
Candidacidal and Antibiofilm Activity of PS1-3 Peptide against Drug-Resistant Candida albicans on Contact Lenses

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Synthesis of PS1 Analog Peptides
2.3. Antifungicidal Activity
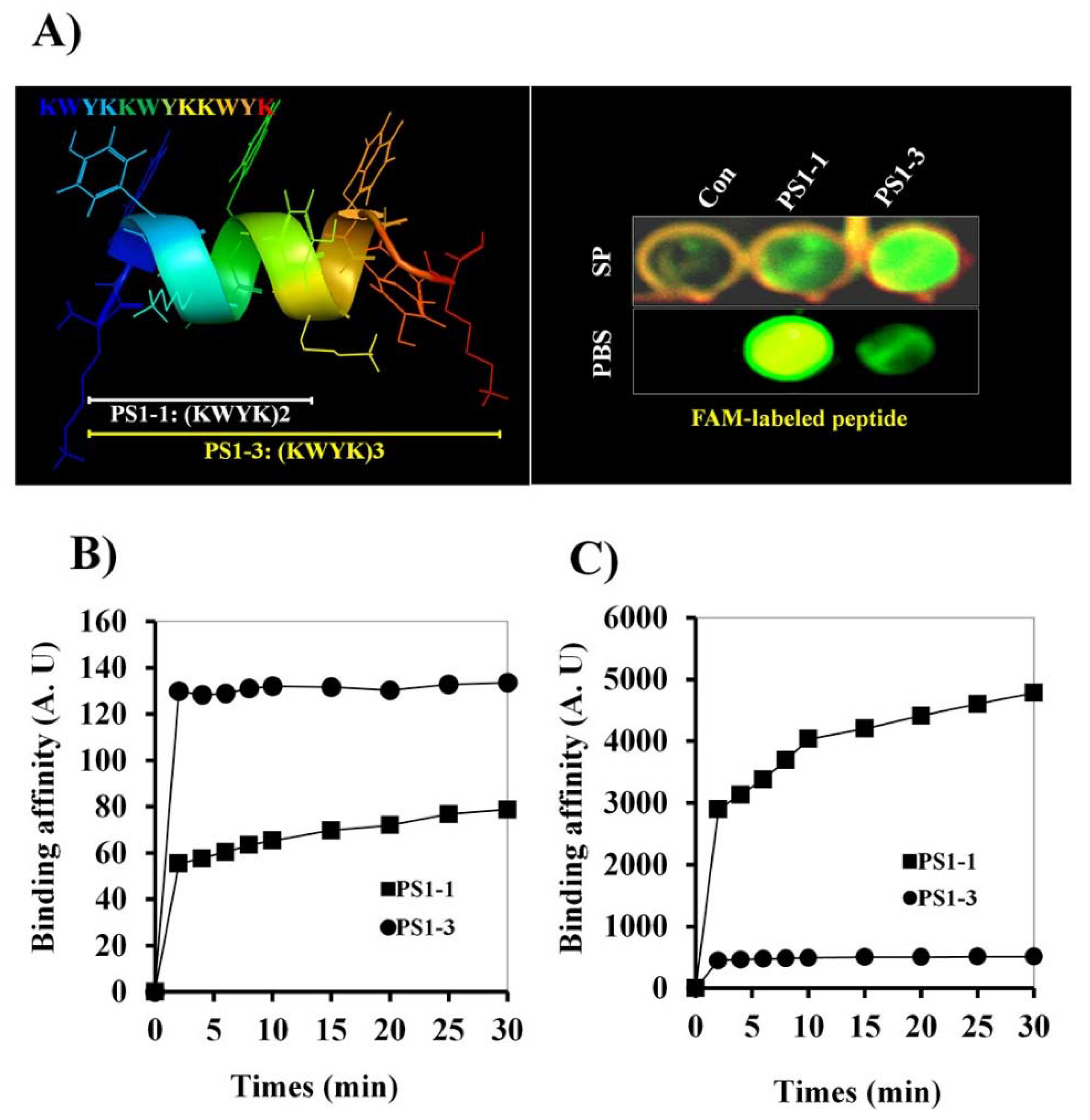
2.4. FAM-PS1-1 and FAM-PS1-3 Peptide Binding Affinity
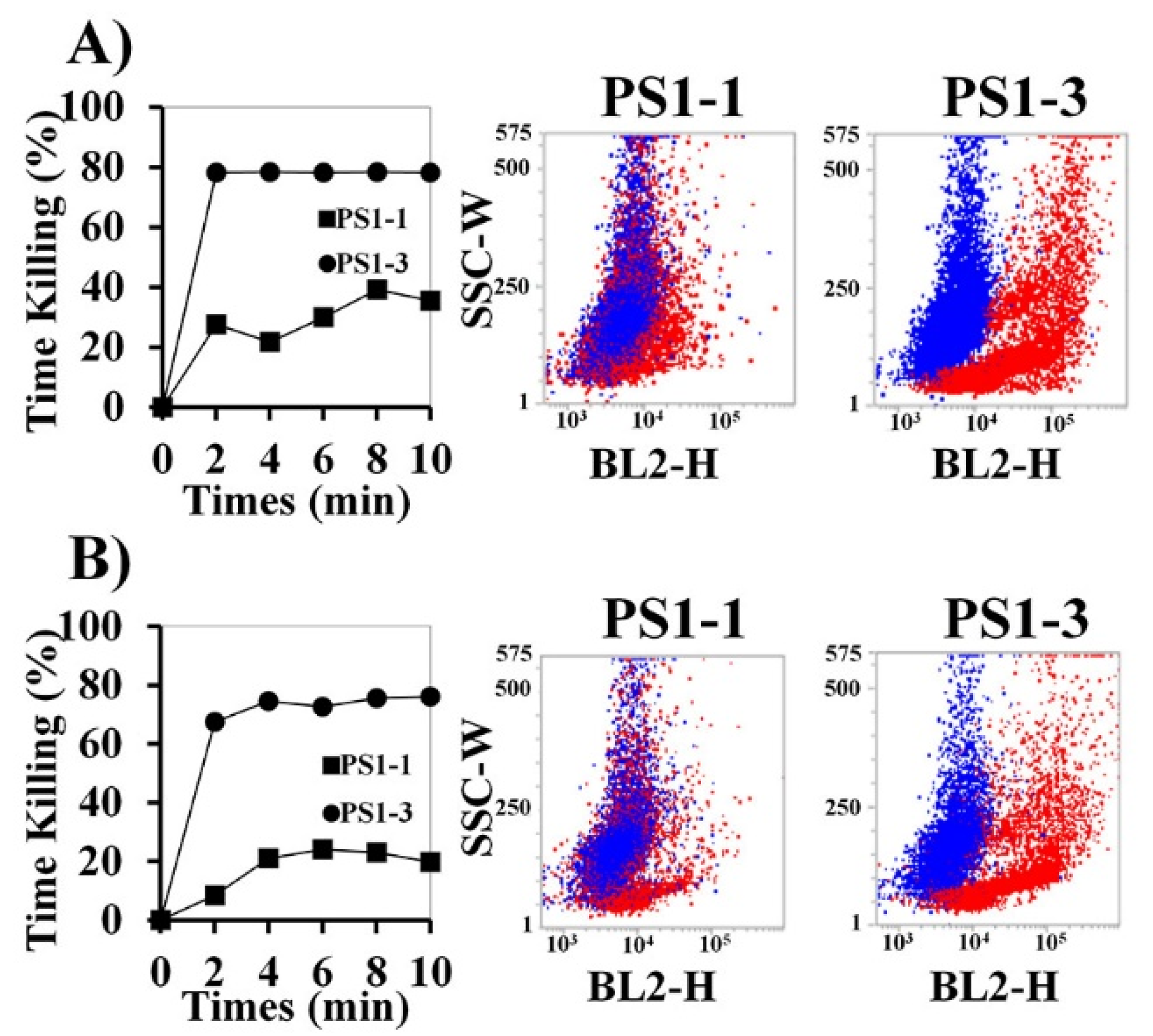
2.5. Assessment of Time Kill-Kinetic Activity Using Fluorescence-Assisted Cell Sorting (FACS) Analysis and Fluorescence Microscopy
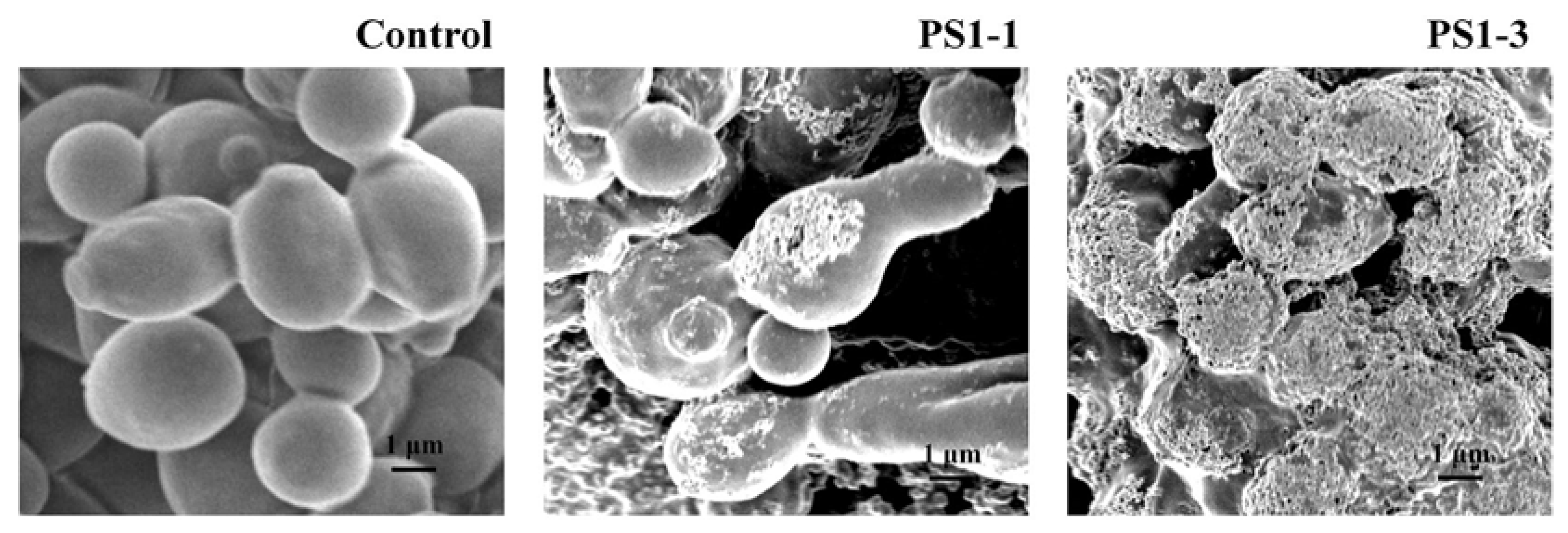
2.6. Analysis of Morphological Changes Using SEM
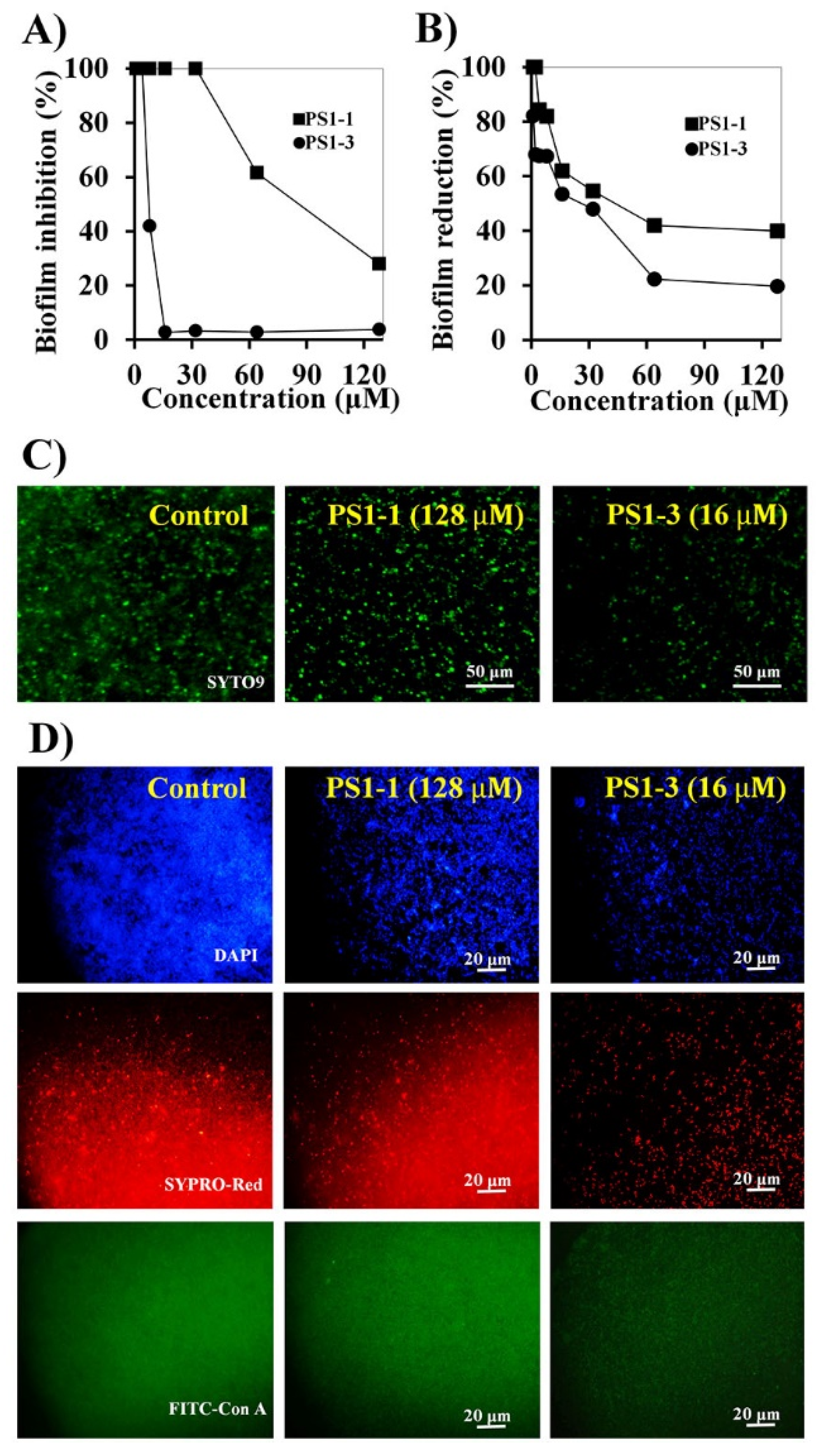
2.7. Biofilm Inhibition Assay
2.8. Biofilm Reduction Assay
2.9. CLSM Analysis
2.10. Biofilm Activity Assays on Coverslips
2.11. Biofilm Activity Assay on Soft CLs (SCLs)
3. Results
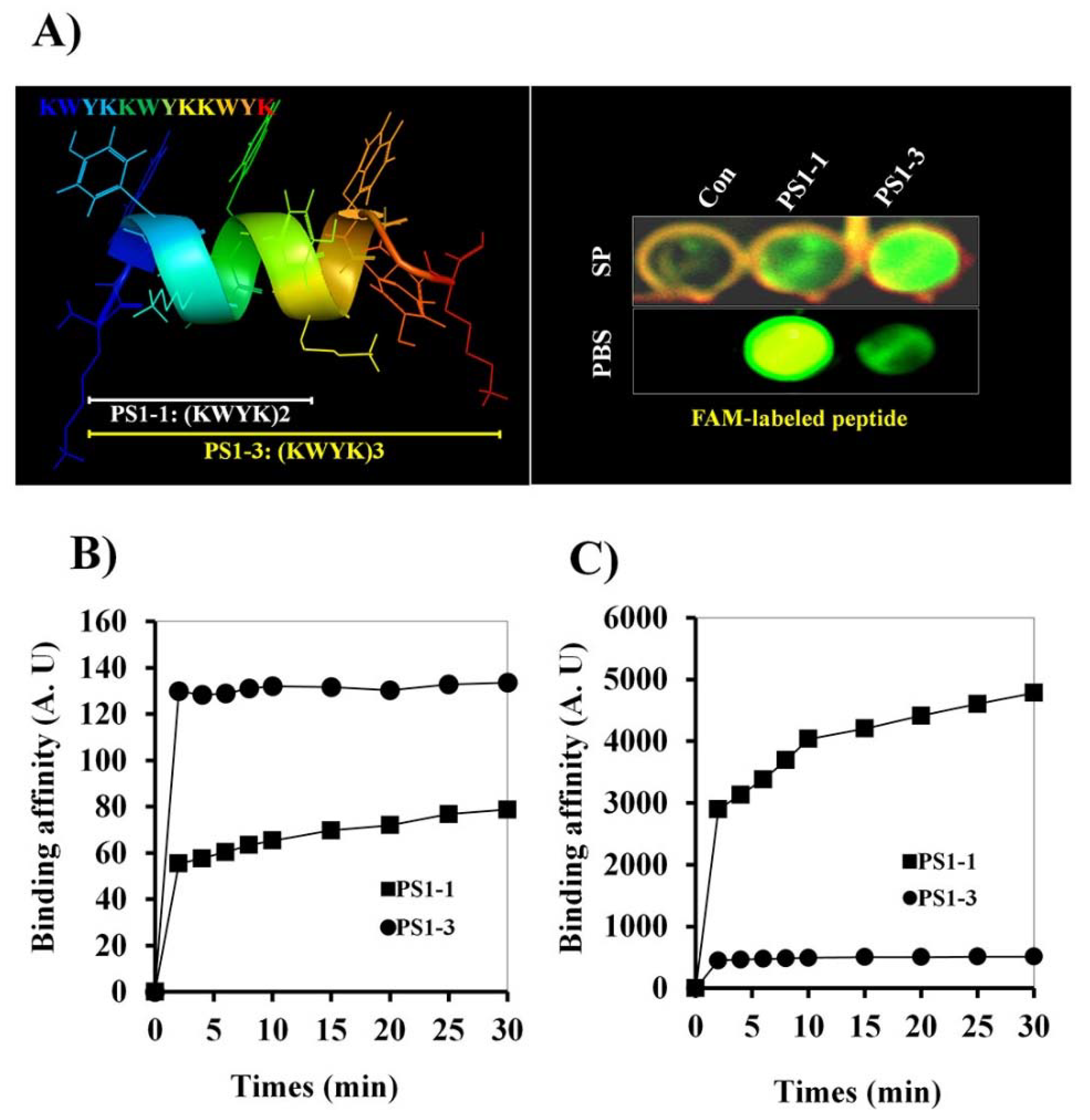
3.1. Candidacidal Activity of PS1 Analog Peptides
3.2. 5’-Fluorescein Phosphoramidite (FAM)-Labeled Peptide Binding Activity
3.3. Time Kill-Kinetic Activity
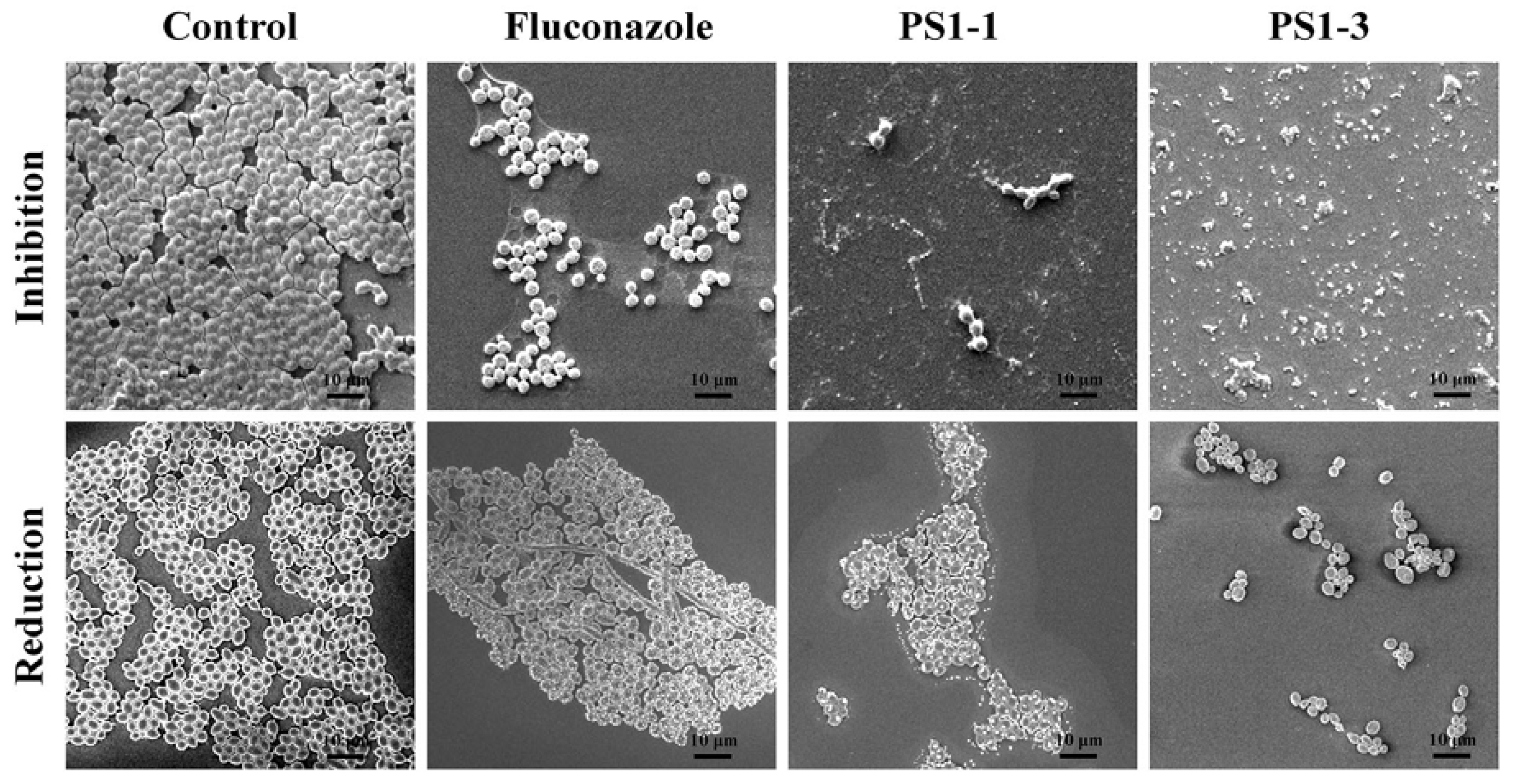
3.4. Analysis of Morphological Changes through Scanning Electron Microscopy (SEM)
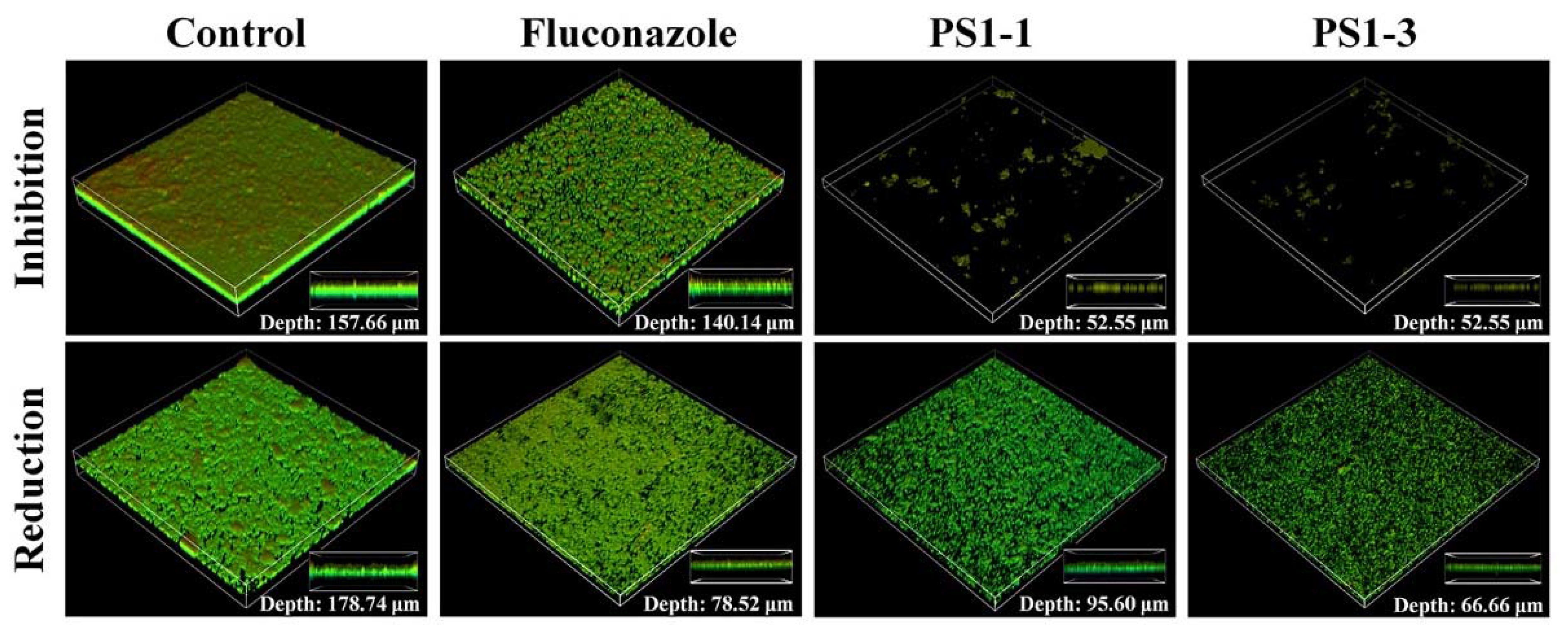
3.5. Antibiofilm Activity of PS1 Analog Peptides
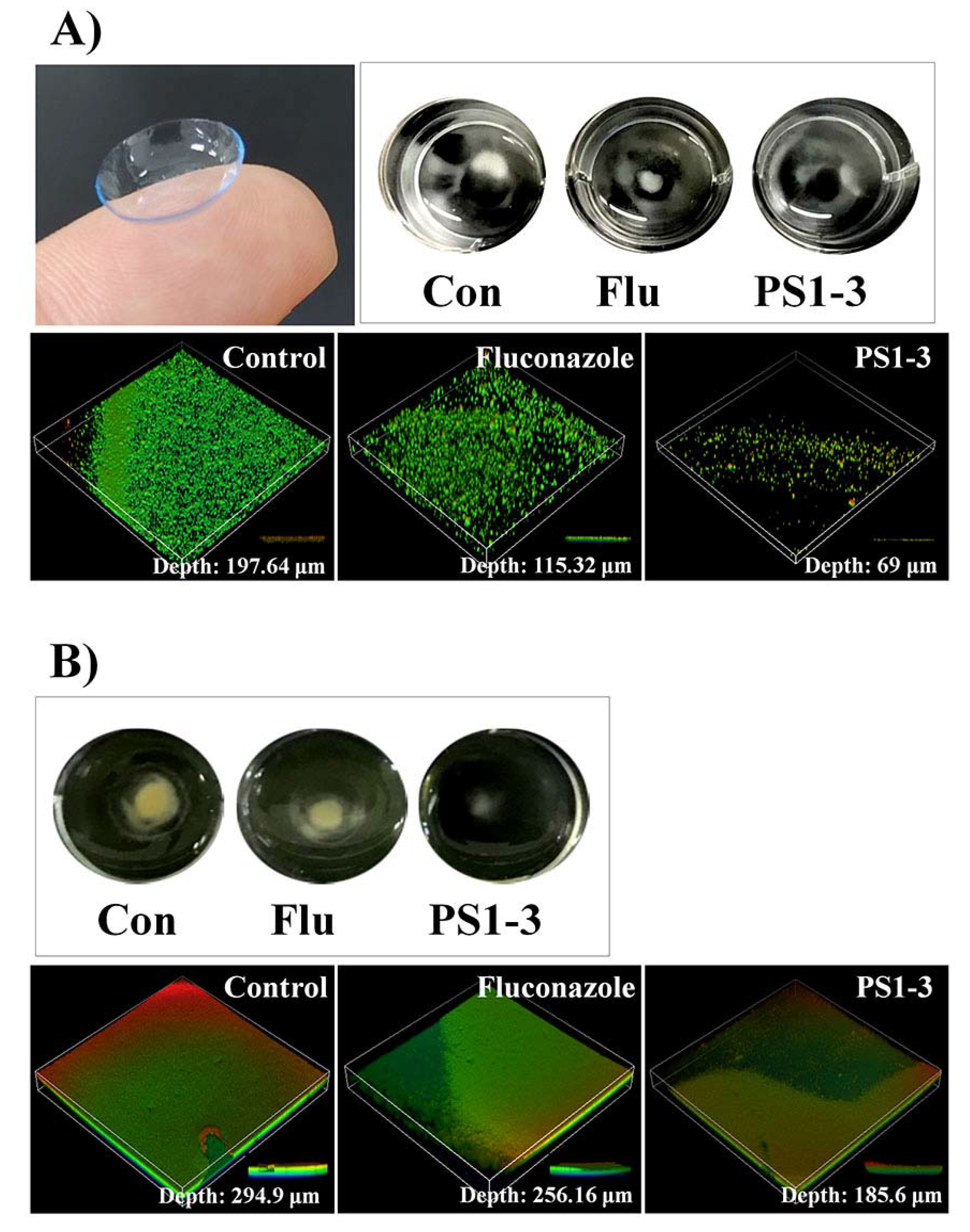
3.6. Ex Vivo Antibiofilm Activities of PS1-1 and PS1-3 Peptides
3.7. Antibiofilm Activity on the CLs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Barbosa, A.; Araújo, D.; Ribeiro, E.; Henriques, M.; Silva, S. Candida albicans Adaptation on Simulated Human Body Fluids under Different pH. Microorganisms 2020, 8, 511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Cheng, L.; Lei, Y.L.; Ren, B.; Zhou, X. The Interactions between Candida albicans and Mucosal Immunity. Front. Microbiol. 2021, 12, 652725. [Google Scholar] [CrossRef] [PubMed]
- Shahid, S.M.; Ahmed, S.N.; Khan, Y. Eye problems in contact lens wearers. BMJ 2019, 27, l6337. [Google Scholar] [CrossRef]
- Pucker, A.D.; Tichenor, A.A. A Review of Contact Lens Dropout. Clin. Optom. 2020, 25, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Boukari, M.; Zhioua-Braham, I.; Souissi, L.; Kaouel, H.; Errais, K.; Ammous, I.; Zhioua, R. Contact lens-related polymicrobial keratitis: Acanthamoeba spp. and Candida albicans. J. Fr. Ophtalmol. 2020, 43, e39–e40. [Google Scholar] [CrossRef] [PubMed]
- Buchele, M.L.C.; Wopereis, D.B.; Casara, F.; de Macedo, J.P.; Rott, M.B.; Monteiro, F.B.F.; Bazzo, M.L.; Dos Reis Spada, F.; Dos Santos, J.I.; Caumo, K.S. Contact lens-related polymicrobial keratitis: Acanthamoeba spp. genotype T4 and Candida albicans. Parasitol. Res. 2018, 117, 3431–3436. [Google Scholar] [CrossRef]
- Imamura, Y.; Chandra, J.; Mukherjee, P.K.; Lattif, A.A.; Szczotka-Flynn, L.B.; Pearlman, E.; Lass, J.H.; O’Donnell, K.; Ghannoum, M.A. Fusarium and Candida albicans biofilms on soft contact lenses: Model development, influence of lens type, and susceptibility to lens care solutions. Antimicrob. Agents Chemother. 2008, 52, 171–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kackar, S.; Suman, E.; Kotian, M.S. Bacterial and fungal biofilm formation on contact lenses and their susceptibility to lens care solutions. Indian J. Med. Microbiol. 2017, 35, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Park, S.C.; Jung, M.; Shin, M.K.; Kang, H.L.; Baik, S.C.; Cheong, G.W.; Jang, M.K.; Lee, W.K. Cell-selectivity of tryptophan and tyrosine in amphiphilic alpha-helical antimicrobial peptides against drug-resistant bacteria. Biochem. Biophys. Res. Commun. 2018, 28, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Kim, Y.M.; Lee, J.K.; Kim, N.H.; Kim, E.J.; Heo, H.; Lee, M.Y.; Lee, J.R.; Jang, M.K. Targeting and synergistic action of an antifungal peptide in an antibiotic drug-delivery system. J. Control. Release 2017, 28, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Park, Y. Anti-endotoxin mechanism of the KW4 peptide in inflammation in RAW 264.7 cells induced by LTA and drug-resistant Staphylococcus aureus 1630. Amino Acids 2018, 50, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Lee, M.Y.; Kim, J.Y.; Kim, H.; Jung, M.; Shin, M.K.; Lee, W.K.; Cheong, G.W.; Lee, J.R.; Jang, M.K. Anti-Biofilm Effects of Synthetic Antimicrobial Peptides Against Drug-Resistant Pseudomonas aeruginosa and Staphylococcus aureus Planktonic Cells and Biofilm. Molecules 2019, 24, 4560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand, M.L. Bacterial and Fungal Endophthalmitis. Clin. Microbiol. Rev. 2017, 30, 597–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Puumala, E.; Robbins, N.; Cowen, L.E. Antifungal Drug Resistance: Molecular Mechanisms in Candida albicans and Beyond. Chem. Rev. 2021, 24, 3390–3411. [Google Scholar] [CrossRef] [PubMed]
- Kabir, M.A.; Hussain, M.A.; Ahmad, Z. Candida albicans: A Model Organism for Studying Fungal Pathogens. ISRN Microbiol. 2012, 29, 538694. [Google Scholar] [CrossRef] [Green Version]
- Nobile, C.J.; Johnson, A.D. Candida albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef] [Green Version]
- Dadar, M.; Tiwari, R.; Karthik, K.; Chakraborty, S.; Shahali, Y.; Dhama, K. Candida albicans—Biology, molecular characterization, pathogenicity, and advances in diagnosis and control—An update. Microb. Pathog. 2018, 117, 128–138. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, Y.G.; Gupta, V.K.; Manoharan, R.K.; Lee, J. Suppression of Fluconazole Resistant Candida albicans Biofilm Formation and Filamentation by Methylindole Derivatives. Front. Microbiol. 2018, 6, 2641. [Google Scholar] [CrossRef]
- Suchodolski, J.; Krasowska, A. Fructose Induces Fluconazole Resistance in Candida albicans through Activation of Mdr1 and Cdr1 Transporters. Int. J. Mol. Sci. 2021, 22, 2127. [Google Scholar] [CrossRef]
- Lu, H.; Shrivastava, M.; Whiteway, M.; Jiang, Y. Candida albicans targets that potentially synergize with Fluconazole. Crit. Rev. Microbiol. 2021, 47, 323–337. [Google Scholar] [CrossRef]
- Paulone, S.; Ardizzoni, A.; Tavanti, A.; Piccinelli, S.; Rizzato, C.; Lupetti, A.; Colombari, B.; Pericolini, E.; Polonelli, L.; Magliani, W.; et al. The synthetic killer peptide KP impairs Candida albicans biofilm in vitro. PLoS ONE 2017, 13, e0181278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakharova, A.A.; Efimova, S.S.; Malev, V.V.; Ostroumova, O.S. Fengycin induces ion channels in lipid bilayers mimicking target fungal cell membranes. Sci. Rep. 2019, 5, 16034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Struyfs, C.; Cammue, B.P.A.; Thevissen, K. Membrane-Interacting Antifungal Peptides. Front. Cell Dev. Biol. 2021, 12, 649875. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Sun, J.; Xia, S.; Tian, X.; Cheserek, M.J.; Le, G. Mechanism of antifungal activity of antimicrobial peptide APP, a cell-penetrating peptide derivative, against Candida albicans: Intracellular DNA binding and cell cycle arrest. Appl. Microbiol. Biotechnol. 2016, 100, 3245–3253. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Raman, N.; Ortiz-Bermúdez, P.; Lynn, D.M.; Palecek, S.P. 14-Helicalb-Peptides Elicit Toxicity against C. albicans by Forming Pores in the Cell Membrane and Subsequently Disrupting Intracellular Organelles. Cell. Chem. Biol. 2019, 21, 289–299.e4. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | Molecular Weight (MW) | MIC/MFC (μM) | |||
|---|---|---|---|---|---|---|
| C. a | C. a (14007) | |||||
| SP | PBS | SP | PBS | |||
| PS1-1 | (KWYK)2 | 1230 Da | 32/32 | 64/64 | 32/32 | 64/128 |
| PS1-3 | (KWYK)4 | 2441 Da | 8/16 | 8/16 | 8/16 | 8/16 |
| Fluconazole | - | - | >128 | >128 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-K.; Park, S.; Kim, Y.-M.; Guk, T.; Lee, M.-Y.; Park, S.-C.; Lee, J.R.; Jang, M.-K. Candidacidal and Antibiofilm Activity of PS1-3 Peptide against Drug-Resistant Candida albicans on Contact Lenses. Pharmaceutics 2022, 14, 1602. https://doi.org/10.3390/pharmaceutics14081602
Lee J-K, Park S, Kim Y-M, Guk T, Lee M-Y, Park S-C, Lee JR, Jang M-K. Candidacidal and Antibiofilm Activity of PS1-3 Peptide against Drug-Resistant Candida albicans on Contact Lenses. Pharmaceutics. 2022; 14(8):1602. https://doi.org/10.3390/pharmaceutics14081602
Chicago/Turabian StyleLee, Jong-Kook, Soyoung Park, Young-Min Kim, Taeuk Guk, Min-Young Lee, Seong-Cheol Park, Jung Ro Lee, and Mi-Kyeong Jang. 2022. "Candidacidal and Antibiofilm Activity of PS1-3 Peptide against Drug-Resistant Candida albicans on Contact Lenses" Pharmaceutics 14, no. 8: 1602. https://doi.org/10.3390/pharmaceutics14081602
APA StyleLee, J.-K., Park, S., Kim, Y.-M., Guk, T., Lee, M.-Y., Park, S.-C., Lee, J. R., & Jang, M.-K. (2022). Candidacidal and Antibiofilm Activity of PS1-3 Peptide against Drug-Resistant Candida albicans on Contact Lenses. Pharmaceutics, 14(8), 1602. https://doi.org/10.3390/pharmaceutics14081602