Perchat, S.; Buisson, C.; Vilas-Bôas, G.; Lereclus, D. A plasmid-borne Rap-Phr system regulates sporulation of Bacillus thuringiensis in insect larvae. Environ. Microbiol. 2018, 20, 145–155. [Google Scholar] [CrossRef]
- Cardoso, P.; Fazion, F.; Perchat, S.; Buisson, C.; Vilas-Bôas, G.; Lereclus, D. RapPhr systems from plasmids pAW63 and pHT8-1 act together to regulate sporulation in the Bacillus thuringiensis serovar kurstaki HD73 strain. Appl. Environ. Microbiol. 2020, 86, e01238–e20. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.H.; Zhang, R.; Lereclus, D.; Peng, Q.; Zhang, J.; Slamti, L.; Song, F. The transcription factor CpcR determines cell fate by modulating the initiation of sporulation in Bacillus thuringiensis. Appl. Environ. Microbiol. 2022, 88, e0237421. [Google Scholar] [CrossRef]
- Gastélum, G.; de la Torre, M.; Rocha, J. Rap protein paralogs of Bacillus thuringiensis: A multifunctional and redundant regulatory repertoire for the control of collective functions. J. Bacteriol. 2019, 202, e00747-19. [Google Scholar] [CrossRef]
- Li, J.D.; Carroll, J.; Ellar, D.J. Crystal structure of insecticidal delta-endotoxin from Bacillus thurigiensis at 2.5 Ǻ resolution. Nature 1991, 352, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A. Phylogenetic relationships of Bacillus thuringiensis δ-endotoxin family proteins and their functional domains. J. Bacteriol. 1997, 179, 2793–2801. [Google Scholar] [CrossRef]
- Grochulski, P.; Masson, L.; Borisova, S.; Pusztai-Carey, M.; Schwartz, J.L.; Brousseau, R.; Cygler, M. Bacillus thuringiensis Cry1A(a) insecticidal toxin: Crystal structure and channel formation. J. Mol. Biol. 1995, 254, 447–464. [Google Scholar] [CrossRef]
- Derbyshire, D.J.; Ellar, D.J.; Li, J. Crystallization of the Bacillus thuringiensis toxin Cry2Ac and its complex with the receptor ligand N-Acetylgalactosamine. Acta Cryst. Sect. 2001, 57, 1938–1944. [Google Scholar] [CrossRef]
- Morse, R.J.; Yamamoto, T.; Stroud, R.M. Structure of Cry2Aa suggests an unexpected receptor binding epitope. Structure 2001, 9, 409–417. [Google Scholar] [CrossRef]
- Galistsky, N.; Cody, V.; Wojtczak, D.; Ghosh, J.R.; Luft, W.P.; English, L. Structure of the insecticidal bacterial δ-endotoxin Cry3Bb1 of Bacillus thuringiensis. Acta Crystallogr. Sect. D 2001, 57, 1101–1109. [Google Scholar] [CrossRef] [PubMed]
- Boonserm, P.; Davis, P.; Ellar, D.J.; Li, J. Crystal structure of the mosquito-larvacidal toxin Cry4Ba and its biological implications. J. Mol. Biol. 2005, 348, 363–382. [Google Scholar] [CrossRef] [PubMed]
- Boonserm, P.; Mo, M.; Angsuthanasombat, A.; Lescar, J. Structure of the functional form of the mosquito larvicidal Cry4Aa toxin from Bacillus thuringiensis at a 2.8 angstrom resolution. J. Bacteriol. 2006, 188, 3391–3401. [Google Scholar] [CrossRef]
- Garczynski, S.F.; Crim, J.W.; Adang, M.J. Identification of putative insect brush border membrane-binding molecules specific to Bacillus thuringiensis δ-endotoxin by protein blot analysis. Appl. Environ. Microbiol. 1991, 57, 2816–2820. [Google Scholar] [CrossRef] [PubMed]
- Fernández, E.; Aimanova, K.G.; Gill, S.S.; Bravo, A.; Soberón, M. A GPI-anchored alkaline phosphatase is a functional midgut receptor of Cry1Aa toxin in Aedes aegypti larvae. Biochem. J. 2006, 394, 77–84. [Google Scholar] [CrossRef]
- Flannagan, R.D.; Yu, C.G.; Mathis, J.P.; Meyer, T.E.; Shi, X.; Siqueira, H.A.A.; Siegfried, B.D. Identification, cloning and expression of Cry1Ab cadherin receptor from European corn borer, Ostrinia nubilalis (Hübner) (Lepidoptera: Crambidae). Insect Biochem. Mol. Biol. 2005, 35, 33–40. [Google Scholar] [CrossRef]
- Burton, S.L.; Ellar, D.J.; Li, J.; Derbyshire, D.J. N-acetylgalactosamine on the putative insect receptor aminopeptidase N is recognized by a site on the domain III lectin-like fold of a Bacillus thuringiensis insecticidal toxin. J. Mol. Biol. 1999, 287, 1011–1022. [Google Scholar] [CrossRef]
- De Maagd, R.A.; Bakker, P.L.; Masson, L.; Adang, M.J.; Sangadala, W.S.; Bosch, D. Domain III of the Bacillus thuringiensis δ-endotoxin Cry1Ac is involved in binding to Manduca sexta brush border membranes and its purifies aminopeptidases N. Mol. Microbiol. 1999, 31, 463–471. [Google Scholar] [CrossRef]
- Kitami, M.; Kadotani, T.; Nakanishi, K.; Atsumi, S.; Higurashi, S.; Ishizaka, T.; Watanabe, A.; Sato, R. Bacillus thuringiensis Cry toxins bound specifically to various proteins via domain III, which had a galactose-binding domain-like fold. Biosci. Biotechnol. Biochem. 2011, 75, 305–312. [Google Scholar] [CrossRef]
- Hernández-Martínez, P.; Khorramnejad, A.; Prentice, K.; Andrés-Garrido, A.; Vera-Velasco, N.M.; Smagghe, G.; Escriche, B. The independent biological activity of Bacillus thuringiensis Cry23Aa protein against Cylas puncticollis. Front. Microbiol. 2020, 11, 1734. [Google Scholar] [CrossRef]
- Jenkins, J.I.; Lee, M.K.; Valaitis, A.P.; Curtiss, A.; Dean, D.H. Bivalent sequential binding model of a Bacillus thuringiensis toxin to gypsy moth aminopeptidase N receptor. J. Biol. Chem. 2000, 275, 14423–14443. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Gómez, I.; Conde, J.; Munoz-Garay, C.; Sánchez, J.; Miranda, R.; Zhuang, M.; Gill, S.S.; Soberón, M. Oligomerization triggers. Binding of a Bacillus thuringiensis Cry1Ab pore-forming toxin to aminopeptidase N receptor leading to insertion into membrane microdomains. Biochim. Biophys. Acta. 2004, 1667, 38–46. [Google Scholar] [CrossRef]
- Jiménez-Juárez, N.; Muñoz-Caray, C.; Gómez, I.; Gill, S.S.; Soberón, M.; Bravo, A. The pre-pore from Bacillus thuringiensis Cry1Ab toxin is necessary to induce insect death in Manduca sexta. Peptides 2008, 29, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Vechon, V.; Laprade, R.; Schwartz, J.L. Current models of the mode of action of Bacillus thuringiensis insecticidal crystal proteins: A critical review. J. Invertebr. Pathol. 2012, 111, 1–12. [Google Scholar] [CrossRef]
- Sánchez, J.; Holmgren, J. Cholera toxin structure, gene regulation and pathophysiological and immunological aspects. Cell. Mol. Life Sci. 2008, 65, 1347–1360. [Google Scholar] [CrossRef] [PubMed]
- Wernick, N.L.; Chinnapen, D.J.; Cho, J.A.; Lencer, W.I. Cholera toxin: An intracellular journey into the cytosol by way of the endoplasmic reticulum. Toxins 2010, 2, 310–325. [Google Scholar] [CrossRef]
- He, X.; Yang, J.; Ji, M.; Chen, Y.; Chen, Y.; Li, H.; Wang, H. A potential delivery system based on cholera toxin: A macromolecule carrier with multiple activities. J. Control. Release 2022, 343, 551–563. [Google Scholar] [CrossRef]
- Setlow, P. I will survive: DNA protection in bacterial spores. Trends Microbiol. 2007, 15, 172–180. [Google Scholar] [CrossRef]
- Yutin, N.; Galperin, M.Y. A genomic update on clostridial phylogeny: Gram-negative spore formers and other misplaced clostridia. Environ. Microbiol. 2013, 15, 2631–2641. [Google Scholar] [CrossRef]
- Veering, J.W.; Stewart, E.J.; Berngruber, T.W.; Taddei, F.; Kuipens, O.; Hamoen, L.W. Bet hedging and epigenetic inheritance in bacterial cell development. Proc. Natl. Acad. Sci. USA 2008, 105, 4393–4398. [Google Scholar] [CrossRef]
- Tan, I.S.; Ramamurthi, K.S. Spore formation in Bacillus subtilis. Environ. Microbiol. Rep. 2014, 6, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Rigden, D.J.; Galperin, M.Y. Sequence analysis of GerM and SpoVS, uncharacterized bacterial sporulation’ proteins with widespread phylogenetic distribution. Bioinformatics 2018, 24, 1793–1797. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Traag, B.A.; Pugliese, A.; Eisen, J.A.; Losick, R. Gene conservation among endospore-forming bacteria reveals additional sporulation genes in Bacillus subtilis. J. Bacteriol. 2013, 195, 253–260. [Google Scholar] [CrossRef]
- Dworkin, J. Protein targeting during Bacillus subtilis sporulation. Microbiol. Spectr. 2014, 2, 1–12. [Google Scholar] [CrossRef]
- Perego, M.; Hoch, J.A. Cell-cell communication regulates the effects of protein aspartate phosphatases on the phosphorelay controlling development in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 1996, 93, 1549–1553. [Google Scholar] [CrossRef]
- Perego, M. A peptide export-import control circuit modulating bacterial development regulates protein phosphatases of the phosphorelay. Proc. Natl. Acad. Sci. USA 1997, 94, 8612–8617. [Google Scholar] [CrossRef]
- Perego, M. A new family of aspartyl phosphate phosphatases targeting the sporulation transcription factor Spo0A of Bacillus subtilis. Mol. Microbiol. 2001, 42, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Shao, W.; Perego, M.; Hoch, J.A. Multiple histidine kinases regulate entry into stationary phase and sporulation in Bacillus subtilis. Mol. Microbiol. 2000, 38, 535–542. [Google Scholar] [CrossRef]
- Grenha, R.; Rzechorzek, N.J.; Brannigan, J.A.; de Jong, R.N.; Ab, E.; Diercks, T.; Truffault, V.; Ladds, J.C.; Fogg, M.J.; Bongiorni, C.; et al. Structural characterization of Spo0E-like protein-aspartic acid phosphatases that regulate sporulation in bacilli. J. Biol. Chem. 2006, 281, 37993–38003. [Google Scholar] [CrossRef] [PubMed]
- Grenha, R.; Slamti, L.; Nicaise, M.; Refes, Y.; Lereclus, D.; Nessler, S. Structural basis for the activation mechanism of the PlcR virulence regulator by the quorum-sensing signal peptide, P.a.p.R. Proc. Natl. Acad. Sci. USA 2013, 110, 1047–1052. [Google Scholar] [CrossRef]
- Hayashi, K.; Kensuke, T.; Kobayashi, K.; Ogasawara, N.; Ogura, M. Bacillus subtilis RghR (YvaN) represses rapG and rapH, which encode inhibitors of expression of the srfA operon. Mol. Microbiol. 2006, 59, 1714–1729. [Google Scholar] [CrossRef] [PubMed]
- Díaz, A.R.; Stephenson, S.; Green, J.M.; Levdikov, V.M.; Wilkinson, A.J.; Perego, M. Functional role for a conserved aspartate in the Spo0E signature motif involved in the dephosphorylation of the Bacillus subtilis sporulation regulator Spo0A. J. Biol. Chem. 2008, 283, 2962–2972. [Google Scholar] [CrossRef] [PubMed]
- Declerck, N.; Bouillaut, L.; Chaix, D.; Rugani, N.; Slamti, L.; Hoh, F.; Lereclus, D.; Arold, S.T. Structure of PlcR: Insights into virulence regulation and evolution of quorum sensing in Gram-positive bacteria. Proc. Natl. Acad. Sci. USA 2007, 104, 18490–18495. [Google Scholar] [CrossRef]
- Slamti, L.; Perchat, S.; Huillet, E.; Lereclus, D. Quorum sensing in Bacillus thuringiensis is required for completion of a full infectious cycle in the insect. Toxins 2014, 6, 2239–2255. [Google Scholar] [CrossRef]
- Ishikawa, S.; Core, L.; Perego, M. Biochemical characterization of aspartyl phosphate phosphatase interaction with a phosphorylated response regulator and its inhibition by a pentapeptide. J. Biol. Chem. 2002, 77, 20483–20489. [Google Scholar] [CrossRef] [PubMed]
- Sonenshein, A.L. Control of sporulation initiation in Bacillus subtilis. Curr. Opin. Microbiol. 2000, 3, 561–566. [Google Scholar] [CrossRef]
- Burbulys, D.; Trach, K.A.; Hoch, J.A. Initiation of sporulation in B. subtilis is controlled by a multicomponent phosphorelay. Cell 1991, 64, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Molle, V.; Fujita, M.; Jensen, S.T.; Eichenberger, P.; González-Pastor, J.E.; Liu, J.S.; Losick, R. The Spo0A regulon of Bacillus subtilis. Mol. Microbiol. 2003, 50, 1683–1701. [Google Scholar] [CrossRef] [PubMed]
- Barák, I.; Youngman, P. SpoIIE mutants of Bacillus subtilis comprise two distinct phenotypic classes consistent with a dual functional role for the SpoIIE protein. J. Bacteriol. 1996, 178, 4984–4989. [Google Scholar] [CrossRef]
- Barák, I.; Muchová, K.; Labajová, N. Asymmetric cell division during Bacillus subtilis sporulation. Future Microbiol. 2019, 14, 353–363. [Google Scholar] [CrossRef]
- Pottathil, M.; Lazazzera, B.A. The extracellular Phr peptide-Rap phosphatase signaling circuit of Bacillus subtilis. Front. Biosci. 2003, 8, d32–d45. [Google Scholar] [CrossRef]
- Fagerlund, A.; Dubois, T.; Okstad, O.A.; Verplaetse, E.; Gilois, N.; Bennaceur, I.; Perchat, M.; Myriam, G.; Aymerich, S.; Kolstø, A.-B.; et al. SinR controls enterotoxin expression in Bacillus thuringiensis biofilms. PLoS ONE 2014, 9, e87532. [Google Scholar] [CrossRef]
- Dyrdahl-Young, R.; Hu, W.; DiGennaro, P. Temporal expression patterns of Pasteuria spp. sporulation genes. J. Nematol. 2019, 29, e2019–e2039. [Google Scholar] [CrossRef]
- Boonstra, M.; de Jong, I.G.; Scholefield, G.; Murray, H.; Kuipers, O.P.; Veering, J.W. SpoOA regulates chromosome copy number during sporulation by directly binding to the origin of replication in Bacillus subtilis. Mol. Microbiol. 2013, 87, 925–938. [Google Scholar] [CrossRef] [PubMed]
- Eichenberger, P.; Jensen, S.T.; Conlon, E.M.; van Ooij, C.; Silvaggi, J.; González-Pastor, J.E.; Fujita, M.; Ben-Yehuda, S.; Stragier, P.; Liu, J.S.; et al. The Sigma E regulon and the identification of additional sporulation genes in Bacillus subtilis. J. Mol. Biol. 2003, 327, 945–972. [Google Scholar] [CrossRef] [PubMed]
- McKenney, P.T.; Driks, A.; Eichenberger, P. The Bacillus subtilis endospore: Assembly and functions of the multilayered coat. Nat. Rev. Microbiol. 2013, 11, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.; Dworkin, J. Recent progress in Bacillus subtilis sporulation. FEMS Microbiol. Rev. 2012, 36, 131–148. [Google Scholar] [CrossRef] [PubMed]
- Pérez, A.R.; Abanes-De Mello, A.; Pogliano, K. Suppression of engulfment defects in Bacillus subtilis by elevated expression of the motility regulon. J. Bacteriol. 2006, 188, 1159–1164. [Google Scholar] [CrossRef]
- Plomp, M.; Carroll, A.M.; Setlow, P.; Malkin, A.J. Architecture and assembly of the Bacillus subtilis spore coat. PLoS ONE 2014, 9, e108560. [Google Scholar] [CrossRef]
- Daniel, R.A.; Drake, S.; Buchanan, C.E.; Scholle, R.; Errington, J. The Bacillus subtilis spoVD gene encodes a mother-cell-specific Penicillin-binding protein required for spore morphogenesis. J. Mol. Biol. 1994, 235, 209–220. [Google Scholar] [CrossRef]
- Resnekov, O.; Driks, A.; Losick, R. Identification and characterization of sporulation gene spoVS from Bacillus subtilis. J. Bacteriol. 1995, 177, 5628–5635. [Google Scholar] [CrossRef][Green Version]
- Matsuno, K.; Sonenshein, A.L. Role of SpoVG in asymmetric septation in Bacillus subtilis. J. Bacteriol. 1999, 181, 3392–3401. [Google Scholar] [CrossRef] [PubMed]
- Radford, D.S.; Wan, Q.; Tzokov, S.; Moir, A.; Bullough, P.A. Molecular tiling on the surface of a bacterial spore- the exosporium of the Bacillus anthracis/cereus/thuringiensis group. Mol Microbiol. 2017, 104, 539–552. [Google Scholar] [CrossRef]
- Henriques, A.O.; Morán Jr, C.H.P. Structure, assembly, and function of the spore surface layers. Annu. Rev. Microbiol. 2007, 61, 555–588. [Google Scholar] [CrossRef]
- Chen, X.; Gao, T.; Peng, Q.; Zhang, J.; Chai, Y.; Song, F. Novel cell wall hydrolase CwlC from Bacillus thuringiensis is essential for mother cell lysis. Appl. Environ. Microbiol. 2018, 84, e02640-17. [Google Scholar] [CrossRef]
- Plomp, M.; Leighton, J.T.; Wheeler, K.E.; Malkin, A.J. Architecture and high-resolution structure of Bacillus thuringiensis and Bacillus cereus spore coat surfaces. Langmuir 2005, 21, 7892–7898. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Wu, J.; Chen, X.; Qiu, L.; Zhang, J.; Tian, H.; Song, F. Disruption of two-component system LytSR affects forespore engulfment in Bacillus thuringiensis. Front. Cell. Infect. Microbiol. 2017, 7, 468. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, R.; Hou, S.; HLiu, H.; Wang, J.; Yu, Q.; Peng, O.; Song, F. Identification and functional characterization of two homologous SpoVS proteins involved in sporulation of Bacillus thuringiensis. Microbiol. Spectr. 2021, 9, e0088121. [Google Scholar] [CrossRef]
- Dubois, T.; Lemy, C.; Perchat, S.; Lereclus, D. The signaling peptide NprX controlling sporulation and necrotrophism is imported into Bacillus thuringiensis by two oligopeptide permease systems. Mol Microbiol. 2019, 112, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yu, Z.; Liu, L.; Chou, S.H.-H.; Wang, J.; He, J. 6S-1 RNA contributes to sporulation and parasporal crystal formation in Bacillus thuringiensis. Front. Microbiol. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Byrd, B.; Camilleri, E.; Korza, G.; Craft, D.L.; Green, J.; Rocha, G.M.; Mok, W.W.K.; Cairmano, M.J.; Setlow, P. Levels and characteristics of mRNAs in Spores of Firmicute Species. J. Bacteriol. 2021, 203, e0001721. [Google Scholar] [CrossRef]
- Riley, E.P.; López-Garrido, J.; Sugie, J.; Liu, R.B.; Pogliano, K. Metabolic differentiation and intercellular nurturing underpin bacterial endospore formation. Sci. Adv. 2021, 7, eabd6385. [Google Scholar] [CrossRef]
- Korza, G.; Camilleri, E.; Green, J.; Robinson, J.; Nagler, K.; Moeller, R.; Caimano, M.J.; Setlow, P. Analysis of mRNAs in spores of Bacillus subtilis. J. Bacteriol. 2019, 201, e00007-19. [Google Scholar] [CrossRef]
- Swarge, B.; Abhyankar, W.; Jonker, M.; Hoefsloot, H.; Kramer, G.; Setlow, P.; Brul, S.; de Koning, L.J. Integrative analysis of proteome and transcriptome dynamics during Bacillus subtilis spore revival. mSphere 2020, 5, e00463-20. [Google Scholar] [CrossRef] [PubMed]
- Setlow, P.; Christie, G. Bacterial spore mRNA: What’s up with that? Front. Microbiol. 2020, 11, 596092. [Google Scholar] [CrossRef] [PubMed]
- Jeng, Y.H.; Doi, R. Messenger ribonucleic acid of dormant spores of Bacillus subtilis. J. Bacteriol. 1974, 119, 514–521. [Google Scholar] [CrossRef]
- Segev, E.; Smith, Y.; Ben-Yehuda, S. RNA dynamics in aging bacterial spores. Cell 2012, 148, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Camp, A.H.; Losick, R. A novel pathway of intercellular signalling in Bacillus subtilis involves a protein with similarity to a component of type III secretion channels. Mol. Microbiol. 2008, 69, 402–417. [Google Scholar] [CrossRef] [PubMed]
- Camp, A.H.; Losick, R. A feeding tube model for activation of a cell specific transcription factor during sporulation in Bacillus subtilis. Genes. Dev. 2009, 23, 1014–1024. [Google Scholar] [CrossRef]
- Crawshaw, A.D.; Serrano, M.; Stanley, W.A.; Henriques, A.O.; Salgado, P.S. A mother cell-to-forespore channel: Current understanding and future challenges. FEMS Microbiol. Lett. 2014, 358, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Meisner, J.; Wang, X.; Serrano, M.; Henriques, A.O.; Moran, C.P., Jr. A channel connecting the mother cell and forespore during bacterial endospore formation. Proc. Natl. Acad. Sci. USA 2008, 105, 15100–15105. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Setlow, B.; Setlow, P. Levels of small molecules and enzymes in the mother cell compartment and the forespore of sporulatin Bacillus megaterium. J. Bacteriol. 1977, 130, 1130–1138. [Google Scholar] [CrossRef]
- Ghosh, A.; Manton, J.D.; Mustafa, A.R.; Gupta, M.; Ayuso-García, A.; Rees, E.J.; Graham, C. Proteins encoded by the gerP operon are localized to the inner coat in Bacillus cereus spores and are dependent on GerPA and SafA for assembly. Appl. Environ. Microbiol. 2018, 84, e00760-18. [Google Scholar] [CrossRef] [PubMed]
- De Francesco, M.; Jacobs, J.Z.; Nunes, F.; Serrano, M.; McKenney, P.T.; Chua, M.H.; Henriques, A.O.; Eichenberger, P. Physical interaction between coat morphogenetic proteins SpoVID and CotE is necessary for spore encasement in Bacillus subtilis. J. Bacteriol. 2012, 194, 4941–4950. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.C. The exosporium layer of bacterial spores: A connection to the environment and the infected host. Microbiol. Mol. Biol. Rev. 2015, 79, 437–457. [Google Scholar] [CrossRef]
- Takamatsu, H.; Watabe, K. Assembly and genetics of spore protective structures. Cell. Mol. Life Sci. 2002, 59, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Popham, D.L.; Bernhards, C.B. Spore peptidoglycan. Microbiol. Spectr. 2015, 3, 1–21. [Google Scholar] [CrossRef]
- Abanes-De Mello, A.; Sun, Y.-I.; Aung, S.; Pogliano, K. A cytoskeleton-like role for the bacterial cell wall during engulfment of the Bacillus subtilis fore-spore. Genes. Dev. 2002, 16, 3253–3264. [Google Scholar] [CrossRef]
- Verplaetse, E.; Slamti, L.; Gohar, M.; Lereclus, D. Cell differentiation in a Bacillus thuringiensis population during planktonic growth, biofilm formation, and host infection. MBio 2015, 6, e00138-15. [Google Scholar] [CrossRef]
- Verplaetse, E.; Slamti, L.; Gohar, M.; Lereclus, D. Two distinct pathways lead Bacillus thuringiensis to commit to sporulation in biofilm. Res. Microbiol. 2016, 168, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Han, G.; Fan, X.; Lv, J.; Zhang, X.; Peng, Q.; Zhang, J.; Xu, J.; Song, F. Characteristics of the sigK deletion mutant from Bacillus thuringiensis var. israelensis strain Bt-59. Curr. Microbiol. 2020, 77, 3422–3429. [Google Scholar] [CrossRef]
- Bechtel, D.B.; Bulla, L.A. Electron microscope study of sporulation and parasporal crystal formation in Bacillus thuringiensis. J. Bacteriol. 1976, 127, 1472–1481. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Agaisse, H.; Salamitou, S.; Lereclus, D. Analysis of cryIAa expression in sigE and sigK mutants of Bacillus thuringiensis. Mol. Gen. Genet. 1996, 250, 734–741. [Google Scholar] [CrossRef] [PubMed]
- Poncet, S.; Dervyn, E.; Klier, A.; Rapoport, G. Spo0A represses transcription of the cry toxin genes in Bacillus thuringiensis. Microbiology 1997, 143, 2743–2751. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Peng, Q.; Song, F.; Lereclus, D. Regulation of cry gene expression in Bacillus thuringiensis. Toxins 2014, 6, 2194–2209. [Google Scholar] [CrossRef]
- Zhang, R.; Slamti, L.; Tong, L.; Verplaetse, E.; Ma, L.; Lemy, C.; Peng, Q.; Guo, S.; Zhang, J.; Song, F.; et al. The stationary phase regulator CpcR activates cry gene expression in non-sporulating cells of Bacillus thuringiensis. Mol. Microbiol. 2020, 113, 740–754. [Google Scholar] [CrossRef]
- Huang, L.; Xu, L.; Han, G.; Crickmore, N.; Song, F.; Xu, J. Characterization of CwlC, an autolysin, and its role in mother cell lysis of Bacillus thuringiensis subsp. israelensis. Lett. Appl. Microbiol. 2022, 74, 92–102. [Google Scholar] [CrossRef]
- Lv, J.; Zhang, X.; Gao, T.; Cui, T.; Peng, Q.; Zhang, J.; Song, F. Effect of the spoIIID mutation on mother cell lysis in Bacillus thuringiensis. Appl. Microbiol. Biotechnol. 2019, 103, 4103–4112. [Google Scholar] [CrossRef] [PubMed]
- Kant, S.; Kapoor, R.; Banerjee, N. Identification of a catabolite-responsive element necessary for regulation of the cry4A gene of Bacillus thuringiensis subsp. israelensis. J. Bacteriol. 2009, 191, 4687–4692. [Google Scholar] [CrossRef]
- Du, L.; Qiu, L.; Peng, Q.; Lereclus, D.; Zhang, J.; Song, F.; Huang, D. Identification of the promoter in the intergenic region between orf1 and cry8Ea1 controlled by Sigma H factor. Appl. Environ. Microbiol. 2012, 78, 4164–4168. [Google Scholar] [CrossRef] [PubMed]
- Komano, T.; Takabe, S.; Sakai, H. Transcription of the insecticidal crystal protein genes of Bacillus thuringiensis. Biotechnol. Annu. Rev. 2000, 5, 131–154. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, P.; Peng, Q.; Rong, R.; Liu, C.; Lereclus, D.; Zhang, J.; Song, F.; Huang, D. Weak transcription of the cry1Ac gene in nonsporulating Bacillus thuringiensis cells. Appl. Environ. Microbiol. 2012, 78, 6466–6474. [Google Scholar] [CrossRef] [PubMed]
- Celandroni, F.; Salvetti, S.; Senesi, S.; Ghelardi, E. Bacillus thuringiensis membrane-damaging toxins acting on mammalian cells. FEMS Microbiol. Lett. 2014, 361, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Nickerson, K.W. Bacillus thuringiensis HD-73 spores have surface-localized Cry1Ac toxin: Physiological and pathogenic consequences. Appl. Environ. Microbiol. 1996, 6, 3722–3726. [Google Scholar] [CrossRef]
- Berry, C.; O’Neil, S.; Ben-Dov, E.; Jones, A.F.; Murphy, L.; Quail, M.A.; Holden, M.T.G.; Harris, D.; Zaritsky, A.; Parkhill, J. Complete sequence and organization of pBtoxis, the toxin-coding plasmid of Bacillus thuringiensis subsp. israelensis. Appl. Environ. Microbiol. 2002, 68, 5082–5095. [Google Scholar] [CrossRef] [PubMed]
- Stein, C.; Jones, G.W.; Chlamers, T.; Berry, C. Transcriptional analysis of the toxin-coding plasmid pBtoxis from Bacillus thuringiensis subsp. israelensis. Appl. Environ. Microbiol. 2006, 72, 1771–1776. [Google Scholar] [CrossRef]
- Fang, J.; Zhu, Y.; Ju, S.H.; Zhang, R.; Yu, Z.; Sun, M. Promoters of crystal protein genes do not control crystal formation inside exosporium of Bacillus thuringiensis sp. finitimus strain YBT-020. FEMS Microbiol Lett. 2009, 300, 11–17. [Google Scholar] [CrossRef]
- Díaz-Mendoza, M.; Bideshi, D.K.; Federici, B.A. A 54-kilodalton protein encoded by pBtoxis is required for parasporal body structural integrity in Bacillus thuringiensis subsp. israelensis. J. Bacteriol. 2012, 194, 1562–1571. [Google Scholar] [CrossRef]
- Sawaya, M.R.; Cascio, D.; Gingery, M.; Rodriguez, J.; Lukasz, G.; Colletier, J.P.; Messerschmidth, M.M.; Boutet, S.; Koglin, J.E.; Willimas, G.J.; et al. Protein crystal structure obtained at 2.9 Å resolution from injecting bacterial cells into an X-ray free-electron laser beam. Proc. Natl. Acad. Sci. USA 2014, 111, 12769–12774. [Google Scholar] [CrossRef]
- Moar, W.J.; Trumble, J.T.; Federici, B.A. Comparative toxicity of spores and crystals from the NRD-12 and HD-1 strains of Bacillus thuringiensis subsp. kurstaki to neonate beet armyworm (Lepidoptera: Noctuidae). J. Econ. Entomol. 1989, 82, 1593–1603. [Google Scholar] [CrossRef]
- Ibarra, J.E.; Federici, B.A. Parasporal bodies of Bacillus thuringiensis subsp. morrisoni (PG-14) and Bacillus thuringiensis subsp. israelensis are similar in protein composition and toxicity. FEMS Microbiol. Lett. 1986, 34, 79–84. [Google Scholar]
- Padua, L.; Federici, B.A. Development of mutants of the mosquitocidal bacterium Bacillus thuringiensis subsp. morrisoni PG-14 toxic to lepidopterous and dipterous insects. FEMS Microbiol. Lett. 1990, 54, 257–262. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lereclus, D.; Arantes, O.; Chaufaux, J.; Lecadet, M. Transformation and expression of a cloned delta-endotoxin gene in Bacillus thuringiensis. FEMS Microbiol. Lett. 1989, 51, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Monro, R.E. Protein turnover and the formation of protein inclusions during sporulation of Bacillus thuringiensis. Biochem. J. 1961, 81, 225–232. [Google Scholar] [CrossRef]
- Ghosh, A.; Chakrabarti, K.; Chattopadhyay, D. Cloning of feather-degrading minor extracellular protease from Bacillus cereus DCUW: Dissection of the structural domains. Microbiology 2009, 155, 2049–2057. [Google Scholar] [CrossRef]
- Nisnevitch, M.; Sigawi, S.; Cahan, R.; Nitzan, Y. Isolation, characterization and biological role of camelysin from Bacillus thuringiensis subsp. israelensis. Curr. Microbiol. 2010, 61, 76–183. [Google Scholar] [CrossRef]
- Frees, D.; Savijoki, K.; Varmanen, P.; Ingmer, H. Clp ATPases and ClpP proteolytic complexes regulate vital biological processes in low GC, Gram-positive bacteria. Mol. Microbiol. 2007, 63, 1285–1295. [Google Scholar] [CrossRef]
- Moliere, N.; Turgay, K. Chaperone-protease systems in regulation and protein quality control in Bacillus subtilis. Res. Microbiol. 2009, 160, 637–644. [Google Scholar] [CrossRef]
- Chen, D.; Xu, D.; Li, M.; He, J.; Gong, Y.; Wu, D.; Sun, M.; Yu, Z. Proteomic analysis of Bacillus thuringiensis phaC mutant BMB171/PHB(-1) reveals that the PHB synthetic pathway warrants normal carbon metabolism. J. Proteom. 2012, 75, 5176–5188. [Google Scholar] [CrossRef]
- Navarro, A.K.; Farrera, R.R.; Lu, P.R.; Prez-Guevara, F. Relationship between poly-beta-hydroxybutyrate production and deltaendotoxin for Bacillus thuringiensis var. kurstaki. Biotechnol. Lett. 2006, 28, 641–644. [Google Scholar] [CrossRef]
- Wu, D.; He, J.; Gong, Y.; Chen, D.; Zhu, X.; Qiu, N.; Sun, M.; Li, M.; Yu, Z. Proteomic analysis reveals the strategies of Bacillus thuringiensis YBT-1520 for survival under long-term heat stress. Proteomics 2011, 11, 2580–2591. [Google Scholar] [CrossRef]
- Fujita, Y.; Fujita, T. The gluconate operon gnt of Bacillus subtilis encodes its own transcriptional negative regulator. Proc. Natl. Acad. Sci. USA 1987, 84, 4524–4528. [Google Scholar] [CrossRef]
- Nickerson, K.W.; St Julian, G.; Bulla, L.A., Jr. Physiology of spore-forming bacteria associated with insects: Radiorespirometric survey of carbohydrate metabolism in the 12 serotypes of Bacillus thuringiensis. Appl. Microbiol. 1974, 28, 129–132. [Google Scholar] [CrossRef]
- Hsieh, Y.C.; Wu, Y.J.; Chiang, T.Y.; Kuo, C.Y.; Shrestha, K.L.; Chao, C.F.; Huang, Y.C.; Chuankhayan, P.; Wu, W.G.; Li, Y.K.; et al. Crystal structures of Bacillus cereus NCTU2 chitinase complexes with chitooligomers reveal novel substrate binding for catalysis: A chitinase without chitin binding and insertion domains. J. Biol. Chem. 2010, 285, 31603–31615. [Google Scholar] [CrossRef]
- Iakiviak, M.; Mackie, R.I.; Cann, I.K. Functional analyses of multiple lichenin-degrading enzymes from the rumen bacterium Ruminococcus albus 8. Appl. Environ. Microbiol. 2011, 77, 7541–7550. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Magge, A.; Granger, A.C.; Wahome, P.G.; Setlow, B.; Vepachedu, V.R.; Loshon, C.A.; Peng, L.; Chen, D.; Li, Y.Q.; Setlow, P. Role of dipicolinic acid in the germination, stability, and viability of spores of Bacillus subtilis. J. Bacteriol. 2008, 190, 4798–4807. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.L.; Chen, H.J.; Shaw, G.C. Identification and characterization of the Bacillus thuringiensis phaZ gene, encoding new intracellular poly-3-hydroxybutyrate depolymerase. J. Bacteriol. 2006, 188, 7592–7599. [Google Scholar] [CrossRef] [PubMed]
- Ireton KJin, S.; Grossman, A.D.; Sonenshein, A.L. Krebs cycle function is required for activation of the Spo0A transcription factorin Bacillus subtilis. Proc. Natl. Acad. Sci. USA 1995, 92, 2845–2849. [Google Scholar] [CrossRef]
- Jin, S.; Sonenshein, A.L. Identification of two distinct Bacillus subtilis citrate synthase genes. J. Bacteriol. 1994, 176, 4669–4679. [Google Scholar] [CrossRef][Green Version]
- Aronson, J.N.; Borris, D.P.; Doerner, J.F.; Akers, E. Gammaaminobutyric acid pathway and modified tricarboxylic acid cycle activity during growth and sporulation of Bacillus thuringiensis. Appl. Microbiol. 1975, 30, 489–492. [Google Scholar] [CrossRef]
- Lohman, J.R.; Olson, A.C.; Remington, S.J. Atomic resolution structures of Escherichia coli and Bacillus anthracis malate synthase A: Comparison with isoform G and implications for structure-based drug discovery. Protein Sci. 2008, 17, 1935–1945. [Google Scholar] [CrossRef]
- Zhu, L.; Peng, Q.; Song, F.; Jiang, Y.; Sun, C.; Zhang, J.; Huang, D. Structure and regulation of the gab gene cluster, involved in the gamma-aminobutyric acid shunt, are controlled by a sigma54 factor in Bacillus thuringiensis. J. Bacteriol. 2010, 192, 346–355. [Google Scholar] [CrossRef]
- Park, K.B.; Oh, S.H. Enhancement of gamma-aminobutyric acid production in Chungkukjang by applying a Bacillus subtilis strainexpressing glutamate decarboxylase from Lactobacillus brevis. Biotechnol.Lett 2006, 28, 1459–1463. [Google Scholar] [CrossRef]
- Lang, D.R.; Felix, J.; Lundgren, D.G. Development of a membrane-bound respiratory system prior to and during sporulation in Bacillus cereus and its relationship to membrane structure. J. Bacteriol. 1972, 110, 968–977. [Google Scholar] [CrossRef]
- Munro, A.W.; Girvan, H.M.; McLean, K.J. Cytochrome P450–redox partner fusion enzymes. Biochim. Biophys. Acta. 2007, 1770, 345–359. [Google Scholar] [CrossRef]
- Jensen, P.R.; Michelsen, O. Carbon and energy metabolism of atp mutants of Escherichia coli. J. Bacteriol. 1992, 174, 7635–7641. [Google Scholar] [CrossRef] [PubMed]
- Santana, M.; Ionescu, M.S.; Vertes, A.; Longin, R.; Kunst, F.; Danchin, A.; Glaser, P. Bacillus subtilis F0F1 ATPase: DNA sequence of the atp operon and characterization of atp mutants. J. Bacteriol. 1994, 176, 6802–6811. [Google Scholar] [CrossRef] [PubMed]
- Paidhungat, M.; Setlow, P. Spore germination and outgrowth. In Bacillus subtilis and Its Relatives: From Genes to Cells; Onenshein, A.L., Hoch, J.A., Losick, R., Eds.; American Society for Microbiology: Washington, WA, USA, 2002; pp. 537–548. [Google Scholar]
- Paredes-Sabja, D.; Setlow, P.; Sarker, M.R. Germination of spores of Bacillales and Clostridiales species: Mechanisms and proteins involved. Trends Microbiol. 2011, 19, 85–94. [Google Scholar] [CrossRef]
- Behravan, J.; Chirakkal, H.; Masson, A.; Moir, A. Mutations in the gerP locus of Bacillus subtilis and Bacillus cereus affect access of germinants to their targets in spores. J. Bacteriol. 2000, 182, 1987–1994. [Google Scholar] [CrossRef] [PubMed]
- Moir, A.; Cooper, G. Spore germination. Microbiol. Spectr. 2015, 3, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Setlow, P.; Wang SHLi, Y.Q. Germination of spores of the orders Bacillales and Clostridiales. Annu. Rev. Microbiol. 2017, 71, 459–477. [Google Scholar] [CrossRef] [PubMed]
- Kaieda, S.; Setlow, B.; Setlow, P.; Halle, B. Mobility of core water in Bacillus subtilis spores by 2H NMR. Biophys. J. 2013, 105, 2016–2123. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.N.; McBride, S.M. Initiation of sporulation in Clostridium difficile: A twist on the classic model. FEMS Microbiol. Lett. 2014, 358, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Christie, G.; Setlow, P. Bacillus spore germination: Knowns, unknowns and what we need to learn. Cell. Signal. 2020, 74, 109729. [Google Scholar] [CrossRef]
- Setlow, P. Spore germination. Curr. Opin. Microbiol. 2003, 6, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Cortezzo, D.E.; Setlow, B.; Setlow, P. Analysis of the action of compounds that inhibit the germination of spores of Bacillus species. J. Appl. Microbiol. 2004, 96, 725–741. [Google Scholar] [CrossRef]
- Chen, D.; Huang, S.S.; Li, Y.Q. Real-time detection of kinetic germination and heterogeneity of single Bacillus spores by laser tweezers Raman spectroscopy. Anal. Chem. 2006, 78, 6936–6941. [Google Scholar] [CrossRef]
- Shah, I.M.; Laaberki, M.H.; Popham, D.L.; Dworkin, J. A eukaryotic Ser/Thr kinase signals bacteria to exit dormancy in response to peptidoglycan fragments. Cell 2008, 135, 486–496. [Google Scholar] [CrossRef]
- Ramírez-Peralta, A.; Zhang, P.; Li, Y.; Setlow, P. Effects of sporulation conditions on the germination and germination protein levels of Bacillus subtilis spores. Appl. Environ. Microbiol. 2012, 78, 2689–2697. [Google Scholar] [CrossRef]
- Setlow, P. When the sleepers wake: The germination of spores of Bacillus species. J. Appl. Microbiol. 2013, 115, 1251–1268. [Google Scholar] [CrossRef]
- Setlow, P. Germination of spores of Bacillus species: What we know and do not know. J. Bacteriol. 2014, 196, 1297–1305. [Google Scholar] [CrossRef]
- Knudsen, S.M.; Cermak, N.; Delgado, F.F.; Setlow, B.; Setlow, P.; Manalis, S.R. Water and small-molecule permeation of dormant Bacillus subtilis spores. J. Bacteriol. 2016, 198, 168–177. [Google Scholar] [CrossRef]
- Hudson, K.D.; Corfe, B.M.; Kemp, E.H.; Coote, P.J.; Moir, A. Localization of GerAA and GerAC germination proteins in the Bacillus subtilis spore. J. Bacteriol. 2001, 183, 4317–4322. [Google Scholar] [CrossRef]
- Wang Yi, X.; Li, Y.Q.; Setlow, P. Germination of individual Bacillus subtilis spores with alterations in the GerD and SpoVA proteins, which are important in spore germination. J. Bacteriol. 2011, 193, 2301–2311. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, K.K.; Zhang, J.; Cowan, A.E.; Yu, J.; Setlow, P. Germination proteins in the inner membrane of dormant Bacillus subtilis spores colocalize in a discrete cluster. Mol. Microbiol. 2011, 81, 1061–1077. [Google Scholar] [CrossRef]
- Cooper, G.R.; Moir, A. Amino acid residues in the GerAB protein important in the function and assembly of the alanine spore germination receptor of Bacillus subtilis 168. J. Bacteriol. 2011, 193, 2261–2267. [Google Scholar] [CrossRef] [PubMed]
- Luu, H.; Akoachere, M.; Patra, M.; Abel-Santos, E. Cooperativity and interference of germination pathways in Bacillus anthracis spores. J. Bacteriol. 2011, 193, 4192–4198. [Google Scholar] [CrossRef]
- Ramírez-Peralta, A.; Gupta, S.; Butzin, X.Y.; Setlow, B.; Korza, G.; Leyva-Vázquez, M.-A.; Christie, G.; Setlow, P. Identification of new proteins that modulate the germination of spores of Bacillus species. J. Bacteriol. 2013, 95, 3009–3021. [Google Scholar] [CrossRef]
- Korza, G.; Setlow, P. Topology and accessibility of germination proteins in the Bacillus subtilis spore inner membrane. J. Bacteriol. 2013, 195, 1484–1491. [Google Scholar] [CrossRef] [PubMed]
- Cowan, A.E.; Olivastro, E.M.; Koppel, D.E.; Loshon, C.A.; Setlow, B.; Setlow, P. Lipids in the inner membrane of dormant spores of Bacillus species are immobile. Proc. Natl. Acad. Sci. USA 2004, 101, 7733–7738. [Google Scholar] [CrossRef] [PubMed]
- Paidhungat, M.; Ragkousi, K.; Setlow, P. Genetic requirements for induction of germination of spores of Bacillus subtilis by Ca+dipicolinate. J. Bacteriol. 2001, 183, 4886–4893. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, B.M.; Setlow, B.; Setlow, P. Levels of H+ and other monovalent cations in dormant and germinated spores of Bacillus megaterium. J. Bacteriol. 1981, 148, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Chen, D.; Setlow, P.; Li, Y.Q. Elastic and inelastic light scattering from single bacterial spores in an optical trap allows the monitoring of spore germination dynamics. Anal. Chem. 2009, 81, 4035–4042. [Google Scholar] [CrossRef]
- Huang, S.S.; Chen, D.; Pelczar, P.L.; Vepachedu, V.R.; Setlow, P.; Li, Y.Q. Levels of Ca-dipicolinic acid in individual Bacillus spores determined using microfluidic Raman tweezers. J. Bacteriol. 2007, 189, 4681–4687. [Google Scholar] [CrossRef]
- Wang, S.; Setlow, P.; Li, Y.Q. Slow leakage of Ca-dipicolinic acid from individual Bacillus spores during initiation of spore germination. J. Bacteriol. 2015, 197, 1095–1103. [Google Scholar] [CrossRef]
- Clements, M.O.; Moir, A. Role of the gerI operon of Bacillus cereus 569 in the response of spores to germinants. J. Bacteriol. 1998, 180, 6729–6735. [Google Scholar] [CrossRef] [PubMed]
- Aspholm, M.; Borch-Pedersen, K.; O’Sullivan, K.; Fiellheim, S.; Bj(ornson, A.I.H.; Granum, P.E.; Lindbäck, T. Importance of individual germination receptor subunits in the cooperative function between GerA and Ynd. J. Bacteriol. 2019, 201, e00451-19. [Google Scholar] [CrossRef]
- Mongkolthanaruk, W.; Cooper, G.R.; Mawer, J.S.; Allan, R.N.; Moir, A. Effect of amino acid substitutions in the GerAA protein on the function of the alanine-responsive germinant receptor of Bacillus subtilis spores. J. Bacteriol. 2011, 193, 2268–2275. [Google Scholar] [CrossRef]
- Ragkousi, K.; Eichenberger, P.; Van Ooij, C.; Setlow, P. Identification of a new gene essential for germination of Bacillus subtilis spores with Ca2+-dipicolinate. J. Bacteriol. 2003, 185, 2315–2329. [Google Scholar] [CrossRef]
- Paidhungar, M.; Setlow, P. Isolation and characterization of mutations in Bacillus subtilis that allow spore germination in the novel germinant D-alanine. J. Bacteriol. 1999, 181, 3341–3350. [Google Scholar] [CrossRef]
- Xiao, Y.; Francke, C.; Abee, T.; Wells-Bennik, M.H. Clostridial spore germination versus bacilli: Genome mining and current insights. Food Microbiol. 2011, 28, 266–274. [Google Scholar] [CrossRef]
- Gai, Y.; Liu, G.; Tan, H. Identification and characterization of a germination operon from Bacillus thuringiensis. Antonie Van Leeuwenhoek 2006, 89, 251–259. [Google Scholar]
- Li, Y.; Butzin, X.Y.; Davis, A.; Setlow, B.; Korza, G.; Ustok, F.I.; Christie, G.; Setlow, P.; Hao, B. Activity and regulation of various forms of CwlJ, SleB, and YpeB proteins in degrading cortex peptidoglycan of spores of Bacillus species in vitro and during spore germination. J. Bacteriol. 2013, 195, 2530–2540. [Google Scholar] [PubMed]
- Yang, J.; Peng, Q.; Chen, Z.; Deng, C.H.; Shu, C.H.; Zhang, J.; Huang, D.; Song, F. Transcriptional Regulation and Characteristics of a Novel N-Acetylmuramoyl-L-Alanine Amidase Gene Involved in Bacillus thuringiensis Mother Cell Lysis. J Bacteriol. 2013, 195, 2887–2897. [Google Scholar] [PubMed]
- Segev, E.; Rosenberg, A.; Mamou, G.; Sinai, L.; Ben-Yehuda, S. Molecular kinetics of reviving bacterial spores. J. Bacteriol. 2013, 195, 1875–1882. [Google Scholar] [CrossRef] [PubMed]
- Heffron, J.D.; Lambert, E.A.; Sherry Nm Popham, D.L. Contributions of four cortex lytic enzymes to germination of Bacillus anthracis spores. J. Bacteriol. 2010, 192, 763–770. [Google Scholar] [CrossRef]
- Yi, X.; Setlow, P. Studies of the commitment step in the germination of spores of Bacillus species. J. Bacteriol. 2010, 192, 3424–3433. [Google Scholar] [CrossRef]
- Yi, X.; Liu, J.; Faeder, J.R.; Setlow, P. Synergism between different germinant receptors in the germination of Bacillus subtilis spores. J. Bacteriol. 2011, 193, 4664–4671. [Google Scholar] [CrossRef]
- Stewart, G.S.A.; Johnstone, K.B.; Hagelberg, F.; Ellar, D.J. Commitment of bacterial spores to germinate: A measure of the trigger reaction. Biochem. J. 1981, 198, 101–106. [Google Scholar] [CrossRef]
- Zhang, J.; Griffiths, K.K.; Cowan, A.; Setlow, P.; Yu, J. Expression level of Bacillus subtilis germinant receptors determines the average rate but not the heterogeneity of spore germination. J Bacteriol 2013, 195, 1735–1740. [Google Scholar] [CrossRef] [PubMed]
- Blinker, S.; Vreede, J.; Setlow, P.; Brul, S. Predicting the structure and dynamics of membrane protein GerAB from Bacillus subtilis. Int. J. Mol. Sci. 2021, 22, 3793. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Gai, Y.; Liang, L.; Liu, G.; Tan, H. A gene encoding alanine racemase is involved in spore germination in Bacillus thuringiensis. Arch. Microbiol. 2007, 187, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Yang, H.; Liu, G.; Tan, H. Identification and characterization of a polysaccharide deacetylase gene from Bacillus thuringiensis. Can. J. Microbiol. 2006, 52, 935–941. [Google Scholar] [CrossRef]
- Liang, L.; Cai, Y.; Hu, K.; Liu, G. The gerA operon is required for spore germination in Bacillus thuringiensis. Wei Sheng Wu Xue Bao 2008, 48, 281–286. [Google Scholar] [PubMed]
- Moir, A. How do spores germinate? J. Appl. Microbiol. 2006, 101, 526–530. [Google Scholar] [CrossRef]
- Setlow, B.; Peng, L.; Loshon, C.A.; Li, Y.; Christie, G.; Setlow, P. Characterization of the germination of Bacillus megaterium spores lacking enzymes that degrade the spore cortex. J. Appl. Microbiol. 2009, 107, 318–328. [Google Scholar]
- Tehri, N.; Kumar, N.; Raghu, H.V.; Thakur, G.; Kumar-Sharma, K. Role of stereospecific nature of germinants in Bacillus megaterium spores germination. J. Biotech. 2017, 7, 259. [Google Scholar] [CrossRef]
- Setlow, P.; Christie, G. What’s new and notable in bacterial spore killing! World J. Microbiol. Biotechnol. 2021, 37, 144. [Google Scholar] [CrossRef]
- Andryukov, B.G.; Karpenko, A.A.; Lyapun, I.N. Learning from nature: Bacterial spores as a target for current technologies in medicine. Sovrem. Tekhnologii Med. 2021, 12, 105–122. [Google Scholar] [CrossRef]
- Moreno-Fierros, L.; García, N.; Gutiérrez, R.; López-Revilla, R.; Vázquez-Padrón, R.I. Intranasal, rectal and intraperitoneal immunization with protoxin Cry1Ac from Bacillus thurengiensis induces compartmentalized serum, intestinal, vaginal and pulmonary immune responses in Balb/c mice. Microbes Infect. 2000, 2, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Favela-Hernández, J.M.; Balderas, R.I.; Guerrero, G.G. The potential of a commercial product based on Bacillus thuringiensis Cry1A-Cry2A as a immunogen and adjuvant. Madridge J. Immunol. 2018, 2, 58–64. [Google Scholar] [CrossRef]
- Guerrero, G.G.; Tuero, I. Adjuvant compounds: Friends in vaccine formulations against infectious diseases. Hum. Vaccines Immunother. 2021, 17, 3539–3550. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Yu, Q.; Song, F. Expression of cry genes in Bacillus thuringiensis biotechnology. Appl. Microbiol. Biotechnol. 2019, 103, 1617–1626. [Google Scholar] [CrossRef] [PubMed]
- Ibuki, T.; Iwasawa, S.; Lian, A.A.; Lye, P.Y.; Maruta, R.; Asano, S.I.; Kotani, E.; Mori, H. Development of a cypovirus proteins microcrystal-encapsulated Bacillus thuringiensis UV-tolerant and mosqutocidal delta-endotoxin. Biol. Open. 2022, 11, bio059363. [Google Scholar] [CrossRef]
- Wang, F.F.; Qu, S.X.; Lin, J.S.H.; Li, H.P.; Hou, L.J.; Jiang, N.; Luo, X.; Ma, L. Identification of Cyt2Ba from a new strain of Bacillus thuringiensis and its toxicity in Bradysia difformis. Curr. Microbiol. 2020, 77, 2859–2866. [Google Scholar]
- El-Khoury, N.; Majad, R.; Perchat, S.; Kallasay, M.; Lereclus, D.; Gohar, M. Spatio-temporal evolution of sporulation in Bacillus thuringiensis biofilm. Front. Microb. 2016, 7, 1222. [Google Scholar] [CrossRef]
- Jouzani, G.S.; Valijanina, E.; Sharafi, R. Bacillus thuringiensis: A successful insecticide with new environmental features and tidings. Appl. Microbiol. Biotechnol. 2017, 101, 2691–2711. [Google Scholar] [CrossRef]
- Tetreau, G.; Andreeva, E.A.; Banneville, A.S.; De Zitter, E.; Colletier, J.P. Can (we make) Bacillus thuringiensis crystallize more than its toxins? Toxins 2021, 13, 441–456. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).