
C3-Liposome Delivery of MUC1 Peptide and TLR Agonists Enhances Adaptive Immunity and Results in Sex-Based Tumor Growth Differences

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Tumor Cell Line
2.3. Liposome Preparation
2.4. Mouse Model, Vaccination Groups, and Immunization
2.5. Flow Cytometry Analysis of Spleen Cells
2.6. ELISA Assay
2.7. ELISPOT Assay
2.8. Liver Toxicity Assays
2.9. Statistical Analysis
3. Results
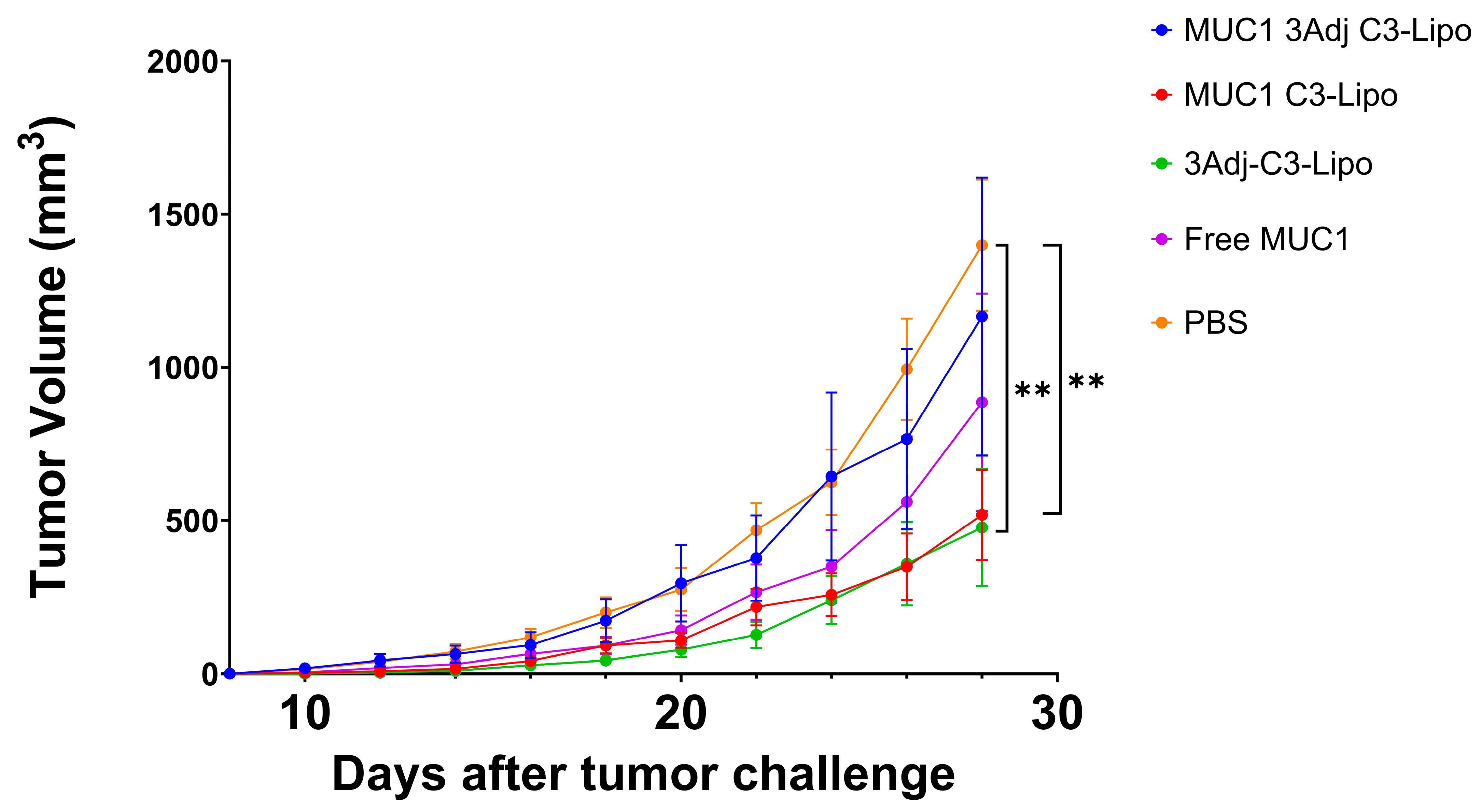
3.1. MUC1 C3-Liposomes Slow Tumor Growth in MUC1.Tg Mice
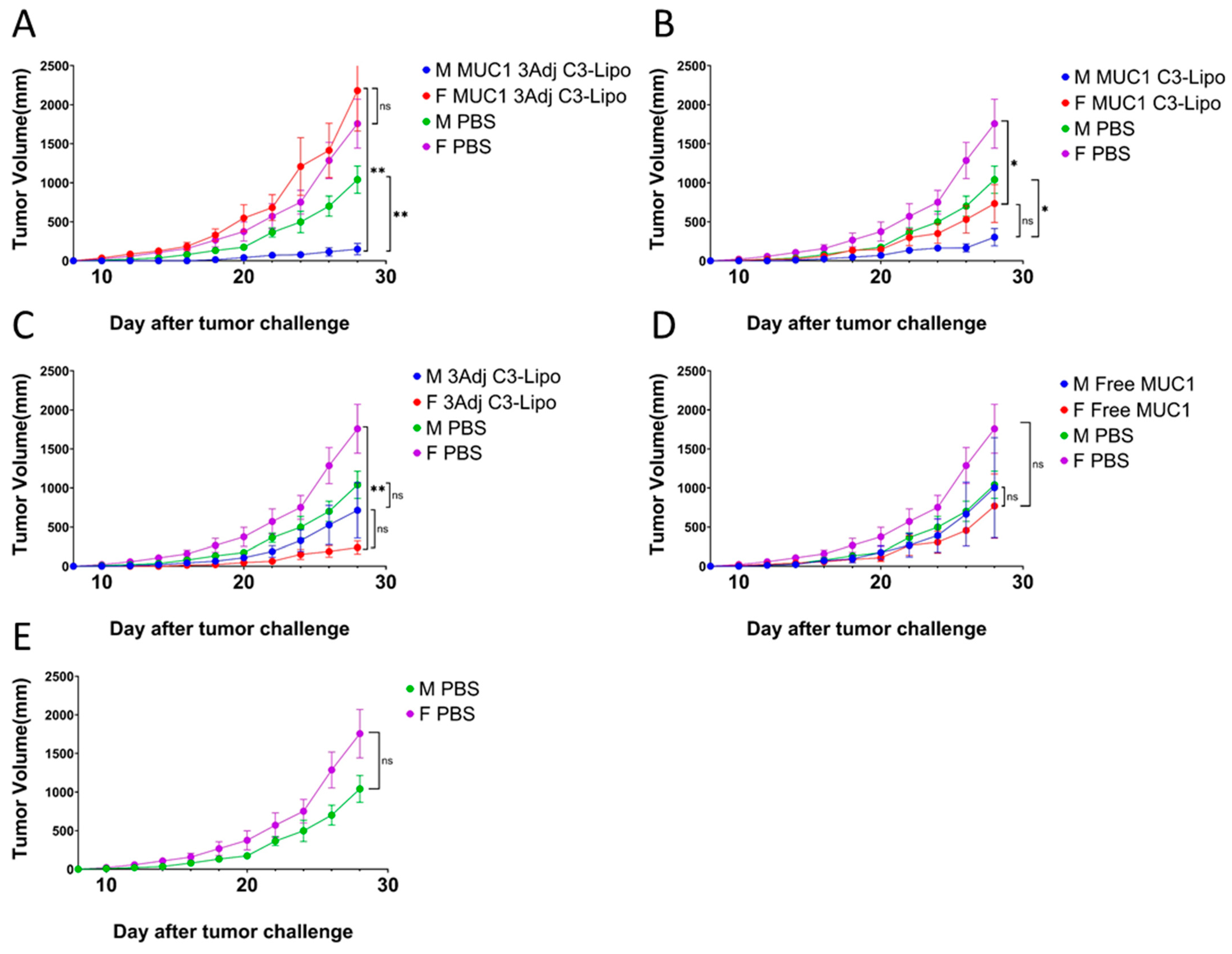
3.2. C3-Liposome Delivery of MUC1 Antigen and TLR Agonists Resulted in Sex Differences in Tumor Growth
3.3. MUC1-Specific T-Cell Response in Mice Vaccinated with MUC1 C3-Liposome Formulations
3.4. C3-Liposomes Enhance a MUC1-Specific IgG Antibody Response
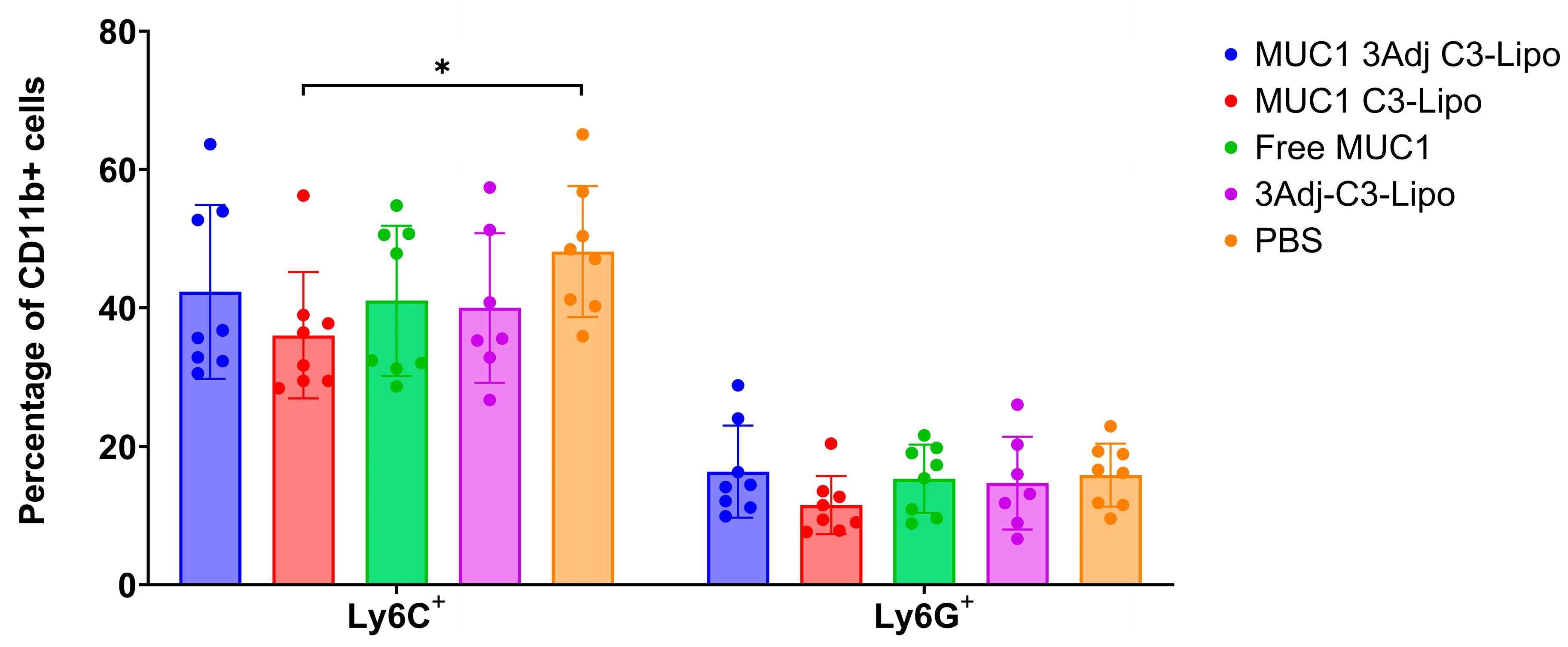
3.5. Vaccination with MUC1 3Adj C3-Liposomes Results in Decreased Levels of Systemic Monocytic MDSCs
3.6. Sex Differences in MDSC Levels in Mice Vaccinated with MUC1 3Adj C3-Liposomes
3.7. No Cytotoxicity Was Observed in Vaccinated MUC1.Tg Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Finn, O.J. Cancer vaccines: Between the idea and the reality. Nat. Rev. Immunol. 2003, 3, 630–641. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, R.E.; Kathrin, J. Turning the corner on therapeutic cancer vaccines. NPJ Vaccines 2019, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Barnd, D.L.; Lan, M.S.; Metzgar, R.S.; Finn, O.J. Specific, major histocompatibility complex-unrestricted recognition of tumor-associated mucins by human cytotoxic T cells. Proc. Natl. Acad. Sci. USA 1989, 86, 7159–7163. [Google Scholar] [CrossRef]
- Finn, O.J.; Gantt, K.R.; Lepisto, A.J.; Pejawar-Gaddy, S.; Xue, J.; Beatty, P.L. Importance of MUC1 and spontaneous mouse tumor models for understanding the immunobiology of human adenocarcinomas. Immunol. Res. 2011, 50, 261–268. [Google Scholar] [CrossRef]
- Gao, T.; Qianhong, C.; Han, L. A review on development of MUC1-based cancer vaccine. Biomed. Pharmacother. 2020, 132, 110888. [Google Scholar] [CrossRef] [PubMed]
- Acres, B.; Apostolopoulos, V.; Balloul, J.-M.; Wreschner, D.; Xing, P.-X.; Ali-Hadji, D.; Bizouarne, N.; Kieny, M.P.; McKenzie, I.F. MUC1-specific immune responses in human MUC1 transgenic mice immunized with various human MUC1 vaccines. Cancer Immunol. Immunother. 2000, 48, 588–594. [Google Scholar] [CrossRef]
- Beatty, P.L.; Narayanan, S.; Gariépy, J.; Ranganathan, S.; Finn, O.J. Vaccine against MUC1 antigen expressed in inflammatory bowel disease and cancer lessens colonic inflammation and prevents progression to colitis-associated colon cancer. Cancer Prev. Res. 2010, 3, 438–446. [Google Scholar] [CrossRef]
- Coffman, R.L.; Sher, A.; Seder, R.A. Vaccine adjuvants: Putting innate immunity to work. Immunity 2010, 33, 492–503. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, L.; Gao, N.; Diao, Y.; Zhong, J.; Deng, Y.; Wang, Z.; Jin, G.; Wang, X. Synthetic MUC1 breast cancer vaccine containing a Toll-like receptor 7 agonist exerts antitumor effects. Oncol. Lett. 2020, 20, 2369–2377. [Google Scholar] [CrossRef]
- Ramanathan, R.K.; Lee, K.M.; McKolanis, J.; Hitbold, E.; Schraut, W.; Moser, A.J.; Warnick, E.; Whiteside, T.; Osborne, J.; Kim, H.; et al. Phase I study of a MUC1 vaccine composed of different doses of MUC1 peptide with SB-AS2 adjuvant in resected and locally advanced pancreatic cancer. Cancer Immunol. Immunother. 2005, 54, 254–264. [Google Scholar] [CrossRef]
- Du, J.-J.; Zhou, S.-H.; Cheng, Z.-R.; Xu, W.-B.; Zhang, R.-Y.; Wang, L.-S.; Guo, J. MUC1-specific immune responses enhanced by coadministration of liposomal DDA/MPLA and lipoglycopeptide. Front. Chem. 2022, 10, 814880. [Google Scholar]
- Zhou, S.-H.; Li, Y.-T.; Zhang, R.-Y.; Liu, Y.-L.; You, Z.-W.; Bian, M.-M.; Wen, Y.; Wang, J.; Du, J.-J.; Guo, J. Alum adjuvant and built-in TLR7 agonist synergistically enhance anti-MUC1 immune responses for cancer vaccine. Front. Immunol. 2022, 13, 857779. [Google Scholar] [CrossRef]
- Du, J.-J.; Wang, C.-W.; Xu, W.-B.; Zhang, L.; Tang, Y.-K.; Zhou, S.-H.; Gao, X.-F.; Yang, G.-F.; Guo, J. Multifunctional protein conjugates with built-in adjuvant(Adjuvant-Protein-Antigen) as cancer vaccines boost potent immune responses. iScience 2020, 23, 100935. [Google Scholar]
- Paston, S.J.; Brentville, V.A.; Symonds, P.; Durrant, L.G. Cancer vaccines, adjuvants, and delivery systems. Front. Immunol. 2022, 12, 627932. [Google Scholar]
- Wen, R.; Banik, B.; Pathak, R.K.; Kumar, A.; Kolishetti, N.; Dhar, S. Nanotechnology-inspired tools for mitochondrial dysfunction-related diseases. Adv. Drug Deliv. Rev. 2016, 99, 52–69. [Google Scholar] [CrossRef] [PubMed]
- Francian, A.; Mann, K.; Kullberg, M. Complement C3-dependent uptake of targeted liposomes into human macrophages, B cells, dendritic cells, neutrophils, and MDSCs. Int. J. Nanomed. 2017, 12, 5149–5161. [Google Scholar]
- Francian, A.; Namen, S.; Stanley, M.; Mann, K.; Martinson, H.; Kullberg, M. Intratumoral delivery of antigen with complement C3-bound liposomes reduces tumor growth in mice. Nanomedicine 2019, 18, 326–335. [Google Scholar] [CrossRef]
- Joshi, M.D.; Unger, W.J.; Storm, G.; van Kooyk, Y.; Mastrobattista, E. Targeting tumor antigens to dendritic cells using particulate carriers. J. Control. Release 2012, 25, 37. [Google Scholar]
- Irache, J.M.; Salman, H.H.; Gamazo, C.; Espuelas, S. Mannose-targeted systems for the delivery of therapeutics. Expert Opin. Drug Deliv. 2008, 6, 703–724. [Google Scholar] [CrossRef]
- Fan, Y.; Moon, J.J. Nanoparticle drug delivery systems designed to improve cancer vaccines and immunotherapy. Vaccines 2015, 3, 662–685. [Google Scholar] [CrossRef]
- Arabi, A.; Soltani, S.A.; Maniaci, B.; Mann, K.; Martinson, H.; Kullberg, M. Enhancing T cell and antibody response in Mucin-1 transgenic mice through co-delivery of tumor-associated Mucin-1 antigen and TLR agonists in C3-liposomes. Pharmaceutics 2023, 15, 2774. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; McKolanis, J.R.; Dzubinski, L.A.; Islam, K.; Potter, D.M.; Salazar, A.M.; Schoen, R.E.; Finn, O.J. MUC1 vaccine for individuals with advanced adenoma of the colon: A cancer immunoprevention feasibility study. Cancer Prev. Res. 2013, 6, 18–26. [Google Scholar]
- Marigo, I.; Dolcetti, L.; Serafini, P.; Zanovello, P.; Bronte, V. Tumor-induced tolerance and immune suppression by myeloid-derived suppressor cells. Immunol. Rev. 2008, 222, 162–179. [Google Scholar]
- Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Bronte, V. Coordinated regulation of myeloid cells by tumors. Nat. Rev. Immunol. 2012, 4, 253–268. [Google Scholar]
- Pan, J.; Zeng, W.; Jia, J.; Shi, Y.; Wang, D.; Dong, J.; Fang, Z.; He, J.; Yang, X.; Zhang, R.; et al. A novel therapeutic tumor vaccine targeting MUC1 in combination with PD-L1 elicits specific anti-tumor immunity in mice. Vaccines 2022, 10, 1092. [Google Scholar] [CrossRef]
- Gilewski, T.A.; Ragupathi, G.; Dickler, M.; Powell, S.; Bhuta, S.; Panageas, K.; Koganty, R.R.; Chin-Eng, J.; Hudis, C.; Norton, L.; et al. Immunization of high-risk breast cancer patients with clustered sTn-KLH conjugate plus the immunologic adjuvant QS-22. Clin. Cancer Res. 2007, 10, 2977–2985. [Google Scholar]
- Akporiaye, E.T.; Bradleydunlop, D.; Gendler, S.J.; Mukherjee, P.; Madsen, C.S.; Hahn, T.; Besselsen, D.G.; Dial, S.M.; Cui, H.; Trevor, K. Characterization of the MUC1.Tg/MIN transgenic mouse as a model for studying antigen-specific immunotherapy of adenomas. Vaccine 2007, 28, 6965–6974. [Google Scholar]
- Zhu, H.; Wang, K.; Wang, Z.; Wang, D.; Yin, X.; Liu, Y.; Yu, F.; Zhao, W. An efficient and safe MUC1-dendritic cell-derived exosome conjugate vaccine elicits potent cellular and humoral immunity and tumor inhibition in vivo. Acta Biomater. 2022, 138, 491–504. [Google Scholar] [PubMed]
- Dougan, M.; Dranoff, G. Immune therapy for cancer. Annu. Rev. Immunol. 2009, 27, 83–117. [Google Scholar]
- Samuel, J.; Budzynski, W.A.; Reddish, M.A.; Ding, L.; Zimmermann, G.L.; Krantz, M.J.; Koganty, R.R.; Longenecker, B.M. Immunogenicity and antitumor activity of a liposomal MUC1 peptide-based vaccine. Int. J. Cancer 1998, 2, 295–302. [Google Scholar]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 10, 626–638. [Google Scholar]
- Durgeau, A.; Virk, Y.; Corgnac, S.; Mami-Chouaib, F. Recent advances in targeting CD8 T-cell immunity for more effective cancer immunotherapy. Front. Immunol. 2018, 9, 14. [Google Scholar]
- Appay, V.; Douek, D.C.; Price, D.A. CD8+ T cell efficacy in vaccination and disease. Nat. Med. 2008, 14, 623–628. [Google Scholar]
- Cook, I.F. Sexual dimorphism of humoral immunity with human vaccines. Vaccine 2008, 26, 3551–3555. [Google Scholar] [PubMed]
- Weinstein, Y.; Ran, S.; Segal, S. Sex-associated differences in the regulation of immune responses controlled by the MHC of the mouse. J. Immunol. 1984, 132, 656–661. [Google Scholar]
- Hamilton, G.; Plangger, A. The impact of NK cell-based therapeutics for the treatment of lung cancer for biologics: Targets and therapy. Biologics 2021, 7, 265–277. [Google Scholar]
- Itahashi, K.; Irie, T.; Nishikawa, H. Regulatory T cell development in the tumor microenvironment. Eur. J. Immunol. 2022, 52, 1216–1227. [Google Scholar]
- Tam, J.W.; Kullas, A.L.; Mena, P.J.; van der Velden, M. CD11b+ Ly6Chi Ly6G− immature myeloid cells recruited in response to Salmonella enterica serovar Typhimurium infection exhibit protective and immunosuppressive properties. Infect. Immun. 2014, 82, 2606–2614. [Google Scholar]
- Höchst, B.; Mikulec, J.; Baccega, T.; Metzger, C.; Welz, M.; Peusquens, J.; Tacke, F.; Knolle, P.; Kurts, C.; Diehl, L.; et al. Differential induction of Ly6G and Ly6C positive myeloid-derived suppressor cells in chronic kidney and liver inflammation and fibrosis. PLoS ONE 2015, 10, e0119662. [Google Scholar]
- McGill, M.R. The past and present of serum aminotransferases and the future of liver injury biomarkers. EXCLI J. 2016, 15, 817–828. [Google Scholar]
- Ameneh, A. Development of a MUC1 Cancer Immunotherapy Using Complement Targeted Liposomes and Organoplatinum(IV) Complex as an Effective Anticancer Drug. Doctoral Dissertation, University of Alaska Fairbanks, Fairbanks, AK, USA, 2022. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccination Group | ALT * (U/L) | AST † (U/L) |
|---|---|---|
| MUC1 3Adj C3-liposomes | 37.25 ± 11.37 | 23.53 ± 5.10 |
| MC1 C3-liposomes | 56.06 ± 26.82 | 15.34 ± 5.21 |
| Free MUC1 | 26.05 ± 8.19 | 22.17 ± 4.28 |
| 3Adj C3-liposomes | 67.20 ± 31.23 | 20.08 ± 5.74 |
| PBS | 39.74 ± 12.90 | 10.12 ± 6.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soltani, S.; Arabi, A.; Mann, K.; Hess, A.; Martinson, H.A.; Kullberg, M. C3-Liposome Delivery of MUC1 Peptide and TLR Agonists Enhances Adaptive Immunity and Results in Sex-Based Tumor Growth Differences. Pharmaceutics 2025, 17, 468. https://doi.org/10.3390/pharmaceutics17040468
Soltani S, Arabi A, Mann K, Hess A, Martinson HA, Kullberg M. C3-Liposome Delivery of MUC1 Peptide and TLR Agonists Enhances Adaptive Immunity and Results in Sex-Based Tumor Growth Differences. Pharmaceutics. 2025; 17(4):468. https://doi.org/10.3390/pharmaceutics17040468
Chicago/Turabian StyleSoltani, Shahab, Ameneh Arabi, Kristine Mann, Austin Hess, Holly A. Martinson, and Max Kullberg. 2025. "C3-Liposome Delivery of MUC1 Peptide and TLR Agonists Enhances Adaptive Immunity and Results in Sex-Based Tumor Growth Differences" Pharmaceutics 17, no. 4: 468. https://doi.org/10.3390/pharmaceutics17040468
APA StyleSoltani, S., Arabi, A., Mann, K., Hess, A., Martinson, H. A., & Kullberg, M. (2025). C3-Liposome Delivery of MUC1 Peptide and TLR Agonists Enhances Adaptive Immunity and Results in Sex-Based Tumor Growth Differences. Pharmaceutics, 17(4), 468. https://doi.org/10.3390/pharmaceutics17040468