
Integrin-αvβ3 as a Therapeutic Target in Glioblastoma: Back to the Future?


{kind=link}
Abstract
1. Introduction
2. Role of Integrin-αvβ3 in GBM Progression
2.1. Integrin Signaling
2.2. Expression of Integrin-αvβ3 in GBM
2.3. Roles of Integrin-αvβ3 in Angiogenesis
2.4. Roles of Integrin in Migration and Invasion Processes
2.5. Integrin-αvβ3, Stemness and Drug Resistance
3. Anti-Integrin-αvβ3 Agents for Anti-Cancer Therapy
3.1. RGD-Derived Antagonists
3.2. Integrin-αvβ3 and Drug Delivery
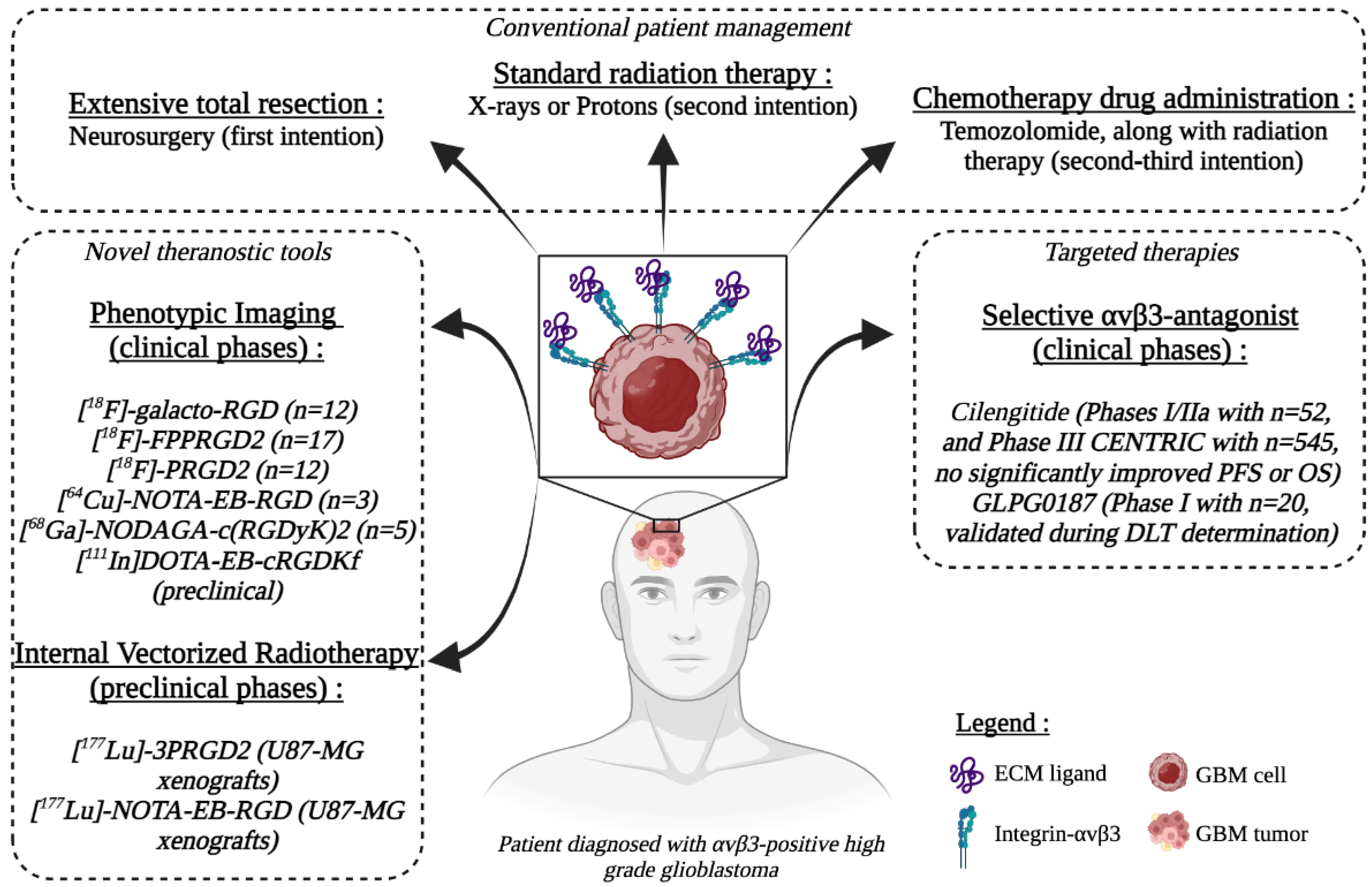
3.3. Theranostics Development of αvβ3-Targeting Agents
3.3.1. Integrin-αvβ3 Targeting for Phenotypic Imaging
3.3.2. Integrin-αvβ3-Targeted Radiotherapy
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ostrom, Q.T.; Gittleman, H.; Liao, P.; Rouse, C.; Chen, Y.; Dowling, J.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J. CBTRUS Statistical Report: Primary Brain and Central Nervous System Tumors Diagnosed in the United States in 2007–2011. Neuro-Oncology 2014, 16, iv1–iv63. [Google Scholar] [CrossRef] [PubMed]
- Koshy, M.; Villano, J.L.; Dolecek, T.A.; Howard, A.; Mahmood, U.; Chmura, S.J.; Weichselbaum, R.R.; McCarthy, B.J. Improved Survival Time Trends for Glioblastoma Using the SEER 17 Population-Based Registries. J. Neuro-Oncol. 2012, 107, 207–212. [Google Scholar] [CrossRef]
- Wen, P.Y.; Kesari, S. Malignant Gliomas in Adults. N. Engl. J. Med. 2008, 359, 492–507. [Google Scholar] [CrossRef] [PubMed]
- Ostrom, Q.T.; Truitt, G.; Gittleman, H.; Brat, D.J.; Kruchko, C.; Wilson, R.; Barnholtz-Sloan, J.S. Relative Survival after Diagnosis with a Primary Brain or Other Central Nervous System Tumor in the National Program of Cancer Registries, 2004 to 2014. Neuro-Oncol. Pract. 2020, 7, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Tan, A.C.; Ashley, D.M.; López, G.Y.; Malinzak, M.; Friedman, H.S.; Khasraw, M. Management of Glioblastoma: State of the Art and Future Directions. CA Cancer J. Clin. 2020, 70, 299–312. [Google Scholar] [CrossRef]
- Nabors, L.B.; Portnow, J.; Ahluwalia, M.; Baehring, J.; Brem, H.; Brem, S.; Butowski, N.; Campian, J.L.; Clark, S.W.; Fabiano, A.J. Central Nervous System Cancers, Version 3.2020, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. JNCCN 2020, 18, 1537–1570. [Google Scholar] [CrossRef]
- Davis, M.E. Glioblastoma: Overview of Disease and Treatment. Clin. J. Oncol. Nurs. 2016, 20, S2–S8. [Google Scholar] [CrossRef]
- Audureau, E.; Chivet, A.; Ursu, R.; Corns, R.; Metellus, P.; Noel, G.; Zouaoui, S.; Guyotat, J.; Le Reste, P.-J.; Faillot, T.; et al. Prognostic Factors for Survival in Adult Patients with Recurrent Glioblastoma: A Decision-Tree-Based Model. J. Neuro-Oncol. 2018, 136, 565–576. [Google Scholar] [CrossRef]
- Qazi, M.A.; Vora, P.; Venugopal, C.; Sidhu, S.S.; Moffat, J.; Swanton, C.; Singh, S.K. Intratumoral Heterogeneity: Pathways to Treatment Resistance and Relapse in Human Glioblastoma. Ann. Oncol. 2017, 28, 1448–1456. [Google Scholar] [CrossRef]
- Tirosh, I.; Suvà, M.L. Tackling the Many Facets of Glioblastoma Heterogeneity. Cell Stem Cell 2020, 26, 303–304. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO Classification of Tumours of the Central Nervous System. Acta Neuropathol. 2007, 114, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Ou, A.; Yung, W.K.A.; Majd, N. Molecular Mechanisms of Treatment Resistance in Glioblastoma. Int. J. Mol. Sci. 2020, 22, 351. [Google Scholar] [CrossRef] [PubMed]
- Vollmann-Zwerenz, A.; Leidgens, V.; Feliciello, G.; Klein, C.A.; Hau, P. Tumor Cell Invasion in Glioblastoma. Int. J. Mol. Sci. 2020, 21, 1932. [Google Scholar] [CrossRef] [PubMed]
- Roth, P. The Role of Integrins in Glioma Biology and Anti-Glioma Therapies. SpringerPlus 2015, 4, L12. [Google Scholar] [CrossRef][Green Version]
- Takada, Y.; Ye, X.; Simon, S. The Integrins. Genome Biol. 2007, 8, 215. [Google Scholar] [CrossRef]
- Schnell, O.; Krebs, B.; Wagner, E.; Romagna, A.; Beer, A.J.; Grau, S.J.; Thon, N.; Goetz, C.; Kretzschmar, H.A.; Tonn, J.; et al. Expression of Integrin Avβ3 in Gliomas Correlates with Tumor Grade and Is Not Restricted to Tumor Vasculature. Brain Pathol. 2008, 18, 378–386. [Google Scholar] [CrossRef]
- Gladson, C.L. Expression of Integrin Alpha v Beta 3 in Small Blood Vessels of Glioblastoma Tumors. J. Neuropathol. Exp. Neurol. 1996, 55, 1143–1149. [Google Scholar] [CrossRef]
- Bello, L.; Francolini, M.; Marthyn, P.; Zhang, J.; Carroll, R.S.; Nikas, D.C.; Strasser, J.F.; Villani, R.; Cheresh, D.A.; Black, P.M. Alpha(v)Beta3 and Alpha(v)Beta5 Integrin Expression in Glioma Periphery. Neurosurgery 2001, 49, 380–389; discussion 390. [Google Scholar] [CrossRef]
- Lorger, M.; Krueger, J.S.; O’Neal, M.; Staflin, K.; Felding-Habermann, B. Activation of Tumor Cell Integrin Avβ3 Controls Angiogenesis and Metastatic Growth in the Brain. Proc. Natl. Acad. Sci. USA 2009, 106, 10666–10671. [Google Scholar] [CrossRef]
- Gingras, M.C.; Roussel, E.; Bruner, J.M.; Branch, C.D.; Moser, R.P. Comparison of Cell Adhesion Molecule Expression between Glioblastoma Multiforme and Autologous Normal Brain Tissue. J. Neuroimmunol. 1995, 57, 143–153. [Google Scholar] [CrossRef]
- Raab-Westphal, S.; Marshall, J.F.; Goodman, S.L. Integrins as Therapeutic Targets: Successes and Cancers. Cancers 2017, 9, 110. [Google Scholar] [CrossRef] [PubMed]
- Beer, A.J.; Schwaiger, M. Imaging of Integrin Alphavbeta3 Expression. Cancer Metastasis Rev. 2008, 27, 631–644. [Google Scholar] [CrossRef] [PubMed]
- Gomes Marin, J.F.; Nunes, R.F.; Coutinho, A.M.; Zaniboni, E.C.; Costa, L.B.; Barbosa, F.G.; Queiroz, M.A.; Cerri, G.G.; Buchpiguel, C.A. Theranostics in Nuclear Medicine: Emerging and Re-Emerging Integrated Imaging and Therapies in the Era of Precision Oncology. RadioGraphics 2020, 40, 1715–1740. [Google Scholar] [CrossRef]
- Niu, G.; Chen, X. Why Integrin as a Primary Target for Imaging and Therapy. Theranostics 2011, 1, 30–47. [Google Scholar] [CrossRef]
- Tamkun, J.W.; DeSimone, D.W.; Fonda, D.; Patel, R.S.; Buck, C.; Horwitz, A.F.; Hynes, R.O. Structure of Integrin, a Glycoprotein Involved in the Transmembrane Linkage between Fibronectin and Actin. Cell 1986, 46, 271–282. [Google Scholar] [CrossRef]
- Geiger, B.; Bershadsky, A.; Pankov, R.; Yamada, K.M. Transmembrane Crosstalk between the Extracellular Matrix and the Cytoskeleton. Nat. Rev. Mol. Cell Biol. 2001, 2, 793–805. [Google Scholar] [CrossRef]
- Munson, J.; Bonner, M.; Fried, L.; Hofmekler, J.; Arbiser, J.; Bellamkonda, R. Identifying New Small Molecule Anti-Invasive Compounds for Glioma Treatment. Cell Cycle 2013, 12, 2200–2209. [Google Scholar] [CrossRef]
- Zhao, X.; Guan, J.-L. Focal Adhesion Kinase and Its Signaling Pathways in Cell Migration and Angiogenesis. Adv. Drug Deliv. Rev. 2011, 63, 610–615. [Google Scholar] [CrossRef]
- Seker-Polat, F.; Pinarbasi Degirmenci, N.; Solaroglu, I.; Bagci-Onder, T. Tumor Cell Infiltration into the Brain in Glioblastoma: From Mechanisms to Clinical Perspectives. Cancers 2022, 14, 443. [Google Scholar] [CrossRef]
- Owens, L.V.; Xu, L.; Craven, R.J.; Dent, G.A.; Weiner, T.M.; Kornberg, L.; Liu, E.T.; Cance, W.G. Overexpression of the Focal Adhesion Kinase (P125FAK) in Invasive Human Tumors. Cancer Res. 1995, 55, 2752–2755. [Google Scholar] [PubMed]
- Zagzag, D.; Friedlander, D.R.; Margolis, B.; Grumet, M.; Semenza, G.L.; Zhong, H.; Simons, J.W.; Holash, J.; Wiegand, S.J.; Yancopoulos, G.D. Molecular Events Implicated in Brain Tumor Angiogenesis and Invasion. Pediatr. Neurosurg. 2000, 33, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Legate, K.R.; Wickström, S.A.; Fässler, R. Genetic and Cell Biological Analysis of Integrin Outside-in Signaling. Genes Dev. 2009, 23, 397–418. [Google Scholar] [CrossRef] [PubMed]
- Ellert-Miklaszewska, A.; Poleszak, K.; Pasierbinska, M.; Kaminska, B. Integrin Signaling in Glioma Pathogenesis: From Biology to Therapy. Int. J. Mol. Sci. 2020, 21, 888. [Google Scholar] [CrossRef] [PubMed]
- Danen, E.H.J. Integrin Signaling as a Cancer Drug Target. ISRN Cell Biol. 2013, 2013, e135164. [Google Scholar] [CrossRef]
- Delamarre, E.; Taboubi, S.; Mathieu, S.; Bérenguer, C.; Rigot, V.; Lissitzky, J.-C.; Figarella-Branger, D.; Ouafik, L.; Luis, J. Expression of Integrin Alpha6beta1 Enhances Tumorigenesis in Glioma Cells. Am. J. Pathol. 2009, 175, 844–855. [Google Scholar] [CrossRef]
- Riemenschneider, M.J.; Mueller, W.; Betensky, R.A.; Mohapatra, G.; Louis, D.N. In Situ Analysis of Integrin and Growth Factor Receptor Signaling Pathways in Human Glioblastomas Suggests Overlapping Relationships with Focal Adhesion Kinase Activation. Am. J. Pathol. 2005, 167, 1379–1387. [Google Scholar] [CrossRef]
- Ducassou, A.; Uro-Coste, E.; Verrelle, P.; Filleron, T.; Benouaich-Amiel, A.; Lubrano, V.; Sol, J.-C.; Delisle, M.-B.; Favre, G.; Ken, S.; et al. Avβ3 Integrin and Fibroblast Growth Factor Receptor 1 (FGFR1): Prognostic Factors in a Phase I-II Clinical Trial Associating Continuous Administration of Tipifarnib with Radiotherapy for Patients with Newly Diagnosed Glioblastoma. Eur. J. Cancer Oxf. Engl. 2013, 49, 2161–2169. [Google Scholar] [CrossRef]
- Brooks, P.C.; Clark, R.A.; Cheresh, D.A. Requirement of Vascular Integrin Alpha v Beta 3 for Angiogenesis. Science 1994, 264, 569–571. [Google Scholar] [CrossRef]
- Gladson, C.L.; Cheresh, D.A. Glioblastoma Expression of Vitronectin and the Alpha v Beta 3 Integrin. Adhesion Mechanism for Transformed Glial Cells. J. Clin. Investig. 1991, 88, 1924–1932. [Google Scholar] [CrossRef]
- Serres, E.; Debarbieux, F.; Stanchi, F.; Maggiorella, L.; Grall, D.; Turchi, L.; Burel-Vandenbos, F.; Figarella-Branger, D.; Virolle, T.; Rougon, G.; et al. Fibronectin Expression in Glioblastomas Promotes Cell Cohesion, Collective Invasion of Basement Membrane in Vitro and Orthotopic Tumor Growth in Mice. Oncogene 2014, 33, 3451–3462. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-H.; Lu, C.; Sun, J.; Chen, X.-D.; Dai, J.-X.; Cai, J.-Y.; Chen, X.-L. Diagnostic and Prognostic Value of Serum Vitronectin Levels in Human Glioma. J. Neurol. Sci. 2016, 371, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Schittenhelm, J.; Schwab, E.I.; Sperveslage, J.; Tatagiba, M.; Meyermann, R.; Fend, F.; Goodman, S.L.; Sipos, B. Longitudinal Expression Analysis of Av Integrins in Human Gliomas Reveals Upregulation of Integrin Avβ3 as a Negative Prognostic Factor. J. Neuropathol. Exp. Neurol. 2013, 72, 194–210. [Google Scholar] [CrossRef] [PubMed]
- Demircioglu, F.; Hodivala-Dilke, K. Avβ3 Integrin and Tumour Blood Vessels-Learning from the Past to Shape the Future. Curr. Opin. Cell Biol. 2016, 42, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Bussolati, B.; Deambrosis, I.; Russo, S.; Deregibus, M.C.; Camussi, G. Altered Angiogenesis and Survival in Human Tumor-Derived Endothelial Cells. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2003, 17, 1159–1161. [Google Scholar] [CrossRef]
- Avraamides, C.J.; Garmy-Susini, B.; Varner, J.A. Integrins in Angiogenesis and Lymphangiogenesis. Nat. Rev. Cancer 2008, 8, 604–617. [Google Scholar] [CrossRef]
- Brooks, P.C.; Montgomery, A.M.; Rosenfeld, M.; Reisfeld, R.A.; Hu, T.; Klier, G.; Cheresh, D.A. Integrin Alpha v Beta 3 Antagonists Promote Tumor Regression by Inducing Apoptosis of Angiogenic Blood Vessels. Cell 1994, 79, 1157–1164. [Google Scholar] [CrossRef]
- Nisato, R.E.; Tille, J.-C.; Jonczyk, A.; Goodman, S.L.; Pepper, M.S. Alphav Beta 3 and Alphav Beta 5 Integrin Antagonists Inhibit Angiogenesis in Vitro. Angiogenesis 2003, 6, 105–119. [Google Scholar] [CrossRef]
- Soldi, R.; Mitola, S.; Strasly, M.; Defilippi, P.; Tarone, G.; Bussolino, F. Role of Alphavbeta3 Integrin in the Activation of Vascular Endothelial Growth Factor Receptor-2. EMBO J. 1999, 18, 882–892. [Google Scholar] [CrossRef]
- Cui, X.; Morales, R.-T.T.; Qian, W.; Wang, H.; Gagner, J.-P.; Dolgalev, I.; Placantonakis, D.; Zagzag, D.; Cimmino, L.; Snuderl, M.; et al. Hacking Macrophage-Associated Immunosuppression for Regulating Glioblastoma Angiogenesis. Biomaterials 2018, 161, 164–178. [Google Scholar] [CrossRef]
- Steri, V.; Ellison, T.S.; Gontarczyk, A.M.; Weilbaecher, K.; Schneider, J.G.; Edwards, D.; Fruttiger, M.; Hodivala-Dilke, K.M.; Robinson, S.D. Acute Depletion of Endothelial Β3-Integrin Transiently Inhibits Tumor Growth and Angiogenesis in Mice. Circ. Res. 2014, 114, 79–91. [Google Scholar] [CrossRef] [PubMed]
- D’Abaco, G.M.; Kaye, A.H. Integrins: Molecular Determinants of Glioma Invasion. J. Clin. Neurosci. 2007, 14, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Conte, M.; Casas-Tintò, S.; Soler, J. Modeling Invasion Patterns in the Glioblastoma Battlefield. PLOS Comput. Biol. 2021, 17, e1008632. [Google Scholar] [CrossRef] [PubMed]
- Paulus, W.; Tonn, J.C. Basement Membrane Invasion of Glioma Cells Mediated by Integrin Receptors. J. Neurosurg. 1994, 80, 515–519. [Google Scholar] [CrossRef]
- Lal, A.; Lash, A.E.; Altschul, S.F.; Velculescu, V.; Zhang, L.; McLendon, R.E.; Marra, M.A.; Prange, C.; Morin, P.J.; Polyak, K.; et al. A Public Database for Gene Expression in Human Cancers. Cancer Res. 1999, 59, 5403–5407. [Google Scholar]
- Colin, C.; Baeza, N.; Bartoli, C.; Fina, F.; Eudes, N.; Nanni, I.; Martin, P.-M.; Ouafik, L.; Figarella-Branger, D. Identification of Genes Differentially Expressed in Glioblastoma versus Pilocytic Astrocytoma Using Suppression Subtractive Hybridization. Oncogene 2006, 25, 2818–2826. [Google Scholar] [CrossRef]
- Mahesparan, R.; Read, T.-A.; Lund-Johansen, M.; Skaftnesmo, K.O.; Bjerkvig, R.; Engebraaten, O. Expression of Extracellular Matrix Components in a Highly Infiltrative in Vivo Glioma Model. Acta Neuropathol. 2003, 105, 49–57. [Google Scholar] [CrossRef]
- Ohnishi, T.; Hiraga, S.; Izumoto, S.; Matsumura, H.; Kanemura, Y.; Arita, N.; Hayakawa, T. Role of Fibronectin-Stimulated Tumor Cell Migration in Glioma Invasion in Vivo: Clinical Significance of Fibronectin and Fibronectin Receptor Expressed in Human Glioma Tissues. Clin. Exp. Metastasis 1998, 16, 729–741. [Google Scholar] [CrossRef]
- Fabian, C.; Han, M.; Bjerkvig, R.; Niclou, S.P. Novel Facets of Glioma Invasion. Int. Rev. Cell Mol. Biol. 2021, 360, 33–64. [Google Scholar] [CrossRef]
- Hagemann, C.; Anacker, J.; Ernestus, R.-I.; Vince, G.H. A Complete Compilation of Matrix Metalloproteinase Expression in Human Malignant Gliomas. World J. Clin. Oncol. 2012, 3, 67–79. [Google Scholar] [CrossRef]
- Yosef, G.; Arkadash, V.; Papo, N. Targeting the MMP-14/MMP-2/Integrin Avβ3 Axis with Multispecific N-TIMP2-Based Antagonists for Cancer Therapy. J. Biol. Chem. 2018, 293, 13310–13326. [Google Scholar] [CrossRef] [PubMed]
- Kesanakurti, D.; Chetty, C.; Rajasekhar Maddirela, D.; Gujrati, M.; Rao, J.S. Functional Cooperativity by Direct Interaction between PAK4 and MMP-2 in the Regulation of Anoikis Resistance, Migration and Invasion in Glioma. Cell Death Dis. 2012, 3, e445. [Google Scholar] [CrossRef] [PubMed]
- Osuka, S.; Meir, E.G.V. Overcoming Therapeutic Resistance in Glioblastoma: The Way Forward. J. Clin. Investig. 2017, 127, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Dymova, M.A.; Kuligina, E.V.; Richter, V.A. Molecular Mechanisms of Drug Resistance in Glioblastoma. Int. J. Mol. Sci. 2021, 22, 6385. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Xiao, W.; Sun, S.; Sohrabi, A.; Liang, J.; Seidlits, S.K. Extracellular Matrix Proteins Confer Cell Adhesion-Mediated Drug Resistance Through Integrin Av in Glioblastoma Cells. Front. Cell Dev. Biol. 2021, 9, 616580. [Google Scholar] [CrossRef] [PubMed]
- Ponomarev, A.; Gilazieva, Z.; Solovyeva, V.; Allegrucci, C.; Rizvanov, A. Intrinsic and Extrinsic Factors Impacting Cancer Stemness and Tumor Progression. Cancers 2022, 14, 970. [Google Scholar] [CrossRef]
- Sattiraju, A.; Sai, K.K.S.; Mintz, A. Glioblastoma Stem Cells and Their Microenvironment. Adv. Exp. Med. Biol. 2017, 1041, 119–140. [Google Scholar] [CrossRef]
- Filatova, A.; Acker, T.; Garvalov, B.K. The Cancer Stem Cell Niche(s): The Crosstalk between Glioma Stem Cells and Their Microenvironment. Biochim. Biophys. Acta 2013, 1830, 2496–2508. [Google Scholar] [CrossRef]
- Yu, Q.; Xue, Y.; Liu, J.; Xi, Z.; Li, Z.; Liu, Y. Fibronectin Promotes the Malignancy of Glioma Stem-Like Cells Via Modulation of Cell Adhesion, Differentiation, Proliferation and Chemoresistance. Front. Mol. Neurosci. 2018, 11, 130. [Google Scholar] [CrossRef]
- Deininger, M.H.; Pater, S.; Strik, H.; Meyermann, R. Macrophage/Microglial Cell Subpopulations in Glioblastoma Multiforme Relapses Are Differentially Altered by Radiochemotherapy. J. Neurooncol. 2001, 55, 141–147. [Google Scholar] [CrossRef]
- Zhou, W.; Ke, S.Q.; Huang, Z.; Flavahan, W.; Fang, X.; Paul, J.; Wu, L.; Sloan, A.E.; McLendon, R.E.; Li, X.; et al. Periostin Secreted by Glioblastoma Stem Cells Recruits M2 Tumour-Associated Macrophages and Promotes Malignant Growth. Nat. Cell Biol. 2015, 17, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Seguin, L.; Kato, S.; Franovic, A.; Camargo, M.F.; Lesperance, J.; Elliott, K.C.; Yebra, M.; Mielgo, A.; Lowy, A.M.; Husain, H.; et al. An Integrin Β3;-KRAS-RalB Complex Drives Tumour Stemness and Resistance to EGFR Inhibition. Nat. Cell Biol. 2014, 16, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Visvader, J.E.; Stingl, J. Mammary Stem Cells and the Differentiation Hierarchy: Current Status and Perspectives. Genes Dev. 2014, 28, 1143–1158. [Google Scholar] [CrossRef]
- Monferran, S.; Skuli, N.; Delmas, C.; Favre, G.; Bonnet, J.; Cohen-Jonathan-Moyal, E.; Toulas, C. Alphavbeta3 and Alphavbeta5 Integrins Control Glioma Cell Response to Ionising Radiation through ILK and RhoB. Int. J. Cancer 2008, 123, 357–364. [Google Scholar] [CrossRef]
- Mikkelsen, T.; Brodie, C.; Finniss, S.; Berens, M.E.; Rennert, J.L.; Nelson, K.; Lemke, N.; Brown, S.L.; Hahn, D.; Neuteboom, B.; et al. Radiation Sensitization of Glioblastoma by Cilengitide Has Unanticipated Schedule-Dependency. Int. J. Cancer 2009, 124, 2719–2727. [Google Scholar] [CrossRef]
- Christmann, M.; Diesler, K.; Majhen, D.; Steigerwald, C.; Berte, N.; Freund, H.; Stojanović, N.; Kaina, B.; Osmak, M.; Ambriović-Ristov, A.; et al. Integrin AVβ3 Silencing Sensitizes Malignant Glioma Cells to Temozolomide by Suppression of Homologous Recombination Repair. Oncotarget 2017, 8, 27754–27771. [Google Scholar] [CrossRef] [PubMed]
- Skuli, N.; Monferran, S.; Delmas, C.; Favre, G.; Bonnet, J.; Toulas, C.; Cohen-Jonathan Moyal, E. Alphavbeta3/Alphavbeta5 Integrins-FAK-RhoB: A Novel Pathway for Hypoxia Regulation in Glioblastoma. Cancer Res. 2009, 69, 3308–3316. [Google Scholar] [CrossRef] [PubMed]
- Maurer, G.D.; Tritschler, I.; Adams, B.; Tabatabai, G.; Wick, W.; Stupp, R.; Weller, M. Cilengitide Modulates Attachment and Viability of Human Glioma Cells, but Not Sensitivity to Irradiation or Temozolomide in Vitro. Neuro-Oncolog 2009, 11, 747–756. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Lee, J.K.; Kim, B.; DeWitt, J.P.; Lee, J.E.; Han, J.H.; Kim, S.-K.; Oh, C.W.; Kim, C.-Y. Combination Therapy of Cilengitide with Belotecan against Experimental Glioblastoma. Int. J. Cancer 2013, 133, 749–756. [Google Scholar] [CrossRef]
- Yamada, S.; Bu, X.-Y.; Khankaldyyan, V.; Gonzales-Gomez, I.; McComb, J.G.; Laug, W.E. Effect of the Angiogenesis Inhibitor Cilengitide (EMD 121974) on Glioblastoma Growth in Nude Mice. Neurosurgery 2006, 59, 1304–1312; discussion 1312. [Google Scholar] [CrossRef]
- Stupp, R.; Hegi, M.E.; Neyns, B.; Goldbrunner, R.; Schlegel, U.; Clement, P.M.J.; Grabenbauer, G.G.; Ochsenbein, A.F.; Simon, M.; Dietrich, P.-Y.; et al. Phase I/IIa Study of Cilengitide and Temozolomide with Concomitant Radiotherapy Followed by Cilengitide and Temozolomide Maintenance Therapy in Patients with Newly Diagnosed Glioblastoma. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2010, 28, 2712–2718. [Google Scholar] [CrossRef]
- Nabors, L.B.; Mikkelsen, T.; Hegi, M.E.; Ye, X.; Batchelor, T.; Lesser, G.; Peereboom, D.; Rosenfeld, M.R.; Olsen, J.; Brem, S.; et al. A Safety Run-in and Randomized Phase II Study of Cilengitide Combined with Chemoradiation for Newly Diagnosed Glioblastoma (NABTT 0306). Cancer 2012, 118, 5601–5607. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.; Nabors, L.B.; Gorlia, T.; Leske, H.; Rushing, E.; Bady, P.; Hicking, C.; Perry, J.; Hong, Y.-K.; Roth, P.; et al. Cilengitide in Newly Diagnosed Glioblastoma: Biomarker Expression and Outcome. Oncotarget 2016, 7, 15018–15032. [Google Scholar] [CrossRef] [PubMed]
- Chinot, O.L. Cilengitide in Glioblastoma: When Did It Fail? Lancet Oncol. 2014, 15, 1044–1045. [Google Scholar] [CrossRef]
- Reynolds, A.R.; Hart, I.R.; Watson, A.R.; Welti, J.C.; Silva, R.G.; Robinson, S.D.; Da Violante, G.; Gourlaouen, M.; Salih, M.; Jones, M.C.; et al. Stimulation of Tumor Growth and Angiogenesis by Low Concentrations of RGD-Mimetic Integrin Inhibitors. Nat. Med. 2009, 15, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Silginer, M.; Weller, M.; Ziegler, U.; Roth, P. Integrin Inhibition Promotes Atypical Anoikis in Glioma Cells. Cell Death Dis. 2014, 5, e1012. [Google Scholar] [CrossRef]
- Cirkel, G.A.; Kerklaan, B.M.; Vanhoutte, F.; Van der Aa, A.; Lorenzon, G.; Namour, F.; Pujuguet, P.; Darquenne, S.; de Vos, F.Y.F.; Snijders, T.J.; et al. A Dose Escalating Phase I Study of GLPG0187, a Broad Spectrum Integrin Receptor Antagonist, in Adult Patients with Progressive High-Grade Glioma and Other Advanced Solid Malignancies. Invest. New Drugs 2016, 34, 184–192. [Google Scholar] [CrossRef]
- Battistini, L.; Bugatti, K.; Sartori, A.; Curti, C.; Zanardi, F. RGD Peptide-Drug Conjugates as Effective Dual Targeting Platforms: Recent Advances. Eur. J. Org. Chem. 2021, 2021, 2506–2528. [Google Scholar] [CrossRef]
- Chen, Z.; Deng, J.; Zhao, Y.; Tao, T. Cyclic RGD Peptide-Modified Liposomal Drug Delivery System: Enhanced Cellular Uptake in Vitro and Improved Pharmacokinetics in Rats. Int. J. Nanomedicine 2012, 7, 3803–3811. [Google Scholar] [CrossRef]
- Katsamakas, S.; Chatzisideri, T.; Thysiadis, S.; Sarli, V. RGD-Mediated Delivery of Small-Molecule Drugs. Future Med. Chem. 2017, 9, 579–604. [Google Scholar] [CrossRef]
- Zhan, C.; Gu, B.; Xie, C.; Li, J.; Liu, Y.; Lu, W. Cyclic RGD Conjugated Poly(Ethylene Glycol)-Co-Poly(Lactic Acid) Micelle Enhances Paclitaxel Anti-Glioblastoma Effect. J. Control. Release Off. J. Control. Release Soc. 2010, 143, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Belhadj, Z.; Zhan, C.; Ying, M.; Wei, X.; Xie, C.; Yan, Z.; Lu, W. Multifunctional Targeted Liposomal Drug Delivery for Efficient Glioblastoma Treatment. Oncotarget 2017, 8, 66889–66900. [Google Scholar] [CrossRef] [PubMed]
- McNerny, D.Q.; Kukowska-Latallo, J.F.; Mullen, D.G.; Wallace, J.M.; Desai, A.M.; Shukla, R.; Huang, B.; Banaszak Holl, M.M.; Baker, J.R. RGD Dendron Bodies; Synthetic Avidity Agents with Defined and Potentially Interchangeable Effector Sites That Can Substitute for Antibodies. Bioconjug. Chem. 2009, 20, 1853–1859. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Liu, W.; Gao, F.; Fang, X.; Chen, Y. C(RGDyK)-Decorated Pluronic Micelles for Enhanced Doxorubicin and Paclitaxel Delivery to Brain Glioma. Int. J. Nanomedicine 2016, 11, 1629–1641. [Google Scholar] [CrossRef] [PubMed]
- Quader, S.; Liu, X.; Chen, Y.; Mi, P.; Chida, T.; Ishii, T.; Miura, Y.; Nishiyama, N.; Cabral, H.; Kataoka, K. CRGD Peptide-Installed Epirubicin-Loaded Polymeric Micelles for Effective Targeted Therapy against Brain Tumors. J. Control. Release Off. J. Control. Release Soc. 2017, 258, 56–66. [Google Scholar] [CrossRef]
- Schnell, O.; Krebs, B.; Carlsen, J.; Miederer, I.; Goetz, C.; Goldbrunner, R.H.; Wester, H.-J.; Haubner, R.; Pöpperl, G.; Holtmannspötter, M.; et al. Imaging of Integrin Avβ3 Expression in Patients with Malignant Glioma by [18F] Galacto-RGD Positron Emission Tomography. Neuro-Oncology 2009, 11, 861–870. [Google Scholar] [CrossRef]
- Iagaru, A.; Mosci, C.; Mittra, E.; Zaharchuk, G.; Fischbein, N.; Harsh, G.; Li, G.; Nagpal, S.; Recht, L.; Gambhir, S.S. Glioblastoma Multiforme Recurrence: An Exploratory Study of (18)F FPPRGD2 PET/CT. Radiology 2015, 277, 497–506. [Google Scholar] [CrossRef]
- Li, D.; Zhao, X.; Zhang, L.; Li, F.; Ji, N.; Gao, Z.; Wang, J.; Kang, P.; Liu, Z.; Shi, J.; et al. 68Ga-PRGD2 PET/CT in the Evaluation of Glioma: A Prospective Study. Mol. Pharm. 2014, 11, 3923–3929. [Google Scholar] [CrossRef]
- Zhang, J.; Li, D.; Niu, G.; Baum, R.; Zhu, Z.; Chen, X. First-in-Human Study of A64Cu-Labeled Long-Acting Integrin Avβ3 Targeting Molecule64Cu-NOTA-EB-RGD in Healthy Volunteers and GBM Patients. J. Nucl. Med. 2020, 61, 349. [Google Scholar]
- Novy, Z.; Stepankova, J.; Hola, M.; Flasarova, D.; Popper, M.; Petrik, M. Preclinical Evaluation of Radiolabeled Peptides for PET Imaging of Glioblastoma Multiforme. Molecules 2019, 14, 2496. [Google Scholar] [CrossRef]
- Lo, W.-L.; Lo, S.-W.; Chen, S.-J.; Chen, M.-W.; Huang, Y.-R.; Chen, L.-C.; Chang, C.-H.; Li, M.-H. Molecular Imaging and Preclinical Studies of Radiolabeled Long-Term RGD Peptides in U-87 MG Tumor-Bearing Mice. Int. J. Mol. Sci. 2021, 22, 5459. [Google Scholar] [CrossRef]
- Shi, J.; Fan, D.; Dong, C.; Liu, H.; Jia, B.; Zhao, H.; Jin, X.; Liu, Z.; Li, F.; Wang, F. Anti-Tumor Effect of Integrin Targeted (177)Lu-3PRGD2 and Combined Therapy with Endostar. Theranostics 2014, 4, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Chen, H.; Guo, Z.; Fu, K.; Yao, L.; Fu, L.; Guo, W.; Wen, X.; Jacobson, O.; Zhang, X.; et al. Targeted Radionuclide Therapy in Patient-Derived Xenografts Using 177Lu-EB-RGD. Mol. Cancer Ther. 2020, 19, 2034–2043. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Echavidre, W.; Picco, V.; Faraggi, M.; Montemagno, C. Integrin-αvβ3 as a Therapeutic Target in Glioblastoma: Back to the Future? Pharmaceutics 2022, 14, 1053. https://doi.org/10.3390/pharmaceutics14051053
Echavidre W, Picco V, Faraggi M, Montemagno C. Integrin-αvβ3 as a Therapeutic Target in Glioblastoma: Back to the Future? Pharmaceutics. 2022; 14(5):1053. https://doi.org/10.3390/pharmaceutics14051053
Chicago/Turabian StyleEchavidre, William, Vincent Picco, Marc Faraggi, and Christopher Montemagno. 2022. "Integrin-αvβ3 as a Therapeutic Target in Glioblastoma: Back to the Future?" Pharmaceutics 14, no. 5: 1053. https://doi.org/10.3390/pharmaceutics14051053
APA StyleEchavidre, W., Picco, V., Faraggi, M., & Montemagno, C. (2022). Integrin-αvβ3 as a Therapeutic Target in Glioblastoma: Back to the Future? Pharmaceutics, 14(5), 1053. https://doi.org/10.3390/pharmaceutics14051053