
Antipsoriatic Potential of Quebecol and Its Derivatives

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
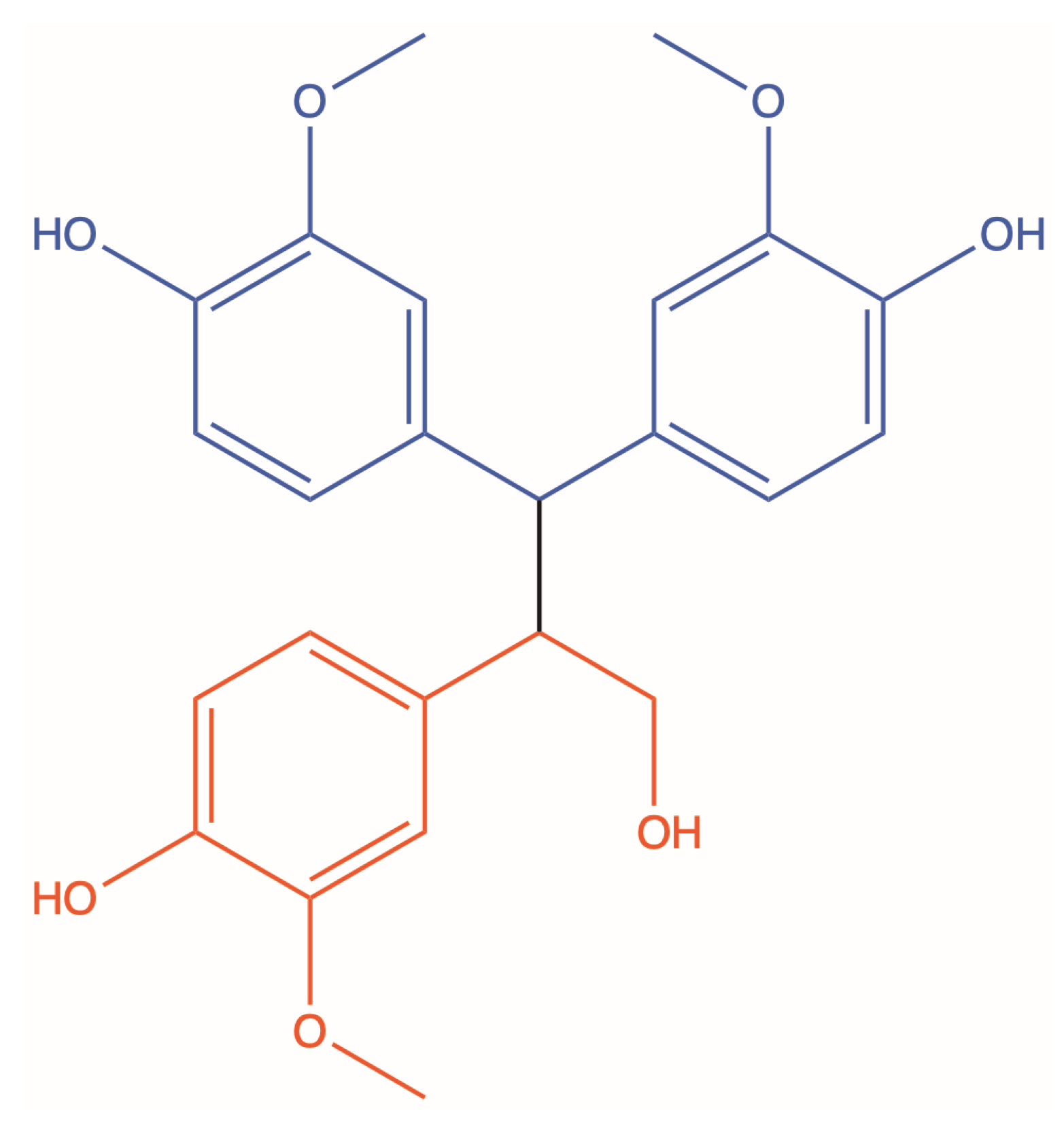
2.1. Synthesis of CPD1, CPD2 and CPD3
2.2. Biological Testing
2.3. Biopsies
2.4. Cell Extraction
2.5. Cell Culture
2.6. Sulforhodamine B Assay
2.7. Viability Assay
2.8. Skin Substitute Production
2.9. Histological Analyses
2.10. Immunofluorescence Staining
2.11. Statistical Analysis
3. Results
3.1. Antiproliferative Potential
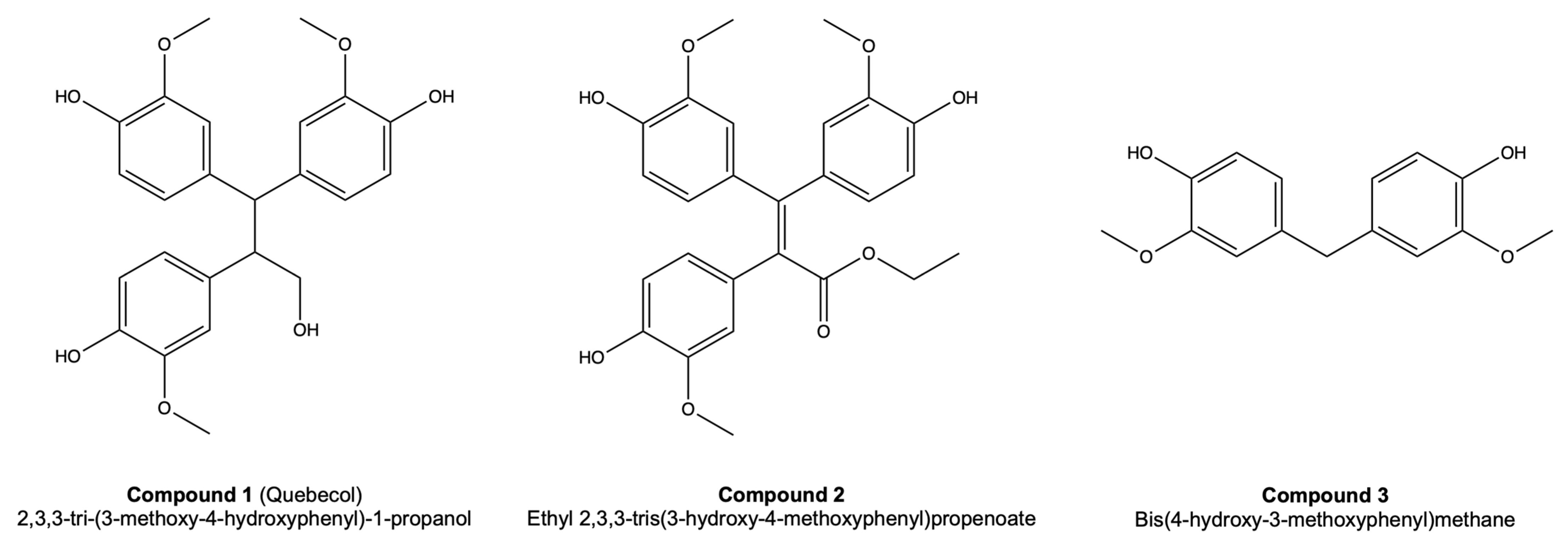
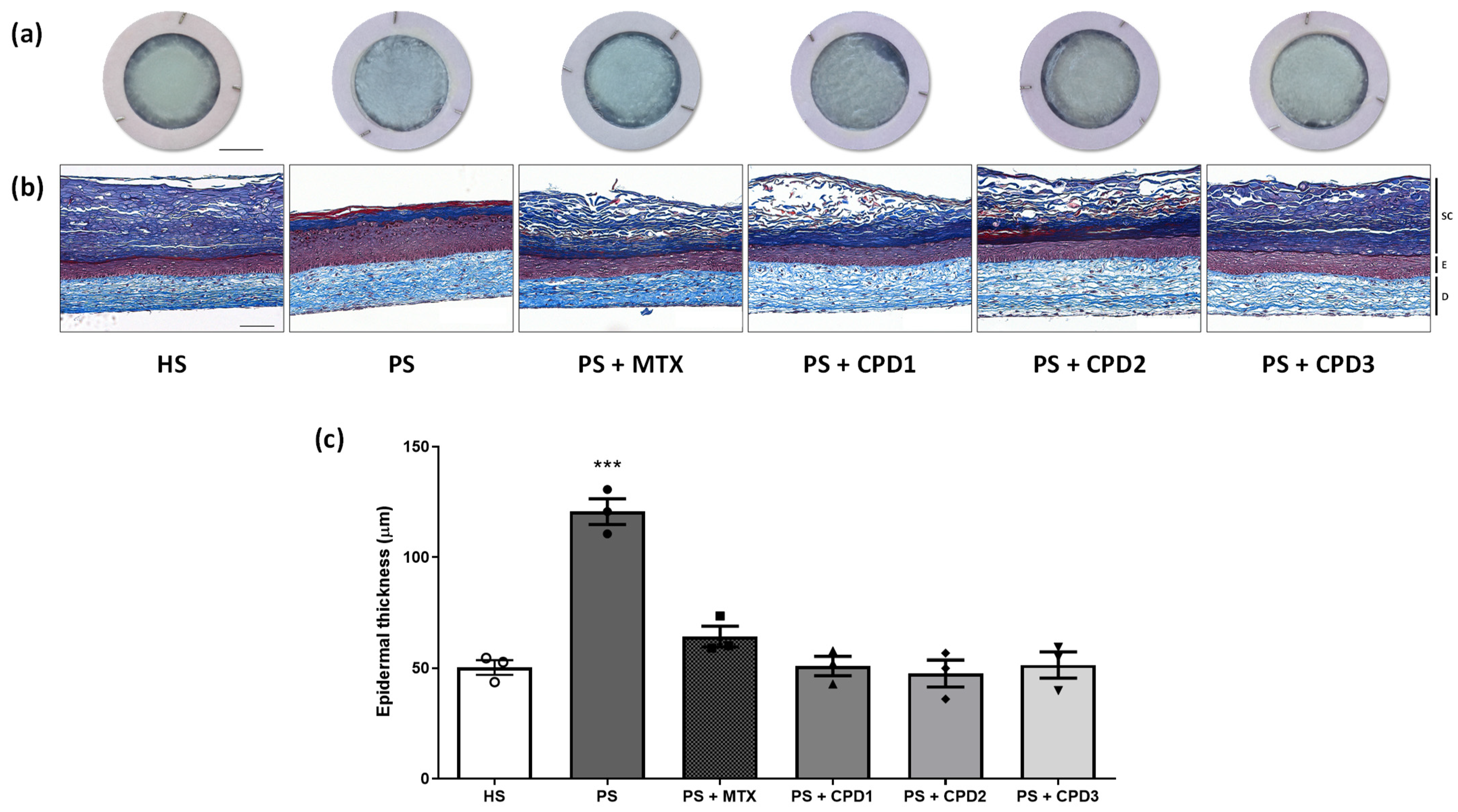
3.2. Skin Substitute Morphology
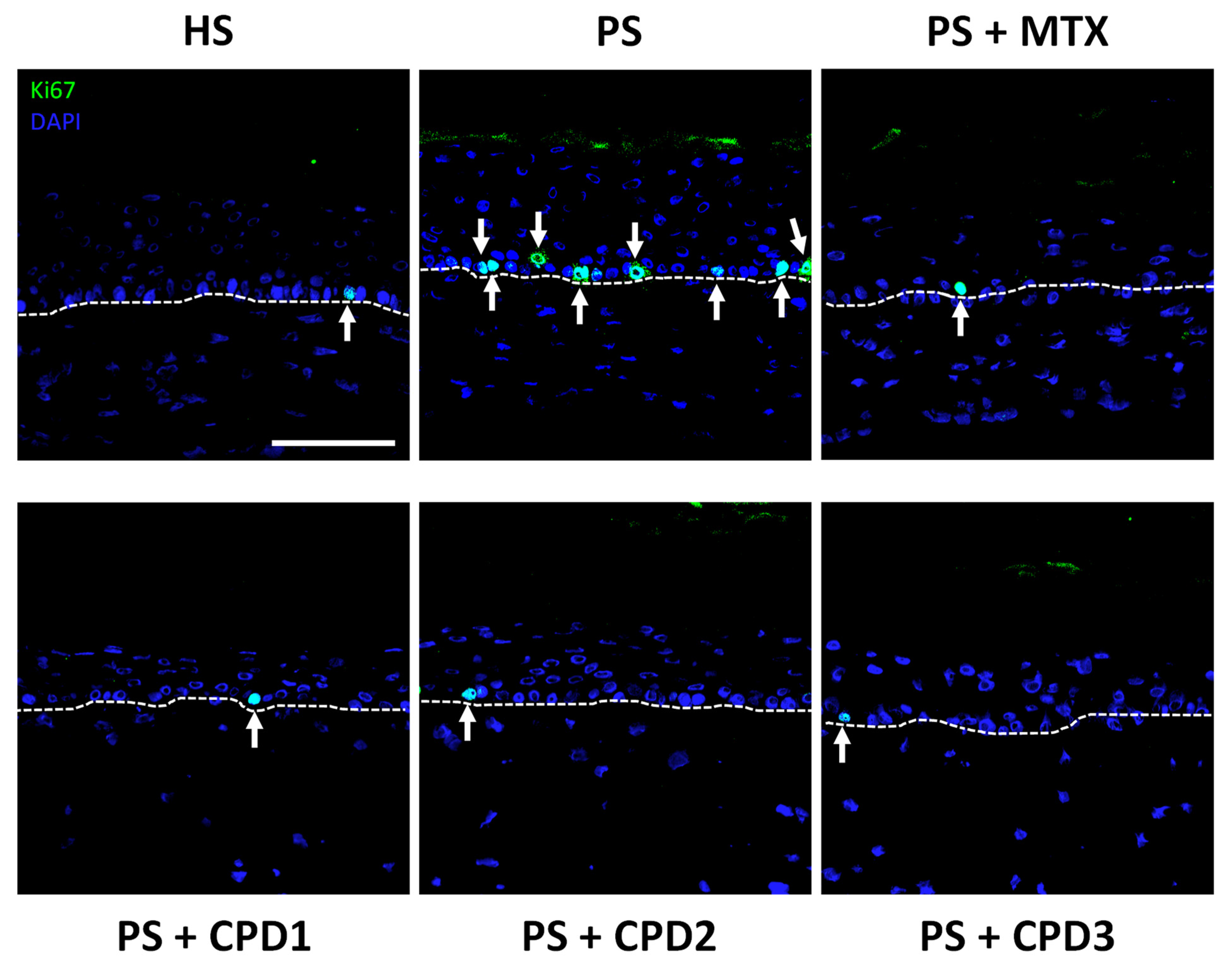
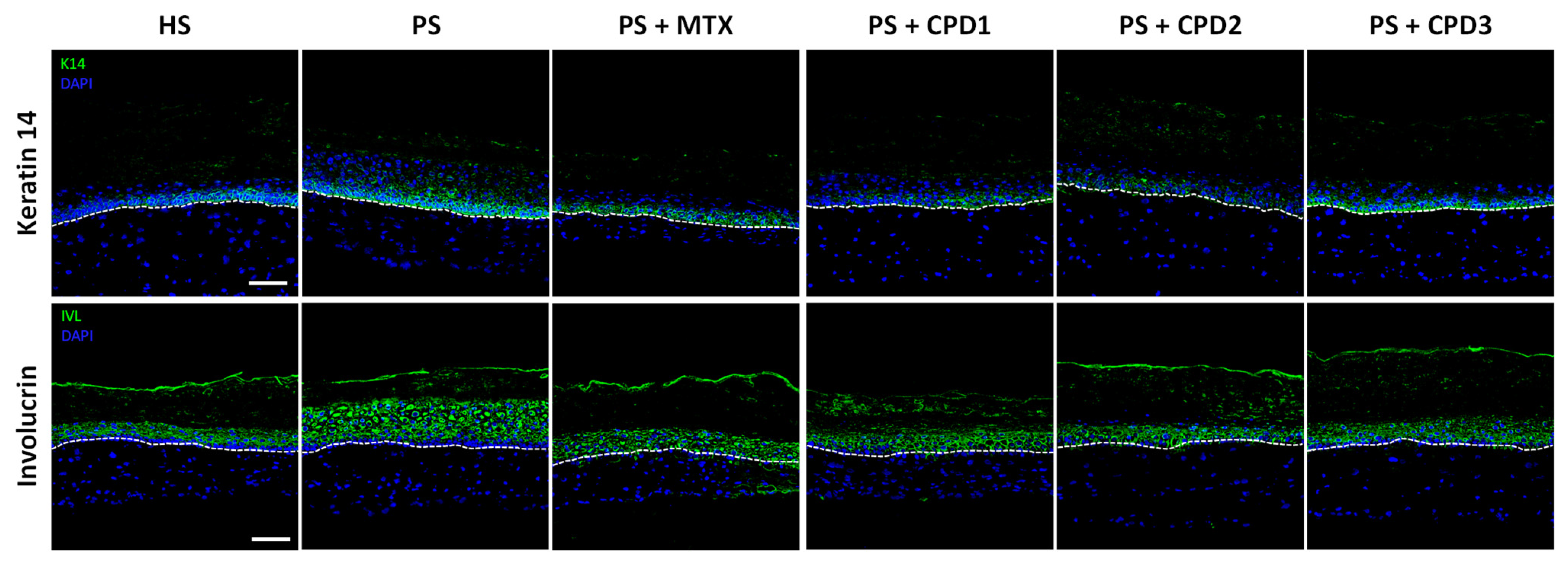
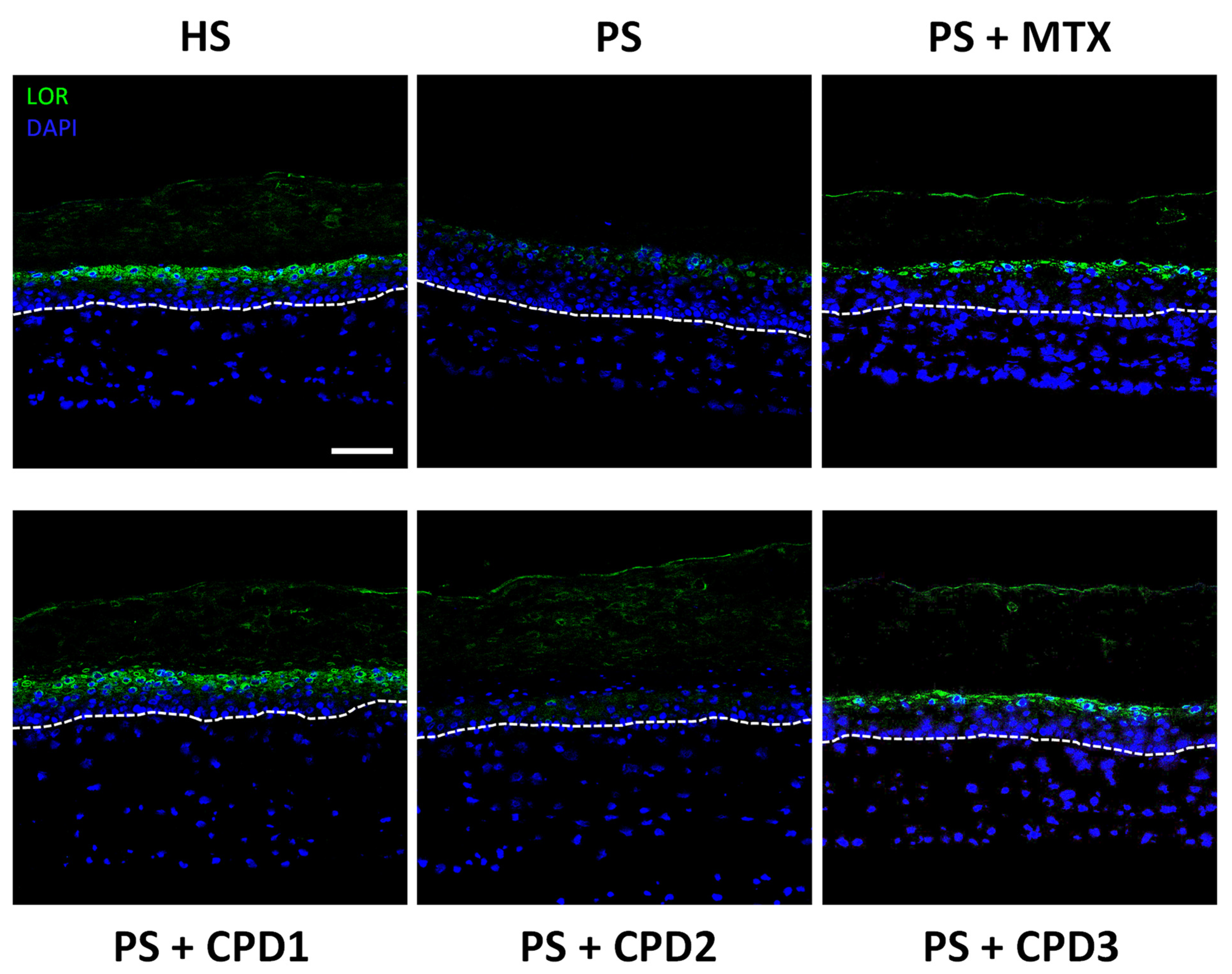
3.3. Regulation of Hyperproliferation and Abnormal Differentiation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raychaudhuri, S.; Farber, E. The prevalence of psoriasis in the world. J. Eur. Acad. Dermatol. Venereol. 2001, 15, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Srinivasan, S. Histopathologic diagnostic parameters of psoriasis; a clinicopathological study. Int. J. Res. Med. Sci. 2016, 4, 1915–1920. [Google Scholar] [CrossRef]
- Camisa, C. Pathogenesis of Psoriasis. In Handbook of Psoriasis, 2nd ed.; Camisa, C., Ed.; Blackwell Publishing: Malden, MA, USA, 2004; pp. 45–60. [Google Scholar]
- Eroschenko, V.P. DiFiore’s Atlas of Histology with Functional Correlations, 11th ed.; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2008; p. 532. [Google Scholar]
- Stern, R.S.; Nijsten, T.; Feldman, S.R.; Margolis, D.J.; Rolstad, T. Psoriasis Is Common, Carries a Substantial Burden Even When Not Extensive, and Is Associated with Widespread Treatment Dissatisfaction. J. Investig. Dermatol. Symp. Proc. 2004, 9, 136–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tveit, K.S.; Duvetorp, A.; Østergaard, M.; Skov, L.; Danielsen, K.; Iversen, L.; Seifert, O. Treatment use and satisfaction among patients with psoriasis and psoriatic arthritis: Results from the NORdic PAtient survey of Psoriasis and Psoriatic arthritis (NORPAPP). J. Eur. Acad. Dermatol. Venereol. 2019, 33, 340–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legault, J.; Girard-Lalancette, K.; Grenon, C.; Dussault, C.; Pichette, A. Antioxidant Activity, Inhibition of Nitric Oxide Overproduction, and In Vitro Antiproliferative Effect of Maple Sap and Syrup from Acer saccharum. J. Med. Food 2010, 13, 460–468. [Google Scholar] [CrossRef]
- Thériault, M.; Caillet, S.; Kermasha, S.; Lacroix, M. Antioxidant, antiradical and antimutagenic activities of phenolic compounds present in maple products. Food Chem. 2006, 98, 490–501. [Google Scholar] [CrossRef]
- Nahar, P.P.; Driscoll, M.V.; Li, L.; Slitt, A.L.; Seeram, N.P. Phenolic mediated anti-inflammatory properties of a maple syrup extract in RAW 264.7 murine macrophages. J. Funct. Foods 2014, 6, 126–136. [Google Scholar] [CrossRef]
- González-Sarrías, A.; Li, L.; Seeram, N.P. Anticancer effects of maple syrup phenolics and extracts on proliferation, apoptosis, and cell cycle arrest of human colon cells. J. Funct. Foods 2012, 4, 185–196. [Google Scholar] [CrossRef]
- Li, L.; Seeram, N.P. Quebecol, a novel phenolic compound isolated from Canadian maple syrup. J. Funct. Foods 2011, 3, 125–128. [Google Scholar] [CrossRef]
- Cardinal, S.; Voyer, N. Total synthesis of quebecol. Tetrahedron Lett. 2013, 54, 5178–5180. [Google Scholar] [CrossRef]
- Pericherla, K.; Shirazi, A.N.; Rao, V.K.; Tiwari, R.K.; DaSilva, N.; McCaffrey, K.T.; Beni, Y.A.; González-Sarrías, A.; Seeram, N.P.; Parang, K.; et al. Synthesis and antiproliferative activities of quebecol and its analogs. Bioorg. Med. Chem. Lett. 2013, 23, 5329–5331. [Google Scholar] [CrossRef] [Green Version]
- Cardinal, S.; Azelmat, J.; Grenier, D.; Voyer, N. Anti-inflammatory properties of quebecol and its derivatives. Bioorg. Med. Chem. Lett. 2016, 26, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Cardinal, S.; Paquet-Côté, P.-A.; Azelmat, J.; Bouchard, C.; Grenier, D.; Voyer, N. Synthesis and anti-inflammatory activity of isoquebecol. Bioorg. Med. Chem. 2017, 25, 2043–2056. [Google Scholar] [CrossRef] [PubMed]
- Czarnecka-Operacz, M.; Sadowska-Przytocka, A. The possibilities and principles of methotrexate treatment of psoriasis—The updated knowledge. Postepy. Dermatol. Alergol. 2014, 31, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Menting, S.P.; Dekker, P.M.; Limpens, J.; Hooft, L.; Spuls, P.I. Methotrexate Dosing Regimen for Plaque-type Psoriasis: A Systematic Review of the Use of Test-dose, Start-dose, Dosing Scheme, Dose Adjustments, Maximum Dose and Folic Acid Supplementation. Acta Derm. Venereol. 2016, 96, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Bélanger, A.; Grenier, A.; Simard, F.; Gendreau, I.; Pichette, A.; Legault, J.; Pouliot, R. Dihydrochalcone Derivatives from Populus balsamifera L. Buds for the Treatment of Psoriasis. Int. J. Mol. Sci. 2019, 21, 256. [Google Scholar] [CrossRef] [Green Version]
- Germain, L.; Rouabhia, M.; Guignard, R.; Carrier, L.; Bouvard, V.; Auger, F.A. Improvement of human keratinocyte isolation and culture using thermolysin. Burns 1993, 19, 99–104. [Google Scholar] [CrossRef]
- Auger, F.A.; López Valle, C.A.; Guignard, R.; Tremblay, N.; Noel, B.; Goulet, F.; Germain, L. Skin equivalent produced with human collagen. In Vitro Cell. Dev. Biol. Anim. 1995, 31, 432–439. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Larouche, D.; Jean, J.; Berthod, F.; Germain, L.; Pouliot, R. Markers for an in vitro skin substitute. In Methods in Bioengineering: Alternative Technologies to Animal Testing; Maguire, T., Novik, E., Eds.; Artech House: Boston, MA, USA, 2010; pp. 183–203. [Google Scholar]
- Jean, J.; García-Pérez, M.-E.; Pouliot, R. Bioengineered Skin: The Self- Assembly Approach. J. Tissue Sci. Eng. 2011, S5, 001. [Google Scholar] [CrossRef]
- Jean, J.; Lapointe, M.; Soucy, J.; Pouliot, R. Development of an in vitro psoriatic skin model by tissue engineering. J. Dermatol. Sci. 2009, 53, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Jean, J.; Soucy, J.; Pouliot, R. Effects of Retinoic Acid on Keratinocyte Proliferation and Differentiation in a Psoriatic Skin Model. Tissue Eng. Part A 2011, 17, 1859–1868. [Google Scholar] [CrossRef] [PubMed]
- Jean, J.; Leroy, M.; Duque-Fernandez, A.; Bernard, G.; Soucy, J.; Pouliot, R. Characterization of a psoriatic skin model produced with involved or uninvolved cells. J. Tissue Eng. Regen. Med. 2015, 9, 789–798. [Google Scholar] [CrossRef]
- Morin, A.; Simard, M.; Rioux, G.; Grenier, A.; Morin, S.; Pouliot, R. Application of an In Vitro Psoriatic Skin Model to Study Cutaneous Metabolization of Tazarotene. Processes 2019, 7, 871. [Google Scholar] [CrossRef] [Green Version]
- Simard, M.; Rioux, G.; Morin, S.; Martin, C.; Guérin, S.L.; Flamand, N.; Julien, P.; Fradette, J.; Pouliot, R. Investigation of Omega-3 Polyunsaturated Fatty Acid Biological Activity in a Tissue-Engineered Skin Model Involving Psoriatic Cells. J. Investig. Dermatol. 2021, 141, 2391–2401.e2313. [Google Scholar] [CrossRef]
- Morin, S.; Simard, M.; Flamand, N.; Pouliot, R. Biological action of docosahexaenoic acid in a 3D tissue-engineered psoriatic skin model: Focus on the PPAR signaling pathway. Biochim. Biophys. Acta 2021, 1866, 159032. [Google Scholar] [CrossRef] [PubMed]
- Grenier, A.; Legault, J.; Pichette, A.; Jean, L.; Bélanger, A.; Pouliot, R. Antioxidant, Anti-Inflammatory, and Anti-Aging Potential of a Kalmia angustifolia Extract and Identification of Some Major Compounds. Antioxidants 2021, 10, 1373. [Google Scholar] [CrossRef]
- Niehues, H.; van den Bogaard, E.H. Past, present and future of in vitro 3D reconstructed inflammatory skin models to study psoriasis. Exp. Dermatol. 2018, 27, 512–519. [Google Scholar] [CrossRef] [Green Version]
- Blauvelt, A. IL-6 Differs from TNF-α: Unpredicted Clinical Effects Caused by IL-6 Blockade in Psoriasis. J. Investig. Dermatol. 2017, 137, 541–542. [Google Scholar] [CrossRef] [Green Version]
- Grossman, R.M.; Krueger, J.; Yourish, D.; Granelli-Piperno, A.; Murphy, D.P.; May, L.T.; Kupper, T.S.; Sehgal, P.B.; Gottlieb, A.B. Interleukin 6 is expressed in high levels in psoriatic skin and stimulates proliferation of cultured human keratinocytes. Proc. Natl. Acad. Sci. USA 1989, 86, 6367–6371. [Google Scholar] [CrossRef] [Green Version]
- Ettehadi, P.; Greaves, M.W.; Wallach, D.; Aderka, D.; Camp, R.D.R. Elevated tumour necrosis factor-alpha (TNF-α) biological activity in psoriatic skin lesions. Clin. Exp. Immunol. 1994, 96, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Boyman, O.; Hefti, H.P.; Conrad, C.; Nickoloff, B.J.; Suter, M.; Nestle, F.O. Spontaneous Development of Psoriasis in a New Animal Model Shows an Essential Role for Resident T Cells and Tumor Necrosis Factor-α. J. Exp. Med. 2004, 199, 731–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashir, M.M.; Sharma, M.R.; Werth, V.P. TNF-α production in the skin. Arch. Dermatol. Res. 2009, 301, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Köck, A.; Schwarz, T.; Kirnbauer, R.; Urbanski, A.; Perry, P.; Ansel, J.C.; Luger, T.A. Human keratinocytes are a source for tumor necrosis factor alpha: Evidence for synthesis and release upon stimulation with endotoxin or ultraviolet light. J. Exp. Med. 1990, 172, 1609–1614. [Google Scholar] [CrossRef] [Green Version]
- Goldminz, A.M.; Au, S.C.; Kim, N.; Gottlieb, A.B.; Lizzul, P.F. NF-κB: An essential transcription factor in psoriasis. J. Dermatol. Sci. 2013, 69, 89–94. [Google Scholar] [CrossRef]
- Lizzul, P.F.; Aphale, A.; Malaviya, R.; Sun, Y.; Masud, S.; Dombrovskiy, V.; Gottlieb, A.B. Differential Expression of Phosphorylated NF-κB/RelA in Normal and Psoriatic Epidermis and Downregulation of NF-κB in Response to Treatment with Etanercept. J. Investig. Dermatol. 2005, 124, 1275–1283. [Google Scholar] [CrossRef] [Green Version]
- Rebholz, B.; Haase, I.; Eckelt, B.; Paxian, S.; Flaig, M.J.; Ghoreschi, K.; Nedospasov, S.A.; Mailhammer, R.; Debey-Pascher, S.; Schultze, J.L.; et al. Crosstalk between Keratinocytes and Adaptive Immune Cells in an IκBα Protein-Mediated Inflammatory Disease of the Skin. Immunity 2007, 27, 296–307. [Google Scholar] [CrossRef] [Green Version]
- Moorchung, N.; Kulaar, J.S.; Chatterjee, M.; Vasudevan, B.; Tripathi, T.; Dutta, V. Role of NF-κB in the pathogenesis of psoriasis elucidated by its staining in skin biopsy specimens. Int. J. Dermatol. 2014, 53, 570–574. [Google Scholar] [CrossRef]
- García-Pérez, M.-E.; Allaeys, I.; Rusu, D.; Pouliot, R.; Janezic, T.S.; Poubelle, P.E. Picea mariana polyphenolic extract inhibits phlogogenic mediators produced by TNF-α-activated psoriatic keratinocytes: Impact on NF-κB pathway. J. Ethnopharmacol. 2014, 151, 265–278. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef]
- Huang, T.-H.; Lin, C.-F.; Alalaiwe, A.; Yang, S.-C.; Fang, J.-Y. Apoptotic or Antiproliferative Activity of Natural Products against Keratinocytes for the Treatment of Psoriasis. Int. J. Mol. Sci. 2019, 20, 2558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thewes, M.; Stadler, R.; Korge, B.; Mischke, D. Normal psoriatic epidermis expression of hyperproliferation-associated keratins. Arch. Dermatol. Res. 1991, 283, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Rice, R.H.; Green, H. Presence in human epidermal cells of a soluble protein precursor of the cross-linked envelope: Activation of the cross-linking by calcium ions. Cell 1979, 18, 681–694. [Google Scholar] [CrossRef]
- Steinert, P.M.; Marekov, L.N. Direct Evidence That Involucrin Is a Major Early Isopeptide Cross-linked Component of the Keratinocyte Cornified Cell Envelope. J. Biol. Chem. 1997, 272, 2021–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida-Yamamoto, A.; Iizuka, H. Differences in Involucrin Immunolabeling Within Cornified Cell Envelopes in Normal and Psoriatic Epidermis. J. Investig. Dermatol. 1995, 104, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Steven, A.C.; Bisher, M.E.; Roop, D.R.; Steinert, P.M. Biosynthetic pathways of filaggrin and loricrin—two major proteins expressed by terminally differentiated epidermal keratinocytes. J. Struct. Biol. 1990, 104, 150–162. [Google Scholar] [CrossRef]
- Murphy, M.; Kerr, P.; Grant-Kels, J.M. The histopathologic spectrum of psoriasis. Clin. Dermatol. 2007, 25, 524–528. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC20 (μM) | Cell Viability (%) |
|---|---|---|
| CPD1 | 400 | 97 |
| CPD2 | 150 | 94 |
| CPD3 | 350 | 97 |
| MTX | 734 1 | 85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouchard, C.; Grenier, A.; Cardinal, S.; Bélanger, S.; Voyer, N.; Pouliot, R. Antipsoriatic Potential of Quebecol and Its Derivatives. Pharmaceutics 2022, 14, 1129. https://doi.org/10.3390/pharmaceutics14061129
Bouchard C, Grenier A, Cardinal S, Bélanger S, Voyer N, Pouliot R. Antipsoriatic Potential of Quebecol and Its Derivatives. Pharmaceutics. 2022; 14(6):1129. https://doi.org/10.3390/pharmaceutics14061129
Chicago/Turabian StyleBouchard, Corinne, Alexe Grenier, Sébastien Cardinal, Sarah Bélanger, Normand Voyer, and Roxane Pouliot. 2022. "Antipsoriatic Potential of Quebecol and Its Derivatives" Pharmaceutics 14, no. 6: 1129. https://doi.org/10.3390/pharmaceutics14061129
APA StyleBouchard, C., Grenier, A., Cardinal, S., Bélanger, S., Voyer, N., & Pouliot, R. (2022). Antipsoriatic Potential of Quebecol and Its Derivatives. Pharmaceutics, 14(6), 1129. https://doi.org/10.3390/pharmaceutics14061129