
Effects of the Interleukin-6 Receptor Blocker Sarilumab on Metabolic Activity and Differentiation Capacity of Primary Human Osteoblasts

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Osteoblastic Cell Culture and Stimulation Experiments
2.2. Determination of Cell Morphology, Viability, and Osteoblastic Differentiation Capacity
2.3. Gene Expression Analysis
2.4. Transcriptome Analysis
2.5. Mineralization Assay
2.6. Graphical Illustration and Statistics
3. Results
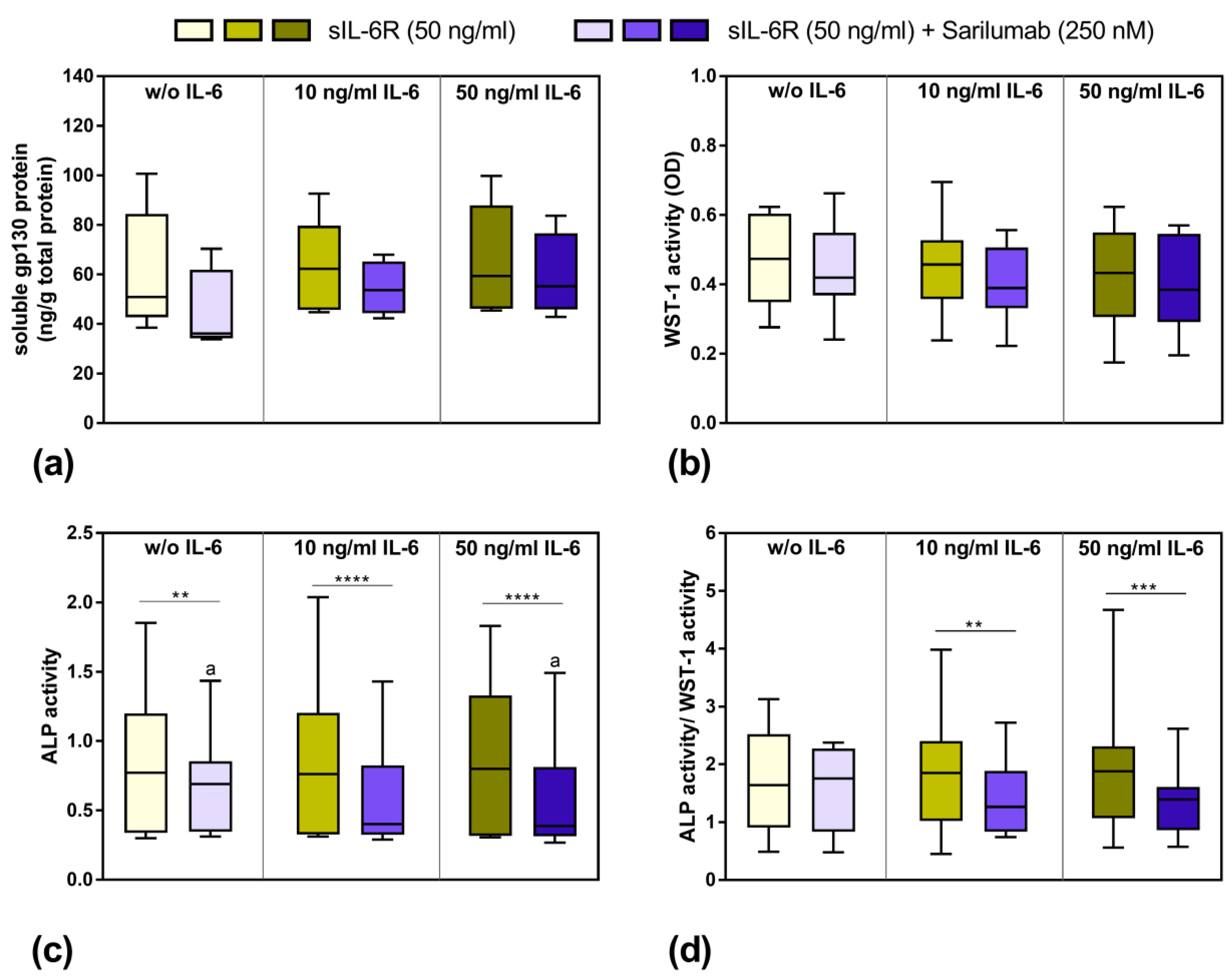
3.1. Influence of Exogenous IL-6 on the Secretion of sIL-6R and sgp130
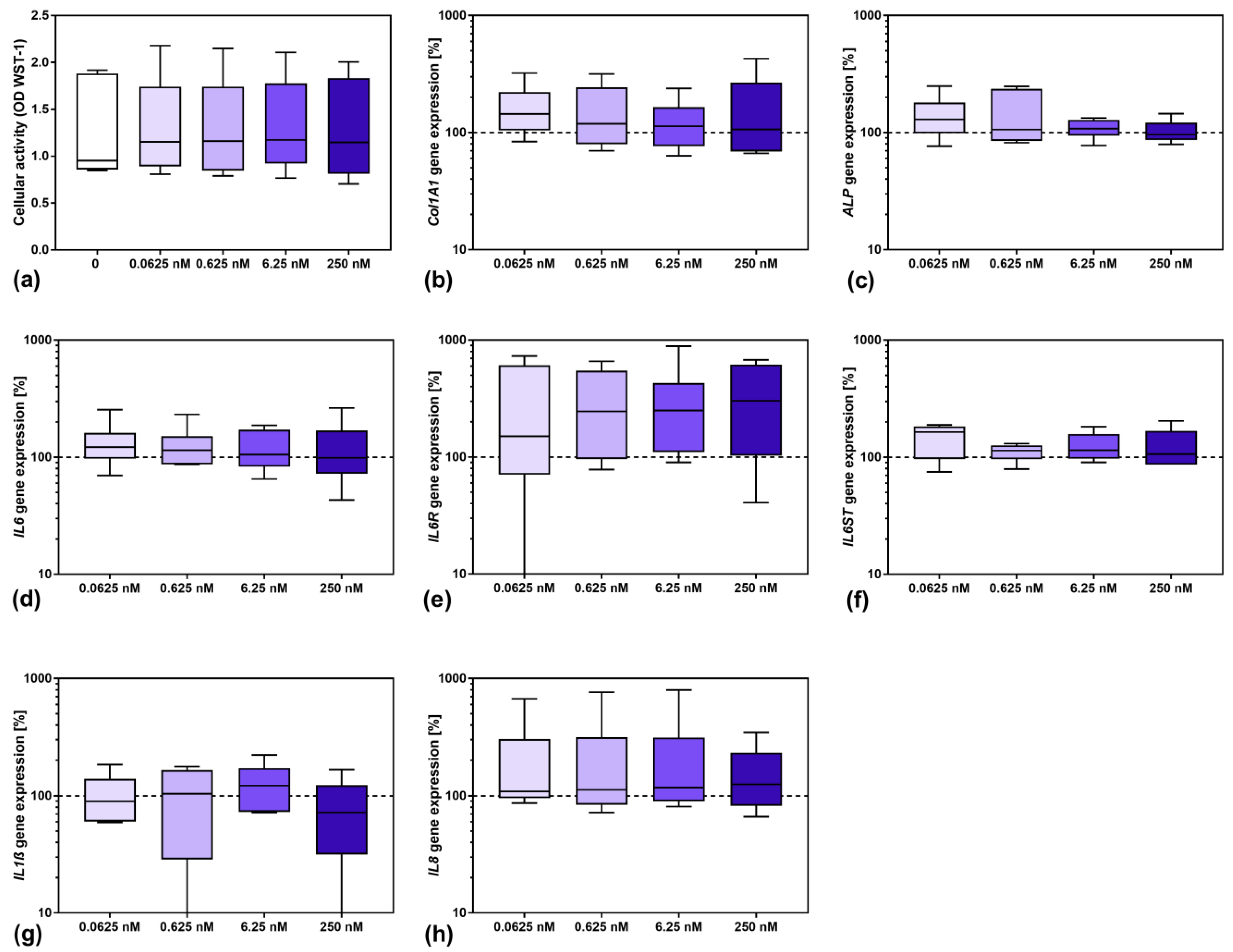
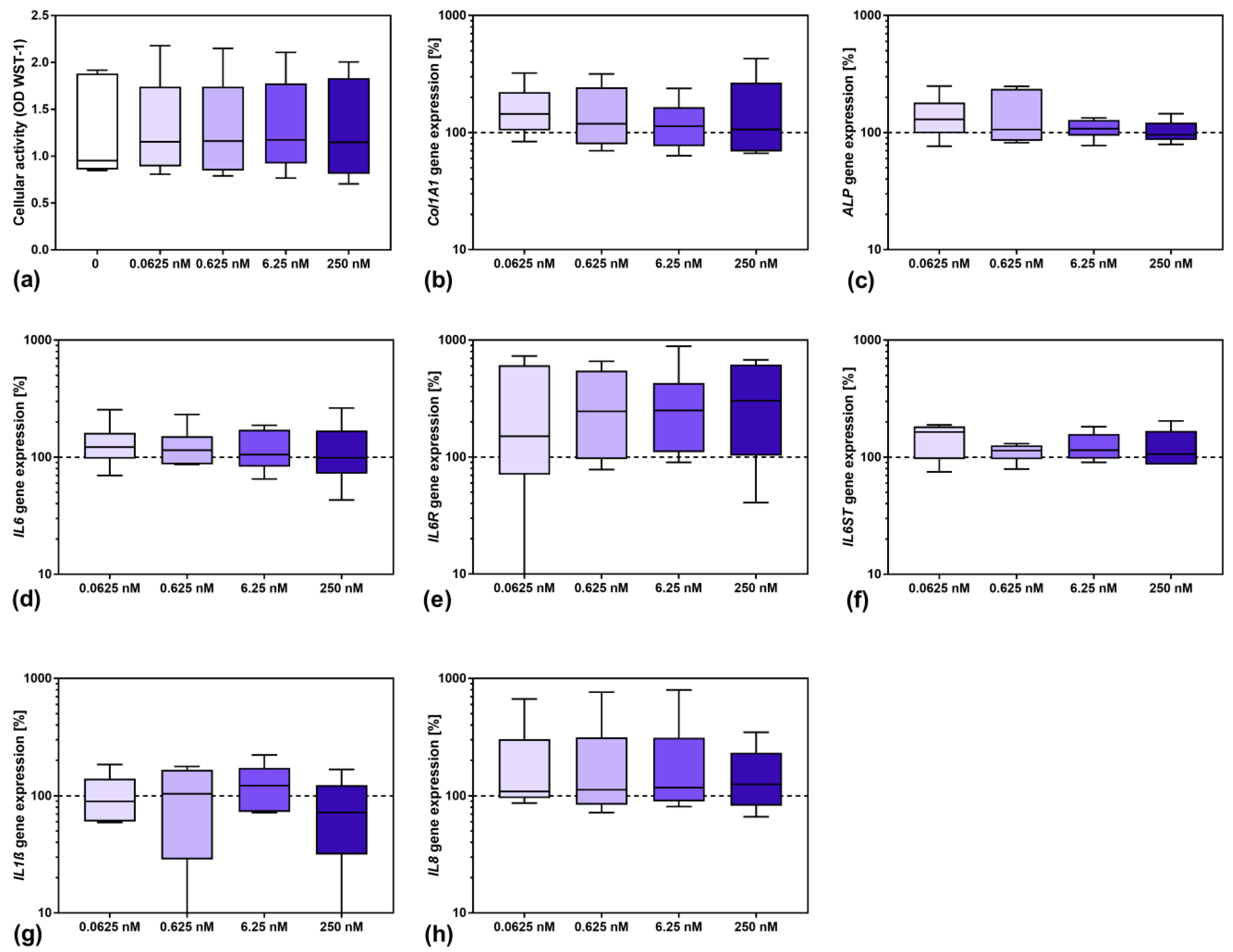
3.2. Influence of Sarilumab on Osteoblastic Viability and Gene Expression
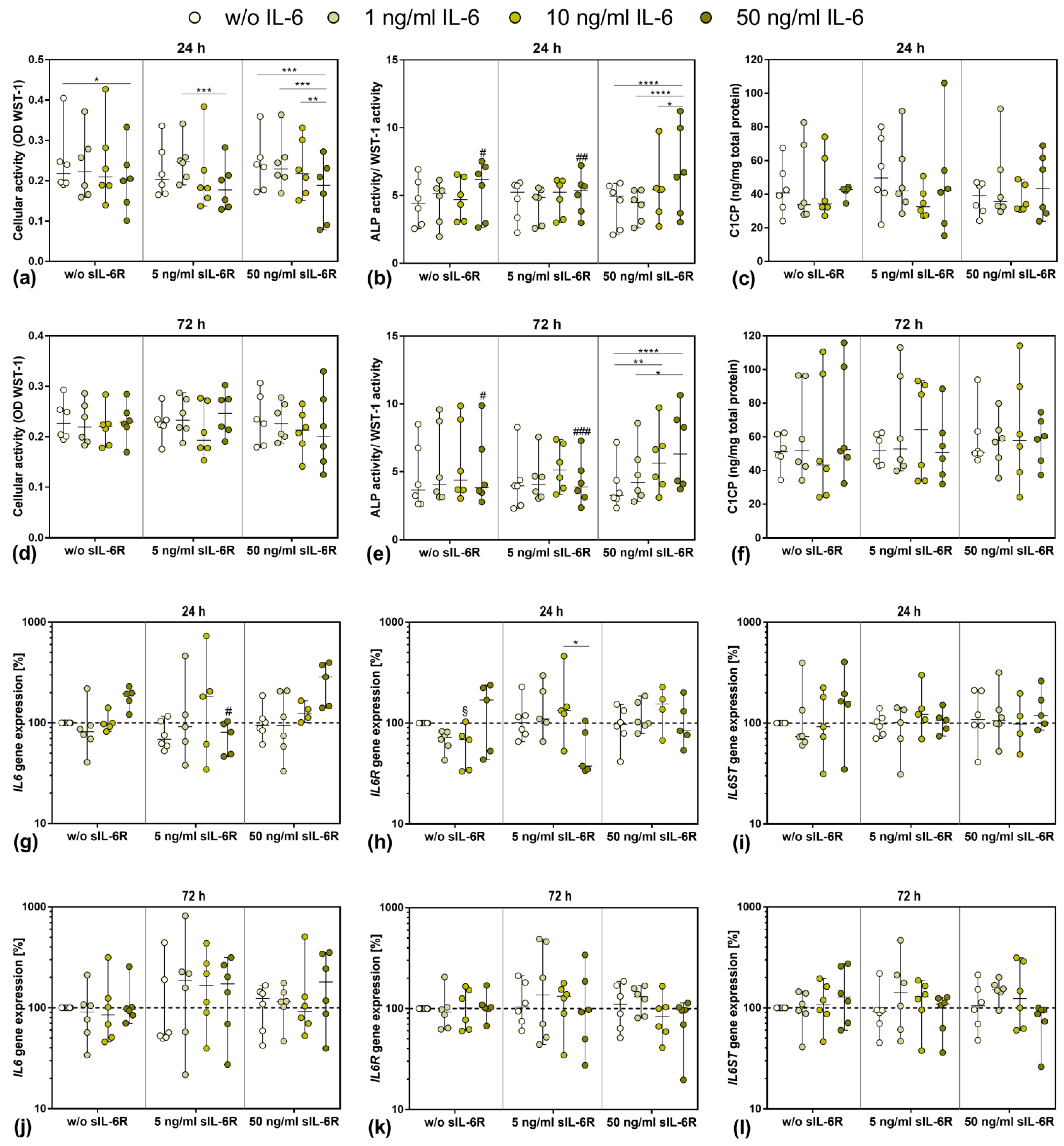
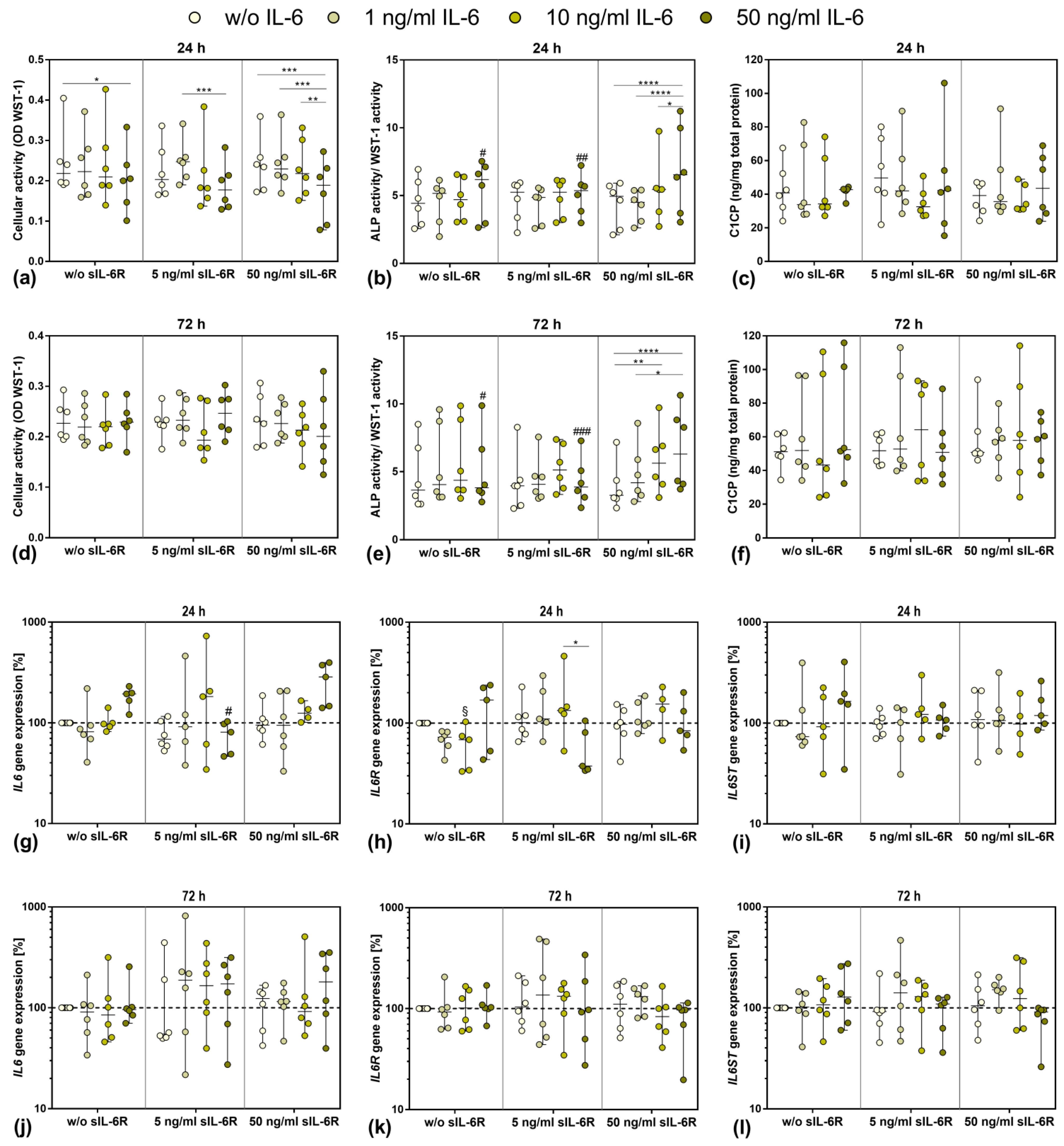
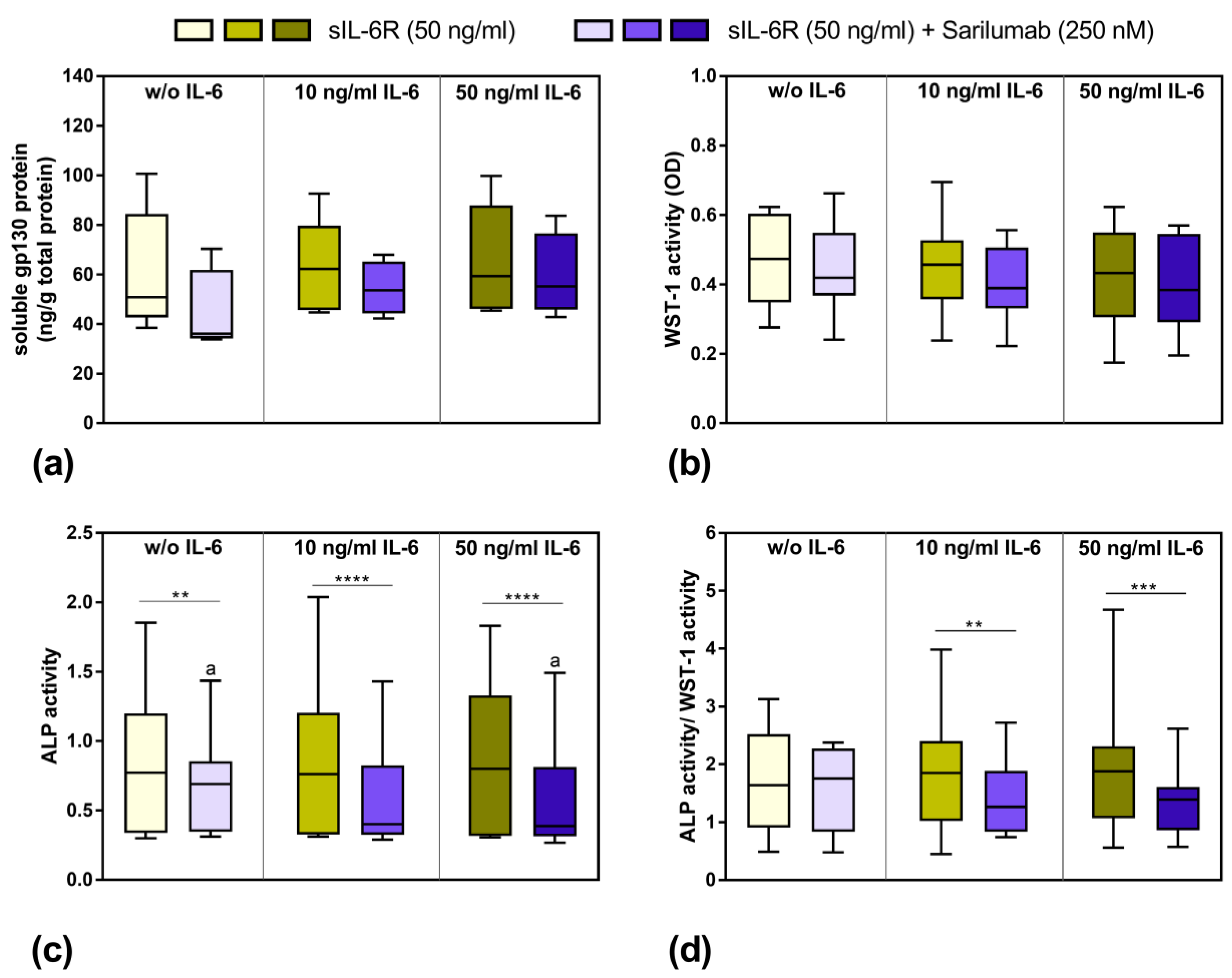
3.3. Influence of Stimulation with Exogenous IL-6 and sIL-6R on Osteoblastic Viability and Differentiation
3.4. Gene Expression of IL6, IL6R, and IL6ST after Exposure to Exogenous IL-6 and sIL-6R
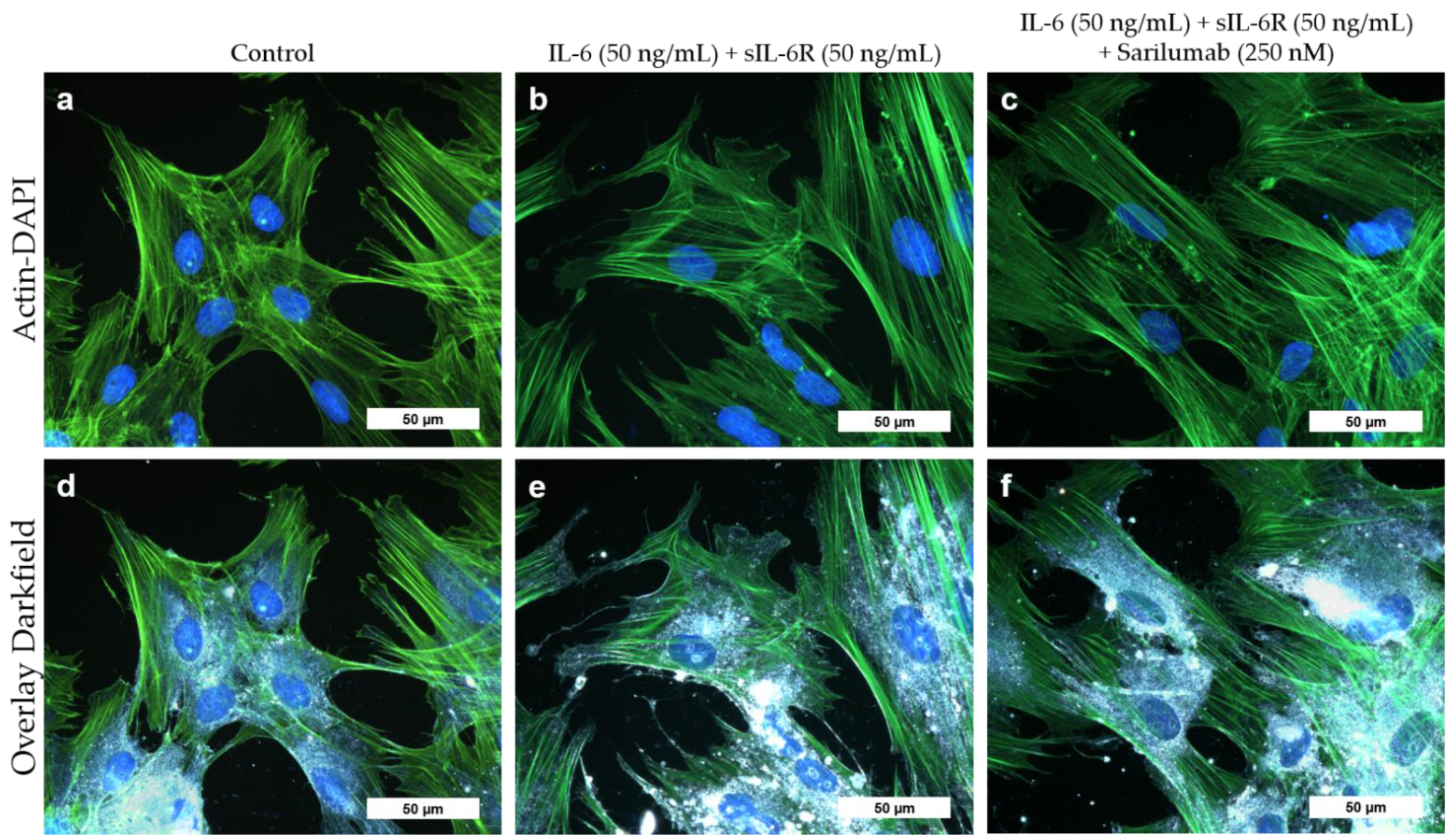
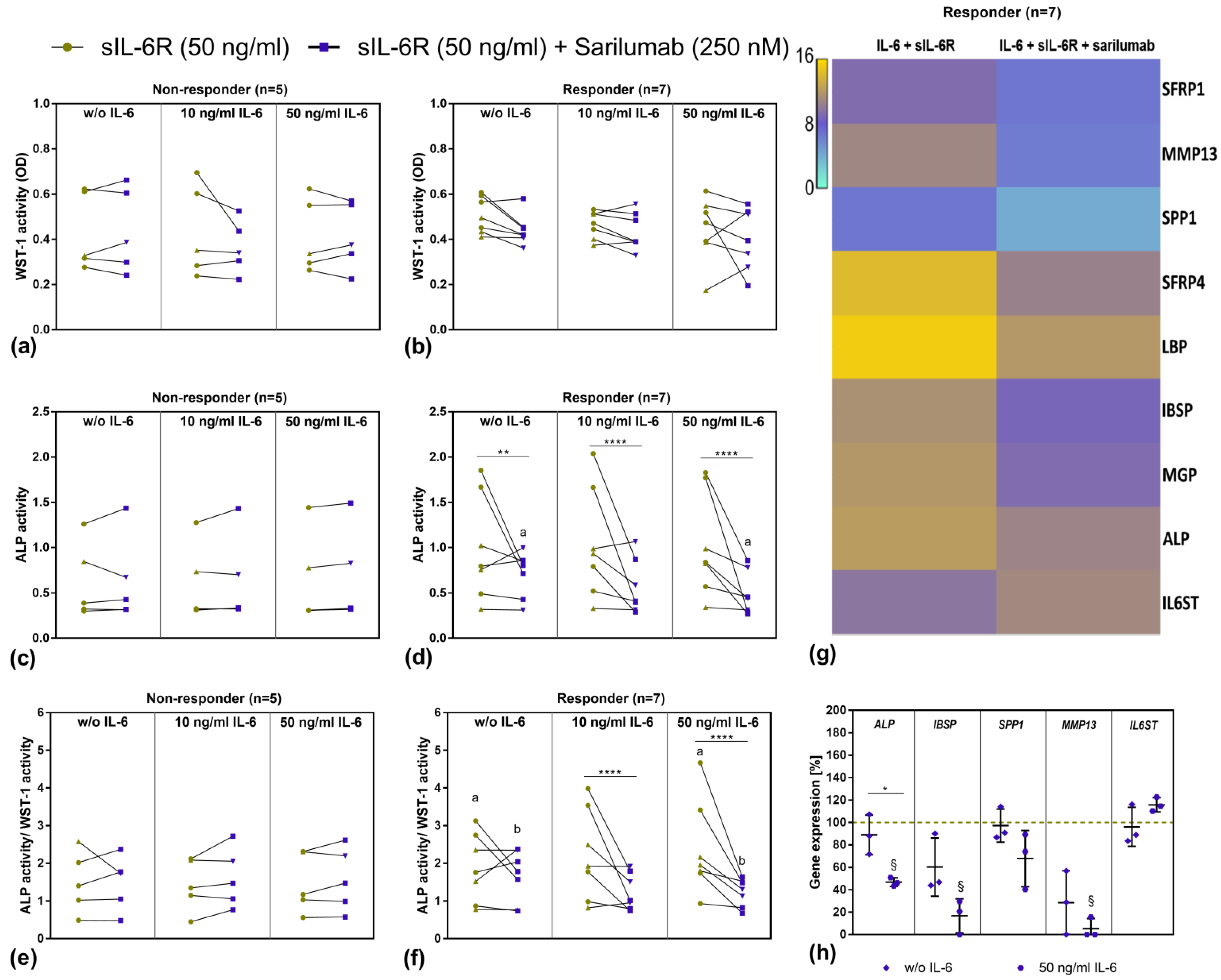
3.5. Viability and Differentiation Capacity of Human Osteoblasts after Exposure to IL-6, sIL-6R, and Sarilumab Treatment
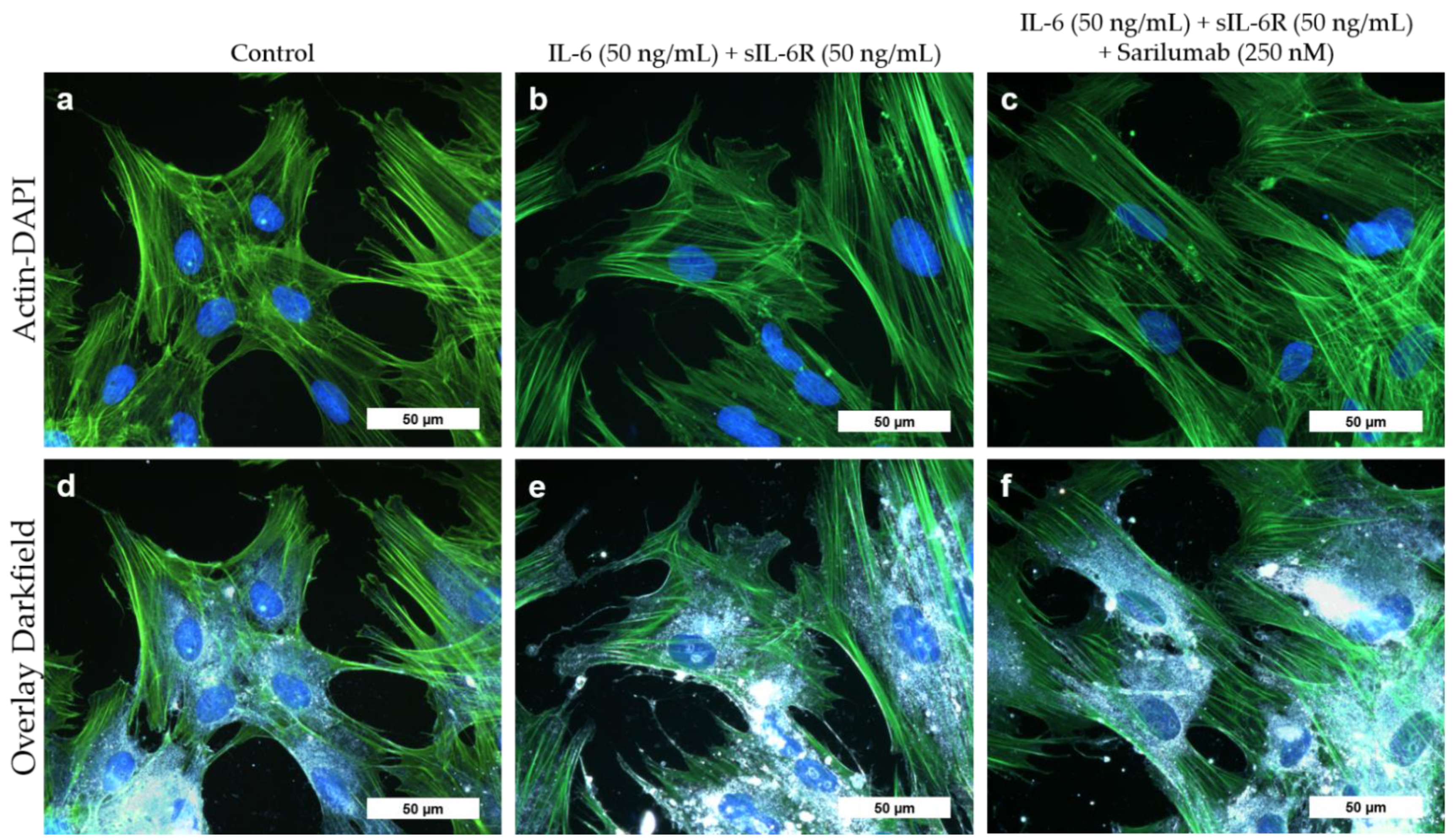
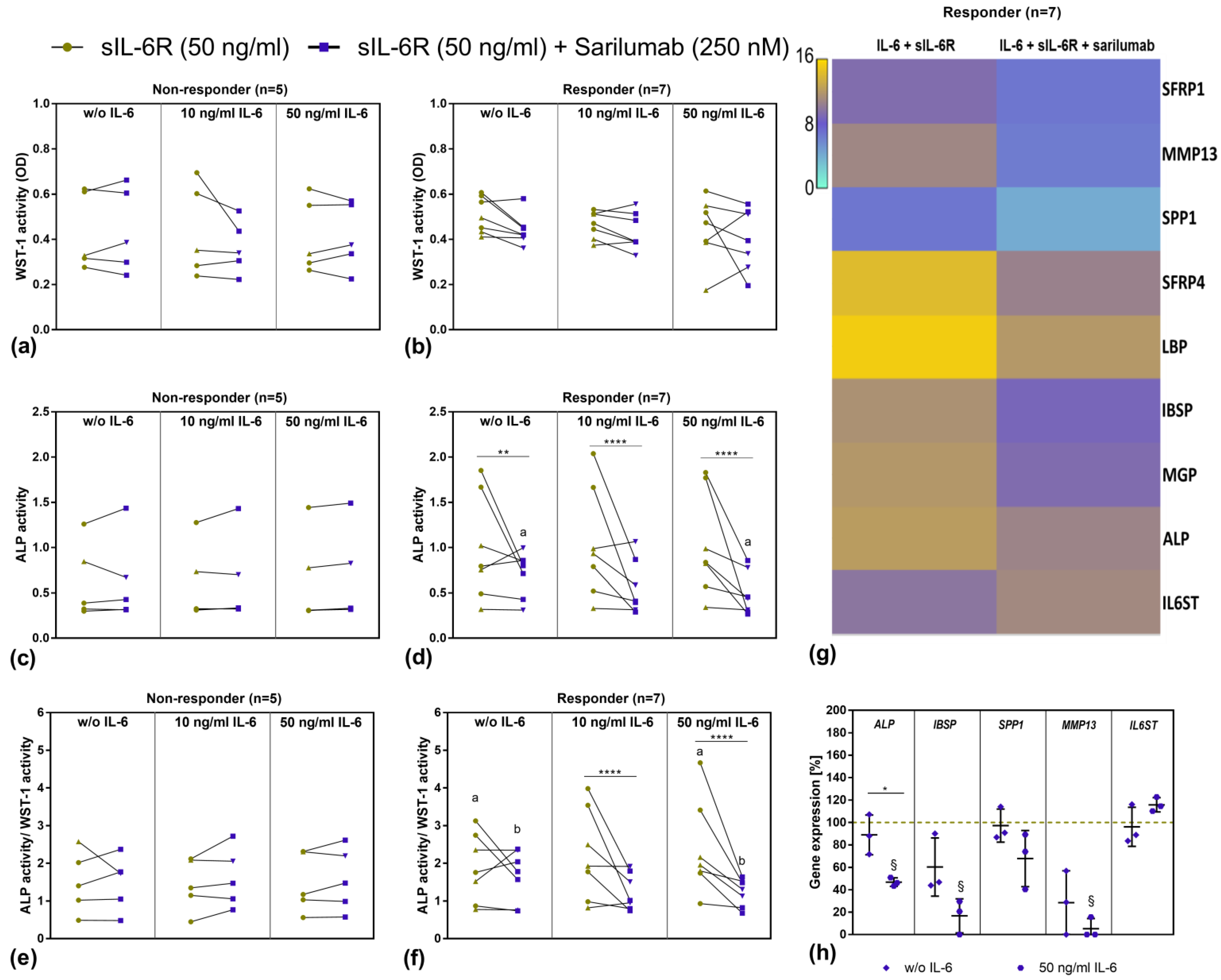
3.6. Transcriptome Analysis of Sarilumab-Responsive Osteoblasts and Confirmation of Results by Independent qPCR Experiments
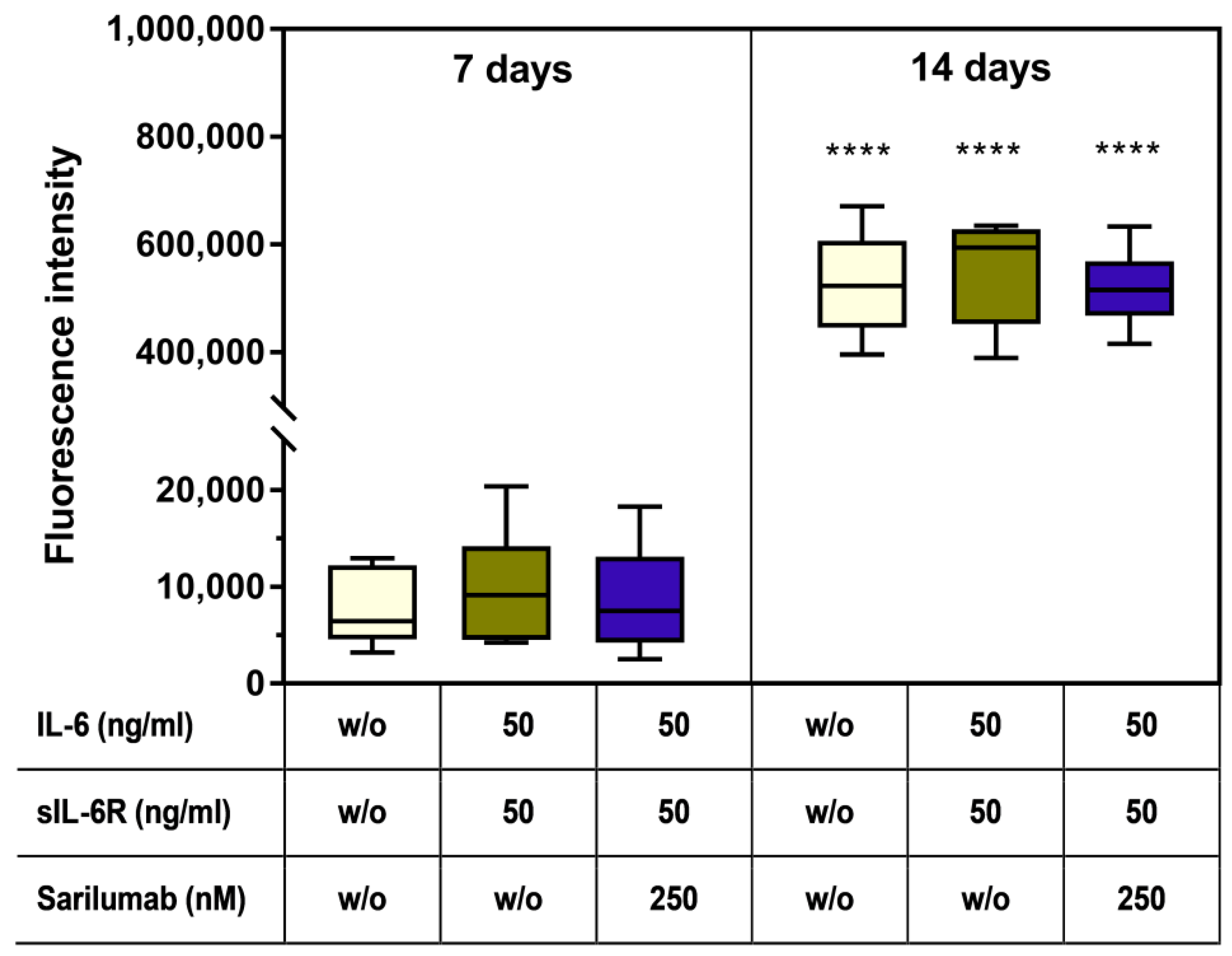
3.7. Mineralization Capacity of Human Osteoblasts Treated with IL-6, sIL-6R ± Sarilumab
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vermes, C.; Jacobs, J.J.; Zhang, J.; Firneisz, G.; Roebuck, K.A.; Glant, T.T. Shedding of the interleukin-6 (IL-6) receptor (gp80) determines the ability of IL-6 to induce gp130 phosphorylation in human osteoblasts. J. Biol. Chem. 2002, 277, 16879–16887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel Meguid, M.H.; Hamad, Y.H.; Swilam, R.S.; Barakat, M.S. Relation of interleukin-6 in rheumatoid arthritis patients to systemic bone loss and structural bone damage. Rheumatol. Int. 2013, 33, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Kaneshiro, S.; Ebina, K.; Shi, K.; Higuchi, C.; Hirao, M.; Okamoto, M.; Koizumi, K.; Morimoto, T.; Yoshikawa, H.; Hashimoto, J. IL-6 negatively regulates osteoblast differentiation through the SHP2/MEK2 and SHP2/Akt2 pathways in vitro. J. Bone Miner. Metab. 2014, 32, 378–392. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.W.; Brennan, H.J.; Vrahnas, C.; Poulton, I.J.; McGregor, N.E.; Standal, T.; Walker, E.C.; Koh, T.T.; Nguyen, H.; Walsh, N.C.; et al. The primary function of gp130 signaling in osteoblasts is to maintain bone formation and strength, rather than promote osteoclast formation. J. Bone Miner. Res. 2014, 29, 1492–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franchimont, N.; Wertz, S.; Malaise, M. Interleukin-6: An osteotropic factor influencing bone formation? Bone 2005, 37, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Sims, N.A.; Jenkins, B.J.; Quinn, J.M.; Nakamura, A.; Glatt, M.; Gillespie, M.T.; Ernst, M.; Martin, T.J. Glycoprotein 130 regulates bone turnover and bone size by distinct downstream signaling pathways. J. Clin. Investig. 2004, 113, 379–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, S.; Schulz, L.; Palmisano, B.; Singh, P.; Berger, J.M.; Yadav, V.K.; Mera, P.; Ellingsgaard, H.; Hidalgo, J.; Brüning, J.; et al. Muscle-derived interleukin 6 increases exercise capacity by signaling in osteoblasts. J. Clin. Investig. 2020, 130, 2888–2902. [Google Scholar] [CrossRef] [Green Version]
- Scheller, J.; Grötzinger, J.; Rose-John, S. Updating interleukin-6 classic- and trans-signaling. Signal. Transduct. 2006, 6, 240–259. [Google Scholar] [CrossRef]
- Sims, N.A.; Walsh, N.C. GP130 cytokines and bone remodelling in health and disease. BMB Rep. 2010, 43, 513–523. [Google Scholar] [CrossRef]
- Raimondo, M.G.; Biggioggero, M.; Crotti, C.; Becciolini, A.; Favalli, E.G. Profile of sarilumab and its potential in the treatment of rheumatoid arthritis. Drug Des. Devel. Ther. 2017, 11, 1593–1603. [Google Scholar] [CrossRef] [Green Version]
- Baran, P.; Hansen, S.; Waetzig, G.H.; Akbarzadeh, M.; Lamertz, L.; Huber, H.J.; Ahmadian, M.R.; Moll, J.M.; Scheller, J. The balance of interleukin (IL)-6, IL-6·soluble IL-6 receptor (sIL-6R), and IL-6·sIL-6R·sgp130 complexes allows simultaneous classic and trans-signaling. J. Biol. Chem. 2018, 293, 6762–6775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbers, C.; Hermanns, H.M.; Schaper, F.; Müller-Newen, G.; Grötzinger, J.; Rose-John, S.; Scheller, J. Plasticity and cross-talk of interleukin 6-type cytokines. Cytokine Growth Factor Rev. 2012, 23, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Rose-John, S.; Scheller, J.; Elson, G.; Jones, S.A. Interleukin-6 biology is coordinated by membrane-bound and soluble receptors: Role in inflammation and cancer. J. Leukoc. Biol. 2006, 80, 227–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGregor, N.E.; Murat, M.; Elango, J.; Poulton, I.J.; Walker, E.C.; Crimeen-Irwin, B.; Ho, P.W.M.; Gooi, J.H.; Martin, T.J.; Sims, N.A. IL-6 exhibits both cis- and trans-signaling in osteocytes and osteoblasts, but only trans-signaling promotes bone formation and osteoclastogenesis. J. Biol. Chem. 2019, 294, 7850–7863. [Google Scholar] [CrossRef] [PubMed]
- Hirabayashi, Y. Tocilizumab can efficiently prevent bone destruction in patients with recent-onset rheumatoid arthritis. Mod. Rheumatol. 2021, 31, 966–971. [Google Scholar] [CrossRef] [PubMed]
- Boyapati, A.; Msihid, J.; Fiore, S.; van Adelsberg, J.; Graham, N.M.; Hamilton, J.D. Sarilumab plus methotrexate suppresses circulating biomarkers of bone resorption and synovial damage in patients with rheumatoid arthritis and inadequate response to methotrexate: A biomarker study of MOBILITY. Arthritis Res. Ther. 2016, 18, 225. [Google Scholar] [CrossRef] [Green Version]
- Schulze, C.; Lochner, K.; Jonitz, A.; Lenz, R.; Duettmann, O.; Hansmann, D.; Bader, R. Cell viability, collagen synthesis and cytokine expression in human osteoblasts following incubation with generated wear particles using different bone cements. Int. J. Mol. Med. 2013, 32, 227–234. [Google Scholar] [CrossRef]
- Burmester, G.R.; Lin, Y.; Patel, R.; van Adelsberg, J.; Mangan, E.K.; Graham, N.M.; van Hoogstraten, H.; Bauer, D.; Ignacio Vargas, J.; Lee, E.B. Efficacy and safety of sarilumab monotherapy versus adalimumab monotherapy for the treatment of patients with active rheumatoid arthritis (MONARCH): A randomised, double-blind, parallel-group phase III trial. Ann. Rheum. Dis. 2017, 76, 840–847. [Google Scholar] [CrossRef]
- Paccaly, A.J.; Kovalenko, P.; Parrino, J.; Boyapati, A.; Xu, C.; van Hoogstraten, H.; Ishii, T.; Davis, J.D.; DiCioccio, A.T. Pharmacokinetics and Pharmacodynamics of Subcutaneous Sarilumab and Intravenous Tocilizumab Following Single-Dose Administration in Patients With Active Rheumatoid Arthritis on Stable Methotrexate. J. Clin. Pharmacol. 2021, 61, 90–104. [Google Scholar] [CrossRef]
- Kivitz, A.; Baret-Cormel, L.; van Hoogstraten, H.; Wang, S.; Parrino, J.; Xu, C.; Stanislav, M. Usability and Patient Preference Phase 3 Study of the Sarilumab Pen in Patients with Active Moderate-to-Severe Rheumatoid Arthritis. Rheumatol. Ther. 2018, 5, 231–242. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Rafique, A.; Potocky, T.; Paccaly, A.; Nolain, P.; Lu, Q.; Iglesias-Rodriguez, M.; St John, G.; Nivens, M.C.; Kanamaluru, V.; et al. Differential Binding of Sarilumab and Tocilizumab to IL-6Rα and Effects of Receptor Occupancy on Clinical Parameters. J. Clin. Pharmacol. 2021, 61, 714–724. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sims, N.A. Cell-specific paracrine actions of IL-6 family cytokines from bone, marrow and muscle that control bone formation and resorption. Int. J. Biochem. Cell Biol. 2016, 79, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Bellido, T.; Borba, V.Z.; Roberson, P.; Manolagas, S.C. Activation of the Janus kinase/STAT (signal transducer and activator of transcription) signal transduction pathway by interleukin-6-type cytokines promotes osteoblast differentiation. Endocrinology 1997, 138, 3666–3676. [Google Scholar] [CrossRef] [PubMed]
- Sims, N.A. The JAK1/STAT3/SOCS3 axis in bone development, physiology, and pathology. Exp. Mol. Med. 2020, 52, 1185–1197. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, F.; Duplomb, L.; Baud’huin, M.; Brounais, B. The dual role of IL-6-type cytokines on bone remodeling and bone tumors. Cytokine Growth Factor Rev. 2009, 20, 19–28. [Google Scholar] [CrossRef]
- Nishimura, R.; Moriyama, K.; Yasukawa, K.; Mundy, G.R.; Yoneda, T. Combination of interleukin-6 and soluble interleukin-6 receptors induces differentiation and activation of JAK-STAT and MAP kinase pathways in MG-63 human osteoblastic cells. J. Bone Miner. Res. 1998, 13, 777–785. [Google Scholar] [CrossRef]
- Liedtke, D.; Hofmann, C.; Jakob, F.; Klopocki, E.; Graser, S. Tissue-Nonspecific Alkaline Phosphatase-A Gatekeeper of Physiological Conditions in Health and a Modulator of Biological Environments in Disease. Biomolecules 2020, 10, 1648. [Google Scholar] [CrossRef]
- Lund, S.A.; Giachelli, C.M.; Scatena, M. The role of osteopontin in inflammatory processes. J. Cell Commun. Signal. 2009, 3, 311–322. [Google Scholar] [CrossRef] [Green Version]
- Graser, S.; Liedtke, D.; Jakob, F. TNAP as a New Player in Chronic Inflammatory Conditions and Metabolism. Int. J. Mol. Sci. 2021, 22, 919. [Google Scholar] [CrossRef]
- Singh, A.; Gill, G.; Kaur, H.; Amhmed, M.; Jakhu, H. Role of osteopontin in bone remodeling and orthodontic tooth movement: A review. Prog. Orthod. 2018, 19, 18. [Google Scholar] [CrossRef] [PubMed]
- Luxembourger, C.; Ruyssen-Witrand, A.; Ladhari, C.; Rittore, C.; Degboe, Y.; Maillefert, J.F.; Gaudin, P.; Marotte, H.; Wendling, D.; Jorgensen, C.; et al. A single nucleotide polymorphism of IL6-receptor is associated with response to tocilizumab in rheumatoid arthritis patients. Pharm. J. 2019, 19, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Montoro, M.; Cañadas-Garre, M.; González-Utrilla, A.; Ángel Calleja-Hernández, M. Influence of IL6R gene polymorphisms in the effectiveness to treatment with tocilizumab in rheumatoid arthritis. Pharm. J. 2018, 18, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Enevold, C.; Baslund, B.; Linde, L.; Josephsen, N.L.; Tarp, U.; Lindegaard, H.; Jacobsen, S.; Nielsen, C.H. Interleukin-6-receptor polymorphisms rs12083537, rs2228145, and rs4329505 as predictors of response to tocilizumab in rheumatoid arthritis. Pharmacogenet. Genom. 2014, 24, 401–405. [Google Scholar] [CrossRef]
- Garbers, C.; Monhasery, N.; Aparicio-Siegmund, S.; Lokau, J.; Baran, P.; Nowell, M.A.; Jones, S.A.; Rose-John, S.; Scheller, J. The interleukin-6 receptor Asp358Ala single nucleotide polymorphism rs2228145 confers increased proteolytic conversion rates by ADAM proteases. Biochim. Biophys. Acta 2014, 1842, 1485–1494. [Google Scholar] [CrossRef] [Green Version]
- Peruzzi, B.; Cappariello, A.; Del Fattore, A.; Rucci, N.; Benedetti, F.D.; Teti, A. c-Src and IL-6 inhibit osteoblast differentiation and integrate IGFBP5 signalling. Nat. Commun. 2012, 3, 630. [Google Scholar] [CrossRef] [Green Version]
- Auréal, M.; Machuca-Gayet, I.; Coury, F. Rheumatoid Arthritis in the View of Osteoimmunology. Biomolecules 2020, 11, 48. [Google Scholar] [CrossRef]
- Terpos, E.; Fragiadaki, K.; Konsta, M.; Bratengeier, C.; Papatheodorou, A.; Sfikakis, P.P. Early effects of IL-6 receptor inhibition on bone homeostasis: A pilot study in women with rheumatoid arthritis. Clin. Exp. Rheumatol. 2011, 29, 921–925. [Google Scholar]
- Houschyar, K.S.; Tapking, C.; Borrelli, M.R.; Popp, D.; Duscher, D.; Maan, Z.N.; Chelliah, M.P.; Li, J.; Harati, K.; Wallner, C.; et al. Wnt Pathway in Bone Repair and Regeneration—What Do We Know So Far. Front. Cell Dev. Biol. 2018, 6, 170. [Google Scholar] [CrossRef]
- Tang, S.Y.; Herber, R.-P.; Ho, S.P.; Alliston, T. Matrix metalloproteinase-13 is required for osteocytic perilacunar remodeling and maintains bone fracture resistance. J. Bone Miner. Res. 2012, 27, 1936–1950. [Google Scholar] [CrossRef] [Green Version]
- Noack, M.; Miossec, P. Selected cytokine pathways in rheumatoid arthritis. Semin. Immunopathol. 2017, 39, 365–383. [Google Scholar] [CrossRef] [PubMed]
- Di Pompo, G.; Errani, C.; Gillies, R.; Mercatali, L.; Ibrahim, T.; Tamanti, J.; Baldini, N.; Avnet, S. Acid-Induced Inflammatory Cytokines in Osteoblasts: A Guided Path to Osteolysis in Bone Metastasis. Front. Cell Dev. Biol. 2021, 9, 678532. [Google Scholar] [CrossRef] [PubMed]
- Palmqvist, P.; Persson, E.; Conaway, H.H.; Lerner, U.H. IL-6, leukemia inhibitory factor, and oncostatin M stimulate bone resorption and regulate the expression of receptor activator of NF-kappa B ligand, osteoprotegerin, and receptor activator of NF-kappa B in mouse calvariae. J. Immunol. 2002, 169, 3353–3362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garnero, P.; Thompson, E.; Woodworth, T.; Smolen, J.S. Rapid and sustained improvement in bone and cartilage turnover markers with the anti-interleukin-6 receptor inhibitor tocilizumab plus methotrexate in rheumatoid arthritis patients with an inadequate response to methotrexate: Results from a substudy of the multicenter double-blind, placebo-controlled trial of tocilizumab in inadequate responders to methotrexate alone. Arthritis Rheum. 2010, 62, 33–43. [Google Scholar] [CrossRef]
- Gnudi, S.; Butturini, L.; Ripamonti, C.; Avella, M.; Bacci, G. The effects of methotrexate (MTX) on bone. A densitometric study conducted on 59 patients with MTX administered at different doses. Ital. J. Orthop. Traumatol. 1988, 14, 227–231. [Google Scholar]
- Robin, F.; Cadiou, S.; Albert, J.-D.; Bart, G.; Coiffier, G.; Guggenbuhl, P. Methotrexate osteopathy: Five cases and systematic literature review. Osteoporos. Int. 2021, 32, 225–232. [Google Scholar] [CrossRef]
- Bourne, L.E.; Wheeler-Jones, C.P.; Orriss, I.R. Regulation of mineralisation in bone and vascular tissue: A comparative review. J. Endocrinol. 2021, 248, R51–R65. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| Alkaline phosphatase (ALP) | CATTGTGACCACCACGAGAG | CCATGATCACGTCAATGTCC |
| Integrin-binding sialoprotein (IBSP) | ATTTTGGGAATGGCCTGTGC | GTCACTACTGCCCTGAACTGG |
| Collagen type 1 (COL1A1) | ACGAAGACATCCCACCAATC | AGATCACGTCATCGCACAAC |
| Glycoprotein 130 (IL6ST) | ACTGTACAACTCGTGTGGAAGAC | TGCTCTCTGCTAAGTTCCCTTG |
| Hypoxanthine-guanine phosphoribosyltransferase (HPRT) | CCCTGGCGTCGTGATTAGTG | TCGAGCAAGACGTTCAGTCC |
| Interleukin 1β (IL1β) | TACTCACTTAAAGCCCGCCT | ATGTGGGAGCGAATGACAGA |
| Interleukin 6 (IL6) | TGGATTCAATGAGGAGACTTGCC | CTGGCATTTGTGGTTGGGTC |
| Interleukin 8 (IL8) | TCTGTGTGAAGGTGCAGTTTTG | ATTTCTGTGTTGGCGCAGTG |
| Matrix metalloprotease 13 (MMP13) | CACGCATAGTCATATAGATACT | CTGGAGATATGATGATACTAAC |
| Interleukin 6 receptor (IL6R) | CTCCTCTGCATTGCCATTGT | TGTGGCTCGAGGTATTGTCA |
| Osteopontin (SPP1) | AACGCCGACCAAGGAAAACT | GCACAGGTGATGCCTAGGAG |
| IL-6 | 0 ng/mL | 1 ng/mL | 10 ng/mL | 50 ng/mL |
|---|---|---|---|---|
| sIL-6R [ng/g] | 1.0 (0.7; 2.3) | 0.9 (0.4; 2.2) | 0.9 (0.5; 1.6) | 0.8 (0.3; 1.8) |
| sgp130 [ng/g] | 76 (39; 90) | 43 (31; 122) | 46 (40; 84) | 41 (28; 94) |
| 50 ng/mL sIL-6R | 50 ng/mL sIL-6R + Sarilumab | |||||
|---|---|---|---|---|---|---|
| IL-6 | 0 ng/mL | 10 ng/mL | 50 ng/mL | 0 ng/mL | 10 ng/mL | 50 ng/mL |
| C1CP [ng/mg] | 56 (29; 85) | 56 (30; 75) | 55 (36; 84) | 55 (35; 111) | 48 (32; 122) | 54 (35; 94) |
| OPN [ng/g] | 31 (18; 64) | 19 (18; 52) | 29 (0; 59) | 0 (0; 192) | 21 (0; 58) | 17 (0; 302) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klinder, A.; Waletzko-Hellwig, J.; Sellin, M.-L.; Seyfarth-Sehlke, A.; Wolfien, M.; Prehn, F.; Bader, R.; Jonitz-Heincke, A. Effects of the Interleukin-6 Receptor Blocker Sarilumab on Metabolic Activity and Differentiation Capacity of Primary Human Osteoblasts. Pharmaceutics 2022, 14, 1390. https://doi.org/10.3390/pharmaceutics14071390
Klinder A, Waletzko-Hellwig J, Sellin M-L, Seyfarth-Sehlke A, Wolfien M, Prehn F, Bader R, Jonitz-Heincke A. Effects of the Interleukin-6 Receptor Blocker Sarilumab on Metabolic Activity and Differentiation Capacity of Primary Human Osteoblasts. Pharmaceutics. 2022; 14(7):1390. https://doi.org/10.3390/pharmaceutics14071390
Chicago/Turabian StyleKlinder, Annett, Janine Waletzko-Hellwig, Marie-Luise Sellin, Anika Seyfarth-Sehlke, Markus Wolfien, Franziska Prehn, Rainer Bader, and Anika Jonitz-Heincke. 2022. "Effects of the Interleukin-6 Receptor Blocker Sarilumab on Metabolic Activity and Differentiation Capacity of Primary Human Osteoblasts" Pharmaceutics 14, no. 7: 1390. https://doi.org/10.3390/pharmaceutics14071390
APA StyleKlinder, A., Waletzko-Hellwig, J., Sellin, M.-L., Seyfarth-Sehlke, A., Wolfien, M., Prehn, F., Bader, R., & Jonitz-Heincke, A. (2022). Effects of the Interleukin-6 Receptor Blocker Sarilumab on Metabolic Activity and Differentiation Capacity of Primary Human Osteoblasts. Pharmaceutics, 14(7), 1390. https://doi.org/10.3390/pharmaceutics14071390