
Adenoviruses Encapsulated in PEGylated DOTAP-Folate Liposomes Are Protected from the Pre-Existing Humoral Immune Response

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Cell Lines
2.2. Synthesis of Liposome-Encapsulated Ad-GFP
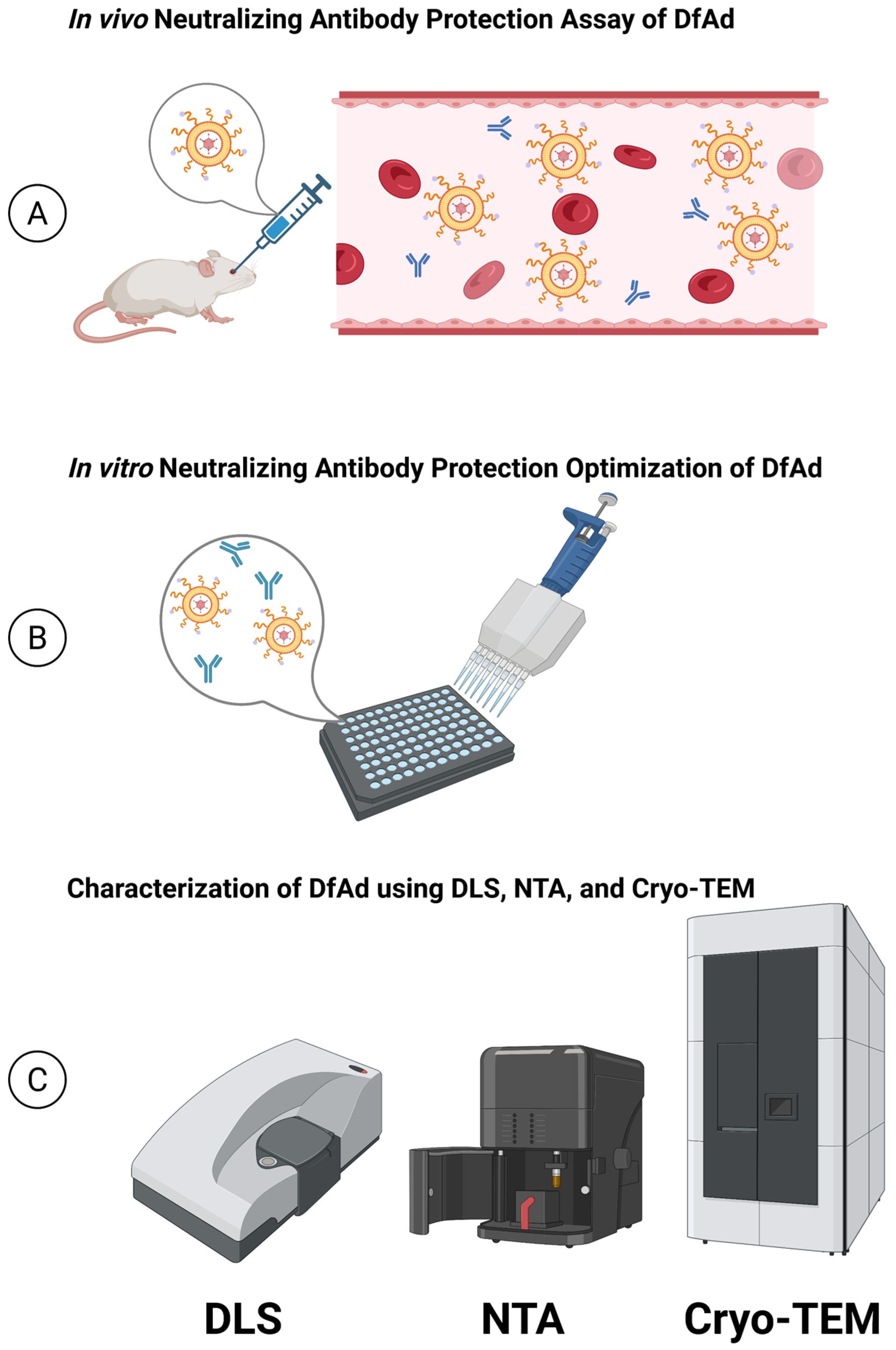
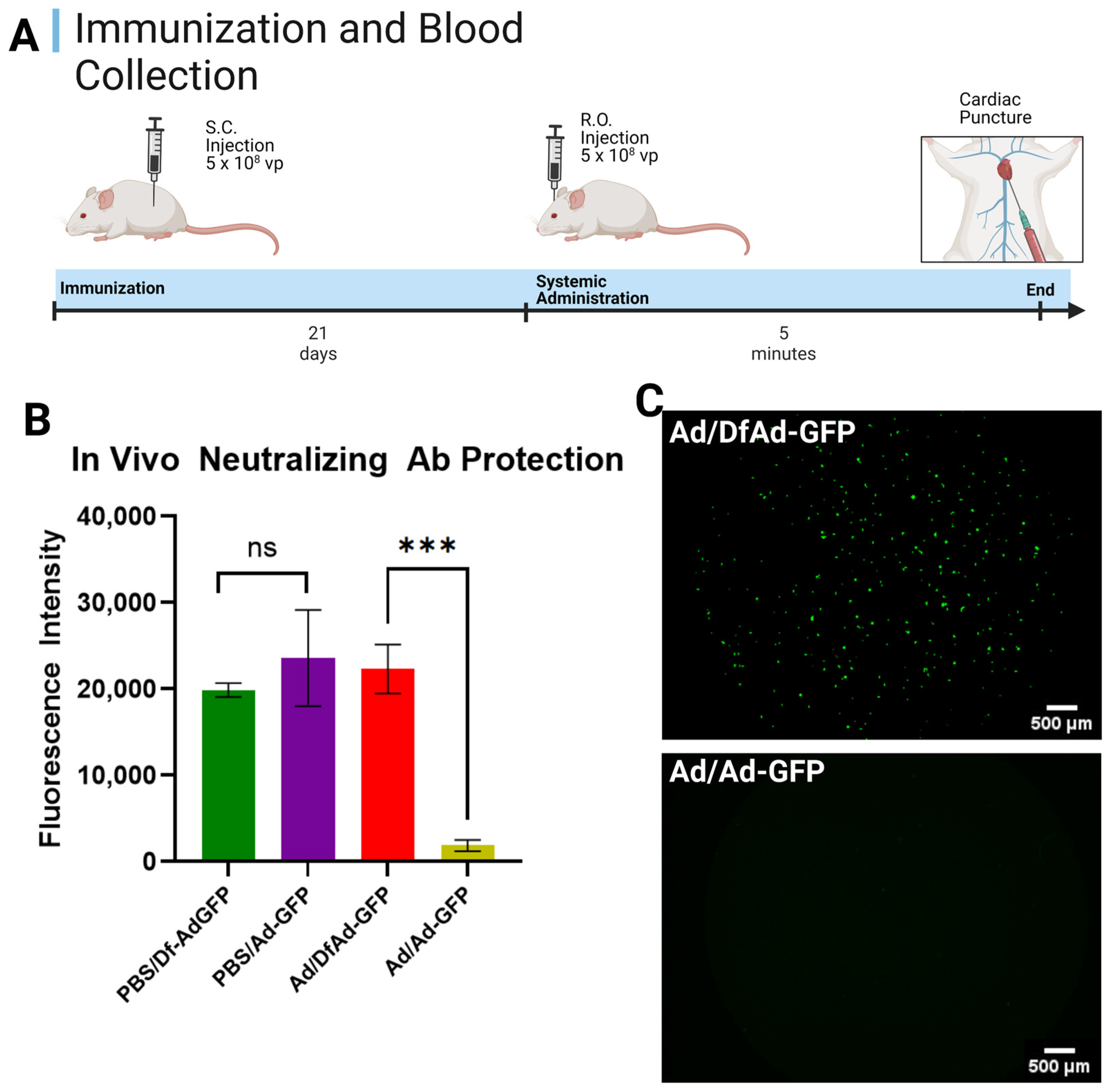
2.3. In Vivo Neutralizing Antibody Protection Assay
2.4. Fluorescence Microscopy
2.5. In Vitro Neutralizing Antibody Protection Assay
2.6. Cryo-Transmission Electron Microscopy (Cryo-TEM)
2.7. Statistical Analysis
3. Results
3.1. In Vivo Neutralizing Antibody Protection Assay
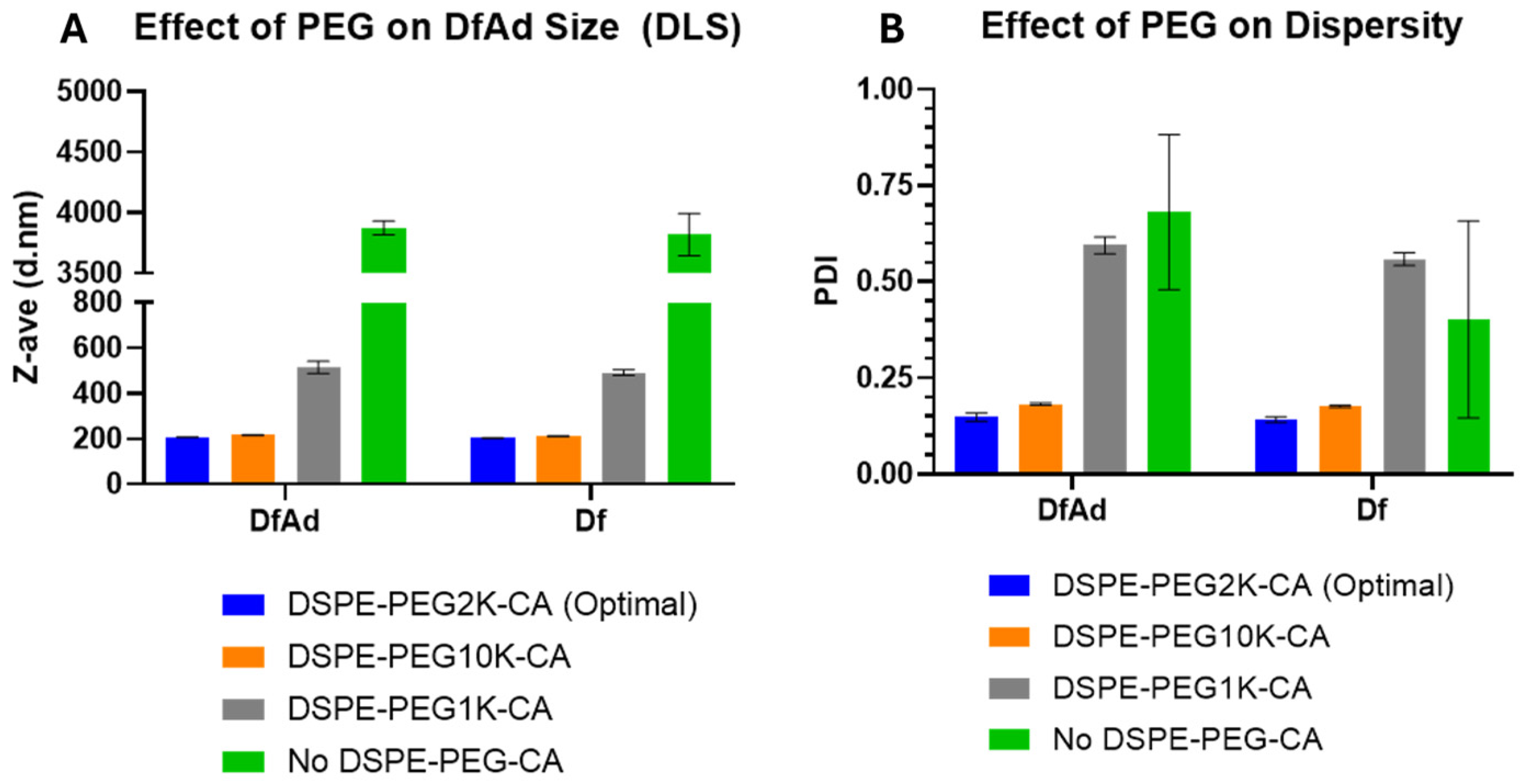
3.2. In Vitro Optimization of DfAd for Protection Against Neutralizing Antibodies
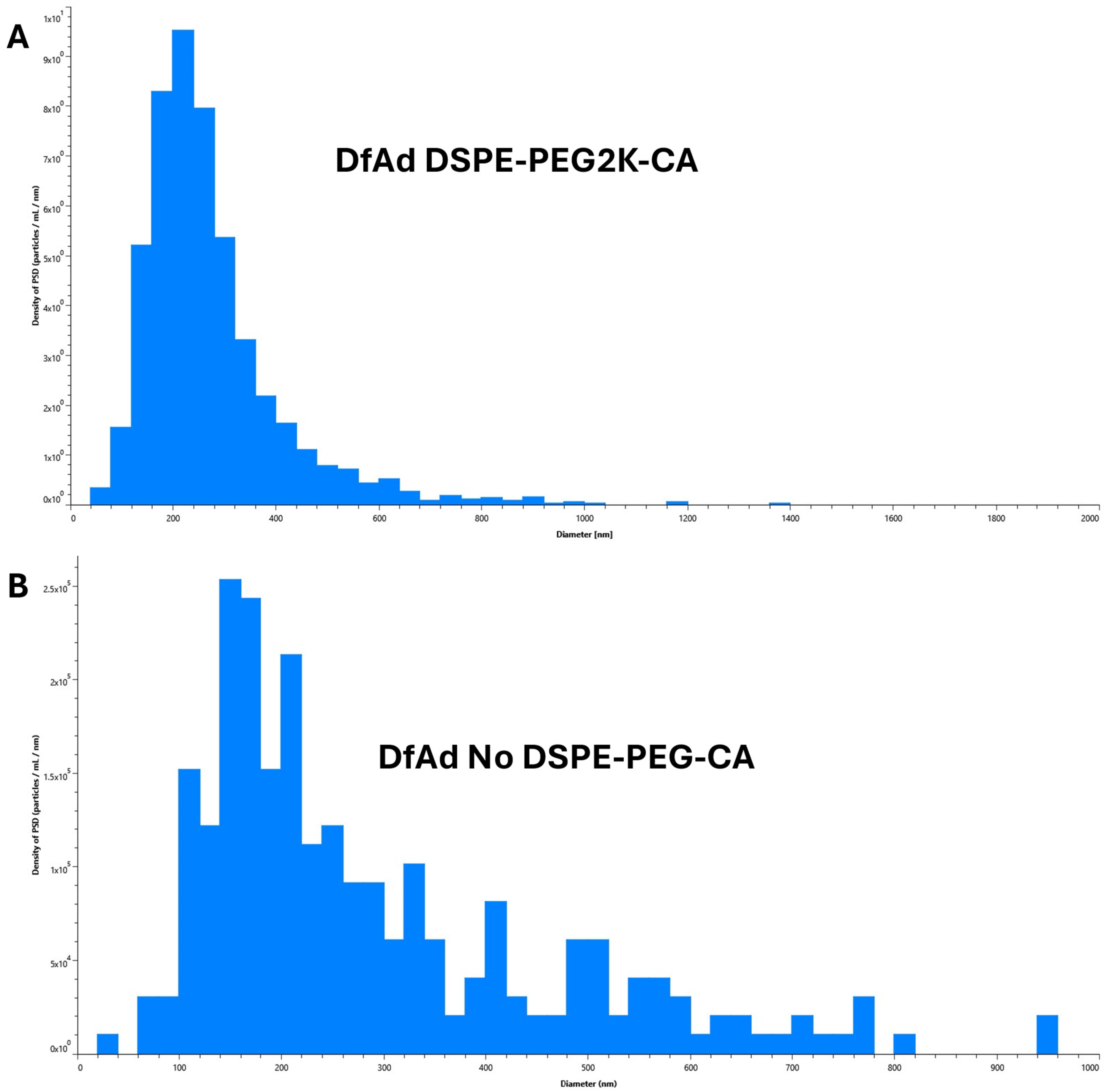
3.3. Characterization of DfAd
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yan, Y.; Xu, H.; Wang, J.; Wu, X.; Wen, W.; Liang, Y.; Wang, L.; Liu, F.; Du, X. Inhibition of breast cancer cells by targeting E2F-1 gene and expressing IL15 oncolytic adenovirus. Biosci. Rep. 2019, 39, BSR20190384. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.E.; Koch, A.; Lauer, U.M.; Hartkopf, A.D. Clinical Trials of Oncolytic Viruses in Breast Cancer. Front. Oncol. 2021, 11, 803050. [Google Scholar] [CrossRef] [PubMed]
- Ang, L.; Guo, L.; Wang, J.; Huang, J.; Lou, X.; Zhao, M. Oncolytic virotherapy armed with an engineered interfering lncRNA exhibits antitumor activity by blocking the epithelial mesenchymal transition in triple-negative breast cancer. Cancer Lett. 2020, 479, 42–53. [Google Scholar] [CrossRef]
- Ishikawa, W.; Kikuchi, S.; Ogawa, T.; Tabuchi, M.; Tazawa, H.; Kuroda, S.; Noma, K.; Nishizaki, M.; Kagawa, S.; Urata, Y.; et al. Boosting Replication and Penetration of Oncolytic Adenovirus by Paclitaxel Eradicate Peritoneal Metastasis of Gastric Cancer. Mol. Ther.-Oncolytics 2020, 18, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Quixabeira, D.C.A.; Jirovec, E.; Pakola, S.; Havunen, R.; Basnet, S.; Santos, J.M.; Kudling, T.V.; Clubb, J.H.A.; Haybout, L.; Arias, V.; et al. Improving the cytotoxic response of tumor-infiltrating lymphocytes towards advanced stage ovarian cancer with an oncolytic adenovirus expressing a human vIL-2 cytokine. Cancer Gene Ther. 2023, 30, 1543–1553. [Google Scholar] [CrossRef]
- Nassiri, F.; Patil, V.; Yefet, L.S.; Singh, O.; Liu, J.; Dang, R.M.A.; Yamaguchi, T.N.; Daras, M.; Cloughesy, T.F.; Colman, H.; et al. Oncolytic DNX-2401 virotherapy plus pembrolizumab in recurrent glioblastoma: A phase 1/2 trial. Nat. Med. 2023, 29, 1370–1378. [Google Scholar] [CrossRef]
- Xie, D.; Tian, Y.; Hu, D.; Wang, Y.; Yang, Y.; Zhou, B.; Zhang, R.; Ren, Z.; Liu, M.; Xu, J.; et al. Oncolytic adenoviruses expressing checkpoint inhibitors for cancer therapy. Signal Transduct. Target. Ther. 2023, 8, 436. [Google Scholar] [CrossRef]
- Quixabeira, D.C.A.; Zafar, S.; Santos, J.M.; Cervera-Carrascon, V.; Havunen, R.; Kudling, T.V.; Basnet, S.; Anttila, M.; Kanerva, A.; Hemminki, A. Oncolytic Adenovirus Coding for a Variant Interleukin 2 (vIL-2) Cytokine Re-Programs the Tumor Microenvironment and Confers Enhanced Tumor Control. Front. Immunol. 2021, 12, 674400. [Google Scholar] [CrossRef]
- Yi, L.; Ning, Z.; Xu, L.; Shen, Y.; Zhu, X.; Yu, W.; Xie, J.; Meng, Z. The combination treatment of oncolytic adenovirus H101 with nivolumab for refractory advanced hepatocellular carcinoma: An open-label, single-arm, pilot study. ESMO Open 2024, 9, 102239. [Google Scholar] [CrossRef]
- Atasheva, S.; Shayakhmetov, D.M. Oncolytic Viruses for Systemic Administration: Engineering a Whole Different Animal. Mol. Ther. 2021, 29, 904–907. [Google Scholar] [CrossRef]
- Andtbacka, R.H.; Ross, M.; Puzanov, I.; Milhem, M.; Collichio, F.; Delman, K.A.; Amatruda, T.; Zager, J.S.; Cranmer, L.; Hsueh, E.; et al. Patterns of Clinical Response with Talimogene Laherparepvec (T-VEC) in Patients with Melanoma Treated in the OPTiM Phase III Clinical Trial. Ann. Surg. Oncol. 2016, 23, 4169–4177. [Google Scholar] [CrossRef] [PubMed]
- Gómez, A.; Sardón, D.; Cejalvo, T.; Vázquez, F.; García-Castro, J.; Perisé-Barrios, A.J. Biodistribution Analysis of Oncolytic Adenoviruses in Canine Patient Necropsy Samples Treated with Cellular Virotherapy. Mol. Ther. Oncolytics 2020, 18, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.; Zhang, W.; Li, Y.; Yu, L. Harnessing adenovirus in cancer immunotherapy: Evoking cellular immunity and targeting delivery in cell-specific manner. Biomark. Res. 2024, 12, 36. [Google Scholar] [CrossRef]
- Khare, R.; Hillestad, M.L.; Xu, Z.; Byrnes, A.P.; Barry, M.A. Circulating antibodies and macrophages as modulators of adenovirus pharmacology. J. Virol. 2013, 87, 3678–3686. [Google Scholar] [CrossRef]
- Hemminki, O.; Dos Santos, J.M.; Hemminki, A. Oncolytic viruses for cancer immunotherapy. J. Hematol. Oncol. 2020, 13, 84. [Google Scholar] [CrossRef]
- Jirovec, E.; Quixabeira, D.C.A.; Clubb, J.H.A.; Pakola, S.A.; Kudling, T.; Arias, V.; Haybout, L.; Jalkanen, K.; Alanko, T.; Monberg, T.; et al. Single intravenous administration of oncolytic adenovirus TILT-123 results in systemic tumor transduction and immune response in patients with advanced solid tumors. J. Exp. Clin. Cancer Res. 2024, 43, 297. [Google Scholar] [CrossRef]
- Heo, J.; Liang, J.D.; Kim, C.W.; Woo, H.Y.; Shih, I.L.; Su, T.H.; Lin, Z.Z.; Yoo, S.Y.; Chang, S.; Urata, Y.; et al. Safety and dose escalation of the targeted oncolytic adenovirus OBP-301 for refractory advanced liver cancer: Phase I clinical trial. Mol. Ther. 2023, 31, 2077–2088. [Google Scholar] [CrossRef]
- Garcia-Carbonero, R.; Bazan-Peregrino, M.; Gil-Martín, M.; Álvarez, R.; Macarulla, T.; Riesco-Martinez, M.C.; Verdaguer, H.; Guillén-Ponce, C.; Farrera-Sal, M.; Moreno, R.; et al. Phase I, multicenter, open-label study of intravenous VCN-01 oncolytic adenovirus with or without nab-paclitaxel plus gemcitabine in patients with advanced solid tumors. J. Immunother. Cancer 2022, 10, e003255. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.; Heiniö, C.; Quixabeira, D.; Zafar, S.; Clubb, J.; Pakola, S.; Cervera-Carrascon, V.; Havunen, R.; Kanerva, A.; Hemminki, A. Systemic Delivery of Oncolytic Adenovirus to Tumors Using Tumor-Infiltrating Lymphocytes as Carriers. Cells 2021, 10, 978. [Google Scholar] [CrossRef]
- Thambi, T.; Hong, J.; Yoon, A.R.; Yun, C.-O. Challenges and progress toward tumor-targeted therapy by systemic delivery of polymer-complexed oncolytic adenoviruses. Cancer Gene Ther. 2022, 29, 1321–1331. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, Z.; Li, L.; Wu, J.; Zhang, H.; Lei, T.; Xu, B. Oncolytic Adenovirus: Prospects for Cancer Immunotherapy. Front. Microbiol. 2021, 12, 707290. [Google Scholar] [CrossRef] [PubMed]
- Mendez, N.; Herrera, V.; Zhang, L.; Hedjran, F.; Feuer, R.; Blair, S.L.; Trogler, W.C.; Reid, T.R.; Kummel, A.C. Encapsulation of adenovirus serotype 5 in anionic lecithin liposomes using a bead-based immunoprecipitation technique enhances transfection efficiency. Biomaterials 2014, 35, 9554–9561. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-H.; Dong, T.; Phung, A.T.; Shah, J.R.; Larson, C.; Sanchez, A.B.; Blair, S.L.; Oronsky, B.; Trogler, W.C.; Reid, T.; et al. Full Remission of CAR-Deficient Tumors by DOTAP-Folate Liposome Encapsulation of Adenovirus. ACS Biomater. Sci. Eng. 2022, 8, 5199–5209. [Google Scholar] [CrossRef]
- Shah, J.R.; Dong, T.; Phung, A.T.; Reid, T.; Larson, C.; Sanchez, A.B.; Oronsky, B.; Blair, S.L.; Aisagbonhi, O.; Trogler, W.C.; et al. Development of Adenovirus Containing Liposomes Produced by Extrusion vs. Homogenization: A Comparison for Scale-Up Purposes. Bioengineering 2022, 9, 620. [Google Scholar] [CrossRef]
- Dong, T.; Shah, J.R.; Phung, A.T.; Larson, C.; Sanchez, A.B.; Aisagbonhi, O.; Blair, S.L.; Oronsky, B.; Trogler, W.C.; Reid, T.; et al. A Local and Abscopal Effect Observed with Liposomal Encapsulation of Intratumorally Injected Oncolytic Adenoviral Therapy. Cancers 2023, 15, 3157. [Google Scholar] [CrossRef]
- Phung, A.T.; Shah, J.R.; Dong, T.; Reid, T.; Larson, C.; Sanchez, A.B.; Oronsky, B.; Trogler, W.C.; Kummel, A.C.; Aisagbonhi, O.; et al. CAR expression in invasive breast carcinoma and its effect on adenovirus transduction efficiency. Breast Cancer Res. 2024, 26, 131. [Google Scholar] [CrossRef]
- Yotnda, P.; Chen, D.H.; Chiu, W.; Piedra, P.A.; Davis, A.; Templeton, N.S.; Brenner, M.K. Bilamellar cationic liposomes protect adenovectors from preexisting humoral immune responses. Mol. Ther. 2002, 5, 233–241. [Google Scholar] [CrossRef]
- Steel, J.C.; Cavanagh, H.M.; Burton, M.A.; Dingwall, D.J.; Kalle, W.H. Modification of liposomal concentration in liposome/adenoviral complexes allows significant protection of adenoviral vectors from neutralising antibody, in vitro. J. Virol. Methods 2005, 126, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-H.; Smyth-Templeton, N.; Davis, A.R.; Davis, E.A.; Ballian, N.; Li, M.; Liu, H.; Fisher, W.; Brunicardi, F.C. Multiple treatment cycles of liposome-encapsulated adenoviral RIP-TK gene therapy effectively ablate human pancreatic cancer cells in SCID mice. Surgery 2011, 149, 484–495. [Google Scholar] [CrossRef]
- Ganesan, L.P.; Mohanty, S.; Kim, J.; Clark, K.R.; Robinson, J.M.; Anderson, C.L. Rapid and efficient clearance of blood-borne virus by liver sinusoidal endothelium. PLoS Pathog. 2011, 7, e1002281. [Google Scholar] [CrossRef]
- Welsher, K.; Sherlock, S.P.; Dai, H. Deep-tissue anatomical imaging of mice using carbon nanotube fluorophores in the second near-infrared window. Proc. Natl. Acad. Sci. USA 2011, 108, 8943–8948. [Google Scholar] [CrossRef] [PubMed]
- Naumenko, V.A.; Vishnevskiy, D.A.; Stepanenko, A.A.; Sosnovtseva, A.O.; Chernysheva, A.A.; Abakumova, T.O.; Valikhov, M.P.; Lipatova, A.V.; Abakumov, M.A.; Chekhonin, V.P. In Vivo Tracking for Oncolytic Adenovirus Interactions with Liver Cells. Biomedicines 2022, 10, 1697. [Google Scholar] [CrossRef] [PubMed]
- Shayakhmetov, D.M.; Li, Z.Y.; Ni, S.; Lieber, A. Analysis of adenovirus sequestration in the liver, transduction of hepatic cells, and innate toxicity after injection of fiber-modified vectors. J. Virol. 2004, 78, 5368–5381. [Google Scholar] [CrossRef]
- Tao, N.; Gao, G.P.; Parr, M.; Johnston, J.; Baradet, T.; Wilson, J.M.; Barsoum, J.; Fawell, S.E. Sequestration of adenoviral vector by Kupffer cells leads to a nonlinear dose response of transduction in liver. Mol. Ther. 2001, 3, 28–35. [Google Scholar] [CrossRef]
- Yoon, A.R.; Hong, J.; Kim, M.; Yun, C.-O. Hepatocellular carcinoma-targeting oncolytic adenovirus overcomes hypoxic tumor microenvironment and effectively disperses through both central and peripheral tumor regions. Sci. Rep. 2018, 8, 2233. [Google Scholar] [CrossRef]
- Zhang, Z.; Krimmel, J.; Hu, Z.; Seth, P. Systemic delivery of a novel liver-detargeted oncolytic adenovirus causes reduced liver toxicity but maintains the antitumor response in a breast cancer bone metastasis model. Hum. Gene Ther. 2011, 22, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Le, T.M.D.; Hong, J.; Jiao, A.; Yoon, A.R.; Yun, C.-O. Smart Accumulating Dual-Targeting Lipid Envelopes Equipping Oncolytic Adenovirus for Enhancing Cancer Gene Therapeutic Efficacy. ACS Nano 2024, 18, 27869–27890. [Google Scholar] [CrossRef]
- Shah, J.R.; Dong, T.; Phung, A.T.; Khan, S.; Aisagbonhi, O.; Blair, S.L.; Bouvet, M.; Trogler, W.C.; Kummel, A.C. Liposomal oncolytic adenovirus as a neoadjuvant therapy for triple-negative breast cancer. Sci. Rep. 2025, 15, 16737. [Google Scholar] [CrossRef]
- Azumah, J.; Smistad, G.; Hiorth, M. Preparation of stable polymer-liposome complexes by a novel approach employing a one-pot method. Colloids Surf. A Physicochem. Eng. Asp. 2022, 653, 129924. [Google Scholar] [CrossRef]
- Suk, J.S.; Xu, Q.; Kim, N.; Hanes, J.; Ensign, L.M. PEGylation as a strategy for improving nanoparticle-based drug and gene delivery. Adv. Drug Deliv. Rev. 2016, 99, 28–51. [Google Scholar] [CrossRef]
- Santhanakrishnan, K.R.; Koilpillai, J.; Narayanasamy, D. PEGylation in Pharmaceutical Development: Current Status and Emerging Trends in Macromolecular and Immunotherapeutic Drugs. Cureus 2024, 16, e66669. [Google Scholar] [CrossRef] [PubMed]
- Nag, O.K.; Awasthi, V. Surface engineering of liposomes for stealth behavior. Pharmaceutics 2013, 5, 542–569. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, M.; Broniatowski, M.; Mach, M.; Płachta, Ł.; Wydro, P. The effect of the polyethylene glycol chain length of a lipopolymer (DSPE-PEGn) on the properties of DPPC monolayers and bilayers. J. Mol. Liq. 2021, 335, 116529. [Google Scholar] [CrossRef]
- El-Baz, N.; Nunn, B.M.; Bates, P.J.; O’Toole, M.G. The Impact of PEGylation on Cellular Uptake and In Vivo Biodistribution of Gold Nanoparticle MRI Contrast Agents. Bioengineering 2022, 9, 766. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Barz, M. Investigating the stability of RNA-lipid nanoparticles in biological fluids: Unveiling its crucial role for understanding LNP performance. J. Control. Release 2025, 381, 113559. [Google Scholar] [CrossRef]
- Marín, D.; Alemán, A.; Sánchez-Faure, A.; Montero, P.; Gómez-Guillén, M.C. Freeze-dried phosphatidylcholine liposomes encapsulating various antioxidant extracts from natural waste as functional ingredients in surimi gels. Food Chem. 2018, 245, 525–535. [Google Scholar] [CrossRef]
- Xia, Y.; Sun, J.; Liang, D. Aggregation, Fusion, and Leakage of Liposomes Induced by Peptides. Langmuir 2014, 30, 7334–7342. [Google Scholar] [CrossRef]
- Xu, X.; Tian, F.; Pan, Y.; Zhang, T.; Deng, L.; Jiang, H.; Han, J.; Liu, J.; Zhao, Y.; Liu, W. Emerging mechanistic insights into liposomal stability: Full process management from production and storage to food application. Chem. Eng. J. 2025, 505, 159552. [Google Scholar] [CrossRef]
- Yu, J.Y.; Chuesiang, P.; Shin, G.H.; Park, H.J. Post-Processing Techniques for the Improvement of Liposome Stability. Pharmaceutics 2021, 13, 1023. [Google Scholar] [CrossRef]



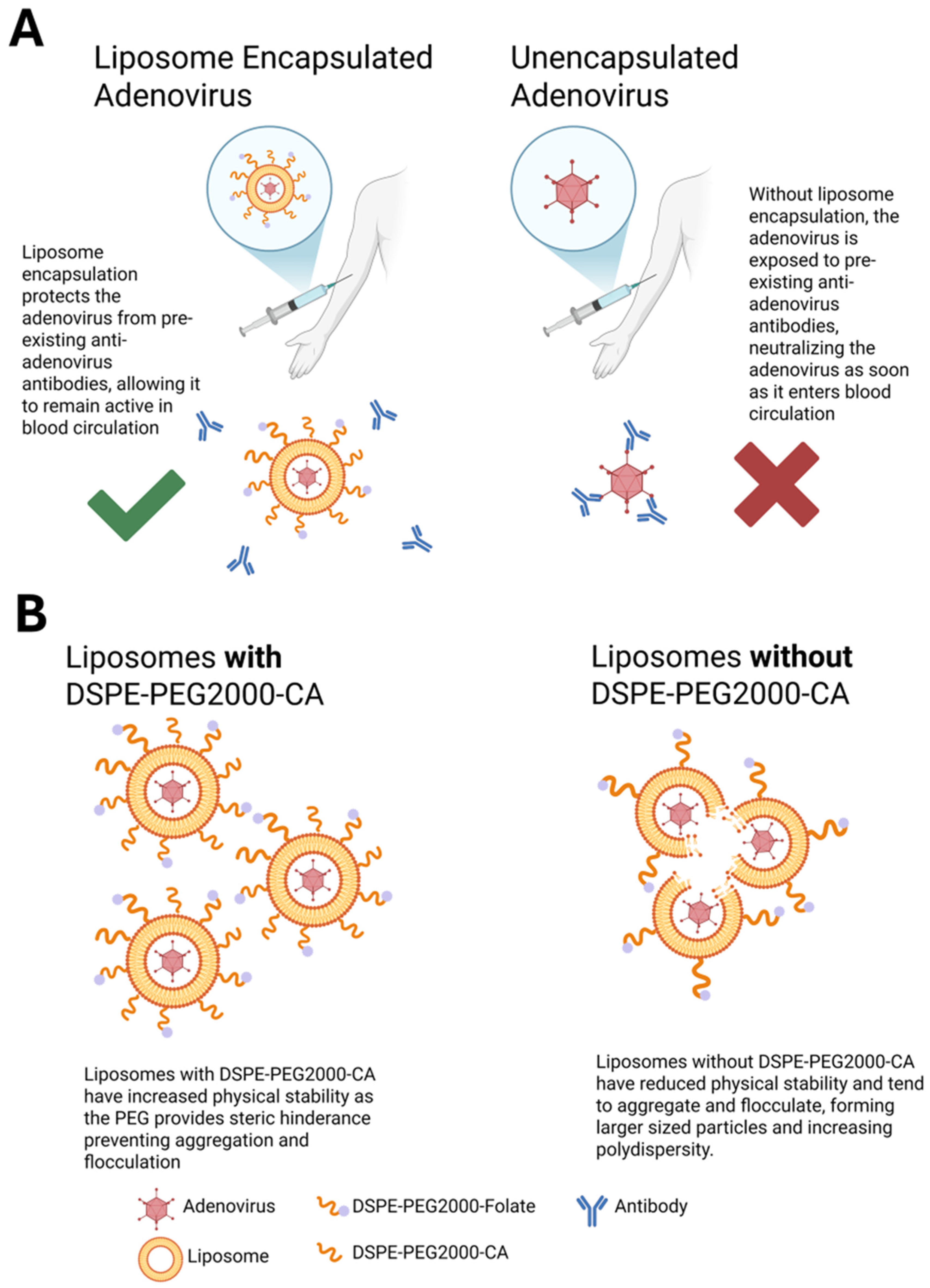

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | z-Average (nm) | Fold Change (z-Average) | Polydispersity Index (PDI) | Fold Change (PI) |
|---|---|---|---|---|
| DfAd DSPE-PEG2K-CA | 206 ± 2 | 1 | 0.1 ± 0.0 | 1 |
| Df DSPE-PEG2K-CA (Empty Liposomes) | 203 ± 2 | 1 | 0.1 ± 0.0 | 1 |
| DfAd No DSPE-PEG-CA | 3870 ± 57 | 19 | 0.7 ± 0.2 | 5 |
| Df No DSPE-PEG-CA (Empty Liposomes) | 3820 ± 175 | 19 | 0.4 ± 0.3 | 3 |
| DfAd DSPE-PEG1K-CA | 515 ± 27 | 3 | 0.6 ± 0.0 | 4 |
| Df DSPE-PEG1K-CA (Empty Liposomes) | 492 ± 12 | 2 | 0.6 ± 0.0 | 4 |
| DfAd DSPE-PEG10K-CA | 216 ± 1 | 1 | 0.2 ± 0.0 | 1 |
| Df DSPE-PEG10K-CA (Empty Liposomes) | 211 ± 2 | 1 | 0.2 ± 0.0 | 1 |
| Sample | Mean Diameter (nm) | Fold Change (Diameter) | D10, D50, D90 (nm) |
|---|---|---|---|
| DfAd DSPE-PEG2K-CA | 136 | 1.0 | 73, 120, 215 |
| DfAd No DSPE-PEG-CA | 291 | 2.1 | 127, 230. 560 |
| Sample | Mean Diameter (nm) | Fold Change (Diameter) | D10, D50, D90 (nm) |
|---|---|---|---|
| DfAd DSPE-PEG2K-CA | 145 | 1.0 | 82, 125, 234 |
| DfAd No DSPE-PEG-CA | 134 | 0.9 | 79, 121, 202 |
| DfAd DSPE-PEG1K-CA | 128 | 0.9 | 68, 105, 223 |
| DfAd DSPE-PEG5K-CA | 165 | 1.1 | 90, 148, 264 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phung, A.T.; Shah, J.R.; Dong, T.; Aisagbonhi, O.; Trogler, W.C.; Kummel, A.C.; Blair, S.L. Adenoviruses Encapsulated in PEGylated DOTAP-Folate Liposomes Are Protected from the Pre-Existing Humoral Immune Response. Pharmaceutics 2025, 17, 769. https://doi.org/10.3390/pharmaceutics17060769
Phung AT, Shah JR, Dong T, Aisagbonhi O, Trogler WC, Kummel AC, Blair SL. Adenoviruses Encapsulated in PEGylated DOTAP-Folate Liposomes Are Protected from the Pre-Existing Humoral Immune Response. Pharmaceutics. 2025; 17(6):769. https://doi.org/10.3390/pharmaceutics17060769
Chicago/Turabian StylePhung, Abraham T., Jaimin R. Shah, Tao Dong, Omonigho Aisagbonhi, William C. Trogler, Andrew C. Kummel, and Sarah L. Blair. 2025. "Adenoviruses Encapsulated in PEGylated DOTAP-Folate Liposomes Are Protected from the Pre-Existing Humoral Immune Response" Pharmaceutics 17, no. 6: 769. https://doi.org/10.3390/pharmaceutics17060769
APA StylePhung, A. T., Shah, J. R., Dong, T., Aisagbonhi, O., Trogler, W. C., Kummel, A. C., & Blair, S. L. (2025). Adenoviruses Encapsulated in PEGylated DOTAP-Folate Liposomes Are Protected from the Pre-Existing Humoral Immune Response. Pharmaceutics, 17(6), 769. https://doi.org/10.3390/pharmaceutics17060769