

Flavonoids as Potential Modulators of Pancreatic Lipase Catalytic Activity

 ,
, 

 , , ,
, , ,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. In Vitro Assay Using Porcine Pancreatic Lipase
2.3. In Vitro Assay Using Human Pancreatic Lipase
2.4. Inhibition Kinetic of Pancreatic Lipase
2.5. Statistical Analysis
2.6. Computational Studies
2.6.1. Protein Preparation and Binding Pocket Analysis
2.6.2. Ligand Preparation
2.6.3. Molecular Docking Protocol (Validation and Execution)
2.6.4. Visualization of Docking Poses and Protein–Ligand Interaction Analysis
3. Results
3.1. Porcine Pancreatic Lipase—In Vitro Inhibition
3.2. Human Pancreatic Lipase—In Vitro Inhibition
3.3. Porcine Pancreatic Lipase—Inhibitory Kinetics
3.4. Human Pancreatic Lipase—Inhibitory Kinetics
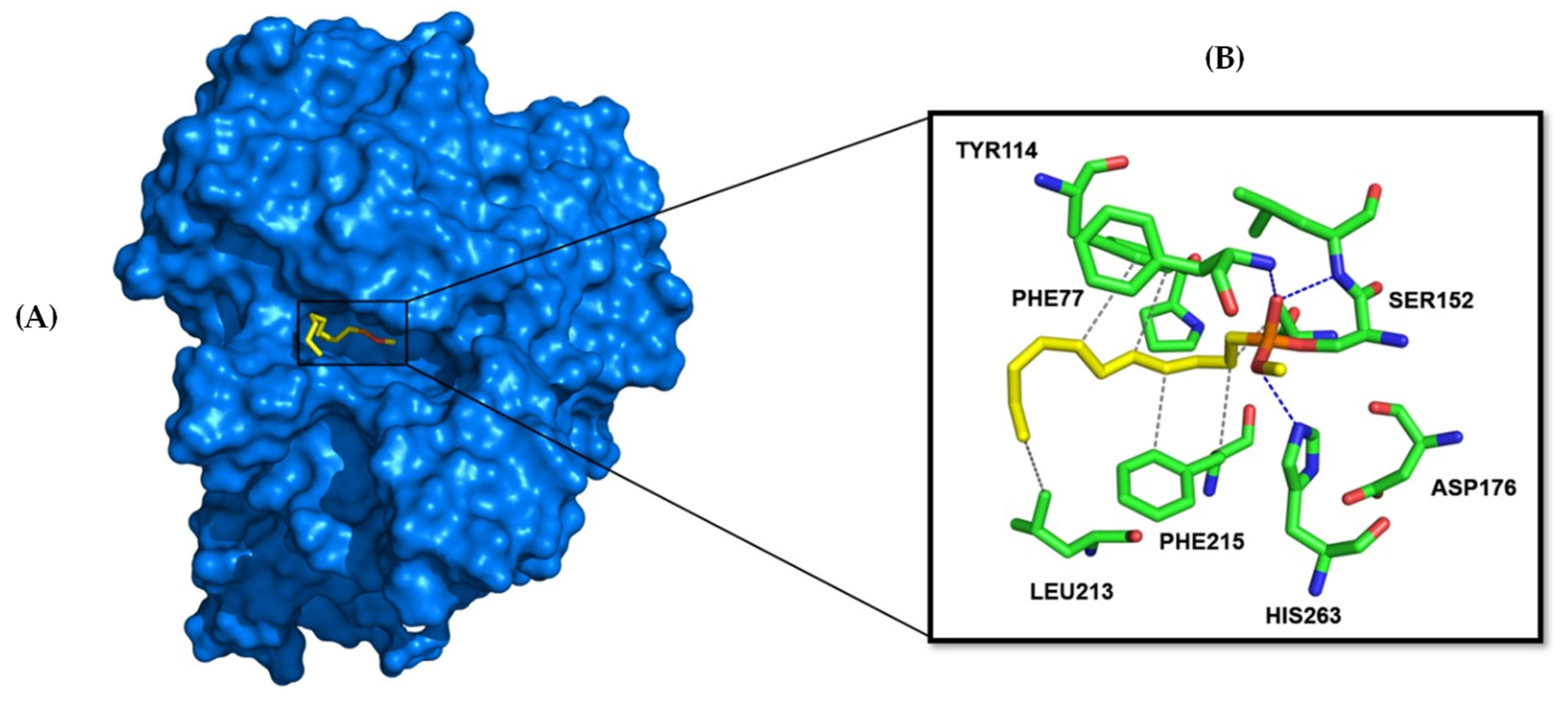
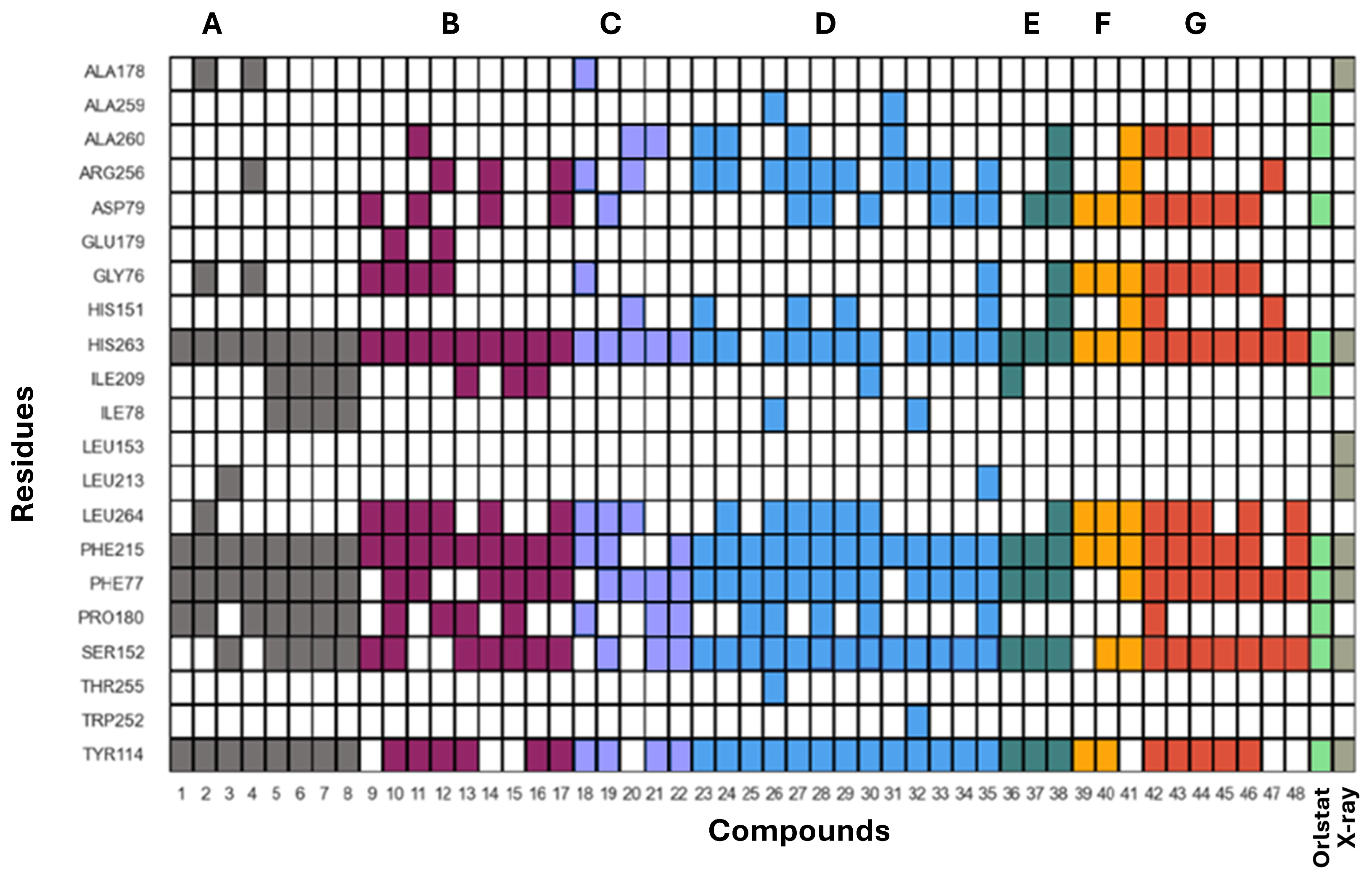
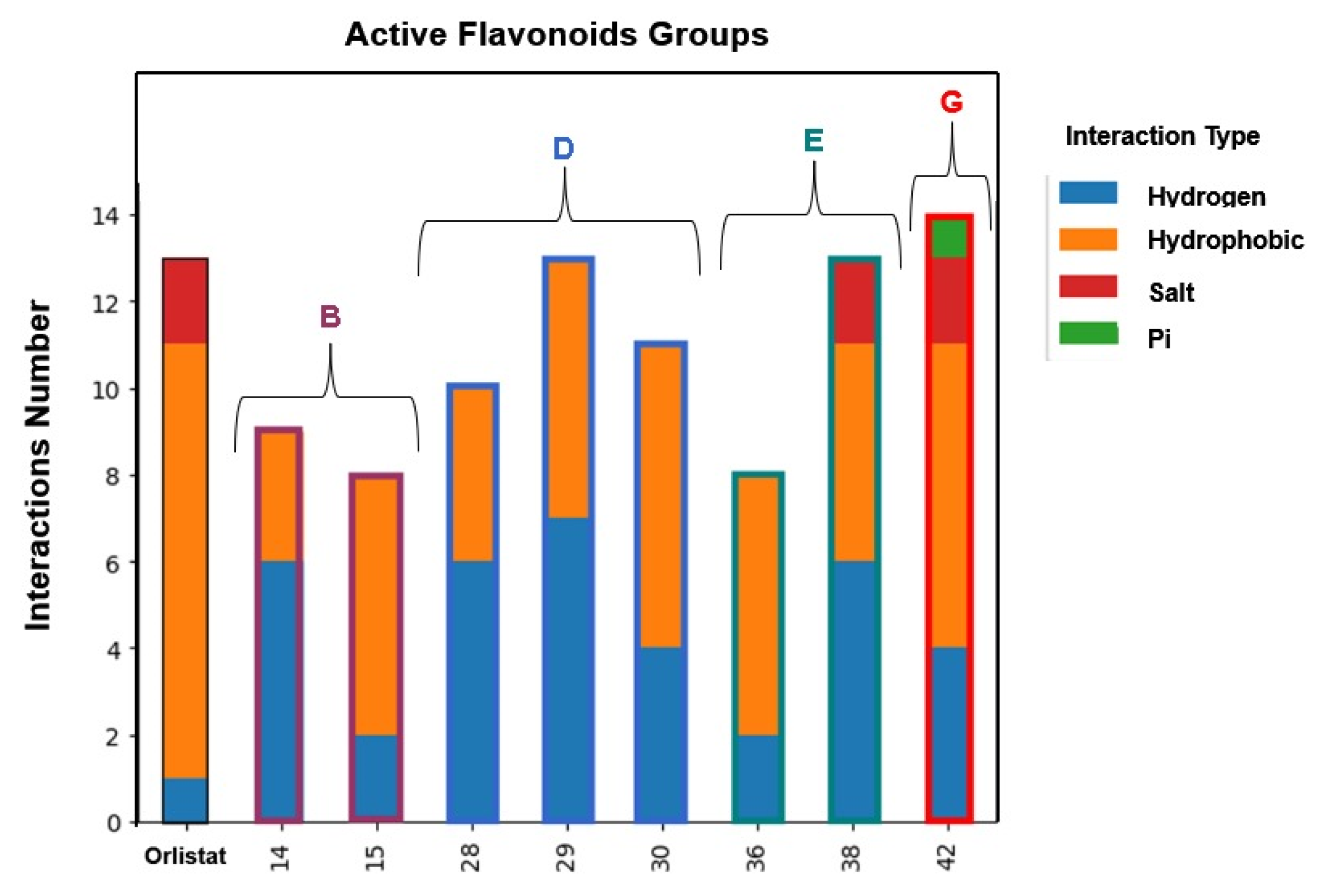
3.5. Human Pancreatic Lipase—Computational Studies
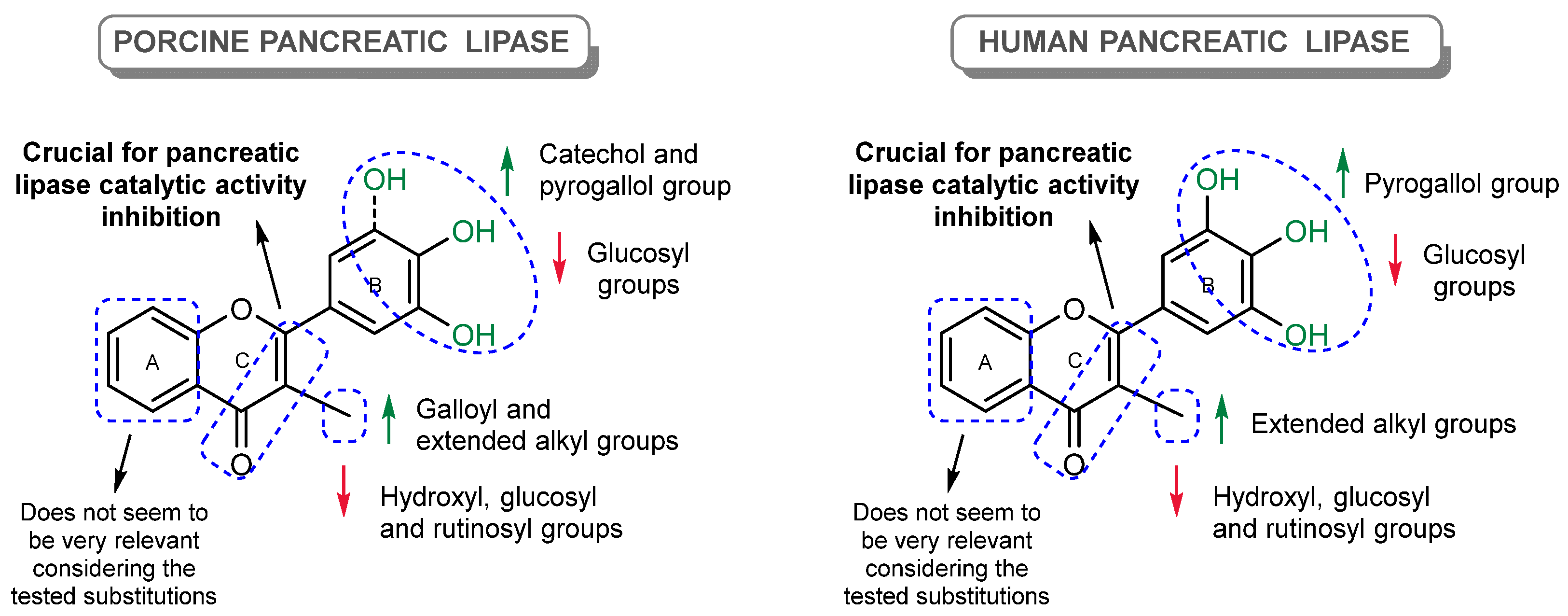
4. Discussion
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 4-MU | 4-methylumbeliferone |
| 4-MUO | 4-methylumbelliferyl oleate |
| AUC | Area under the curve |
| BMI | Body mass index |
| DMSO | Dimethyl sulfoxide |
| EGCG | (–)-epigallocatechin gallate |
| HPL | Human pancreatic lipase |
| PL | Pancreatic lipase |
| PPL | Porcine pancreatic lipase |
| RFU | Relative fluorescence unit |
| RMSD | Root mean square deviation |
| ROC | Receiver operating characteristic |
| Tris | Tris(hydroxymethyl)aminomethane |
| WHO | World Health Organization |
| WOF | World Obesity Federation |
References
- Jensen, M.D.; Ryan, D.H.; Apovian, C.M.; Ard, J.D.; Comuzzie, A.G.; Donato, K.A.; Hu, F.B.; Hubbard, V.S.; Jakicic, J.M.; Kushner, R.F. 2013 AHA/ACC/TOS guideline for the management of overweight and obesity in adults: A report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines and The Obesity Society. J. Am. Coll. Cardiol. 2014, 129, S102–S138. [Google Scholar] [CrossRef]
- Bray, G.A.; Kim, K.K.; Wilding, J.P.H.; World Obesity Federation. Obesity: A chronic relapsing progressive disease process. A position statement of the World Obesity Federation. Obes. Rev. 2017, 18, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Collaboration, P.S. Body-mass index and cause-specific mortality in 900 000 adults: Collaborative analyses of 57 prospective studies. Lancet 2009, 373, 1083–1096. [Google Scholar] [CrossRef] [PubMed]
- Apovian, C.M. Obesity: Definition, comorbidities, causes, and burden. AJMC 2016, 22, s176–s185. [Google Scholar]
- World Obesity Federation. World Obesity Atlas 2022; World Obesity Federation: London, UK, 2022; Available online: https://data.worldobesity.org/publications/World-Obesity-Atlas-2022-updated.pdf (accessed on 17 November 2024).
- World Obesity Federation. World Obesity Atlas 2023; World Obesity Federation: London, UK, 2023; Available online: https://www.worldobesity.org/resources/resource-library/world-obesity-atlas-2023 (accessed on 17 November 2024).
- Engin, A. The definition and prevalence of obesity and metabolic syndrome. In Obesity and Lipotoxicity; Springer: Cham, Switzerland, 2017; pp. 1–17. [Google Scholar]
- World Health Organization. Global Action Plan on Physical Activity 2018–2030: More Active People for a Healthier World; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Demirci, N.; Demirci, N.; Yildirim, İ.; Demirci, P.T.; Ersroz, Y. Why should we do physical activity? More active people for a healthier world. Int. J. Disabil. Sports Heal. Sci. 2018, 1, 1–14. [Google Scholar] [CrossRef]
- Clinical Guidelines on the Identification, Evaluation, and Treatment of Overweight and Obesity in Adults: The Evidence Report; National Heart, Lung, and Blood Institute: Bethesda, MD, USA, 1998.
- Wadden, T.A.; Bray, G.A. Handbook of Obesity Treatment; Guilford Publications: New York, NY, USA, 2018. [Google Scholar]
- Thomson, A.B.R.; De Pover, A.; Keelan, M.; Jarocka-Cyrta, E.; Clandinin, M.T. Inhibition of lipid absorption as an approach to the treatment of obesity. Meth. Enzymol. 1997, 286, 3–44. [Google Scholar]
- Lowe, M.E. Structure and function of pancreatic lipase and colipase. Annu. Rev. Nutr. 1997, 17, 141–158. [Google Scholar] [CrossRef] [PubMed]
- Lowe, M.E.; Rosenblum, J.L.; Strauss, A.W. Cloning and characterization of human pancreatic lipase cDNA. J. Biol. Chem. 1989, 264, 20042–20048. [Google Scholar] [CrossRef] [PubMed]
- Chapus, C.; Rovery, M.; Sarda, L.; Verger, R. Minireview on pancreatic lipase and colipase. Biochimie 1988, 70, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Gargouri, Y.; Bensalah, A.; Douchet, I.; Verger, R. Kinetic behaviour of pancreatic lipase in five species using emulsions and monomolecular films of synthetic glycerides. Biochim. Biophys. Acta. 1995, 1257, 223–229. [Google Scholar] [CrossRef]
- Abousalham, A.; Verger, R. Egg yolk lipoproteins as substrates for lipases. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2000, 1485, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Chanoine, J.-P.; Hampl, S.; Jensen, C.; Boldrin, M.; Hauptman, J. Effect of orlistat on weight and body composition in obese adolescents: A randomized controlled trial. JAMA 2005, 293, 2873–2883. [Google Scholar] [CrossRef]
- Tan, H.C.; Dampil, O.A.; Marquez, M.M. Efficacy and safety of semaglutide for weight loss in obesity without diabetes: A systematic review and meta-analysis. J. ASEAN Fed. Endocr. Soc. 2022, 37, 65. [Google Scholar] [CrossRef]
- Qi, X. Review of the clinical effect of orlistat. IOP Conf. Ser. Mater. Sci. Eng. 2018, 301, 012063. [Google Scholar] [CrossRef]
- Yao, L.H.; Jiang, Y.M.; Tomás-Barberán, F.A.; Datta, N.; Singanusong, R.; Chen, S.S. Flavonoids in food and their health benefits. Plant Foods Hum. Nutr. 2004, 59, 113–122. [Google Scholar] [CrossRef]
- Li, A.-N.; Li, S.; Zhang, Y.-J.; Xu, X.-R.; Chen, Y.-M.; Li, H.-B. Resources and biological activities of natural polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef] [PubMed]
- Hasnat, H.; Shompa, S.A.; Islam, M.M.; Alam, S.; Richi, F.T.; Emon, N.U.; Ashrafi, S.; Ahmed, N.U.; Chowdhury, N.R.; Fatema, N.; et al. Flavonoids: A treasure house of prospective pharmacological potentials. Heliyon 2024, 10, e27533. [Google Scholar] [CrossRef] [PubMed]
- Claudine, M.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2024, 79, 727–747. [Google Scholar]
- Tang, Z.; Zhang, Q. The potential toxic side effects of flavonoids. Biocell 2022, 46, 357–366. [Google Scholar] [CrossRef]
- Rocha, S.; Rufino, A.T.; Freitas, M.; Silva, A.M.S.; Carvalho, F.; Fernandes, E. Methodologies for assessing pancreatic lipase catalytic activity: A review. Crit. Rev. Anal. Chem. 2023, 54, 3038–3065. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, D.; Freitas, M.; Tomé, S.M.; Silva, A.M.S.; Porto, G.; Fernandes, E. Modulation of human neutrophils’ oxidative burst by flavonoids. Eur. J. Med. Chem. 2013, 67, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Seixas, R.S.G.R.; Pinto, D.C.G.A.; Silva, A.M.S.; Cavaleiro, J.A.S. Synthesis of Novel 3-Alkyl-3′, 4′, 5, 7-Tetrahydroxyflavones. Aust. J. Chem. 2008, 61, 718–724. [Google Scholar] [CrossRef]
- Sousa, J.L.C.; Proenca, C.; Freitas, M.; Fernandes, E.; Silva, A.M.S. New polyhydroxylated flavon-3-ols and 3-hydroxy-2-styrylchromones: Synthesis and ROS/RNS scavenging activities. Eur. J. Med. Chem. 2016, 119, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Deng, Z.; Ramdath, D.; Tang, Y.; Chen, P.X.; Liu, R.; Liu, Q.; Tsao, R. Phenolic profiles of 20 Canadian lentil cultivars and their contribution to antioxidant activity and inhibitory effects on α-glucosidase and pancreatic lipase. Food Chem. 2015, 172, 862–872. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, S.A.; Nomura, K.; Malfanov, I.L.; Sklyar, I.V.; Ptitsyn, L.R. Isolation of a novel catechin from Bergenia rhizomes that has pronounced lipase-inhibiting and antioxidative properties. Fitoterapia 2011, 82, 212–218. [Google Scholar] [CrossRef]
- Freitas, M.; Proença, C.; Ribeiro, D.; Quinaz-Garcia, M.B.; Araújo, A.N.; Fernandes, E. Assessment of α-amylase activity in a microanalysis system: Experimental optimization and evaluation of type of inhibition. J. Chem. Educ. 2023, 100, 1237–1245. [Google Scholar] [CrossRef]
- Marchand, L.R.; Pirard, B.; Ertl, P.; Sirockin, F. CAVIAR: A method for automatic cavity detection, description and decomposition into subcavities. J. Comput. Aided Mol. Des. 2021, 35, 737–750. [Google Scholar] [CrossRef]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New docking methods, expanded force field, and python bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra precision glide: Docking and scoring incorporating a model of hydrophobic enclosure for protein– ligand complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [PubMed]
- McNutt, A.T.; Francoeur, P.; Aggarwal, R.; Masuda, T.; Meli, R.; Ragoza, M.; Sunseri, J.; Koes, D.R. GNINA 1.0: Molecular docking with deep learning. J. Cheminf. 2021, 13, 43. [Google Scholar] [CrossRef] [PubMed]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully automated protein–ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef]
- Raschka, S. BioPandas: Working with molecular structures in pandas DataFrames. J. Open Source Softw. 2017, 2, 279. [Google Scholar] [CrossRef]
- Heck, A.M.; Yanovski, J.A.; Calis, K.A. Orlistat, a new lipase inhibitor for the management of obesity. Pharmacotherapy 2000, 20, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Bauer, E.; Jakob, S.; Mosenthin, R. Principles of physiology of lipid digestion. Asian-Australas. J. Anim. Sci. 2005, 18, 282–295. [Google Scholar] [CrossRef]
- Zdrazil, B.; Felix, E.; Hunter, F.; Manners, E.J.; Blackshaw, J.; Corbett, S.; Veij, M.; Ioannidis, H.; Lopez, D.M.; Mosquera, J.F.; et al. The ChEMBL Database in 2023: A drug discovery platform spanning multiple bioactivity data types and time periods. Nucleic Acids Res. 2024, 52, D1180–D1192. [Google Scholar] [CrossRef] [PubMed]
- Bray, G.A. Medical treatment of obesity: The past, the present and the future. Best Pract. Res. Clin. Obstet. Gynaecol. 2014, 28, 665–684. [Google Scholar] [CrossRef] [PubMed]
- Yanovski, S.Z.; Yanovski, J.A. Long-term drug treatment for obesity: A systematic and clinical review. JAMA 2014, 311, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-Y.; Kim, C.S.; Park, K.-M.; Chang, P.-S. Inhibitory characteristics of flavonol-3-O-glycosides from Polygonum aviculare L. (common knotgrass) against porcine pancreatic lipase. Sci. Rep. 2019, 9, 18080. [Google Scholar] [CrossRef] [PubMed]
- Nakai, M.; Fukui, Y.; Asami, S.; Toyoda-Ono, Y.; Iwashita, T.; Shibata, H.; Mitsunaga, T.; Hashimoto, F.; Kiso, Y. Inhibitory effects of oolong tea polyphenols on pancreatic lipase in vitro. J. Agric. Food Chem. 2005, 53, 4593–4598. [Google Scholar] [CrossRef] [PubMed]
- Yuda, N.; Tanaka, M.; Suzuki, M.; Asano, Y.; Ochi, H.; Iwatsuki, K. Polyphenols extracted from black tea (Camellia sinensis) residue by hot-compressed water and their inhibitory effect on pancreatic lipase in vitro. J. Food Sci. 2012, 77, H254–H261. [Google Scholar] [CrossRef] [PubMed]
- Cha, K.H.; Song, D.-G.; Kim, S.M.; Pan, C.-H. Inhibition of gastrointestinal lipolysis by green tea, coffee, and gomchui (Ligularia fischeri) tea polyphenols during simulated digestion. J. Agric. Food Chem. 2012, 60, 7152–7157. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, R.M.F.; Fraga, I.; Dias, A.A. Utilization of integrated Michaelis–Menten equations for enzyme inhibition diagnosis and determination of kinetic constants using Solver supplement of Microsoft Office Excel. Comput. Methods Programs Biomed. 2013, 109, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Gonzalez, A.I.; Alvarez-Parrilla, E.; Díaz-Sánchez, Á.G.; de la Rosa, L.A.; Núñez-Gastélum, J.A.; Vazquez-Flores, A.A.; Gonzalez-Aguilar, G.A. In vitro inhibition of pancreatic lipase by polyphenols: A kinetic, fluorescence spectroscopy and molecular docking study. Food Technol. Biotechnol. 2017, 55, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Rahim, A.T.M.A.; Takahashi, Y.; Yamaki, K. Mode of pancreatic lipase inhibition activity in vitro by some flavonoids and non-flavonoid polyphenols. Food Res. Int. 2015, 75, 289–294. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Porcine Pancreatic Lipase | Human Pancreatic Lipase | |||||||
|---|---|---|---|---|---|---|---|---|
| 4-MUO (μM) | 27 (Quercetin) (μM) | 30 (μM) | 38 (Myricetin) (μM) | 41 (EGCG) (μM) | 4-MUO (μM) | 30 (μM) | 38 (Myricetin) (μM) | |
| 12.5 | 0–50.0 | 0–15.0 | 0–30.0 | 0–60.0 | 12.5 | 0–10.0 | 0–50.0 | |
| 50 | 0–50.0 | 0–15.0 | 0–30.0 | 0–60.0 | 50 | 0–10.0 | 0–50.0 | |
| 200 | 0–50.0 | 0–15.0 | 0–30.0 | 0–60.0 | 200 | 0–10.0 | 0–50.0 | |
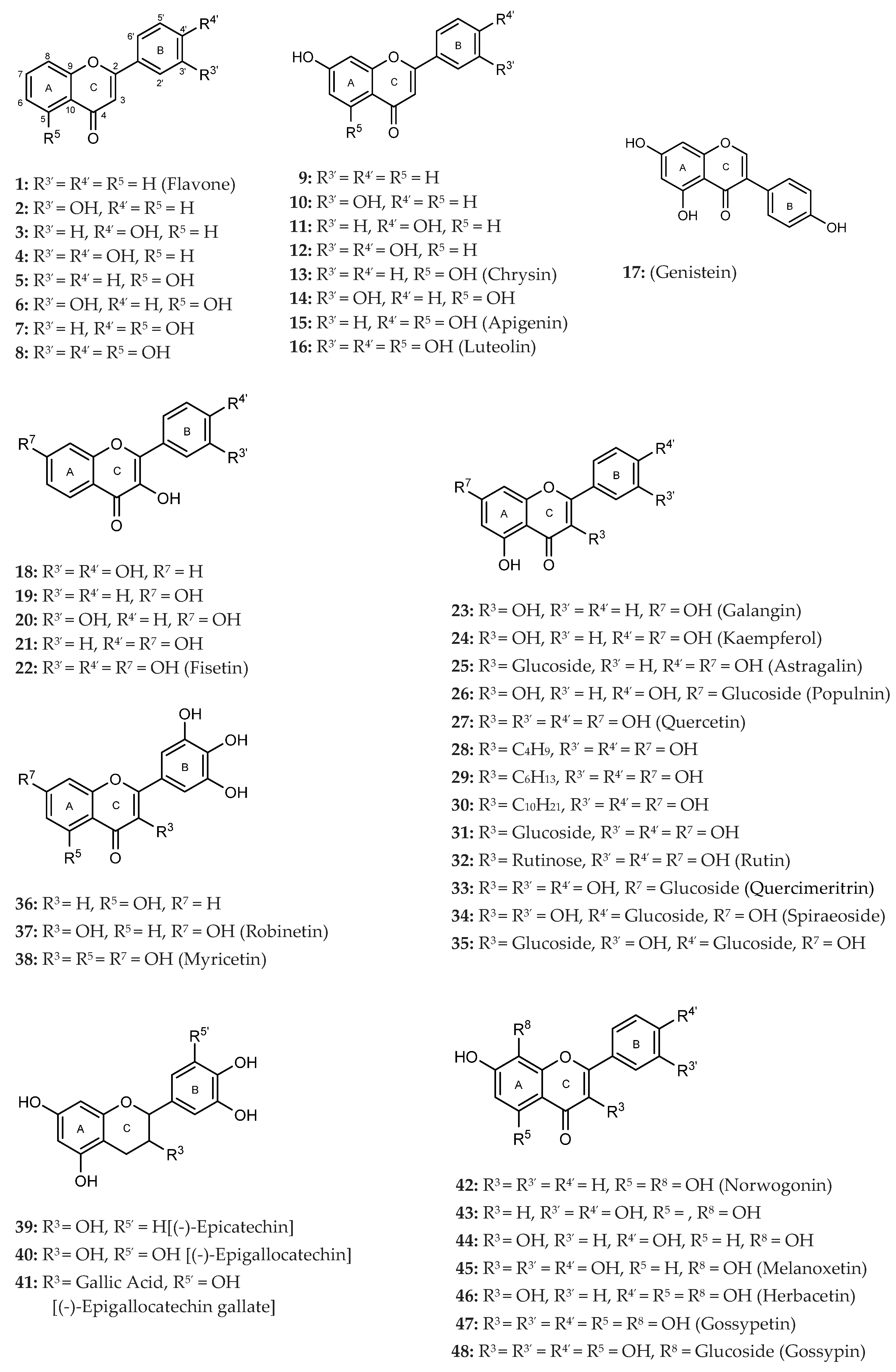
| Flavonoids | Ring C | Ring A | Ring B | IC50 (μM) ± SEM or PL Inhibition (%) ± SEM | IC50 (μM) ± SEM or PL Inhibition (%) ± SEM | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| R3 | R5 | R7 | R8 | R3′ | R4′ | R5′ | Porcine | Human | |||
| GROUP A (Flavones) | 1 Flavone | 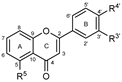 | --- | --- | --- | --- | --- | --- | --- | <30% 100 μM | <30% 100 μM |
| 2 | --- | --- | --- | --- | OH | --- | --- | <30% 100 μM | <30% 100 μM | ||
| 3 | --- | --- | --- | --- | --- | OH | --- | <30% 25 μM | <30% 25 μM | ||
| 4 | --- | --- | --- | --- | OH | OH | --- | <30% 100 μM | <30% 100 μM | ||
| 5 | --- | OH | --- | --- | --- | --- | --- | <30% 50 μM | <30% 50 μM | ||
| 6 | --- | OH | --- | --- | OH | --- | --- | <30% 25 μM | 32.3 ± 0.1% 25 μM | ||
| 7 | --- | OH | --- | --- | --- | OH | --- | <30% 25 μM | <30% 25 μM | ||
| 8 | --- | OH | --- | --- | OH | OH | --- | 46 ± 2% 50 μM | <30% 50 μM | ||
| GROUP B (Flavones) | 9 |  | --- | --- | OH | --- | --- | --- | --- | <30% 50 μM | <30% 50 μM |
| 10 | --- | --- | OH | --- | OH | --- | --- | <30% 100 μM | <30% 100 μM | ||
| 11 | --- | --- | OH | --- | --- | OH | --- | (a) | (a) | ||
| 12 | --- | --- | OH | --- | OH | OH | --- | (a) | (a) | ||
| 13 Chrysin | --- | OH | OH | --- | --- | --- | --- | <30% 50 μM | <30% 50 μM | ||
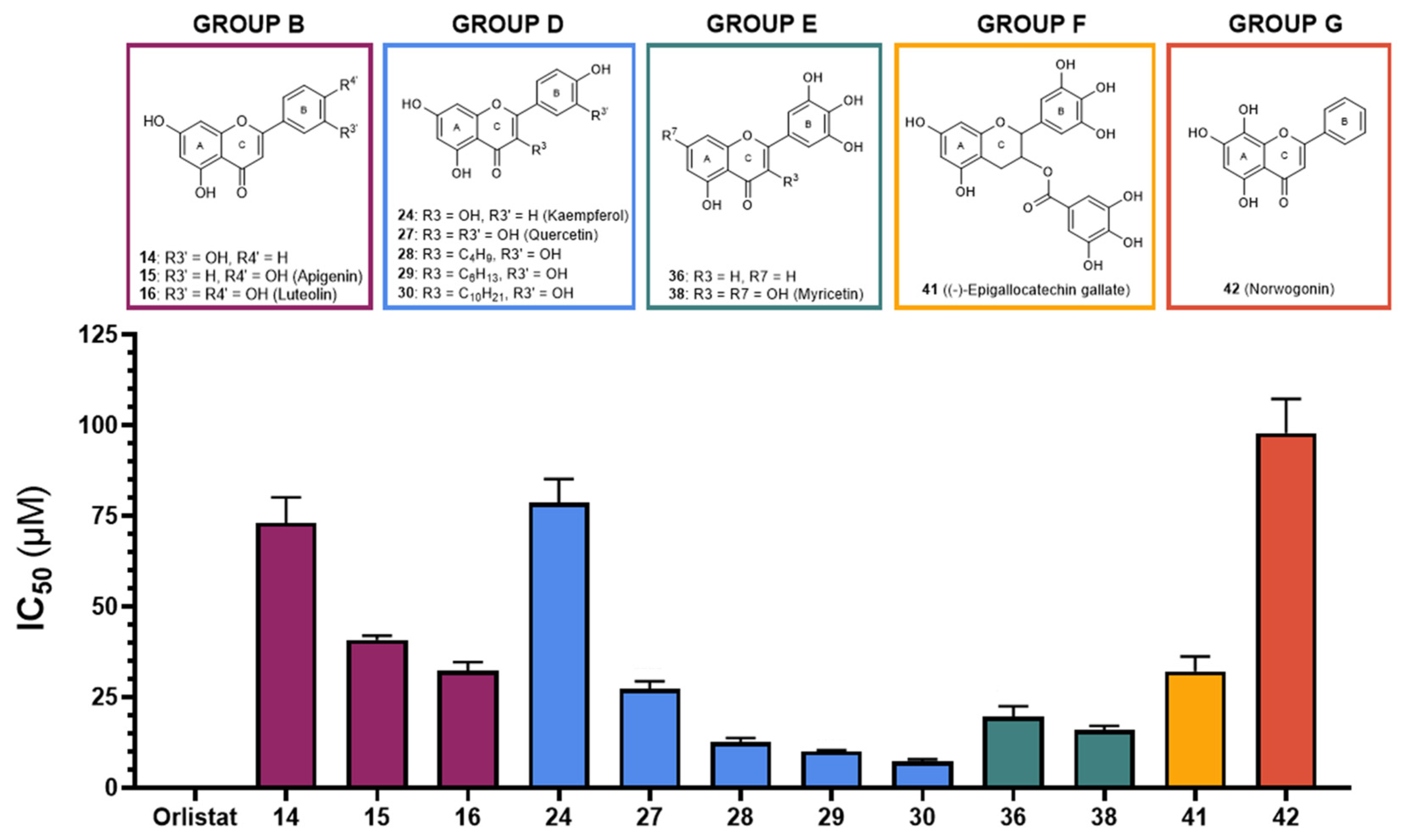
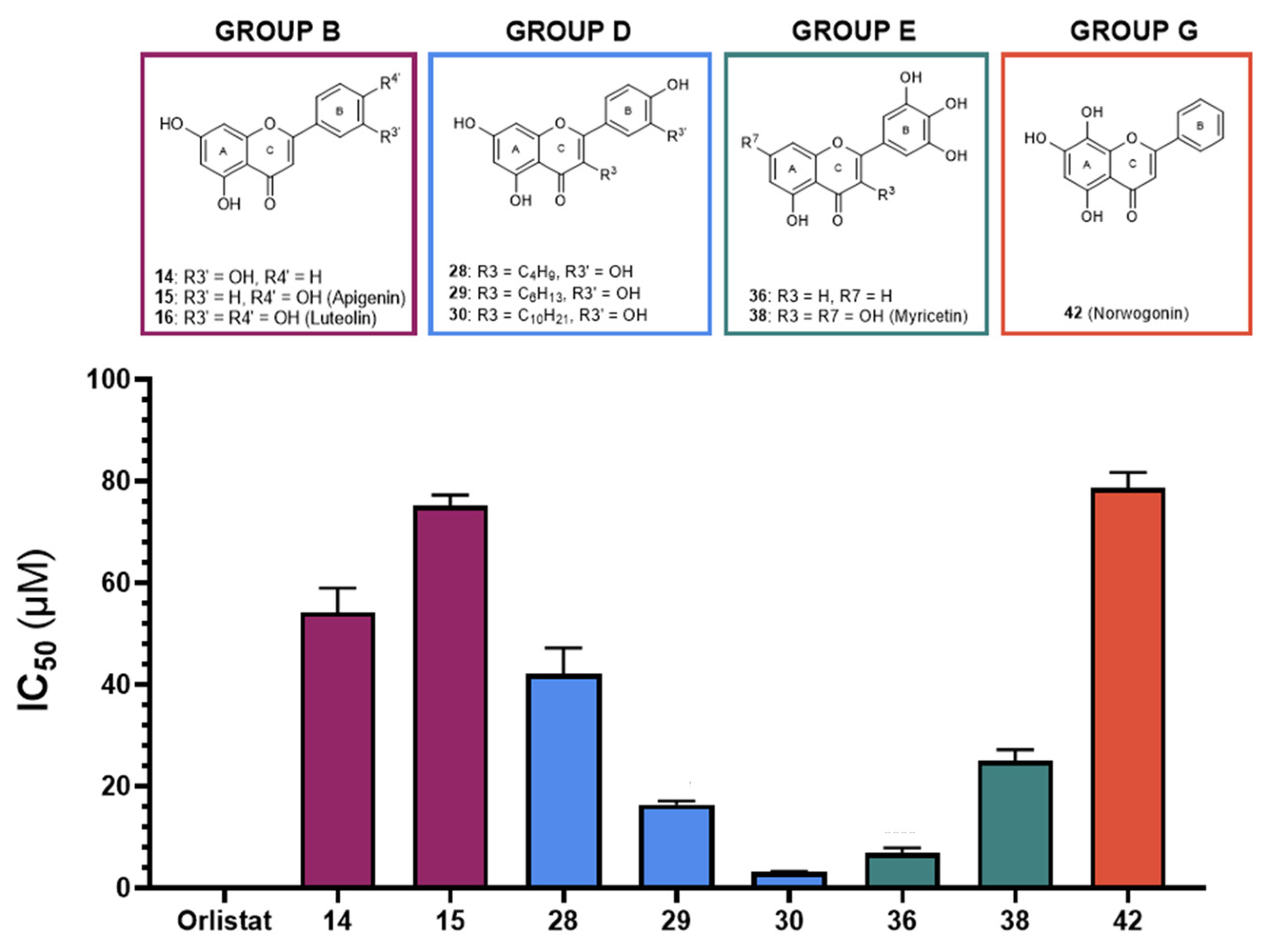
| 14 | --- | OH | OH | --- | OH | --- | --- | 73 ± 7 | 54 ± 5 | ||
| 15 Apigenin | --- | OH | OH | --- | --- | OH | --- | 41 ± 1 | 75 ± 2 | ||
| 16 Luteolin | --- | OH | OH | --- | OH | OH | --- | 32 ± 2 | 54 ± 2% 50 μM | ||
| Isoflavone 17 Genistein | 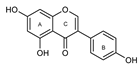 | --- | OH | OH | --- | --- | OH | --- | <30% 100 μM | <30% 100 μM | |
| GROUP C (Flavonols) | 18 |  | OH | --- | --- | --- | OH | OH | --- | 41 ± 4% 50 μM | 44 ± 5% 50 μM |
| 19 | OH | --- | OH | --- | --- | --- | --- | (a) | (a) | ||
| 20 | OH | --- | OH | --- | OH | --- | --- | (a) | (a) | ||
| 21 | OH | --- | OH | --- | --- | OH | --- | (a) | (a) | ||
| 22 Fisetin | OH | --- | OH | --- | OH | OH | --- | (a) | (a) | ||
| GROUP D (Flavones) | 23 Galangin |  | OH | OH | OH | --- | --- | --- | --- | 41 ± 1% 50 μM | <30% 50 μM |
| 24 Kaempferol | OH | OH | OH | --- | --- | OH | --- | 79 ± 6 | 39 ± 3%100 μM | ||
| 25 Astragalin | Glu | OH | OH | --- | --- | OH | --- | <30% 50 μM | 34 ± 1% 50 μM | ||
| 26 Populnin | OH | OH | Glu | --- | --- | OH | --- | <30% 100 μM | <30% 100 μM | ||
| 27 Quercetin | OH | OH | OH | --- | OH | OH | --- | 27 ± 2 | 46% ± 3% 75 μM | ||
| 28 | C4H9 | OH | OH | --- | OH | OH | --- | 13 ± 1 | 42 ± 2 | ||
| 29 | C6H13 | OH | OH | --- | OH | OH | --- | 10.1 ± 0.4 | 16.4 ± 0.7 | ||
| 30 | C10H21 | OH | OH | --- | OH | OH | --- | 7.3 ± 0.6 | 3.2 ± 0.1 | ||
| 31 | Glu | OH | OH | --- | OH | OH | --- | <30% 50 μM | <30% 50 μM | ||
| 32 Rutin | Rut | OH | OH | --- | OH | OH | --- | <30% 50 μM | 34 ± 1% 50 μM | ||
| 33 Quercimeritrin | OH | OH | Glu | --- | OH | OH | --- | <30% 50 μM | <30% 50 μM | ||
| 34 Spiraeoside | OH | OH | OH | --- | OH | Glu | --- | <30% 50 μM | <30% 50 μM | ||
| 35 | Glu | OH | OH | --- | OH | Glu | --- | <30% 50 μM | <30% 50 μM | ||
| GROUP E (Flavones) | 36 |  | --- | OH | --- | --- | OH | OH | OH | 20 ± 3 | 7 ± 1 |
| 37 Robinetin | OH | --- | OH | --- | OH | OH | OH | (a) | (a) | ||
| 38 | OH | OH | OH | --- | OH | OH | OH | 16 ± 1 | 25 ± 2 | ||
| GROUP F (Flavan-3-ols) | 39 (–)-Epicatechin | 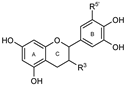 | OH | OH | OH | --- | OH | OH | --- | <30% 100 μM | <30% 100 μM |
| 40 (–)-Epigallocatechin | OH | OH | OH | --- | OH | OH | OH | <30% 100 μM | <30% 100 μM | ||
| 41(–)-Epigallocatechin gallate | Gal | OH | OH | --- | OH | OH | OH | 32 ± 4 | <30% 100 μM | ||
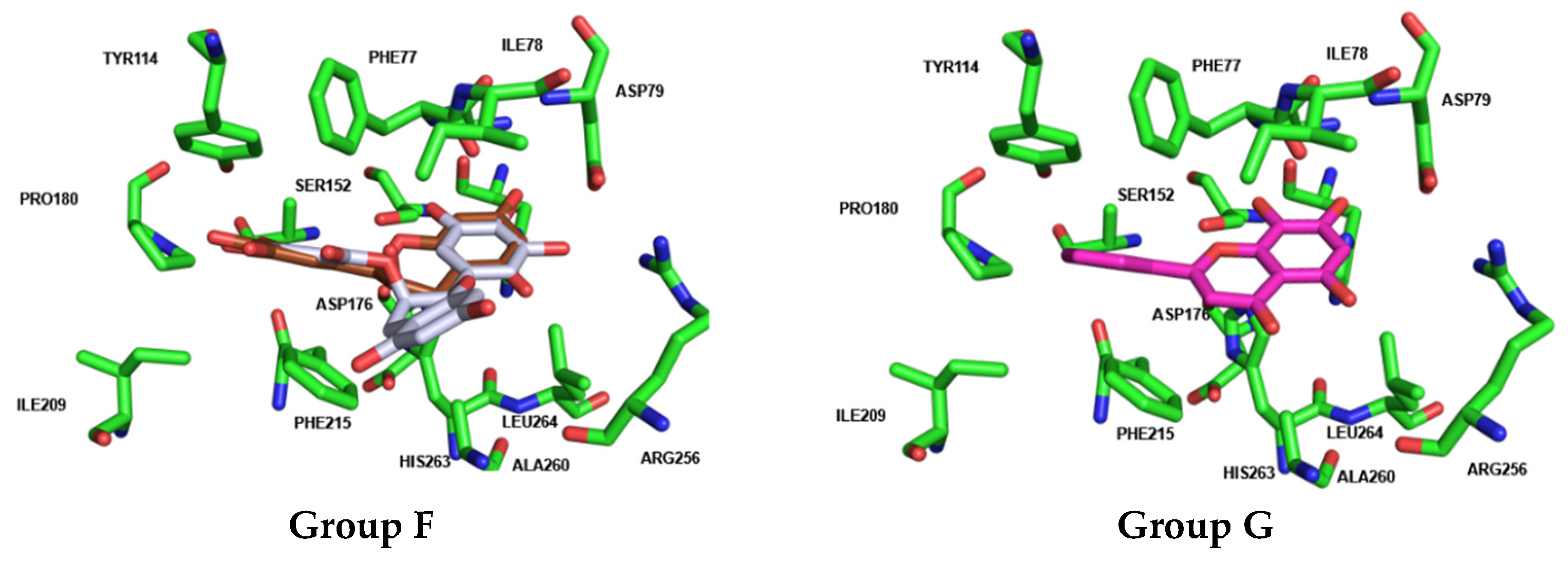
| GROUP G (Flavones) | 42 Norwogonin | 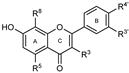 | --- | OH | OH | OH | --- | --- | --- | 98 ± 9 | 79 ± 3 |
| 43 | --- | --- | OH | OH | OH | OH | --- | <30% 50 μM | <30% 50 μM | ||
| 44 | OH | --- | OH | OH | --- | OH | --- | 42 ± 2% 50 μM | <30% 50 μM | ||
| 45 Melanoxetin | OH | --- | OH | OH | OH | OH | --- | <30% 25 μM | <30% 25 μM | ||
| 46 Herbacetin | OH | OH | OH | OH | --- | OH | --- | 46 ± 4% 50 μM | 39 ± 3% 50 μM | ||
| 47 Gossypetin | OH | OH | OH | OH | OH | OH | --- | 36 ± 3% 50 μM | 37 ± 1% 50 μM | ||
| 48 Gossypin | OH | OH | OH | Glu | OH | OH | --- | <30% 12.5 μM | <30% 12.5 μM | ||
| Orlistat |  | 0.0043 ± 0.0003 | 0.00068 ± 0.00003 | ||||||||
Glucosyl (Glu): | Rutinosyl (Rut):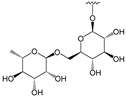 | Galloyl (Gal):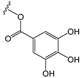 | |||||||||
| Compound | Inhibition Model | Vmax | Km | Kic | Kiu |
|---|---|---|---|---|---|
| 27 (Quercetin) | Competitive | 354 ± 3 | 26.0 ± 0.8 | 13.1 ± 0.5 | - |
| 30 | Competitive | 507 ± 4 | 26.0 ± 0.7 | 1.78 ± 0.04 | - |
| 38 (Myricetin) | Mixed | 655 ± 5 | 23.0 ± 0.7 | 12.0 ± 0.8 | 58 ± 3 |
| 41 (EGCG) | Competitive | 680 ± 7 | 21 ± 2 | 21 ± 3 | - |
| Compound | Inhibition Model | Vmax | Km | Kic | Kic |
|---|---|---|---|---|---|
| 30 | Mixed | 260 ± 10 | 14 ± 2 | 1.3 ± 0.3 | 16 ± 3 |
| 38 (Myricetin) | Noncompetitive | 230 ± 1 | 9.6 ± 0.1 | 74 ± 1 | 74 ± 1 |
| Compound | 15 | 36 | 14 | 38 | 42 | 28 | 29 | 30 | Orlistat |
|---|---|---|---|---|---|---|---|---|---|
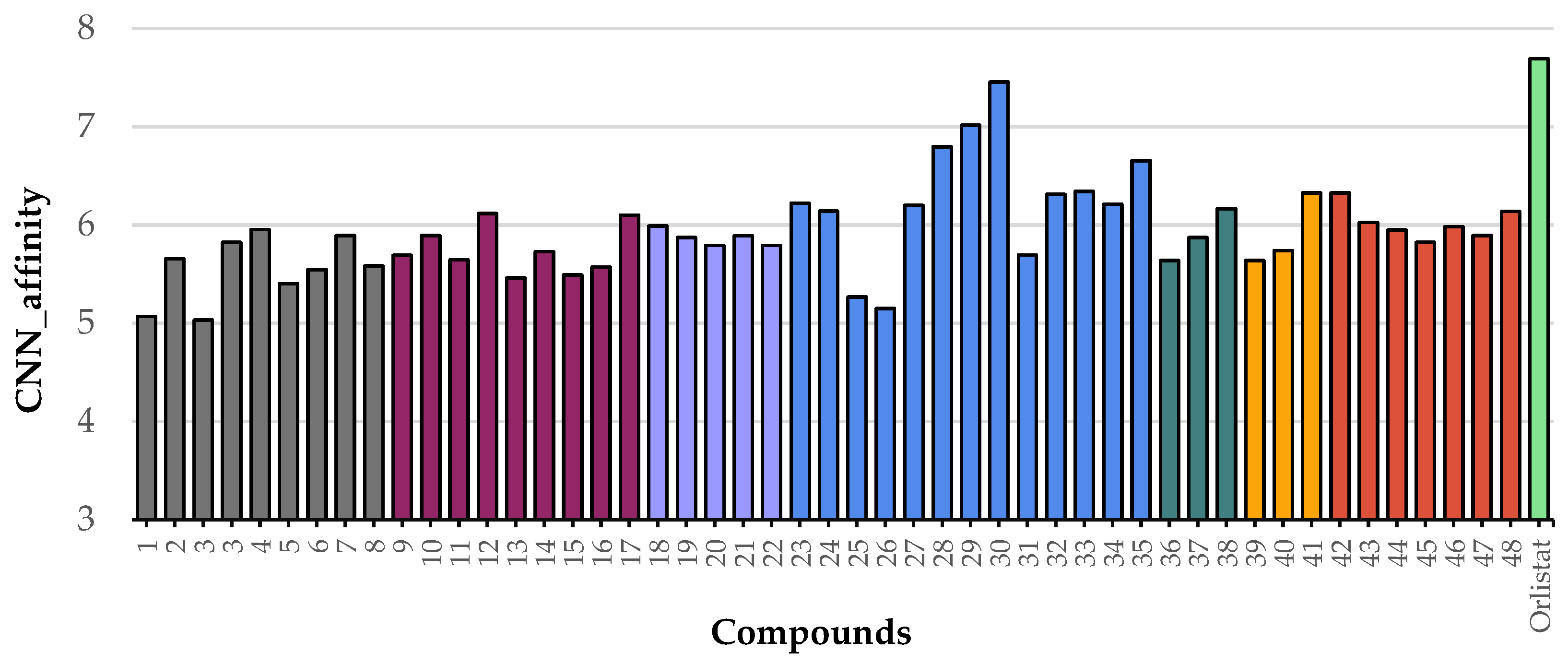
| GNINA Score (CNN affinity) | 5.493 | 5.639 | 5.728 | 6.167 | 6.328 | 6.796 | 7.015 | 7.455 | 7.693 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocha, S.; Proença, C.; Araújo, A.N.; Freitas, M.; Rufino, I.; Aniceto, N.; Silva, A.M.S.; Carvalho, F.; Guedes, R.C.; Fernandes, E. Flavonoids as Potential Modulators of Pancreatic Lipase Catalytic Activity. Pharmaceutics 2025, 17, 163. https://doi.org/10.3390/pharmaceutics17020163
Rocha S, Proença C, Araújo AN, Freitas M, Rufino I, Aniceto N, Silva AMS, Carvalho F, Guedes RC, Fernandes E. Flavonoids as Potential Modulators of Pancreatic Lipase Catalytic Activity. Pharmaceutics. 2025; 17(2):163. https://doi.org/10.3390/pharmaceutics17020163
Chicago/Turabian StyleRocha, Sílvia, Carina Proença, Alberto N. Araújo, Marisa Freitas, Ismael Rufino, Natália Aniceto, Artur M. S. Silva, Félix Carvalho, Rita C. Guedes, and Eduarda Fernandes. 2025. "Flavonoids as Potential Modulators of Pancreatic Lipase Catalytic Activity" Pharmaceutics 17, no. 2: 163. https://doi.org/10.3390/pharmaceutics17020163
APA StyleRocha, S., Proença, C., Araújo, A. N., Freitas, M., Rufino, I., Aniceto, N., Silva, A. M. S., Carvalho, F., Guedes, R. C., & Fernandes, E. (2025). Flavonoids as Potential Modulators of Pancreatic Lipase Catalytic Activity. Pharmaceutics, 17(2), 163. https://doi.org/10.3390/pharmaceutics17020163