
Adiponectin System (Rescue Hormone): The Missing Link between Metabolic and Cardiovascular Diseases


Abstract
1. Introduction
2. Physiology and Pathophysiology of Adiponectin on the Cardiovascular System
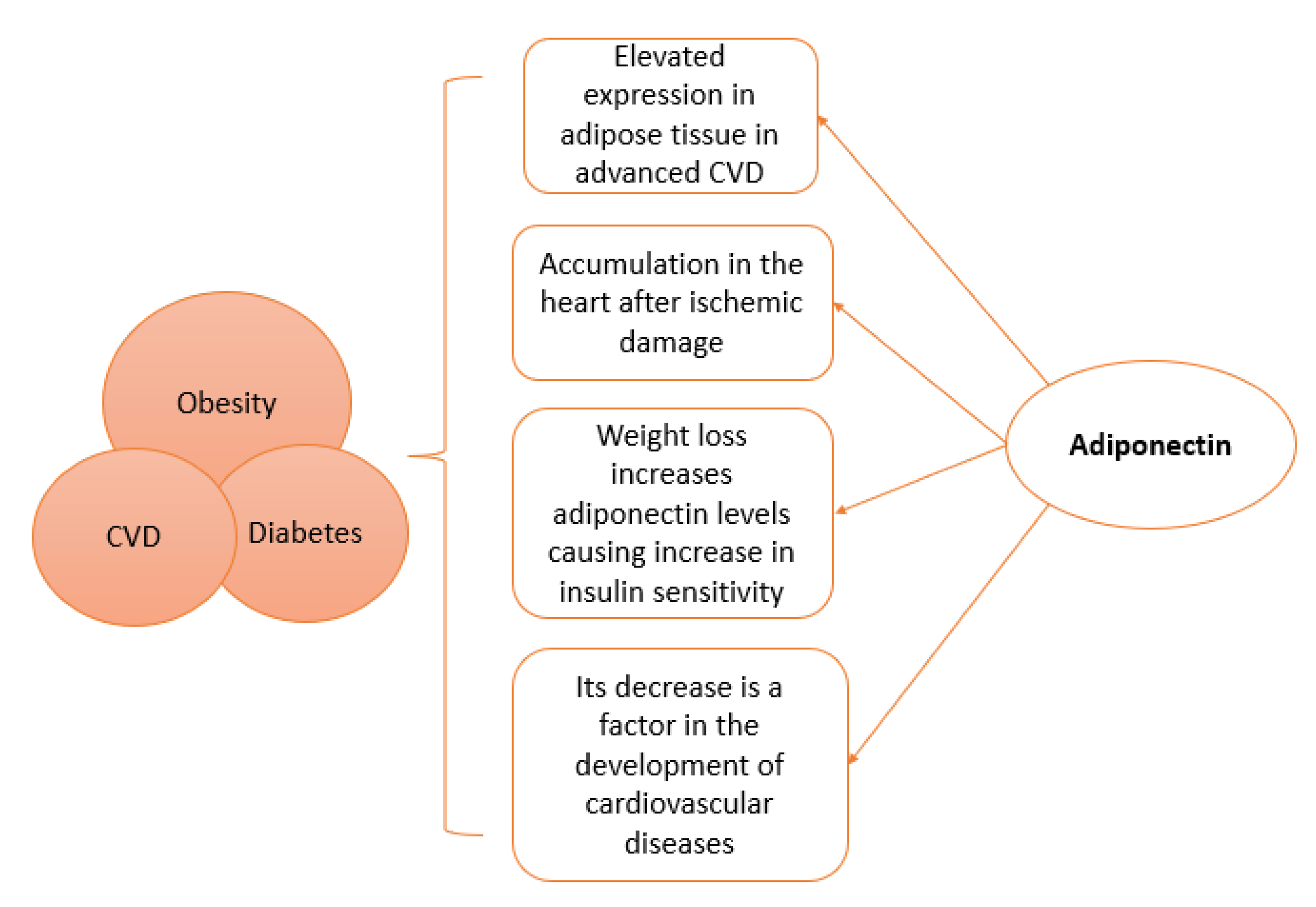
3. Paradoxal Effect of Adiponectin
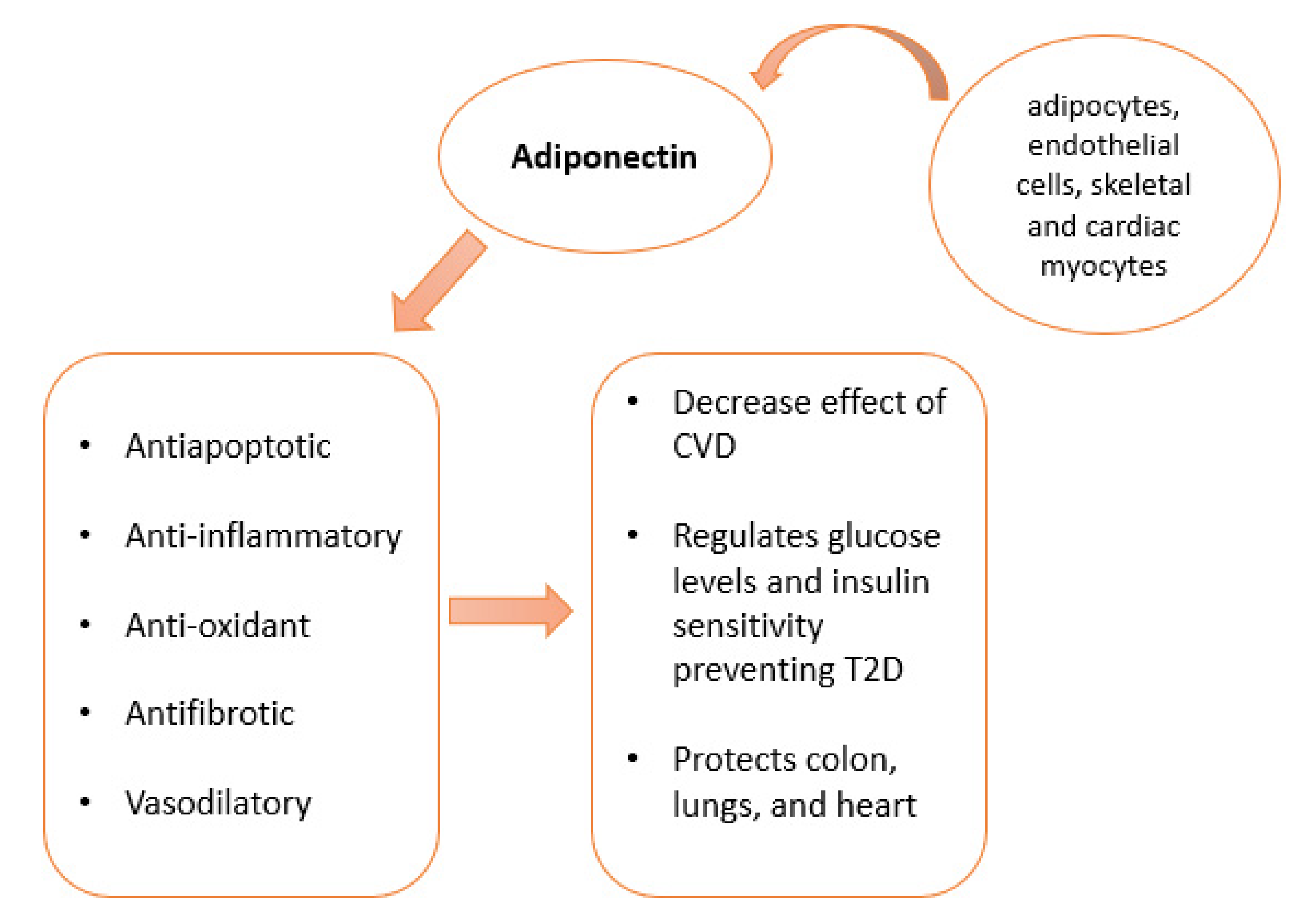
4. Adiponectin Resources and Multiple Functions
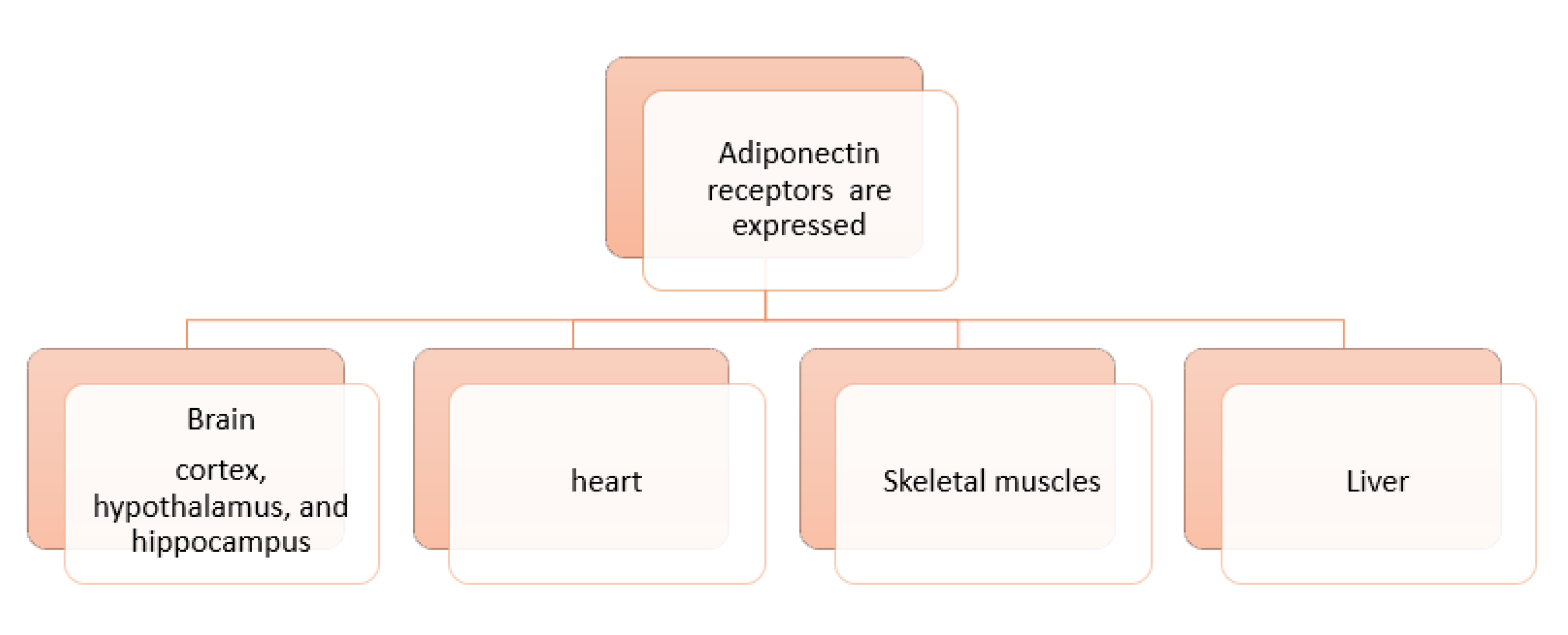
5. Adiponectin and Its Cellular Receptors, AdipoRs “Adiponectin System”
6. Cellular Effects of Adiponectin on Neurophysiology of the Heart
7. Adiponectin Plays the Key Link between Metabolic and Cardiovascular Diseases in Adults
8. Adiponectin Is a Link between Risk Factors of Cardiovascular Diseases in Children and Adolescents
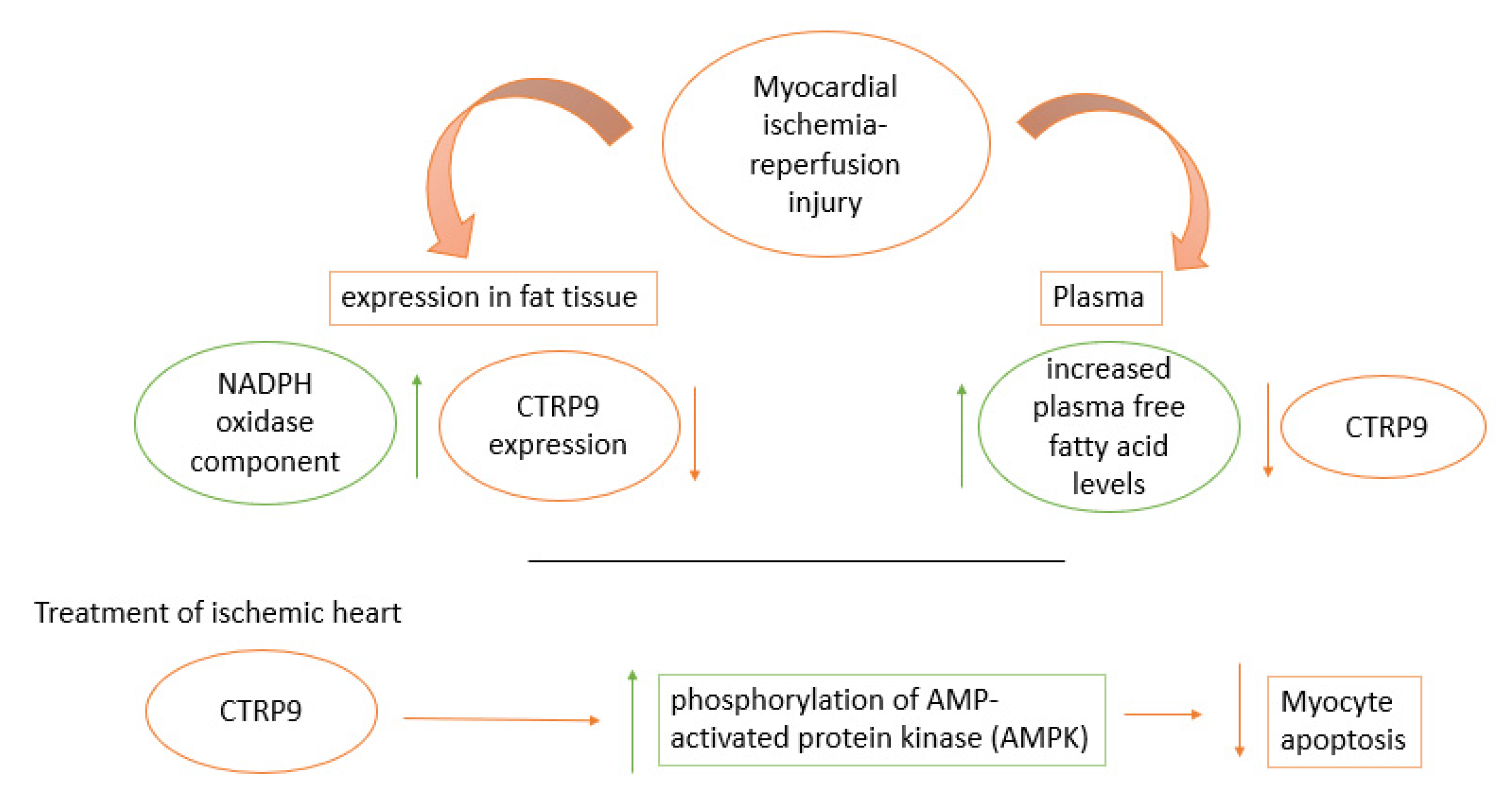
9. Significant Cellular Roles of Adiponectin as a Rescue Hormone In Vivo (Experimental Animals) and In Vitro Studies
10. Adiponectin Effects on Cardiac Cells and Remodeling
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hamdy, O.; Porramatikul, S.; Al-Ozairi, E. Metabolic obesity: The paradox between visceral and subcutaneous fat. Curr. Diabetes Rev. 2006, 2, 367–373. [Google Scholar] [PubMed]
- Fisman, E.Z.; Tenenbaum, A. Adiponectin: A manifold therapeutic target for metabolic syndrome, diabetes, and coronary disease? Cardiovasc. Diabetol. 2014, 13, 103. [Google Scholar] [CrossRef] [PubMed]
- Khoramipour, K.; Chamari, K.; Hekmatikar, A.; Ziyaiyan, A.; Taherkhani, S.; Elguindy, N.; Bragazzi, N. Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition. Nutrients 2021, 13, 1180. [Google Scholar] [CrossRef] [PubMed]
- Shibata, R.; Sato, K.; Kumada, M.; Izumiya, Y.; Sonoda, M.; Kihara, S.; Ouchi, N.; Walsh, K. Adiponectin accumulates in myocardial tissue that has been damaged by ischemia-reperfusion injury via leakage from the vascular compartment. Cardiovasc. Res. 2007, 74, 471–479. [Google Scholar] [CrossRef]
- Farkhondeh, T.; Llorens, S.; Pourbagher-Shahri, A.M.; Ashrafizadeh, M.; Talebi, M.; Shakibaei, M.; Samarghandian, S. An Overview of the Role of Adipokines in Cardiometabolic Diseases. Molecules 2020, 25, 5218. [Google Scholar] [CrossRef]
- Puchałowicz, K.; Kłoda, K.; Dziedziejko, V.; Rać, M.; Wojtarowicz, A.; Chlubek, D.; Safranow, K. Association of Adiponectin, Leptin and Resistin Plasma Concentrations with Echocardiographic Parameters in Patients with Coronary Artery Disease. Diagnostics 2021, 11, 1774. [Google Scholar] [CrossRef]
- Fedorova, T.; Semenenko, N.; Tazina, S.; Mamonov, A.; Sotnikova, T. Metabolic syndrome and chronic heart failure: A modern aspect of the problem. Bangladesh J. Med. Sci. 2022, 21, 105–113. [Google Scholar] [CrossRef]
- Tosun, Ş.A.; Gurbuz, T.; Cebi, A.; Tosun, A.; Gokmen, O.; Usta, M. Association of increased levels of omentin-1 and carotid intima-media thickness with early signs of cardiovascular risk in patients with polycystic ovary syndrome: A prospective case control study. J. Obstet. Gynaecol. Res. 2021, 48, 169–177. [Google Scholar] [CrossRef]
- Song, Y.; Song, F.; Wu, C.; Hong, Y.-X.; Li, G. The roles of epicardial adipose tissue in heart failure. Heart Fail. Rev. 2020, 27, 369–377. [Google Scholar] [CrossRef]
- Ruyvaran, M.; Zamani, A.; Mohamadian, A.; Zarshenas, M.M.; Eftekhari, M.H.; Pourahmad, S.; Abarghooei, E.F.; Akbari, A.; Nimrouzi, M. Safflower (Carthamus tinctorius L.) oil could improve abdominal obesity, blood pressure, and insulin resistance in patients with metabolic syndrome: A randomized, double-blind, placebo-controlled clinical trial. J. Ethnopharmacol. 2021, 282, 114590. [Google Scholar] [CrossRef]
- Kumada, M.; Kihara, S.; Sumitsuji, S.; Kawamoto, T.; Matsumoto, S.; Ouchi, N.; Arita, Y.; Okamoto, Y.; Shimomura, I.; Hiraoka, H.; et al. Coronary artery disease. Association of hypoadiponectinemia with coronary artery disease in men. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Woodward, L.; Akoumianakis, I.; Antoniades, C. Unravelling the adiponectin paradox: Novel roles of adiponectin in the regulation of cardiovascular disease. J. Cereb. Blood Flow Metab. 2016, 174, 4007–4020. [Google Scholar] [CrossRef]
- Zhao, S.; Kusminski, C.M.; Scherer, P.E. Adiponectin, Leptin and Cardiovascular Disorders. Circ. Res. 2021, 128, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Mado, H.; Szczurek, W.; Gąsior, M.; Szyguła-Jurkiewicz, B. Adiponectin in heart failure. Futur. Cardiol. 2021, 17, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Diep Nguyen, T.M. Adiponectin: Role in physiology and pathophysiology. Int. J. Prev. Med. 2020, 11, 136. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Tian, M.; Zhou, Y. The relationship between insulin resistance, adiponectin and C-reactive protein and vascular endothelial injury in diabetic patients with coronary heart disease. Exp. Ther. Med. 2018, 16, 2022–2026. [Google Scholar] [CrossRef] [PubMed]
- Honda, M.; Tsuboi, A.; Minato-Inokawa, S.; Kitaoka, K.; Takeuchi, M.; Yano, M.; Kurata, M.; Wu, B.; Kazumi, T.; Fukuo, K. Association of family history of type 2 diabetes with blood pressure and resting heart rate in young normal weight Japanese women. Diabetol. Int. 2021, 13, 220–225. [Google Scholar] [CrossRef]
- Gar, C.; Thorand, B.; Herder, C.; Sujana, C.; Heier, M.; Meisinger, C.; Peters, A.; Koenig, W.; Rathmann, W.; Roden, M.; et al. Association of circulating MR-proADM with all-cause and cardiovascular mortality in the general population: Results from the KORA F4 cohort study. PLoS ONE 2022, 17, e0262330. [Google Scholar] [CrossRef]
- Kadoglou, N.P.E.; Korakas, E.; Karkos, C.; Maratou, E.; Kanonidis, I.; Plotas, P.; Papanas, N.; Moutsatsou, P.; Ikonomidis, I.; Lambadiari, V. The prognostic role of RBP-4 and adiponectin in patients with peripheral arterial disease undergoing lower limb endovascular revascularization. Cardiovasc. Diabetol. 2021, 20, 221. [Google Scholar] [CrossRef]
- Hao, G.; Li, W.; Guo, R.; Yang, J.-G.; Wang, Y.; Tian, Y.; Liu, M.-Y.; Peng, Y.-G.; Wang, Z.-W. Serum total adiponectin level and the risk of cardiovascular disease in general population: A meta-analysis of 17 prospective studies. Atherosclerosis 2013, 228, 29–35. [Google Scholar] [CrossRef]
- Lawlor, D.A.; Smith, G.D.; Ebrahim, S.; Thompson, C.; Sattar, N. Plasma Adiponectin Levels Are Associated with Insulin Resistance, But Do Not Predict Future Risk of Coronary Heart Disease in Women. J. Clin. Endocrinol. Metab. 2005, 90, 5677–5683. [Google Scholar] [CrossRef] [PubMed]
- Menzaghi, C.; Trischitta, V. The Adiponectin Paradox for All-Cause and Cardiovascular Mortality. Diabetes 2017, 67, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Rojas, E.; Rodríguez-Molina, D.; Bolli, P.; Israili, Z.H.; Faria, J.; Fidilio, E.; Bermúdez, V.; Velasco, M. The Role of Adiponectin in Endothelial Dysfunction and Hypertension. Curr. Hypertens. Rep. 2014, 16, 463. [Google Scholar] [CrossRef] [PubMed]
- Kalaycıoğlu, E.; Çetin, M.; Özyıldız, A.G.; Kırış, T.; Turan, T. Is Adiponectin Elevation Associated with Left Atrial Remodeling and Impaired Mechanical Functions? (a Speckle Tracking Study). Kardiologiia 2021, 61, 65–70. [Google Scholar] [CrossRef]
- Semple, R.K.; Halberg, N.H.; Burling, K.; Soos, M.A.; Schraw, T.; Luan, J.; Cochran, E.K.; Dunger, D.B.; Wareham, N.J.; Scherer, P.E.; et al. Paradoxical Elevation of High–Molecular Weight Adiponectin in Acquired Extreme Insulin Resistance Due to Insulin Receptor Antibodies. Diabetes 2007, 56, 1712–1717. [Google Scholar] [CrossRef][Green Version]
- Ouchi, N.; Walsh, K. Adiponectin as an anti-inflammatory factor. Clin. Chim. Acta 2007, 380, 24–30. [Google Scholar] [CrossRef]
- Yoo, J.-K.; Hwang, M.-H.; Luttrell, M.J.; Kim, H.-K.; Meade, T.H.; English, M.; Segal, M.S.; Christou, D.D. Higher levels of adiponectin in vascular endothelial cells are associated with greater brachial artery flow-mediated dilation in older adults. Exp. Gerontol. 2015, 63, 1–7. [Google Scholar] [CrossRef]
- Parker-Duffen, J.L.; Walsh, K. Cardiometabolic effects of adiponectin. Best Pract. Res. Clin. Endocrinol. Metab. 2013, 28, 81–91. [Google Scholar] [CrossRef]
- Stojanovic, S.; Deljanin-Ilic, M.; Ilic, S.; Stefanovic, M.; Petrovic, D.; Petrovic, V.; Stojanovic, M. Adiponectin resistance parameter as a marker for high normal blood pres-sure and hypertension in patients with metabolic syndrome. Hippokratia 2020, 24, 3–7. [Google Scholar]
- Krause, M.P.; Milne, K.J.; Hawke, T.J. Adiponectin—Consideration for its Role in Skeletal Muscle Health. Int. J. Mol. Sci. 2019, 20, 1528. [Google Scholar] [CrossRef]
- Muldoon, T.G. Hormones and Their Receptors. Arch. Intern. Med. 1988, 148, 961. [Google Scholar] [CrossRef]
- Nussey, S.; Whitehead, S. Chapter 1, Principles of endocrinology. In Endocrinology: An Integrated Approach; BIOS Scientific Publishers: Oxford, UK, 2001. [Google Scholar]
- Vuppaladhadiam, L.; Ehsan, C.; Akkati, M.; Bhargava, A. Corticotropin-Releasing Factor Family: A Stress Hormone-Receptor System’s Emerging Role in Mediating Sex-Specific Signaling. Cells 2020, 9, 839. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chadwick, W.; Park, S.-S.; Zhou, Y.; Silver, N.; Martin, B.; Maudsley, S. Gonadotropin-Releasing Hormone Receptor System: Modulatory Role in Aging and Neurodegeneration. CNS Neurol. Disord.—Drug Targets 2010, 9, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Caselli, C. Role of adiponectin system in insulin resistance. Mol. Genet. Metab. 2014, 113, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Chabrolle, C.; Tosca, L.; Crochet, S.; Tesseraud, S.; Dupont, J. Expression of adiponectin and its receptors (AdipoR1 and AdipoR2) in chicken ovary: Potential role in ovarian steroidogenesis. Domest. Anim. Endocrinol. 2007, 33, 480–487. [Google Scholar] [CrossRef]
- Dall’Aglio, C.; Maranesi, M.; Di Loria, A.; Piantedosi, D.; Ciaramella, P.; Alterisio, M.; Lepri, E.; Mercati, F. Effects of Obesity on Adiponectin System Skin Expression in Dogs: A Comparative Study. Animals 2021, 11, 2308. [Google Scholar] [CrossRef]
- Dobrzyn, K.; Smolinska, N.; Szeszko, K.; Kiezun, M.; Maleszka, A.; Rytelewska, E.; Kaminski, T. Effect of progesterone on adiponectin system in the porcine uterus during early pregnancy1. J. Anim. Sci. 2017, 95, 338–352. [Google Scholar] [CrossRef] [PubMed]
- Dobrzyn, K.; Smolińska, N.; Kiezun, M.; Szeszko, K.; Rytelewska, E.; Kisielewska, K.; Gudelska, M.; Kaminski, T. Adiponectin: A New Regulator of Female Reproductive System. Int. J. Endocrinol. 2018, 2018, 7965071. [Google Scholar] [CrossRef]
- Han, Y.; Zhang, S.; Zhuang, H.; Fan, S.; Yang, J.; Zhao, L.; Bao, W.; Gao, F.; Zhang, H.; Yuan, Z.; et al. The role of the adiponectin system in acute fasting-impaired mouse ovaries. Reproduction 2019, 158, 429–440. [Google Scholar] [CrossRef]
- Kadivar, A.; Khoei, H.H.; Hassanpour, H.; Golestanfar, A.; Ghanaei, H. Correlation of Adiponectin mRNA Abundance and Its Receptors with Quantitative Parameters of Sperm Motility in Rams. Int. J. Fertil Steril. 2016, 10, 127–135. [Google Scholar] [CrossRef]
- Kim, M.J.; Maachi, M.; Debard, C.; Loizon, E.; Clément, K.; Bruckert, E.; Hainque, B.; Capeau, J.; Vidal, H.; Bastard, J.P. Increased adiponectin receptor-1 expression in adipose tissue of impaired glucose-tolerant obese subjects during weight loss. Eur. J. Endocrinol. 2006, 155, 161–165. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nigro, E.; Scudiero, O.; Monaco, M.L.; Palmieri, A.; Mazzarella, G.; Costagliola, C.; Bianco, A.; Daniele, A. New Insight into Adiponectin Role in Obesity and Obesity-Related Diseases. BioMed. Res. Int. 2014, 2014, 658913. [Google Scholar] [CrossRef] [PubMed]
- Paschke, L.; Zemleduch, T.; Rucinski, M.; Ziolkowska, A.; Szyszka, M.; Malendowicz, L.K. Adiponectin and adiponectin receptor system in the rat adrenal gland: Ontogenetic and physiologic regulation, and its involvement in regulating adrenocortical growth and steroidogenesis. Peptides 2010, 31, 1715–1724. [Google Scholar] [CrossRef] [PubMed]
- Skurk, C.; Wittchen, F.; Suckau, L.; Witt, H.; Noutsias, M.; Fechner, H.; Schultheiss, H.-P.; Poller, W. Description of a local cardiac adiponectin system and its deregulation in dilated cardiomyopathy. Eur. Heart J. 2008, 29, 1168–1180. [Google Scholar] [CrossRef]
- Valenzano, A.; Tartaglia, N.; Ambrosi, A.; Tafuri, D.; Monda, M.; Messina, A.; Sessa, F.; Campanozzi, A.; Monda, V.; Cibelli, G.; et al. The Metabolic Rearrangements of Bariatric Surgery: Focus on Orexin-A and the Adiponectin System. J. Clin. Med. 2020, 9, 3327. [Google Scholar] [CrossRef]
- Gardner, D.G.; Anderson, M.; Nissenson, R.A. Chapter 1. Hormones and Hormone Action. In Greenspan’s Basic & Clinical Endocrinology, 9th ed.; Gardner, D.G., Shoback, D., Eds.; McGraw Hill: New York, NY, USA, 2011. [Google Scholar]
- Wong, G.W.; Krawczyk, S.A.; Kitidis-Mitrokostas, C.; Ge, G.; Spooner, E.; Hug, C.; Gimeno, R.; Lodish, H.F. Identification and characterization of CTRP9, a novel secreted glycoprotein, from adipose tissue that reduces serum glucose in mice and forms heterotrimers with adiponectin. FASEB J. 2009, 23, 241–258. [Google Scholar] [CrossRef]
- Kambara, T.; Ohashi, K.; Shibata, R.; Ogura, Y.; Maruyama, S.; Enomoto, T.; Uemura, Y.; Shimizu, Y.; Yuasa, D.; Matsuo, K.; et al. CTRP9 Protein Protects against Myocardial Injury following Ischemia-Reperfusion through AMP-activated Protein Kinase (AMPK)-dependent Mechanism. J. Biol. Chem. 2012, 287, 18965–18973. [Google Scholar] [CrossRef]
- Mohammadi, S.; Arefhosseini, S.R.; Ebrahimi-Mamaeghani, M.; Fallah, P.; Bazi, Z. Adiponectin as a potential biomarker of vascular disease. Vasc. Health Risk Manag. 2015, 11, 55–70. [Google Scholar] [CrossRef]
- Van Raalte, D.H.; Li, M.; Pritchard, P.H.; Wasan, K.M. Peroxisome Proliferator-Activated Receptor (PPAR)-α: A Pharmacological Target with a Promising Future. Pharm. Res. 2004, 21, 1531–1538. [Google Scholar] [CrossRef]
- Tanabe, H.; Fujii, Y.; Okada-Iwabu, M.; Iwabu, M.; Kano, K.; Kawana, H.; Hato, M.; Nakamura, Y.; Terada, T.; Kimura-Someya, T.; et al. Human adiponectin receptor AdipoR1 assumes closed and open structures. Commun. Biol. 2020, 3, 446. [Google Scholar] [CrossRef]
- Rizzo, M.R.; Fasano, R.; Paolisso, G. Adiponectin and Cognitive Decline. Int. J. Mol. Sci. 2020, 21, 2010. [Google Scholar] [CrossRef] [PubMed]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, S.; Huang, H.; Zeng, A.; Han, Y.; Zeng, C.; Zheng, S.; Ren, H.; Wang, Y.; Huang, Y.; et al. GRK4-mediated adiponectin receptor-1 phosphorylative desensitization as a novel mechanism of reduced renal sodium excretion in hypertension. Clin. Sci. 2020, 134, 2453–2467. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Mah, M.; Ritchie, R.H.; De Blasio, M.J. The adiponectin signalling pathway—A therapeutic target for the cardiac complications of type 2 diabetes? Pharmacol. Ther. 2021, 232, 108008. [Google Scholar] [CrossRef]
- Okura, Y.; Imao, T.; Murashima, S.; Shibata, H.; Kamikavwa, A.; Okamatsu-Ogura, Y.; Saito, M.; Kimura, K. Interaction of Nerve Growth Factor β with Adiponectin and SPARC Oppositely Modulates its Biological Activity. Int. J. Mol. Sci. 2019, 20, 1541. [Google Scholar] [CrossRef]
- Hansen, C.S.; Vistisen, D.; Jørgensen, M.E.; Witte, D.R.; Brunner, E.J.; Tabák, A.G.; Kivimäki, M.; Roden, M.; Malik, M.; Herder, C. Adiponectin, biomarkers of inflammation and changes in cardiac autonomic function: Whitehall II study. Cardiovasc. Diabetol. 2017, 16, 153. [Google Scholar] [CrossRef]
- Hotta, K.; Funahashi, T.; Arita, Y.; Takahashi, M.; Matsuda, M.; Okamoto, Y.; Iwahashi, H.; Kuriyama, H.; Ouchi, N.; Maeda, K.; et al. Plasma Concentrations of a Novel, Adipose-Specific Protein, Adiponectin, in Type 2 Diabetic Patients. Arter. Thromb. Vasc. Biol. 2000, 20, 1595–1599. [Google Scholar] [CrossRef]
- Tao, L.; Gao, E.; Jiao, X.; Yuan, Y.; Li, S.; Christopher, T.A.; Lopez, B.L.; Koch, W.; Chan, L.; Goldstein, B.J.; et al. Adiponectin Cardioprotection After Myocardial Ischemia/Reperfusion Involves the Reduction of Oxidative/Nitrative Stress. Circulation 2007, 115, 1408–1416. [Google Scholar] [CrossRef]
- Lee, S.; Kwak, H.-B. Role of adiponectin in metabolic and cardiovascular disease. J. Exerc. Rehabil. 2014, 10, 54–59. [Google Scholar] [CrossRef]
- Orlando, A.; Nava, E.; Giussani, M.; Genovesi, S. Adiponectin and Cardiovascular Risk. From Pathophysiology to Clinic: Focus on Children and Adolescents. Int. J. Mol. Sci. 2019, 20, 3228. [Google Scholar] [CrossRef]
- Ruan, H.; Dong, L.Q. Adiponectin signaling and function in insulin target tissues. J. Mol. Cell Biol. 2016, 8, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Judd, R.L. Adiponectin Regulation and Function. Compr. Physiol. 2018, 8, 1031–1063. [Google Scholar] [PubMed]
- Calle, M.; Fernandez, M. Inflammation and type 2 diabetes. Diabetes Metab. 2012, 38, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Deepa, S.; Dong, L.Q. APPL1: Role in adiponectin signaling and beyond. Am. J. Physiol. Metab. 2009, 296, E22–E36. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Luo, N.; Zhu, X.; Chung, B.-H.; Garvey, W.T.; Fu, Y. Adiponectin-AdipoR1/2-APPL1 signaling axis suppresses human foam cell formation: Differential ability of AdipoR1 and AdipoR2 to regulate inflammatory cytokine responses. Atherosclerosis 2012, 221, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Mayer, O.; Seidlerová, J.; Bruthans, J.; Gelžinský, J.; Rychecká, M.; Mateřánková, M.; Karnosová, P.; Wohlfahrt, P.; Cífková, R.; Filipovský, J. Is There Really an Association of High Circulating Adiponectin Concentration and Mortality or Morbidity Risk in Stable Coronary Artery Disease? Horm. Metab. Res. 2020, 52, 861–868. [Google Scholar] [CrossRef]
- Jang, A.Y.; Scherer, P.E.; Kim, J.Y.; Lim, S.; Koh, K.K. Adiponectin and cardiometabolic trait and mortality: Where do we go? Cardiovasc. Res. 2021, cvab199. [Google Scholar] [CrossRef]
- Nomura, H.; Arashi, H.; Yamaguchi, J.; Ogawa, H.; Hagiwara, N. Relation of Adiponectin to Cardiovascular Events and Mortality in Patients with Acute Coronary Syndrome. Am. J. Cardiol. 2020, 140, 7–12. [Google Scholar] [CrossRef]
- Christen, T.; de Mutsert, R.; Lamb, H.J.; van Dijk, K.W.; le Cessie, S.; Rosendaal, F.R.; Jukema, J.W.; Trompet, S. Mendelian randomization study of the relation between adiponectin and heart function, unravelling the paradox. Peptides 2021, 146, 170664. [Google Scholar] [CrossRef]
- Cohen, K.E.; Katunaric, B.; SenthilKumar, G.; McIntosh, J.J.; Freed, J.K. Vascular endothelial adiponectin signaling across the life span. Am. J. Physiol. Circ. Physiol. 2022, 322, H57–H65. [Google Scholar] [CrossRef]
- Kita, S.; Shimomura, I. Stimulation of exosome biogenesis by adiponectin, a circulating factor secreted from adipocytes. J. Biochem. 2020, 169, 173–179. [Google Scholar] [CrossRef]
- Gan, L.; Liu, D.; Xie, D.; Lau, W.B.; Liu, J.; Christopher, T.A.; Lopez, B.; Liu, L.; Hu, H.; Yao, P.; et al. Ischemic Heart-Derived Small Extracellular Vesicles Impair Adipocyte Function. Circ. Res. 2022, 130, 48–66. [Google Scholar] [CrossRef] [PubMed]
- Da Eira, D.; Jani, S.; Sung, H.; Sweeney, G.; Ceddia, R.B. Effects of the adiponectin mimetic compound ALY688 on glucose and fat metabolism in visceral and subcutaneous rat adipocytes. Adipocyte 2020, 9, 550–562. [Google Scholar] [CrossRef]
- Sung, H.K.; Mitchell, P.L.; Gross, S.; Marette, A.; Sweeney, G. ALY688 elicits adiponectin-mimetic signaling and improves insulin action in skeletal muscle cells. Am. J. Physiol. Physiol. 2022, 322, C151–C163. [Google Scholar] [CrossRef] [PubMed]
- Winer, J.C.; Zern, T.L.; Taksali, S.E.; Dziura, J.; Cali, A.M.G.; Wollschlager, M.; Seyal, A.A.; Weiss, R.; Burgert, T.S.; Caprio, S. Adiponectin in Childhood and Adolescent Obesity and Its Association with Inflammatory Markers and Components of the Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2006, 91, 4415–4423. [Google Scholar] [CrossRef] [PubMed]
- Punthakee, Z.; Delvin, E.E.; O’Loughlin, J.; Paradis, G.; Levy, E.; Platt, R.W.; Lambert, M. Adiponectin, Adiposity, and Insulin Resistance in Children and Adolescents. J. Clin. Endocrinol. Metab. 2006, 91, 2119–2125. [Google Scholar] [CrossRef][Green Version]
- Ohashi, K.; Kihara, S.; Ouchi, N.; Kumada, M.; Fujita, K.; Hiuge, A.; Hibuse, T.; Ryo, M.; Nishizawa, H.; Maeda, N.; et al. Adiponectin Replenishment Ameliorates Obesity-Related Hypertension. Hypertension 2006, 47, 1108–1116. [Google Scholar] [CrossRef]
- Chow, W.S.; Cheung, B.M.; Tso, A.W.; Xu, A.; Wat, N.M.; Fong, C.H.; Ong, L.H.; Tam, S.; Tan, K.C.; Janus, E.D.; et al. Hypoadiponectinemia as a predictor for the development of hypertension: A 5-year prospective study. Hypertension 2007, 49, 1455–1461. [Google Scholar] [CrossRef]
- Iwashima, Y.; Katsuya, T.; Ishikawa, K.; Ouchi, N.; Ohishi, M.; Sugimoto, K.; Fu, Y.; Motone, M.; Yamamoto, K.; Matsuo, A.; et al. Hypoadiponectinemia Is an Independent Risk Factor for Hypertension. Hypertension 2004, 43, 1318–1323. [Google Scholar] [CrossRef]
- Guo, Z.; Xia, Z.; Yuen, V.G.; McNeill, J.H. Cardiac expression of adiponectin and its receptors in streptozotocin-induced diabetic rats. Metabolism 2007, 56, 1363–1371. [Google Scholar] [CrossRef]
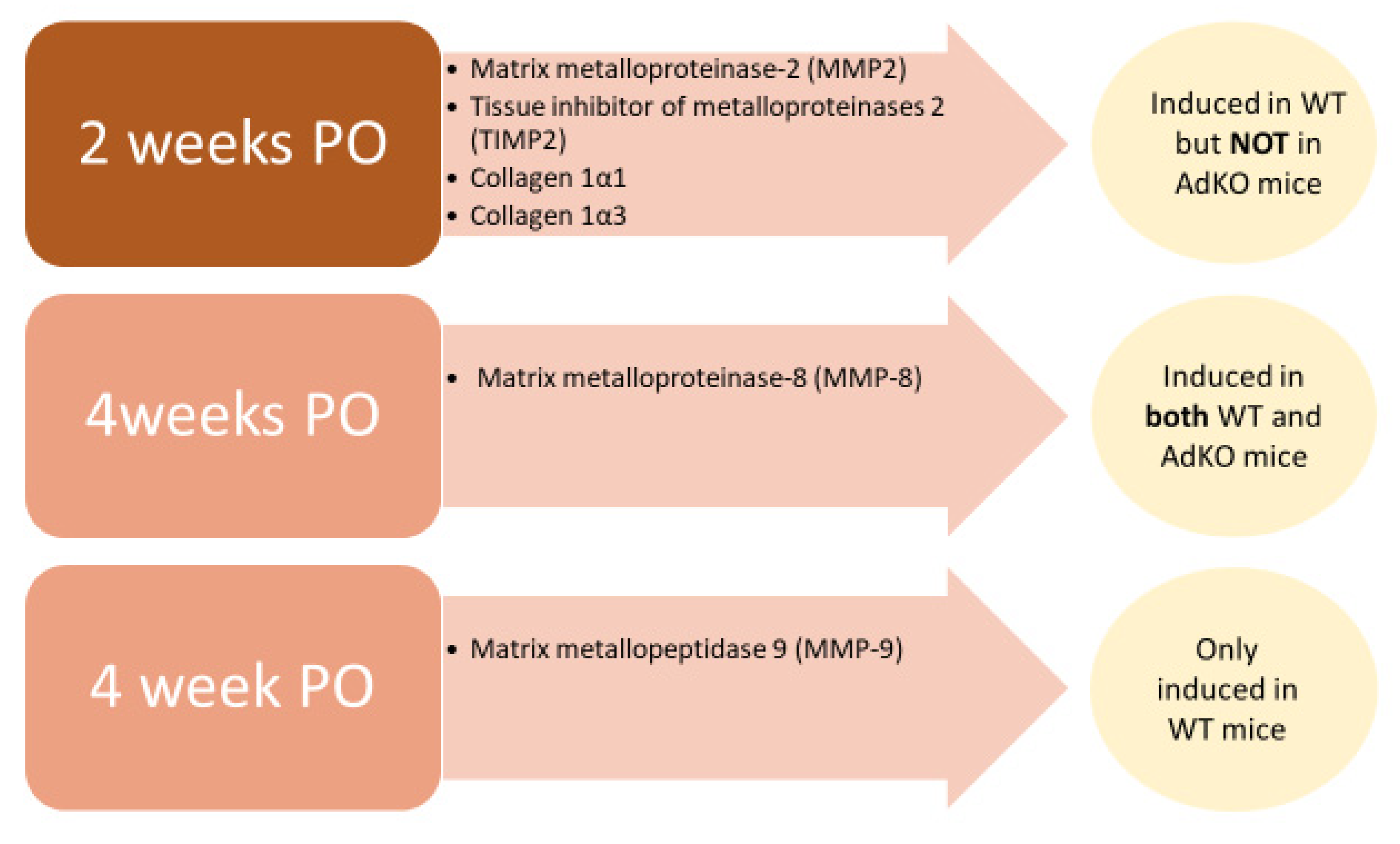
- Dadson, K.; Turdi, S.; Boo, S.; Hinz, B.; Sweeney, G. Temporal and Molecular Analyses of Cardiac Extracellular Matrix Remodeling following Pressure Overload in Adiponectin Deficient Mice. PLoS ONE 2015, 10, e0121049. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Hu, X.; Ge, P.-B.; Chen, Y.; Wu, S.; Zhang, X.-W. CTRP1 Aggravates Cardiac Dysfunction Post Myocardial Infarction by Modulating TLR4 in Macrophages. Front. Immunol. 2021, 12, 635267. [Google Scholar] [CrossRef] [PubMed]
- Jenke, A.; Yazdanyar, M.; Miyahara, S.; Chekhoeva, A.; Immohr, M.B.; Kistner, J.; Boeken, U.; Lichtenberg, A.; Akhyari, P. AdipoRon Attenuates Inflammation and Impairment of Cardiac Function Associated with Cardiopulmonary Bypass–Induced Systemic Inflammatory Response Syndrome. J. Am. Heart Assoc. 2021, 10, e018097. [Google Scholar] [CrossRef] [PubMed]
- Meng, K.; Cai, H.; Cai, S.; Hong, Y.; Zhang, X. Adiponectin Modified BMSCs Alleviate Heart Fibrosis via Inhibition TGF-beta1/Smad in Diabetic Rats. Front. Cell Dev. Biol. 2021, 9, 644160. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, T.A.; Ouchi, N.; Shibata, R.; Walsh, K. Adiponectin actions in the cardiovascular system. Cardiovasc. Res. 2007, 74, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Schram, K.; Sweeney, G. Implications of Myocardial Matrix Remodeling by Adipokines in Obesity-Related Heart Failure. Trends Cardiovasc. Med. 2008, 18, 199–205. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, K.M.; Chess, D.J.; Khairallah, R.J.; Rastogi, S.; Hecker, P.A.; Sabbah, H.N.; Walsh, K.; Stanley, W.C. Effects of adiponectin deficiency on structural and metabolic remodeling in mice subjected to pressure overload. Am. J. Physiol. Circ. Physiol. 2010, 298, H1639–H1645. [Google Scholar] [CrossRef]
- Guo, B.-Y.; Li, Y.-J.; Han, R.; Yang, S.-L.; Shi, Y.-H.; Han, D.-R.; Zhou, H.; Wang, M. Telmisartan attenuates isoproterenol-induced cardiac remodeling in rats via regulation of cardiac adiponectin expression. Acta Pharmacol. Sin. 2011, 32, 449–455. [Google Scholar] [CrossRef]
- Kamareddine, L.; Ghantous, C.M.; Allouch, S.; Al-Ashmar, S.A.; Anlar, G.; Kannan, S.; Djouhri, L.; Korashy, H.M.; Agouni, A.; Zeidan, A. Between Inflammation and Autophagy: The Role of Leptin-Adiponectin Axis in Cardiac Remodeling. J. Inflamm. Res. 2021, 14, 5349–5365. [Google Scholar] [CrossRef]
- Segers, V.F.M.; De Keulenaer, G.W. Autocrine Signaling in Cardiac Remodeling: A Rich Source of Therapeutic Targets. J. Am. Heart Assoc. 2021, 10, e019169. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ALY688 Affect | Epid Adipocytes | Sc Ing Adipocytes |
|---|---|---|
| alter basal lipolysis | Was not affect | Was not affected |
| enhanced isoproterenol-induced lipolysis | affected | - |
| expression of AdipoR2 mRNA | significantly higher expression | Less expression |
| AKTSer473 and p38 mitogen-activated protein kinase (MAPK) phosphorylation | Was not affected | Was not affected |
| Basal and insulin-stimulated rates of glucose uptake | Was not affected | Was not affected |
| glucose incorporation into lipids | Was not affected | Was not affected |
| Research Domain | Adiponectin Effect on Different Health Conditions | Reference |
|---|---|---|
| Clinical research and experimental animals | Decrease concentrations of adiponectin are linked with a rise in
| [59] |
| Clinical research | Decrease of adiponectin concentration in patients with type2 DM was recorded | [60] |
| Clinical research | In apolipoprotein E deficiency, adiponectin was recognized to have direct effect decreasing inflammatory responses caused by mechanical injury in vascular endothelium and increasing endothelium protection through AMP kinase activation | [2] |
| Clinical research | Adiponectin concentration and the increase in BNP increased mortality in patients with stable CAD, whereas the increase in adiponectin was not related to CV death or stroke | [68] |
| Clinical research (children) |
| [77] |
| Clinical research (children) | The connection between obesity and parameters of insulin resistance is reduced in youth of higher adiponectin levels, which made adiponectin a potential mediation goal or risk biomarker | [78] |
| In Vitro studies | Adiponectin modified mesenchymal stem cells revealed protection on cardiac fibrosis so modulated mesenchymal stem cells may be a new therapeutic for the diabetic cardiomyopathy in the future | [86] |
| In Vitro studies | Mesenchymal stem cells have been used therapeutically in mice for pressure overload-led heart failure involved the existence of plasma adiponectin and T-cadherin expression | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aljafary, M.A.; Al-Suhaimi, E.A. Adiponectin System (Rescue Hormone): The Missing Link between Metabolic and Cardiovascular Diseases. Pharmaceutics 2022, 14, 1430. https://doi.org/10.3390/pharmaceutics14071430
Aljafary MA, Al-Suhaimi EA. Adiponectin System (Rescue Hormone): The Missing Link between Metabolic and Cardiovascular Diseases. Pharmaceutics. 2022; 14(7):1430. https://doi.org/10.3390/pharmaceutics14071430
Chicago/Turabian StyleAljafary, Meneerah Abdulrahman, and Ebtesam Abdullah Al-Suhaimi. 2022. "Adiponectin System (Rescue Hormone): The Missing Link between Metabolic and Cardiovascular Diseases" Pharmaceutics 14, no. 7: 1430. https://doi.org/10.3390/pharmaceutics14071430
APA StyleAljafary, M. A., & Al-Suhaimi, E. A. (2022). Adiponectin System (Rescue Hormone): The Missing Link between Metabolic and Cardiovascular Diseases. Pharmaceutics, 14(7), 1430. https://doi.org/10.3390/pharmaceutics14071430