

Antibiofilm Effect of Lavandula multifida Essential Oil: A New Approach for Chronic Infections

 ,
,  ,
,  and
and 
Abstract
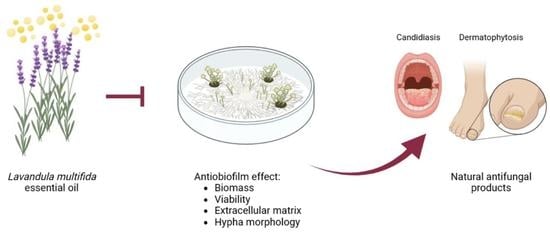
1. Introduction
2. Materials and Methods
2.1. Plant Material and Essential Oil Characterization
2.2. Antifungal Effect of L. multifida Essential Oil
2.2.1. Fungal Strains
2.2.2. Fungal Inoculum
2.2.3. Effect of L. multifida Essential Oil on Biofilm Formation
2.2.4. Effect of the L. multifida Essential Oil on Mature Biofilms
2.2.5. Biofilm Mass
2.2.6. Extracellular Matrix
2.2.7. Biofilm Viability
2.2.8. Biofilm Morphology and Ultrastructure
2.3. Statistical Analysis
3. Results
3.1. Chemical Composition of L. multifida Essential Oil
3.2. Effect of L. multifida Essential Oil on the Formation of Dermatophyte Biofilms
3.3. Effect of L. multifida Essential Oil on the Disruption of Dermatophyte Mature Biofilms
3.4. Morphological Effect of L. multifida Essential Oil on E. floccosum
3.5. Effect of L. multifida Essential Oil on the Formation of Candida albicans Biofilms
3.6. Effect of L. multifida Essential Oil on the Disruption of Candida albicans Mature Biofilms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bongomin, F.; Gago, S.; Oladele, R.; Denning, D. Global and Multi-National Prevalence of Fungal Diseases—Estimate Precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef]
- GAFFI. Global Action Fund for Fungal Infections. Available online: https://gaffi.org/why/fungal-disease-frequency/ (accessed on 6 April 2023).
- World Health Organization. WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action; World Health Organization: Geneva, Switzerland, 2022.
- Daniel, C.R.; Norton, L.A.; Scher, R.K. The Spectrum of Nail Disease in Patients with Human Immunodeficiency Virus Infection. J. Am. Acad. Dermatol. 1992, 27, 93–97. [Google Scholar] [CrossRef]
- Zuzarte, M.; Lopes, G.; Pinto, E.; Salgueiro, L. Are natural products an alternative therapy for dermatophytosis? In Dermatophytes and Dermatophytoses; Springer International Publishing: Cham, Switzerland, 2021; pp. 473–519. [Google Scholar]
- Havlickova, B.; Czaika, V.A.; Friedrich, M. Epidemiological Trends in Skin Mycoses Worldwide. Mycoses 2008, 51, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Cooper, E.A. Update in Antifungal Therapy of Dermatophytosis. Mycopathologia 2008, 166, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Markantonatou, A.-M.; Samaras, K.; Vyzantiadis, T.-A. Dermatophytic Biofilms: Characteristics, Significance and Treatment Approaches. J. Fungi 2023, 9, 228. [Google Scholar] [CrossRef] [PubMed]
- Brilhante, R.S.N.; Correia, E.E.M.; de Guedes, G.M.M.; Pereira, V.S.; de Oliveira, J.S.; Bandeira, S.P.; de Alencar, L.P.; de Andrade, A.R.C.; de Castelo-Branco, D.S.C.M.; de Cordeiro, R.A.; et al. Quantitative and Structural Analyses of the in Vitro and Ex Vivo Biofilm-Forming Ability of Dermatophytes. J. Med. Microbiol. 2017, 66, 1045–1052. [Google Scholar] [CrossRef]
- Kullberg, B.J.; Arendrup, M.C. Invasive Candidiasis. N. Engl. J. Med. 2015, 373, 1445–1456. [Google Scholar] [CrossRef]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive Candidiasis. Nat. Rev. Dis. Primers 2018, 4, 18026. [Google Scholar] [CrossRef]
- Atriwal, T.; Azeem, K.; Husain, F.M.; Hussain, A.; Khan, M.N.; Alajmi, M.F.; Abid, M. Mechanistic Understanding of Candida albicans Biofilm Formation and Approaches for Its Inhibition. Front. Microbiol. 2021, 12, 638609. [Google Scholar] [CrossRef]
- Kuhn, D.M.; Ghannoum, M.A. Candida Biofilms: Antifungal Resistance and Emerging Therapeutic Options. Curr. Opin. Investig. Drugs 2004, 5, 186–197. [Google Scholar] [PubMed]
- Gulati, M.; Nobile, C.J. Candida albicans Biofilms: Development, Regulation, and Molecular Mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Denning, D.W.; Gow, N.A.R.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden Killers: Human Fungal Infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef]
- Rodrigues, M.L.; Nosanchuk, J.D. Fungal Diseases as Neglected Pathogens: A Wake-up Call to Public Health Officials. PLoS Negl. Trop. Dis. 2020, 14, e0007964. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Solache, M.A.; Casadevall, A. Global Warming Will Bring New Fungal Diseases for Mammals. mBio 2010, 1, e00061-10. [Google Scholar] [CrossRef]
- Cohen, J.M.; Venesky, M.D.; Sauer, E.L.; Civitello, D.J.; McMahon, T.A.; Roznik, E.A.; Rohr, J.R. The Thermal Mismatch Hypothesis Explains Host Susceptibility to an Emerging Infectious Disease. Ecol. Lett. 2017, 20, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Zuzarte, M.; Gonçalves, M.J.; Cruz, M.T.; Cavaleiro, C.; Canhoto, J.; Vaz, S.; Pinto, E.; Salgueiro, L. Lavandula Luisieri Essential Oil as a Source of Antifungal Drugs. Food Chem. 2012, 135, 1505–1510. [Google Scholar] [CrossRef]
- Zuzarte, M.; Gonçalves, M.J.; Cavaleiro, C.; Dinis, A.M.; Canhoto, J.M.; Salgueiro, L.R. Chemical Composition and Antifungal Activity of the Essential Oils of Lavandula Pedunculata (Miller) Cav. Chem. Biodivers. 2009, 6, 1283–1292. [Google Scholar] [CrossRef]
- El Abdali, Y.; Agour, A.; Allali, A.; Bourhia, M.; El Moussaoui, A.; Eloutassi, N.; Salamatullah, A.M.; Alzahrani, A.; Ouahmane, L.; Aboul-Soud, M.A.M.; et al. Lavandula Dentata L.: Phytochemical Analysis, Antioxidant, Antifungal and Insecticidal Activities of Its Essential Oil. Plants 2022, 11, 311. [Google Scholar] [CrossRef]
- D’Auria, F.D.; Tecca, M.; Strippoli, V.; Salvatore, G.; Battinelli, L.; Mazzanti, G. Antifungal Activity of Lavandula Angustifolia Essential Oil against Candida albicans Yeast and Mycelial Form. Med. Mycol. 2005, 43, 391–396. [Google Scholar] [CrossRef]
- Zuzarte, M.; Gonçalves, M.J.; Cavaleiro, C.; Cruz, M.T.; Benzarti, A.; Marongiu, B.; Maxia, A.; Piras, A.; Salgueiro, L. Antifungal and Anti-Inflammatory Potential of Lavandula Stoechas and Thymus Herba-Barona Essential Oils. Ind. Crops Prod. 2013, 44, 97–103. [Google Scholar] [CrossRef]
- Minooeianhaghighi, M.H.; Sepehrian, L.; Shokri, H. Antifungal Effects of Lavandula Binaludensis and Cuminum Cyminum Essential Oils against Candida albicans Strains Isolated from Patients with Recurrent Vulvovaginal Candidiasis. J. Mycol. Med. 2017, 27, 65–71. [Google Scholar] [CrossRef]
- Domingues, J.; Goulão, M.; Delgado, F.; Gonçalves, J.C.; Gonçalves, J.; Pintado, C.S. Essential Oils of Two Portuguese Endemic Species of Lavandula as a Source of Antifungal and Antibacterial Agents. Processes 2023, 11, 1165. [Google Scholar] [CrossRef]
- Puškárová, A.; Bučková, M.; Kraková, L.; Pangallo, D.; Kozics, K. The Antibacterial and Antifungal Activity of Six Essential Oils and Their Cyto/Genotoxicity to Human HEL 12469 Cells. Sci. Rep. 2017, 7, 8211. [Google Scholar] [CrossRef] [PubMed]
- Zuzarte, M.; Vale-Silva, L.; Gonçalves, M.J.; Cavaleiro, C.; Vaz, S.; Canhoto, J.; Pinto, E.; Salgueiro, L. Antifungal Activity of Phenolic-Rich Lavandula multifida L. Essential Oil. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Rajendran, R.; Sherry, L.; Williams, C. Fungal Biofilm Resistance. Int. J. Microbiol. 2012, 2012, 528521. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Vande Walle, K.; Wickes, B.L.; López-Ribot, J.L. Standardized Method for In Vitro Antifungal Susceptibility Testing of Candida albicans Biofilms. Antimicrob. Agents Chemother. 2001, 45, 2475–2479. [Google Scholar] [CrossRef]
- Yazdanpanah, S.; Sasanipoor, F.; Khodadadi, H.; Rezaei-Matehkolaei, A.; Jowkar, F.; Zomorodian, K.; Kharazi, M.; Mohammadi, T.; Nouripour-Sisakht, S.; Nasr, R.; et al. Quantitative Analysis of in Vitro Biofilm Formation by Clinical Isolates of Dermatophyte and Antibiofilm Activity of Common Antifungal Drugs. Int. J. Dermatol. 2023, 62, 120–127. [Google Scholar] [CrossRef]
- Brilhante, R.S.N.; Correia, E.E.M.; de Guedes, G.M.M.; de Oliveira, J.S.; de Castelo-Branco, D.S.C.M.; de Cordeiro, R.A.; de Pinheiro, A.Q.; Chaves, L.J.Q.; de Pereira Neto, W.A.; Sidrim, J.J.C.; et al. In Vitro Activity of Azole Derivatives and Griseofulvin against Planktonic and Biofilm Growth of Clinical Isolates of Dermatophytes. Mycoses 2018, 61, 449–454. [Google Scholar] [CrossRef]
- Kernien, J.F.; Snarr, B.D.; Sheppard, D.C.; Nett, J.E. The Interface between Fungal Biofilms and Innate Immunity. Front Immunol 2018, 8, 1968. [Google Scholar] [CrossRef]
- Cavaleiro, C.; Salgueiro, L.R.; Miguel, M.G.; Proença da Cunha, A. Analysis by Gas Chromatography–Mass Spectrometry of the Volatile Components of Teucrium Lusitanicum and Teucrium Algarbiensis. J. Chromatogr. A 2004, 1033, 187–190. [Google Scholar] [CrossRef]
- Ali, I.; Satti, N.K.; Dutt, P.; Prasad, R.; Khan, I.A. Hydroxychavicol: A Phytochemical Targeting Cutaneous Fungal Infections. Sci. Rep. 2016, 6, 37867. [Google Scholar] [CrossRef] [PubMed]
- de Castelo-Branco, D.S.C.M.; de Aguiar, L.; dos Araújo, G.S.; Lopes, R.G.P.; de Sales, J.A.; Pereira-Neto, W.A.; de Pinheiro, A.Q.; Paixão, G.C.; de Cordeiro, R.A.; Sidrim, J.J.C.; et al. In Vitro and Ex Vivo Biofilms of Dermatophytes: A New Panorama for the Study of Antifungal Drugs. Biofouling 2020, 36, 783–791. [Google Scholar] [CrossRef]
- Raut, J.S.; Shinde, R.B.; Chauhan, N.M.; Karuppayil, S.M. Terpenoids of Plant Origin Inhibit Morphogenesis, Adhesion, and Biofilm Formation by Candida albicans. Biofouling 2012, 29, 87–96. [Google Scholar] [CrossRef]
- Costa-Orlandi, C.B.; Sardi, J.C.O.; Santos, C.T.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. In Vitro Characterization of Trichophyton Rubrum and T. Mentagrophytes Biofilms. Biofouling 2014, 30, 719–727. [Google Scholar] [CrossRef]
- Alves, M.; Gonçalves, M.J.; Zuzarte, M.; Alves-Silva, J.M.; Cavaleiro, C.; Cruz, M.T.; Salgueiro, L. Unveiling the Antifungal Potential of Two Iberian Thyme Essential Oils: Effect on C. Albicans Germ Tube and Preformed Biofilms. Front. Pharmacol. 2019, 10, 446. [Google Scholar] [CrossRef]
- Jabra-Rizk, M.A.; Falkler, W.A.; Meiller, T.F. Fungal Biofilms and Drug Resistance. Emerg. Infect. Dis. 2004, 10, 14–19. [Google Scholar] [CrossRef]
- Martinez-Rossi, N.M.; Peres, N.T.A.; Bitencourt, T.A.; Martins, M.P.; Rossi, A. State-of-the-Art Dermatophyte Infections: Epidemiology Aspects, Pathophysiology, and Resistance Mechanisms. J. Fungi 2021, 7, 629. [Google Scholar] [CrossRef]
- Wang, R.; Huang, C.; Zhang, Y.; Li, R. Invasive Dermatophyte Infection: A Systematic Review. Mycoses 2021, 64, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Elmakaoui, A.; Bourais, I.; Oubihi, A.; Nassif, A.; Bezhinar, T.; Shariati, M.A.; Blinov, A.V.; Hleba, L.; Hajjaji, S. El Chemical Composition and Antibacterial Activity of Essential Oil of Lavandula multifida. J. Microbiol. Biotechnol. Food Sci. 2022, 11, e7559. [Google Scholar] [CrossRef]
- Chograni, H.; Zaouali, Y.; Rajeb, C.; Boussaid, M. Essential Oil Variation among Natural Populations of Lavandula multifida L. (Lamiaceae). Chem. Biodivers. 2010, 7, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Messaoud, C.; Chograni, H.; Boussaid, M. Chemical Composition and Antioxidant Activities of Essential Oils and Methanol Extracts of Three Wild Lavandula L. Species. Nat. Prod. Res. 2012, 26, 1976–1984. [Google Scholar] [CrossRef]
- Mostefa, M.B.; Kabouche, A.; Abaza, I.; Aburjai, T.; Touzani, R.; Kabouche, Z. Chemotypes Investigation of Lavandula Essential Oils Growing at Different North African Soils. J. Mater. Environ. Sci 2014, 5, 1896–1901. [Google Scholar]
- Sellam, K.; Ramchoun, M.; Alem, C.; El-Rhaffari, L. Biological Investigations of Antioxidant-Antimicrobial Properties and Chemical Composition of Essential Oil from Lavandula multifida. Oxid. Antioxid. Med. Sci. 2013, 2, 211–216. [Google Scholar] [CrossRef]
- Gupta, A.K.; Carviel, J.; Shear, N.H. Antibiofilm Treatment for Onychomycosis and Chronic Fungal Infections. Ski. Appendage Disord. 2018, 4, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Dalla Lana, D.F.; Batista, B.G.; Alves, S.H.; Fuentefria, A.M. Dermatofitoses: Agentes Etiológicos, Formas Clínicas, Terapêutica e Novas Perspectivas de Tratamento. Clin. Biomed. Res. 2016, 36, 230–241. [Google Scholar] [CrossRef]
- Petrucelli, M.F.; de Abreu, M.H.; Cantelli, B.A.M.; Segura, G.G.; Nishimura, F.G.; Bitencourt, T.A.; Marins, M.; Fachin, A.L. Epidemiology and Diagnostic Perspectives of Dermatophytoses. J. Fungi 2020, 6, 310. [Google Scholar] [CrossRef] [PubMed]
- Alves-Silva, J.M.; Zuzarte, M.; Gonçalves, M.J.; Cruz, M.T.; Cavaleiro, C.; Salgueiro, L. Unveiling the Bioactive Potential of the Essential Oil of a Portuguese Endemism, Santolina Impressa. J. Ethnopharmacol. 2019, 244, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Müller-Sepúlveda, A.; Chevecich, C.C.; Jara, J.A.; Belmar, C.; Sandoval, P.; Meyer, R.S.; Quijada, R.; Moura, S.; López-Muñoz, R.; Díaz-Dosque, M.; et al. Chemical Characterization of Lavandula Dentata Essential Oil Cultivated in Chile and Its Antibiofilm Effect against Candida albicans. Planta Med. 2020, 86, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Karpiński, T.M.; Ożarowski, M.; Seremak-Mrozikiewicz, A.; Wolski, H. Anti-Candida and Antibiofilm Activity of Selected Lamiaceae Essential Oils. Front. Biosci. Landmark 2023, 28, 28. [Google Scholar] [CrossRef] [PubMed]
- Iseppi, R.; Tardugno, R.; Brighenti, V.; Benvenuti, S.; Sabia, C.; Pellati, F.; Messi, P. Phytochemical Composition and In Vitro Antimicrobial Activity of Essential Oils from the Lamiaceae Family against Streptococcus Agalactiae and Candida albicans Biofilms. Antibiotics 2020, 9, 592. [Google Scholar] [CrossRef]
- Dolatabadi, S.; Salari, Z.; Mahboubi, M. Antifungal Effects of Ziziphora Tenuior, Lavandula Angustifolia, Cuminum Cyminum Essential Oils against Clinical Isolates of Candida albicans from Women Suffering from Vulvovaginal Candidiasis. Infectio 2019, 23, 222. [Google Scholar] [CrossRef]
- Ramić, D.; Bucar, F.; Kunej, U.; Dogša, I.; Klančnik, A.; Smole Možina, S. Antibiofilm Potential of Lavandula Preparations against Campylobacter Jejuni. Appl. Environ. Microbiol. 2021, 87, AEM0109921. [Google Scholar] [CrossRef] [PubMed]
- Kot, B.; Wierzchowska, K.; Grużewska, A.; Lohinau, D. The Effects of Selected Phytochemicals on Biofilm Formed by Five Methicillin-Resistant Staphylococcus aureus. Nat. Prod. Res. 2018, 32, 1299–1302. [Google Scholar] [CrossRef] [PubMed]
- Budzyńska, A.; Wieckowska-Szakiel, M.; Sadowska, B.; Kalemba, D.; Rózalska, B. Antibiofilm Activity of Selected Plant Essential Oils and Their Major Components. Pol. J. Microbiol. 2011, 60, 35–41. [Google Scholar] [CrossRef] [PubMed]
- El Kheloui, R.; Laktib, A.; Elmegdar, S.; Fayzi, L.; Zanane, C.; Msanda, F.; Cherifi, K.; Latrache, H.; Mimouni, R.; Hamadi, F. Anti-Adhesion and Antibiofilm Activities of Lavandula Mairei Humbert Essential Oil against Acinetobacter Baumannii Isolated from Hospital Intensive Care Units. Biofouling 2022, 38, 953–964. [Google Scholar] [CrossRef]
- Takarada, K.; Kimizuka, R.; Takahashi, N.; Honma, K.; Okuda, K.; Kato, T. A Comparison of the Antibacterial Efficacies of Essential Oils against Oral Pathogens. Oral. Microbiol. Immunol. 2004, 19, 61–64. [Google Scholar] [CrossRef]
- Kumari, P.; Mishra, R.; Arora, N.; Chatrath, A.; Gangwar, R.; Roy, P.; Prasad, R. Antifungal and Anti-Biofilm Activity of Essential Oil Active Components against Cryptococcus neoformans and Cryptococcus Laurentii. Front. Microbiol. 2017, 8, 2161. [Google Scholar] [CrossRef]
- Kumari, P.; Arora, N.; Chatrath, A.; Gangwar, R.; Pruthi, V.; Poluri, K.M.; Prasad, R. Delineating the Biofilm Inhibition Mechanisms of Phenolic and Aldehydic Terpenes against Cryptococcus neoformans. ACS Omega 2019, 4, 17634–17648. [Google Scholar] [CrossRef]
- Yarkarami, F.; Kazemian, H.; Sadeghifard, N.; Pakzad, R.; Jalilian, F.; Asadollahi, P.; Hematian, A.; Pakzad, I. Inhibitory Effects of Carvacrol on Biofilm Formation and Expression of Biofilm Related Genes in Clinical Isolates of Enterococcus Faecalis. Clin. Lab. 2022, 68. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, P.; Sienkiewicz, M.; Pruss, A.; Łopusiewicz, Ł.; Arszyńska, N.; Wojciechowska-Koszko, I.; Kilanowicz, A.; Kot, B.; Dołęgowska, B. Antibacterial and Anti-Biofilm Activities of Essential Oil Compounds against New Delhi Metallo-β-Lactamase-1-Producing Uropathogenic Klebsiella Pneumoniae Strains. Antibiotics 2022, 11, 147. [Google Scholar] [CrossRef] [PubMed]
- Walczak, M.; Michalska-Sionkowska, M.; Olkiewicz, D.; Tarnawska, P.; Warżyńska, O. Potential of Carvacrol and Thymol in Reducing Biofilm Formation on Technical Surfaces. Molecules 2021, 26, 2723. [Google Scholar] [CrossRef]
- Maquera-Huacho, P.M.; Tonon, C.C.; Correia, M.F.; Francisconi, R.S.; Bordini, E.A.F.; Marcantonio, É.; Spolidorio, D.M.P. In Vitro Antibacterial and Cytotoxic Activities of Carvacrol and Terpinen-4-Ol against Biofilm Formation on Titanium Implant Surfaces. Biofouling 2018, 34, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Ciandrini, E.; Campana, R.; Federici, S.; Manti, A.; Battistelli, M.; Falcieri, E.; Papa, S.; Baffone, W. In Vitro Activity of Carvacrol against Titanium-Adherent Oral Biofilms and Planktonic Cultures. Clin. Oral Investig. 2014, 18, 2001–2013. [Google Scholar] [CrossRef]
- Wang, J.; Qin, T.; Chen, K.; Pan, L.; Xie, J.; Xi, B. Antimicrobial and Antivirulence Activities of Carvacrol against Pathogenic Aeromonas Hydrophila. Microorganisms 2022, 10, 2170. [Google Scholar] [CrossRef]
- Walsh, D.J.; Livinghouse, T.; Goeres, D.M.; Mettler, M.; Stewart, P.S. Antimicrobial Activity of Naturally Occurring Phenols and Derivatives Against Biofilm and Planktonic Bacteria. Front. Chem. 2019, 7, 653. [Google Scholar] [CrossRef] [PubMed]
- Sousa, L.G.V.; Castro, J.; Cavaleiro, C.; Salgueiro, L.; Tomás, M.; Palmeira-Oliveira, R.; Martinez-Oliveira, J.; Cerca, N. Synergistic Effects of Carvacrol, α-Terpinene, γ-Terpinene, ρ-Cymene and Linalool against Gardnerella Species. Sci. Rep. 2022, 12, 4417. [Google Scholar] [CrossRef] [PubMed]
- Marini, E.; Di Giulio, M.; Ginestra, G.; Magi, G.; Di Lodovico, S.; Marino, A.; Facinelli, B.; Cellini, L.; Nostro, A. Efficacy of Carvacrol against Resistant Rapidly Growing Mycobacteria in the Planktonic and Biofilm Growth Mode. PLoS ONE 2019, 14, e0219038. [Google Scholar] [CrossRef]
- Čabarkapa, I.; Čolović, R.; Đuragić, O.; Popović, S.; Kokić, B.; Milanov, D.; Pezo, L. Anti-Biofilm Activities of Essential Oils Rich in Carvacrol and Thymol against Salmonella Enteritidis. Biofouling 2019, 35, 361–375. [Google Scholar] [CrossRef]
- Nostro, A.; Cellini, L.; Zimbalatti, V.; Blanco, A.R.; Marino, A.; Pizzimenti, F.; Di Giulio, M.; Bisignano, G. Enhanced Activity of Carvacrol against Biofilm of Staphylococcus aureus and Staphylococcus Epidermidis in an Acidic Environment. APMIS 2012, 120, 967–973. [Google Scholar] [CrossRef]
- Mechmechani, S.; Gharsallaoui, A.; El Omari, K.; Fadel, A.; Hamze, M.; Chihib, N.-E. Hurdle Technology Based on the Use of Microencapsulated Pepsin, Trypsin and Carvacrol to Eradicate Pseudomonas Aeruginosa and Enterococcus Faecalis Biofilms. Biofouling 2022, 38, 903–915. [Google Scholar] [CrossRef]
- Mechmechani, S.; Gharsallaoui, A.; Fadel, A.; El Omari, K.; Khelissa, S.; Hamze, M.; Chihib, N.-E. Microencapsulation of Carvacrol as an Efficient Tool to Fight Pseudomonas Aeruginosa and Enterococcus Faecalis Biofilms. PLoS ONE 2022, 17, e0270200. [Google Scholar] [CrossRef] [PubMed]
- Bernal-Mercado, A.T.; Juarez, J.; Valdez, M.A.; Ayala-Zavala, J.F.; Del-Toro-Sánchez, C.L.; Encinas-Basurto, D. Hydrophobic Chitosan Nanoparticles Loaded with Carvacrol against Pseudomonas Aeruginosa Biofilms. Molecules 2022, 27, 699. [Google Scholar] [CrossRef]
- Tapia-Rodriguez, M.R.; Bernal-Mercado, A.T.; Gutierrez-Pacheco, M.M.; Vazquez-Armenta, F.J.; Hernandez-Mendoza, A.; Gonzalez-Aguilar, G.A.; Martinez-Tellez, M.A.; Nazzaro, F.; Ayala-Zavala, J.F. Virulence of Pseudomonas Aeruginosa Exposed to Carvacrol: Alterations of the Quorum Sensing at Enzymatic and Gene Levels. J. Cell Commun. Signal. 2019, 13, 531–537. [Google Scholar] [CrossRef]
- Engel, J.B.; Heckler, C.; Tondo, E.C.; Daroit, D.J.; da Silva Malheiros, P. Antimicrobial Activity of Free and Liposome-Encapsulated Thymol and Carvacrol against Salmonella and Staphylococcus aureus Adhered to Stainless Steel. Int. J. Food. Microbiol. 2017, 252, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Nostro, A.; Scaffaro, R.; Botta, L.; Filocamo, A.; Marino, A.; Bisignano, G. Effect of Temperature on the Release of Carvacrol and Cinnamaldehyde Incorporated into Polymeric Systems to Control Growth and Biofilms of Escherichia Coli and Staphylococcus aureus. Biofouling 2015, 31, 639–649. [Google Scholar] [CrossRef]
- Nostro, A.; Scaffaro, R.; D’Arrigo, M.; Botta, L.; Filocamo, A.; Marino, A.; Bisignano, G. Study on Carvacrol and Cinnamaldehyde Polymeric Films: Mechanical Properties, Release Kinetics and Antibacterial and Antibiofilm Activities. Appl. Microbiol. Biotechnol. 2012, 96, 1029–1038. [Google Scholar] [CrossRef]
- Iannitelli, A.; Grande, R.; Di Stefano, A.; Di Giulio, M.; Sozio, P.; Bessa, L.J.; Laserra, S.; Paolini, C.; Protasi, F.; Cellini, L. Potential Antibacterial Activity of Carvacrol-Loaded Poly(DL-Lactide-Co-Glycolide) (PLGA) Nanoparticles against Microbial Biofilm. Int. J. Mol. Sci. 2011, 12, 5039–5051. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Conesa, D.; Cao, J.; Chen, L.; McLandsborough, L.; Weiss, J. Inactivation of Listeria Monocytogenes and Escherichia Coli O157:H7 Biofilms by Micelle-Encapsulated Eugenol and Carvacrol. J. Food Prot. 2011, 74, 55–62. [Google Scholar] [CrossRef]
- Pérez-Conesa, D.; Mclandsborough, L.; Weiss, J. Inhibition and Inactivation of Listeria Monocytogenes and Escherichia Coli O157:H7 Colony Biofilms by Micellar-Encapsulated Eugenol and Carvacrol. J. Food Prot. 2006, 69, 2947–2954. [Google Scholar] [CrossRef] [PubMed]
- Castagliuolo, G.; Di Napoli, M.; Vaglica, A.; Badalamenti, N.; Antonini, D.; Varcamonti, M.; Bruno, M.; Zanfardino, A.; Bazan, G. Thymus richardii subsp. Nitidus (Guss.) Jalas Essential Oil: An Ally against Oral Pathogens and Mouth Health. Molecules 2023, 28, 4803. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RI † | RI ‡ | Compound | % |
|---|---|---|---|
| 931 | 1029 | α-pinene | 1.5 |
| 961 | 1441 | 1-octen-3-ol | 1.9 |
| 972 | 1127 | β-pinene | 0.1 |
| 982 | 1161 | myrcene | 5.9 |
| 1007 | 1152 | 3-carene | 0.7 |
| 1013 | 1272 | ρ-cymene | 1.0 |
| 1022 | 1205 | limonene | 0.5 |
| 1027 | 1233 | cis-β-ocimene | 12.7 |
| 1037 | 1249 | trans-β-ocimene | 0.7 |
| 1054 | octanol | 0.6 | |
| 1067 | 1401 | fenchone | 0.2 |
| 1073 | 1435 | cymenene | 0.7 |
| 1079 | 1285 | terpinolene | 1.3 |
| 1084 | 1381 | nonanal | 0.2 |
| 1084 | 1539 | linalool | 1.5 |
| 1099 | 1577 | fenchol | 0.2 |
| 1118 | 1370 | allo-ocimene | 0.3 |
| 1170 | 1689 | α-terpineol | 0.2 |
| 1281 | 2201 | carvacrol | 46.4 |
| 1411 | 1591 | trans-caryophyllene | 1.4 |
| 1451 | 1637 | allo-aromadendrene | 0.2 |
| 1451 | 1661 | trans-β-farnesene | 0.2 |
| 1469 | 1699 | germacrene D | 0.4 |
| 1495 | 1743 | (E,E)-α-farnesene | 2.3 |
| 1498 | 1720 | β-bisabolene | 10.1 |
| 1510 | 1748 | δ-cadinene | 0.3 |
| 1556 | 2107 | spathulenol | 1.7 |
| 1560 | 1966 | caryophyllene-oxide | 1.0 |
| 1619 | 2170 | T-muurolol | 0.3 |
| 1632 | 2216 | α-cadinol | 0.3 |
| 1662 | 2211 | α-bisabolol | 0.4 |
| Monoterpene hydrocarbons | 25.1 | ||
| Oxygen-containing monoterpenes | 48.8 | ||
| Sesquiterpene hydrocarbons | 14.9 | ||
| Oxygen-containing sesquiterpenes | 3.7 | ||
| Others | 2.7 | ||
| TOTAL | 95.2 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves-Silva, J.; Zuzarte, M.; Cavaleiro, C.; Salgueiro, L. Antibiofilm Effect of Lavandula multifida Essential Oil: A New Approach for Chronic Infections. Pharmaceutics 2023, 15, 2142. https://doi.org/10.3390/pharmaceutics15082142
Alves-Silva J, Zuzarte M, Cavaleiro C, Salgueiro L. Antibiofilm Effect of Lavandula multifida Essential Oil: A New Approach for Chronic Infections. Pharmaceutics. 2023; 15(8):2142. https://doi.org/10.3390/pharmaceutics15082142
Chicago/Turabian StyleAlves-Silva, Jorge, Mónica Zuzarte, Carlos Cavaleiro, and Lígia Salgueiro. 2023. "Antibiofilm Effect of Lavandula multifida Essential Oil: A New Approach for Chronic Infections" Pharmaceutics 15, no. 8: 2142. https://doi.org/10.3390/pharmaceutics15082142
APA StyleAlves-Silva, J., Zuzarte, M., Cavaleiro, C., & Salgueiro, L. (2023). Antibiofilm Effect of Lavandula multifida Essential Oil: A New Approach for Chronic Infections. Pharmaceutics, 15(8), 2142. https://doi.org/10.3390/pharmaceutics15082142