
Irradiation Induces Tuft Cell Hyperplasia and Myenteric Neuronal Loss in the Absence of Dietary Fiber in a Mouse Model of Pelvic Radiotherapy

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Irradiation Procedure
2.3. Diets
2.4. Experimental Design
2.5. Tissue Harvesting
2.6. Histology and Immunohistochemistry
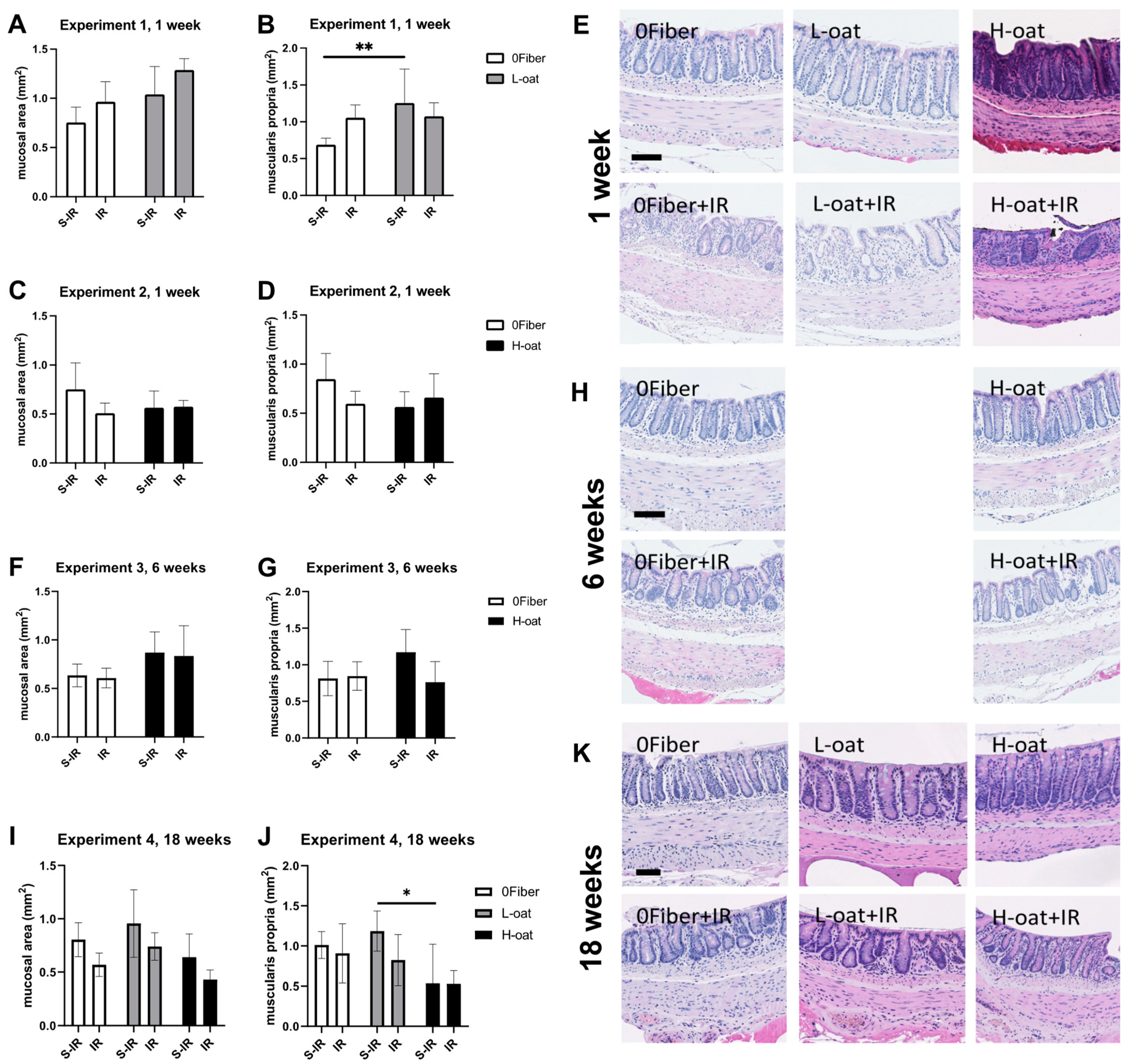
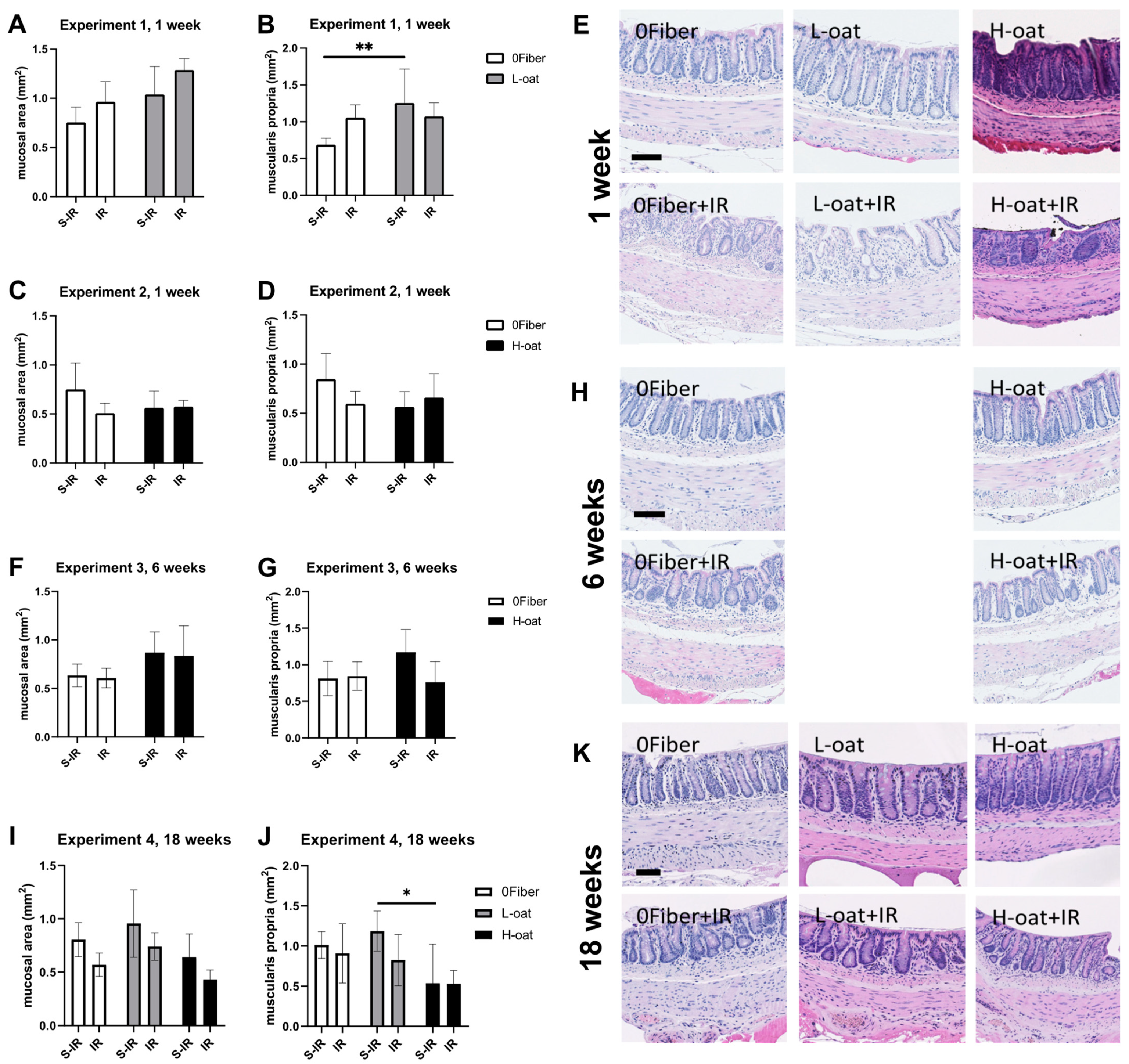
2.7. Morphology
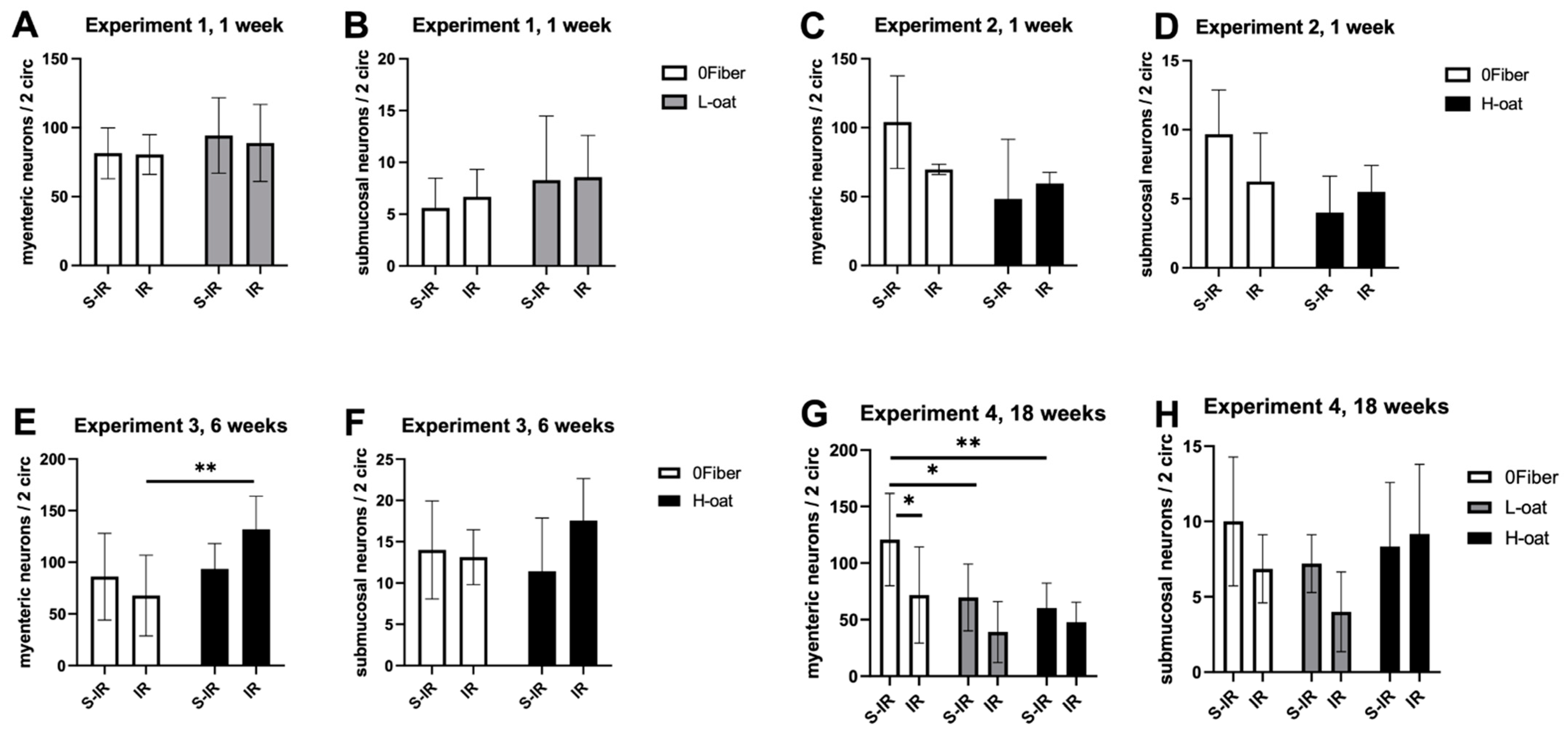
2.8. Neuronal Numbers
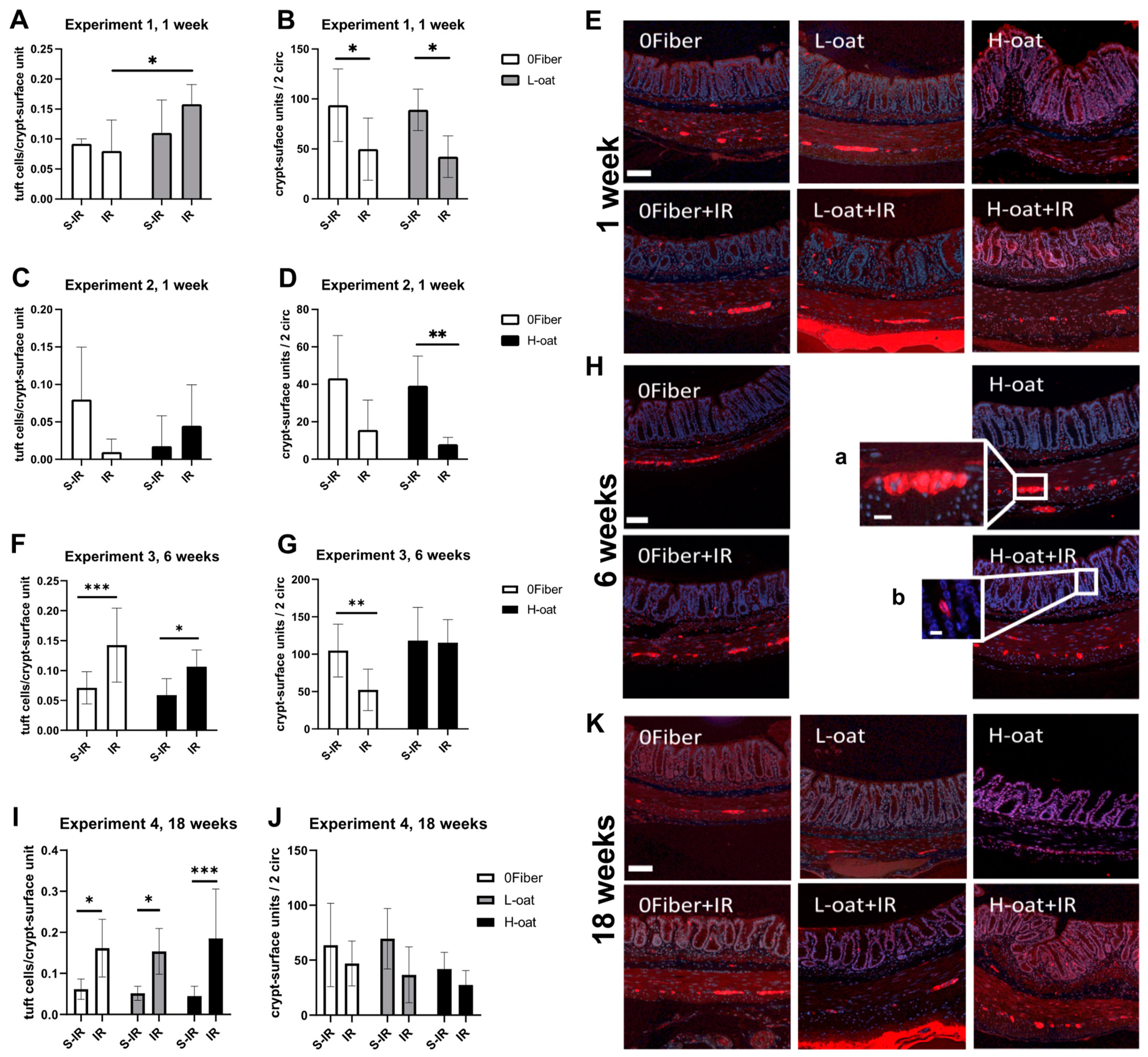
2.9. Tuft Cells and Crypt-Surface Unit Numbers
2.10. Statistical Analyses
3. Results
3.1. General Observations
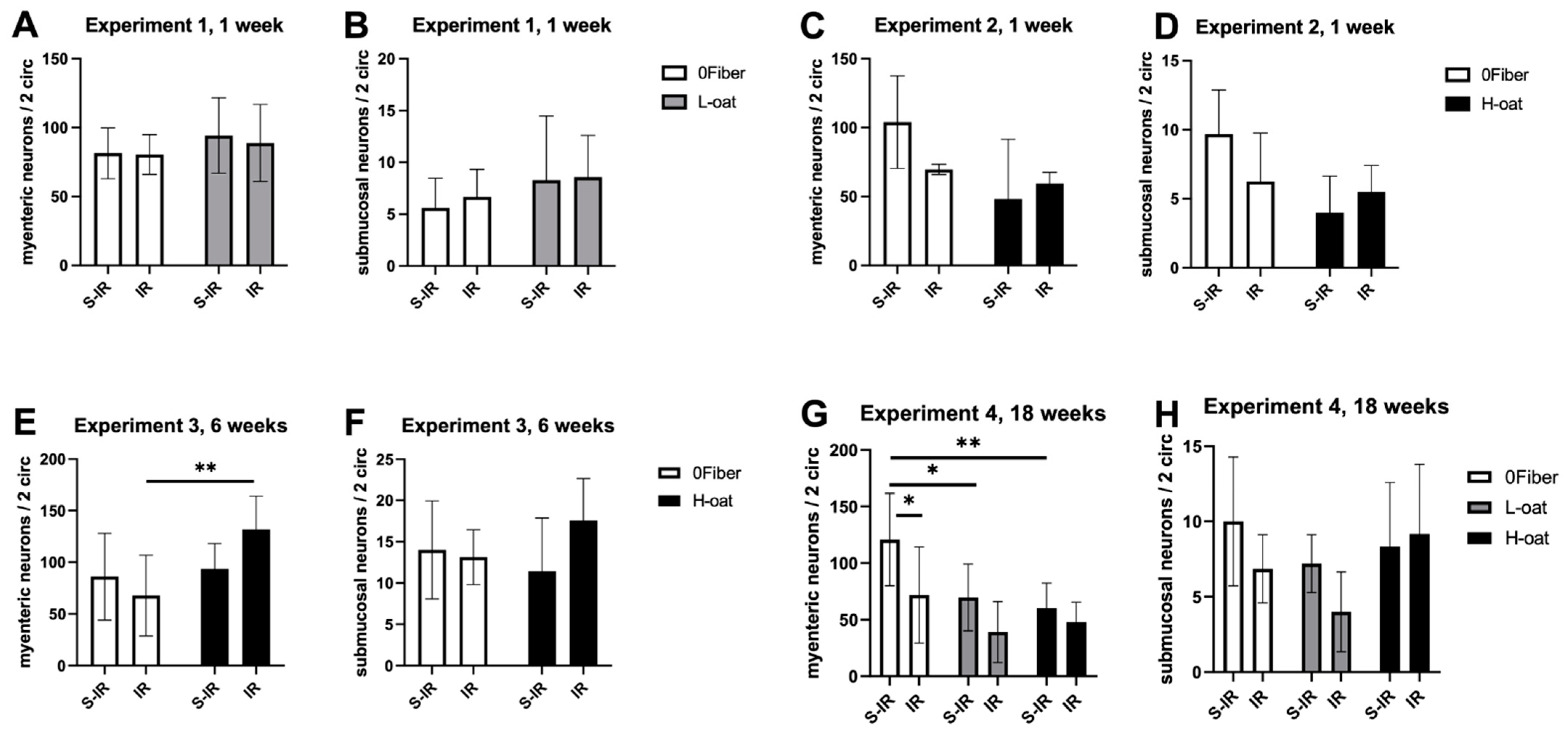
3.2. Myenteric Neurons
3.3. Submucosal Neurons
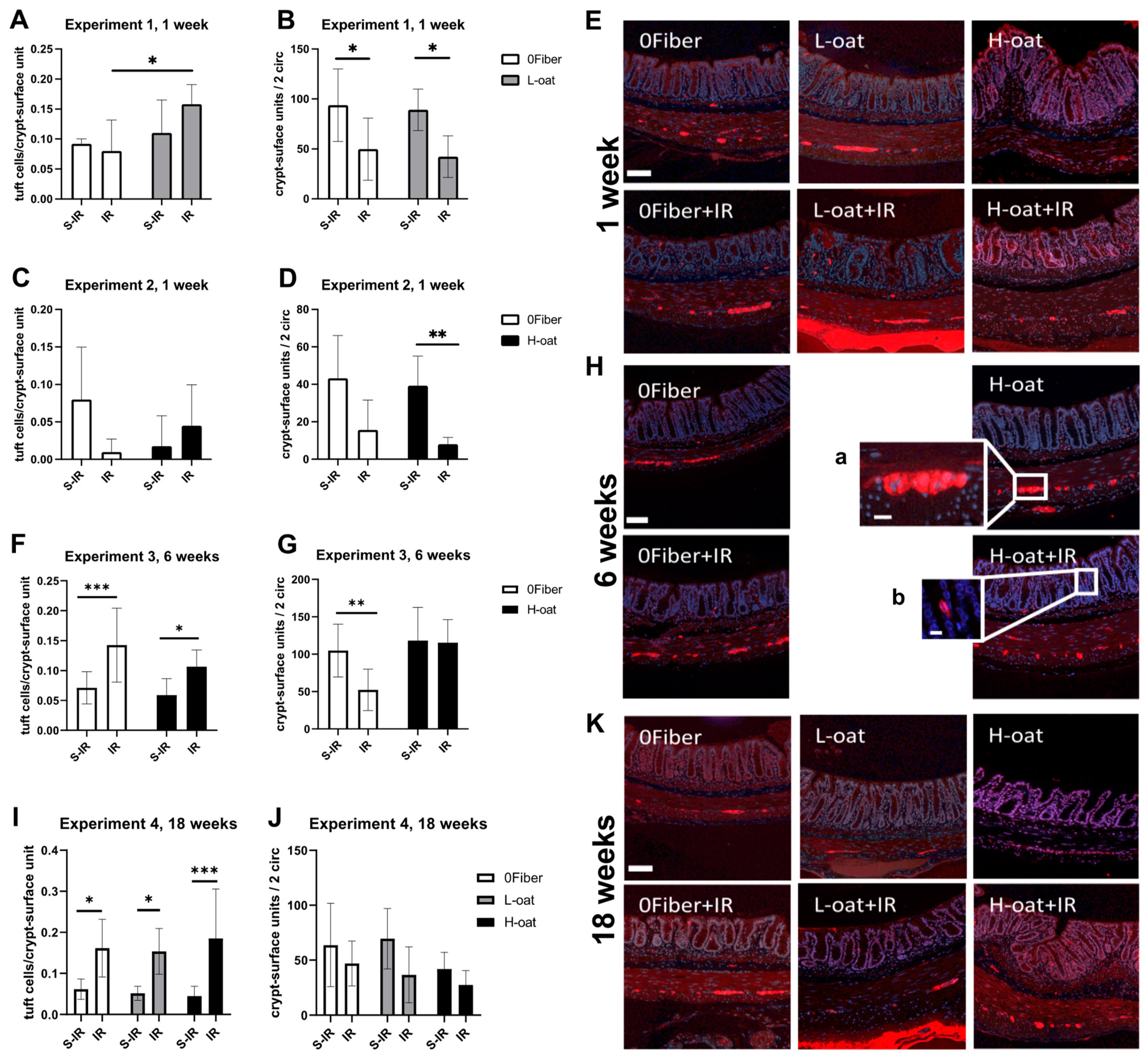
3.4. Tuft Cells
3.5. Crypt-Surface Units
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andreyev, H.J.N.; Wotherspoon, A.; Denham, J.W.; Hauer-Jensen, M. Defining pelvic-radiation disease for the survivorship era. Lancet Oncol. 2010, 11, 310–312. [Google Scholar] [CrossRef]
- Steineck, G.; Skokic, V.; Sjöberg, F.; Bull, C.; Alevronta, E.; Dunberger, G.; Bergmark, K.; Wilderäng, U.; Oh, J.H.; Deasy, J.O.; et al. Identifying radiation-induced survivorship syndromes affecting bowel health in a cohort of gynecological cancer survivors. PLoS ONE 2017, 12, e0171461. [Google Scholar] [CrossRef] [PubMed]
- Andreyev, H. Gastrointestinal Problems after Pelvic Radiotherapy: The Past, the Present and the Future. Clin. Oncol. 2007, 19, 790–799. [Google Scholar] [CrossRef] [PubMed]
- McGough, C.; Baldwin, C.; Frost, G.; Andreyev, H.J.N. Role of nutritional intervention in patients treated with radiotherapy for pelvic malignancy. Br. J. Cancer 2004, 90, 2278–2287. [Google Scholar] [CrossRef] [PubMed]
- Stacey, R.; Green, J.T. Radiation-induced small bowel disease: Latest developments and clinical guidance. Ther. Adv. Chronic Dis. 2014, 5, 15–29. [Google Scholar] [CrossRef] [Green Version]
- Wedlake, L.; Shaw, C.; McNair, H.; Lalji, A.; Mohammed, K.; Klopper, T.; Allan, L.; Tait, D.; Hawkins, M.A.; Somaiah, N.; et al. Randomized controlled trial of dietary fiber for the prevention of radiation-induced gastrointestinal toxicity during pelvic radiotherapy. Am. J. Clin. Nutr. 2017, 106, 849–857. [Google Scholar] [CrossRef]
- Hedelin, M.; Skokic, V.; Wilderäng, U.; Ahlin, R.; Bull, C.; Sjöberg, F.; Dunberger, G.; Bergmark, K.; Stringer, A.; Steineck, G. Intake of citrus fruits and vegetables and the intensity of defecation urgency syndrome among gynecological cancer survivors. PLoS ONE 2019, 14, e0208115. [Google Scholar] [CrossRef]
- Jakobsdottir, G.; Nyman, M.; Fåk, F. Designing future prebiotic fiber to target metabolic syndrome. Nutrition 2014, 30, 497–502. [Google Scholar] [CrossRef]
- El Khoury, D.; Cuda, C.; Luhovyy, B.L.; Anderson, G.H. Beta glucan: Health benefits in obesity and metabolic syndrome. J Nutr. Metab. 2012, 2012, 851362. [Google Scholar] [CrossRef] [Green Version]
- Ramakers, J.D.; Volman, J.J.; Biorklund, M.; Onning, G.; Mensink, R.P.; Plat, J. Fecal water from ileostomic patients consuming oat beta-glucan enhances immune responses in enterocytes. Mol. Nutr. Food Res. 2007, 51, 211–220. [Google Scholar] [CrossRef]
- Wilczak, J.; Błaszczyk, K.; Kamola, D.; Gajewska, M.; Harasym, J.P.; Jałosińska, M.; Gudej, S.; Suchecka, D.; Oczkowski, M.; Gromadzka-Ostrowska, J. The effect of low or high molecular weight oat beta-glucans on the inflammatory and oxidative stress status in the colon of rats with LPS-induced enteritis. Food Funct. 2015, 6, 590–603. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.-H.; Estrada, A.; Van Kessel, A.; Park, B.-C.; Laarveld, B. β-Glucan, extracted from oat, enhances disease resistance against bacterial and parasitic infections. FEMS Immunol. Med. Microbiol. 2003, 35, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.M. CODEX-aligned dietary fiber definitions help to bridge the fiber gap. Nutr. J. 2014, 13, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veronese, N.; Solmi, M.; Caruso, M.G.; Giannelli, G.; Osella, A.R.; Evangelou, E.; Maggi, S.; Fontana, L.; Stubbs, B.; Tzoulaki, I. Dietary fiber and health outcomes: An umbrella review of systematic reviews and meta-analyses. Am. J. Clin. Nutr. 2018, 107, 436–444. [Google Scholar] [CrossRef] [Green Version]
- Hidaka, A.; Harrison, T.A.; Cao, Y.; Sakoda, L.C.; Barfield, R.; Giannakis, M.; Song, M.; Phipps, A.I.; Figueiredo, J.C.; Zaidi, S.H.; et al. Intake of Dietary Fruit, Vegetables, and Fiber and Risk of Colorectal Cancer According to Molecular Subtypes: A Pooled Analysis of 9 Studies. Cancer Res. 2020, 80, 4578–4590. [Google Scholar] [CrossRef]
- King, D.E.; Mainous, A.; Lambourne, C.A. Trends in Dietary Fiber Intake in the United States, 1999-2008. J. Acad. Nutr. Diet. 2012, 112, 642–648. [Google Scholar] [CrossRef]
- Cheng, X.; Voss, U.; Ekblad, E. Tuft cells: Distribution and connections with nerves and endocrine cells in mouse intestine. Exp. Cell Res. 2018, 369, 105–111. [Google Scholar] [CrossRef]
- Middelhoff, M.; Westphalen, C.B.; Hayakawa, Y.; Yan, K.S.; Gershon, M.D.; Wang, T.C.; Quante, M. Dclk1-expressing tuft cells: Critical modulators of the intestinal niche? Am. J. Physiol. Liver Physiol. 2017, 313, G285–G299. [Google Scholar] [CrossRef] [Green Version]
- Westphalen, C.B.; Asfaha, S.; Hayakawa, Y.; Takemoto, Y.; Lukin, D.J.; Nuber, A.H.; Brandtner, A.; Setlik, W.; Remotti, H.; Muley, A.; et al. Long-lived intestinal tuft cells serve as colon cancer–initiating cells. J. Clin. Investig. 2014, 124, 1283–1295. [Google Scholar] [CrossRef] [Green Version]
- Gerbe, F.; Sidot, E.; Smyth, D.J.; Ohmoto, M.; Matsumoto, I.; Dardalhon, V.; Cesses, P.; Garnier, L.; Pouzolles, M.; Brulin, B.; et al. Intestinal epithelial tuft cells initiate type 2 mucosal immunity to helminth parasites. Nature 2016, 529, 226–230. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.; O’Leary, C.E.; Locksley, R.M. Regulation of immune responses by tuft cells. Nat. Rev. Immunol. 2019, 19, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; O’Leary, C.E.; von Moltke, J.; Liang, H.-E.; Ang, Q.Y.; Turnbaugh, P.J.; Radhakrishnan, S.; Pellizzon, M.; Ma, A.; Locksley, R.M. A Metabolite-Triggered Tuft Cell-ILC2 Circuit Drives Small Intestinal Remodeling. Cell 2018, 174, 271–284.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May, R.; Qu, D.; Weygant, N.; Chandrakesan, P.; Ali, N.; Lightfoot, S.A.; Li, L.; Sureban, S.M.; Houchen, C.W. Brief Report: Dclk1 Deletion in Tuft Cells Results in Impaired Epithelial Repair After Radiation Injury. Stem Cells 2013, 32, 822–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrakesan, P.; May, R.; Weygant, N.; Qu, D.; Berry, W.; Sureban, S.; Ali, N.; Rao, C.; Huycke, M.; Bronze, M.S.; et al. Intestinal tuft cells regulate the ATM mediated DNA Damage response via Dclk1 dependent mechanism for crypt restitution following radiation injury. Sci. Rep. 2016, 6, 37667. [Google Scholar] [CrossRef]
- Pearce, S.C.; Weber, G.J.; Van Sambeek, D.M.; Soares, J.W.; Racicot, K.; Breault, D.T. Intestinal enteroids recapitulate the effects of short-chain fatty acids on the intestinal epithelium. PLoS ONE 2020, 15, e0230231. [Google Scholar] [CrossRef]
- McKinley, E.T.; Sui, Y.; Al-Kofahi, Y.; Millis, B.A.; Tyska, M.J.; Roland, J.T.; Santamaria-Pang, A.; Ohland, C.L.; Jobin, C.; Franklin, J.L.; et al. Optimized multiplex immunofluorescence single-cell analysis reveals tuft cell heterogeneity. JCI Insight 2017, 2, e93487. [Google Scholar] [CrossRef]
- Nøhr, M.K.; Pedersen, M.H.; Gille, A.; Egerod, K.L.; Engelstoft, M.S.; Husted, A.S.; Sichlau, R.M.; Grunddal, K.V.; Poulsen, S.S.; Han, S.; et al. GPR41/FFAR3 and GPR43/FFAR2 as cosensors for short-chain fatty acids in enteroendocrine cells vs FFAR3 in enteric neurons and FFAR2 in enteric leukocytes. Endocrinology 2013, 154, 3552–3564. [Google Scholar] [CrossRef]
- Soret, R.; Chevalier, J.; De Coppet, P.; Poupeau, G.; Derkinderen, P.; Segain, J.P.; Neunlist, M. Short-Chain Fatty Acids Regulate the Enteric Neurons and Control Gastrointestinal Motility in Rats. Gastroenterology 2010, 138, 1772–1782.e4. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Sakitani, K.; Konishi, M.; Asfaha, S.; Niikura, R.; Tomita, H.; Renz, B.W.; Tailor, Y.; Macchini, M.; Middelhoff, M.; et al. Nerve Growth Factor Promotes Gastric Tumorigenesis through Aberrant Cholinergic Signaling. Cancer Cell 2017, 31, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Middelhoff, M.; Nienhüser, H.; Valenti, G.; Maurer, H.C.; Hayakawa, Y.; Takahashi, R.; Kim, W.; Jiang, Z.; Malagola, E.; Cuti, K.; et al. Prox1-positive cells monitor and sustain the murine intestinal epithelial cholinergic niche. Nat. Commun. 2020, 11, 111–114. [Google Scholar] [CrossRef]
- Bull, C.; Malipatlolla, D.; Kalm, M.; Sjöberg, F.; Alevronta, E.; Grandér, R.; Sultanian, P.; Persson, L.; Boström, M.; Eriksson, Y.; et al. A novel mouse model of radiation-induced cancer survivorship diseases of the gut. Am. J. Physiol. Liver Physiol. 2017, 313, G456–G466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devarakonda, S.; Malipatlolla, D.K.; Patel, P.; Grandér, R.; Kuhn, H.G.; Steineck, G.; Sjöberg, F.; Rascón, A.; Nyman, M.; Eriksson, Y.; et al. Dietary Fiber and the Hippocampal Neurogenic Niche in a Model of Pelvic Radiotherapy. Neuroscience 2021, 475, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Malipatlolla, D.K.; Patel, P.; Sjöberg, F.; Devarakonda, S.; Kalm, M.; Angenete, E.; Lindskog, E.B.; Grandér, R.; Persson, L.; Stringer, A.; et al. Long-term mucosal injury and repair in a murine model of pelvic radiotherapy. Sci. Rep. 2019, 9, 13803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerbe, F.; Brulin, B.; Makrini, L.; Legraverend, C.; Jay, P. DCAMKL-1 Expression Identifies Tuft Cells Rather Than Stem Cells in the Adult Mouse Intestinal Epithelium. Gastroenterology 2009, 137, 2179–2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voss, U.; Sand, E.; Olde, B.; Ekblad, E. Enteric Neuropathy Can Be Induced by High Fat Diet In Vivo and Palmitic Acid Exposure In Vitro. PLoS ONE 2013, 8, e81413. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Boza-Serrano, A.; Turesson, M.F.; Deierborg, T.; Ekblad, E.; Voss, U. Galectin-3 causes enteric neuronal loss in mice after left sided permanent middle cerebral artery occlusion, a model of stroke. Sci. Rep. 2016, 6, 32893. [Google Scholar] [CrossRef]
- Shen, R.L.; Dang, X.Y.; Dong, J.L.; Hu, X.Z. Effects of oat beta-glucan and barley beta-glucan on fecal characteristics, intestinal microflora, and intestinal bacterial metabolites in rats. J. Agric. Food Chem. 2012, 60, 11301–11308. [Google Scholar] [CrossRef]
- Kristek, A.; Wiese, M.; Heuer, P.; Kosik, O.; Schar, M.Y.; Soycan, G.; Alsharif, S.; Kuhnle, G.G.C.; Walton, G.; Spencer, J.P.E. Oat bran, but not its isolated bioactive beta-glucans or polyphenols, have a bifidogenic effect in an in vitro fermentation model of the gut microbiota. Br. J. Nutr. 2019, 121, 549–559. [Google Scholar] [CrossRef]
- Berggren, A.M.; Björck, I.M.E.; Nyman, E.M.G.L.; Eggum, B.O. Short-chain fatty acid content and pH in caecum of rats given various sources of carbohydrates. J. Sci. Food Agric. 1993, 63, 397–406. [Google Scholar] [CrossRef]
- McIntyre, A.; Young, G.; Taranto, T.; Gibson, P.R.; Ward, P.B. Different fibers have different regional effects on luminal contents of rat colon. Gastroenterology 1991, 101, 1274–1281. [Google Scholar] [CrossRef]
- Stark, A.; Nyska, A.; Madar, Z. Metabolic and Morphometric Changes in Small and Large Intestine in Rats Fed High-Fiber Diets. Toxicol. Pathol. 1996, 24, 166–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsui, R.; Kubo, Y.; Sugiura, Y.; Kuwahara, A.; Karaki, S.-I. Fibre-free diet leads to impairment of neuronally mediated muscle contractile response in rat distal colon. Neurogastroenterol. Motil. 2006, 18, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, S.; Tatewaki, M.; Yamada, T.; Fujimiya, M.; Mantyh, C.; Voss, M.; Eubanks, S.; Harris, M.; Pappas, T.; Takahashi, T. Short-chain fatty acids stimulate colonic transit via intraluminal 5-HT release in rats. Am. J. Physiol. Integr. Comp. Physiol. 2003, 284, R1269–R1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Höckerfelt, U.; Franzén, L.; Norrgård, Ö.; Forsgren, S. Early increase and later decrease in VIP and substance P nerve fiber densities following abdominal radiotherapy: A study on the human colon. Int. J. Radiat. Biol. 2002, 78, 1045–1053. [Google Scholar] [CrossRef]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous Bacteria from the Gut Microbiota Regulate Host Serotonin Biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef] [Green Version]
- de Vadder, F.; Grasset, E.; Holm, L.M.; Karsenty, G.; Macpherson, A.J.; Olofsson, L.E.; Bäckhed, F. Gut microbiota regulates maturation of the adult enteric nervous system via enteric serotonin networks. Proc. Natl. Acad. Sci. USA 2018, 115, 6458–6463. [Google Scholar] [CrossRef] [Green Version]
- Otterson, M.F. Effects of radiation upon gastrointestinal motility. World J. Gastroenterol. 2007, 13, 2684–2692. [Google Scholar] [CrossRef] [Green Version]
- Morris, K.A.; Haboubi, N.Y. Pelvic radiation therapy: Between delight and disaster. World J. Gastrointest. Surg. 2015, 7, 279–288. [Google Scholar] [CrossRef]
- Vasina, V.; Barbara, G.; Talamonti, L.; Stanghellini, V.; Corinaldesi, R.; Tonini, M.; De Ponti, F.; de Giorgio, R. Enteric neuroplasticity evoked by inflammation. Auton. Neurosci. 2006, 126-127, 264–272. [Google Scholar] [CrossRef]
- Brierley, S.M.; Jones, R.W.; Gebhart, G.F.; Blackshaw, A. Splanchnic and pelvic mechanosensory afferents signal different qualities of colonic stimuli in mice. Gastroenterology 2004, 127, 166–178. [Google Scholar] [CrossRef]
- Lynch, A.C.; Antony, A.; Dobbs, B.R.; Frizelle, A.F. Bowel dysfunction following spinal cord injury. Spinal Cord 2001, 39, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Ratto, C.; Grillo, E.; Parello, A.; Petrolino, M.; Costamagna, G.; Doglietto, G.B. Sacral Neuromodulation in Treatment of Fecal Incontinence Following Anterior Resection and Chemoradiation for Rectal Cancer. Dis. Colon Rectum 2005, 48, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Hauer-Jensen, M.; Denham, J.; Andreyev, H.J.N. Radiation enteropathy—pathogenesis, treatment and prevention. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 470–479. [Google Scholar] [CrossRef]
- von Moltke, J.; Ji, M.; Liang, H.E.; Locksley, R.M. Tuft-cell-derived IL-25 regulates an intestinal ILC2-epithelial response circuit. Nature 2016, 529, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.; Malipatlolla, D.K.; Devarakonda, S.; Bull, C.; Rascón, A.; Nyman, M.; Stringer, A.; Tremaroli, V.; Steineck, G.; Sjöberg, F. Dietary Oat Bran Reduces Systemic Inflammation in Mice Subjected to Pelvic Irradiation. Nutrients 2020, 12, 2172. [Google Scholar] [CrossRef] [PubMed]
- Yahyapour, R.; Motevaseli, E.; Rezaeyan, A.; Abdollahi, H.; Farhood, B.; Cheki, M.; Rezapoor, S.; Shabeeb, D.; Musa, A.E.; Najafi, M.; et al. Reduction–oxidation (redox) system in radiation-induced normal tissue injury: Molecular mechanisms and implications in radiation therapeutics. Clin. Transl. Oncol. 2018, 20, 975–988. [Google Scholar] [CrossRef]
- Connors, J.; Dawe, N.; Van Limbergen, J. The Role of Succinate in the Regulation of Intestinal Inflammation. Nutrients 2018, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Lei, W.; Ren, W.; Ohmoto, M.; Urban, J.F.; Matsumoto, I.; Margolskee, R.F.; Jiang, P. Activation of intestinal tuft cell-expressed Sucnr1 triggers type 2 immunity in the mouse small intestine. Proc. Natl. Acad. Sci. USA 2018, 115, 5552–5557. [Google Scholar] [CrossRef] [Green Version]
- Yi, J.; Bergstrom, K.; Fu, J.; Shan, X.; McDaniel, J.M.; McGee, S.; Qu, D.; Houchen, C.W.; Liu, X.; Xia, L. Dclk1 in tuft cells promotes inflammation-driven epithelial restitution and mitigates chronic colitis. Cell Death Differ. 2019, 26, 1656–1669. [Google Scholar] [CrossRef]
- Kawashima, R.; Kawamura, Y.I.; Kato, R.; Mizutani, N.; Toyama–Sorimachi, N.; Dohi, T. IL-13 Receptor α2 Promotes Epithelial Cell Regeneration From Radiation–Induced Small Intestinal Injury in Mice. Gastroenterology 2006, 131, 130–141. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| (a) | |||
| Diet Composition (%) | High Oat 15% Fiber | Low Oat 15% Fiber | No Fiber 1 0% Fiber |
| Bioprocessed oat bran | 28.8 | 9.6 | 0 |
| Microcrystalline cellulose | 0 | 10 | 0 |
| Corn starch | 4.7 | 13.9 | 33.5 |
| Basal mixture | 66.5 | 66.5 | 66.5 |
| Total | 100 | 100 | 100 |
| (b) | |||
| Basal Mixture (g/kg of Total Diet in dwb 1) | |||
| Casein | 133 | ||
| DL-Methionine | 2 | ||
| Corn starch | 250 | ||
| Maltodextrin | 87 | ||
| Sucrose | 106 | ||
| Olive oil | 47 | ||
| Vitamin mixture | 10 | ||
| Choline bitartrate | 2 | ||
| TBHQ 2 | 0.01 | ||
| Mineral mixture | 13 | ||
| Calcium phosphate, dibasic | 11 | ||
| Calcium carbonate | 4 | ||
| (c) | |||
 | |||
| Raised Against | Dilution | Source/RRID |
|---|---|---|
| Double-cortin-like kinase −1 (DCLK1) | 1:250 GR296884 1:2000 GR313013 | Abcam ab37994, RRID:AB_873538 |
| Protein gene product 9.5 (PGP9.5) | 1:2000 | Ultraclone RA95101, RRID:AB_2313685 |
| Alexa Fluor 594 anti-rabbit IgG | 1:1000 | Jackson ImmunoResearch Europe Ltd. 711-515-152 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voss, U.; Malipatlolla, D.K.; Patel, P.; Devarakonda, S.; Sjöberg, F.; Grandér, R.; Rascón, A.; Nyman, M.; Steineck, G.; Bull, C. Irradiation Induces Tuft Cell Hyperplasia and Myenteric Neuronal Loss in the Absence of Dietary Fiber in a Mouse Model of Pelvic Radiotherapy. Gastroenterol. Insights 2022, 13, 87-102. https://doi.org/10.3390/gastroent13010010
Voss U, Malipatlolla DK, Patel P, Devarakonda S, Sjöberg F, Grandér R, Rascón A, Nyman M, Steineck G, Bull C. Irradiation Induces Tuft Cell Hyperplasia and Myenteric Neuronal Loss in the Absence of Dietary Fiber in a Mouse Model of Pelvic Radiotherapy. Gastroenterology Insights. 2022; 13(1):87-102. https://doi.org/10.3390/gastroent13010010
Chicago/Turabian StyleVoss, Ulrikke, Dilip Kumar Malipatlolla, Piyush Patel, Sravani Devarakonda, Fei Sjöberg, Rita Grandér, Ana Rascón, Margareta Nyman, Gunnar Steineck, and Cecilia Bull. 2022. "Irradiation Induces Tuft Cell Hyperplasia and Myenteric Neuronal Loss in the Absence of Dietary Fiber in a Mouse Model of Pelvic Radiotherapy" Gastroenterology Insights 13, no. 1: 87-102. https://doi.org/10.3390/gastroent13010010
APA StyleVoss, U., Malipatlolla, D. K., Patel, P., Devarakonda, S., Sjöberg, F., Grandér, R., Rascón, A., Nyman, M., Steineck, G., & Bull, C. (2022). Irradiation Induces Tuft Cell Hyperplasia and Myenteric Neuronal Loss in the Absence of Dietary Fiber in a Mouse Model of Pelvic Radiotherapy. Gastroenterology Insights, 13(1), 87-102. https://doi.org/10.3390/gastroent13010010