
Recent Advances in the Treatment of Alzheimer’s Disease Using Nanoparticle-Based Drug Delivery Systems

Abstract
:1. Introduction
2. AD Pathology
2.1. Cholinergic Deficit
2.2. Amyloid Hypothesis
2.3. Tau Hypothesis
2.4. Oxidative Stress
2.5. Cellular and Vascular Dysfunction
2.6. Cholesterol
2.7. Inflammation
2.8. Metal Imbalance
3. Commercially Available Medicines and Those in Pipelines
4. BBB Physiology and Structure
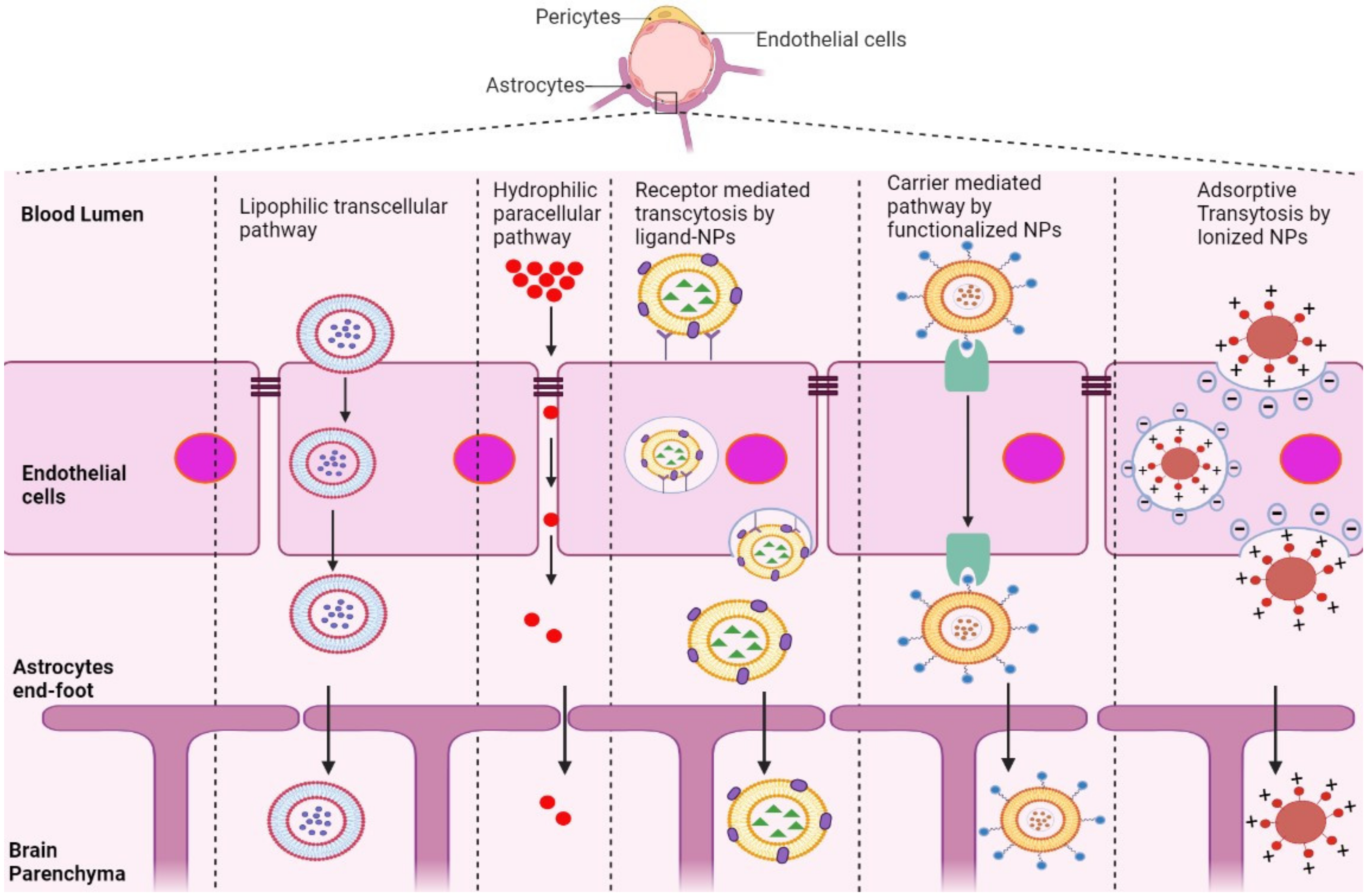
5. Strategies to Overcome the BBB
5.1. Crossing the BBB
5.2. Avoiding the BBB
5.3. Disrupting the BBB
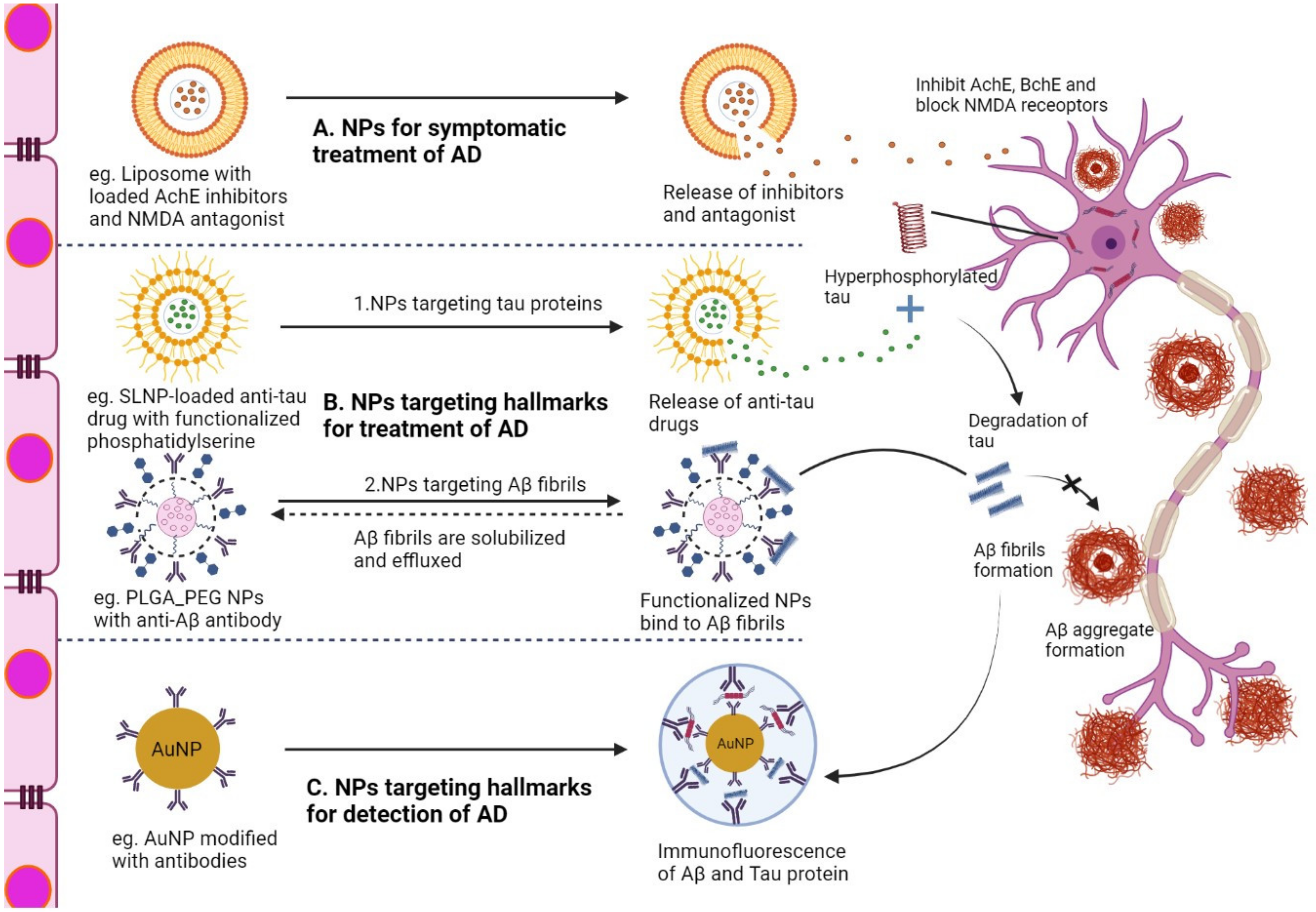
6. NP-Based Delivery System for the Treatment of AD
6.1. Liposomes
6.2. Micelles
6.3. Solid Lipid NPs (SLNs)
6.4. Polymeric NPs (PNPs)
6.5. Dendrimers
6.6. Nanoemulsions (NEs)
6.7. Inorganic NPs
7. Immunotherapy for AD
8. Future Perspectives and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cummings, J.; Lee, G.; Zhong, K.; Fonseca, J.; Taghva, K. Alzheimer’s disease drug development pipeline: 2021. Alzheimers Dement. Transl. Res. Clin. Interv. 2021, 7, e12179. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s Association. 2022 Alzheimer’s Disease Facts and Figures; Alzheimer’s Association: Chicago, IL, USA, 2022. [Google Scholar]
- Alzheimer’s Association. 2021 Alzheimer’s disease facts and figures. Alzheimers Dement. 2021, 17, 327–406. [Google Scholar] [CrossRef] [PubMed]
- Wong, W. Economic burden of Alzheimer disease and managed care considerations. Am. J. Manag. Care 2020, 26 (Suppl. 8), S177–S183. [Google Scholar] [PubMed]
- Yiannopoulou, K.G.; Papageorgiou, S.G. Current and future treatments for Alzheimer’s disease. Adv. Neurol. Disord. 2013, 6, 19–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevigny, J.; Chiao, P.; Bussière, T.; Weinreb, P.H.; Williams, L.; Maier, M.; Dunstan, R.; Salloway, S.; Chen, T.; Ling, Y.; et al. The antibody aducanumab reduces Aβ plaques in Alzheimer’s disease. Nature 2016, 537, 50–56. [Google Scholar] [CrossRef]
- Mehta, D.; Jackson, R.; Paul, G.; Shi, J.; Sabbagh, M. Why do trials for Alzheimer’s disease drugs keep failing? A discontinued drug perspective for 2010-2015. Expert. Opin. Investig. Drugs 2017, 26, 735–739. [Google Scholar] [CrossRef]
- Cai, Z.; Qiao, P.F.; Wan, C.Q.; Cai, M.; Zhou, N.K.; Li, Q. Role of Blood-Brain Barrier in Alzheimer’s Disease. J. Alzheimers Dis. 2018, 63, 1223–1234. [Google Scholar] [CrossRef]
- Teleanu, D.M.; Chircov, C.; Grumezescu, A.M.; Volceanov, A.; Teleanu, R.I. Blood-Brain Delivery Methods Using Nanotechnology. Pharmaceutics 2018, 10, 269. [Google Scholar] [CrossRef] [Green Version]
- Gelperina, S.; Kisich, K.; Iseman, M.D.; Heifets, L. The potential advantages of nanoparticle drug delivery systems in chemotherapy of tuberculosis. Am. J. Respir. Crit. Care Med. 2005, 172, 1487–1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allémann, E.; Leroux, J.C.; Gurny, R.; Doelker, E. In vitro extended-release properties of drug-loaded poly(DL-lactic acid) nanoparticles produced by a salting-out procedure. Pharm. Res. 1993, 10, 1732–1737. [Google Scholar] [CrossRef]
- Banerjee, T.; Mitra, S.; Kumar Singh, A.; Kumar Sharma, R.; Maitra, A. Preparation, characterization and biodistribution of ultrafine chitosan nanoparticles. Int. J. Pharm. 2002, 243, 93–105. [Google Scholar] [CrossRef]
- Topal, G.R.; Meszaros, M.; Porkolab, G.; Szecsko, A.; Polgar, T.F.; Siklos, L.; Deli, M.A.; Veszelka, S.; Bozkir, A. ApoE-Targeting Increases the Transfer of Solid Lipid Nanoparticles with Donepezil Cargo across a Culture Model of the Blood-Brain Barrier. Pharmaceutics 2020, 13, 38. [Google Scholar] [CrossRef] [PubMed]
- Nazıroğlu, M.; Muhamad, S.; Pecze, L. Nanoparticles as potential clinical therapeutic agents in Alzheimer’s disease: Focus on selenium nanoparticles. Expert. Rev. Clin. Pharm. 2017, 10, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Riaz, M.K.; Xie, Y.; Zhang, X.; Liu, Q.; Chen, H.; Bian, Z.; Chen, X.; Lu, A.; Yang, Z. Review of Current Strategies for Delivering Alzheimer’s Disease Drugs across the Blood-Brain Barrier. Int. J. Mol. Sci. 2019, 20, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahni, J.K.; Doggui, S.; Ali, J.; Baboota, S.; Dao, L.; Ramassamy, C. Neurotherapeutic applications of nanoparticles in Alzheimer’s disease. J. Control. Release 2011, 152, 208–231. [Google Scholar] [CrossRef] [PubMed]
- Hroudová, J.; Singh, N.; Fisar, Z.; Ghosh, K. Progress in drug development for Alzheimer’s disease: An overview in relation to mitochondrial energy metabolism. Eur. J. Med. Chem. 2016, 121, 774–784. [Google Scholar] [CrossRef]
- Frölich, L. The cholinergic pathology in Alzheimer’s disease--discrepancies between clinical experience and pathophysiological findings. J. Neural. Transm. 2002, 109, 1003–1013. [Google Scholar] [CrossRef]
- Terry, A.V., Jr.; Buccafusco, J.J. The cholinergic hypothesis of age and Alzheimer’s disease-related cognitive deficits: Recent challenges and their implications for novel drug development. J. Pharm. Exp. 2003, 306, 821–827. [Google Scholar] [CrossRef]
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 2018, 141, 1917–1933. [Google Scholar] [CrossRef]
- Whitehouse, P.J. The cholinergic deficit in Alzheimer’s disease. J. Clin. Psychiatry 1998, 59 (Suppl. 13), 19–22. [Google Scholar]
- DeTure, M.A.; Dickson, D.W. The neuropathological diagnosis of Alzheimer’s disease. Mol. Neurodegener. 2019, 14, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, R.J.; Wong, P.C. Amyloid precursor protein processing and Alzheimer’s disease. Annu. Rev. Neurosci. 2011, 34, 185–204. [Google Scholar] [CrossRef] [Green Version]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Errico, P.; Meyer-Luehmann, M. Mechanisms of Pathogenic Tau and Aβ Protein Spreading in Alzheimer’s Disease. Front. Aging Neurosci. 2020, 12, 265. [Google Scholar] [CrossRef] [PubMed]
- Bloom, G.S. Amyloid-β and tau: The trigger and bullet in Alzheimer disease pathogenesis. JAMA Neurol. 2014, 71, 505–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.H.; Bastianetto, S.; Mennicken, F.; Ma, W.; Kar, S. Amyloid beta peptide induces tau phosphorylation and loss of cholinergic neurons in rat primary septal cultures. Neuroscience 2002, 115, 201–211. [Google Scholar] [CrossRef]
- Méphon-Gaspard, A.; Boca, M.; Pioche-Durieu, C.; Desforges, B.; Burgo, A.; Hamon, L.; Piétrement, O.; Pastré, D. Role of tau in the spatial organization of axonal microtubules: Keeping parallel microtubules evenly distributed despite macromolecular crowding. Cell Mol. Life Sci. 2016, 73, 3745–3760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, A.; Reddy, P.H. Synaptic basis of Alzheimer’s disease: Focus on synaptic amyloid beta, P-tau and mitochondria. Ageing Res. Rev. 2021, 65, 101208. [Google Scholar] [CrossRef]
- Gong, C.X.; Iqbal, K. Hyperphosphorylation of microtubule-associated protein tau: A promising therapeutic target for Alzheimer disease. Curr. Med. Chem. 2008, 15, 2321–2328. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Latypova, X.; Wilson, C.M.; Magnaudeix, A.; Perrin, M.L.; Yardin, C.; Terro, F. Tau protein kinases: Involvement in Alzheimer’s disease. Ageing Res. Rev. 2013, 12, 289–309. [Google Scholar] [CrossRef]
- Arnsten, A.F.T.; Datta, D.; Del Tredici, K.; Braak, H. Hypothesis: Tau pathology is an initiating factor in sporadic Alzheimer’s disease. Alzheimers Dement. 2021, 17, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Mohandas, E.; Rajmohan, V.; Raghunath, B. Neurobiology of Alzheimer’s disease. Indian J. Psychiatry 2009, 51, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Reed, T.; Newman, S.F.; Sultana, R. Roles of amyloid beta-peptide-associated oxidative stress and brain protein modifications in the pathogenesis of Alzheimer’s disease and mild cognitive impairment. Free. Radic. Biol. Med. 2007, 43, 658–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butterfield, D.A.; Lauderback, C.M. Lipid peroxidation and protein oxidation in Alzheimer’s disease brain: Potential causes and consequences involving amyloid beta-peptide-associated free radical oxidative stress. Free. Radic. Biol. Med. 2002, 32, 1050–1060. [Google Scholar] [CrossRef]
- Cheignon, C.; Tomas, M.; Bonnefont-Rousselot, D.; Faller, P.; Hureau, C.; Collin, F. Oxidative stress and the amyloid beta peptide in Alzheimer’s disease. Redox. Biol. 2018, 14, 450–464. [Google Scholar] [CrossRef]
- Trevisan, K.; Cristina-Pereira, R.; Silva-Amaral, D.; Aversi-Ferreira, T.A. Theories of Aging and the Prevalence of Alzheimer’s Disease. Biomed. Res. Int. 2019, 2019, 9171424. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, R.A. Risk factors for Alzheimer’s disease. Folia Neuropathol. 2019, 57, 87–105. [Google Scholar] [CrossRef] [Green Version]
- Farkas, E.; Luiten, P.G. Cerebral microvascular pathology in aging and Alzheimer’s disease. Prog. Neurobiol. 2001, 64, 575–611. [Google Scholar] [CrossRef] [Green Version]
- Daulatzai, M.A. Cerebral hypoperfusion and glucose hypometabolism: Key pathophysiological modulators promote neurodegeneration, cognitive impairment, and Alzheimer’s disease. J. Neurosci. Res. 2017, 95, 943–972. [Google Scholar] [CrossRef]
- Mann, K.M.; Thorngate, F.E.; Katoh-Fukui, Y.; Hamanaka, H.; Williams, D.L.; Fujita, S.; Lamb, B.T. Independent effects of APOE on cholesterol metabolism and brain Abeta levels in an Alzheimer disease mouse model. Hum. Mol. Genet. 2004, 13, 1959–1968. [Google Scholar] [CrossRef] [Green Version]
- Fong, L.K.; Yang, M.M.; Dos Santos Chaves, R.; Reyna, S.M.; Langness, V.F.; Woodruff, G.; Roberts, E.A.; Young, J.E.; Goldstein, L.S.B. Full-length amyloid precursor protein regulates lipoprotein metabolism and amyloid-β clearance in human astrocytes. J. Biol. Chem. 2018, 293, 11341–11357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puglielli, L.; Tanzi, R.E.; Kovacs, D.M. Alzheimer’s disease: The cholesterol connection. Nat. Neurosci. 2003, 6, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Langness, V.F.; van der Kant, R.; Das, U.; Wang, L.; Chaves, R.D.S.; Goldstein, L.S.B. Cholesterol-lowering drugs reduce APP processing to Aβ by inducing APP dimerization. Mol. Biol. Cell 2021, 32, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Feringa, F.M.; van der Kant, R. Cholesterol and Alzheimer’s Disease; From Risk Genes to Pathological Effects. Front. Aging Neurosci. 2021, 13, 690372. [Google Scholar] [CrossRef]
- Tuppo, E.E.; Arias, H.R. The role of inflammation in Alzheimer’s disease. Int. J. Biochem. Cell Biol. 2005, 37, 289–305. [Google Scholar] [CrossRef]
- Moore, A.H.; O’Banion, M.K. Neuroinflammation and anti-inflammatory therapy for Alzheimer’s disease. Adv. Drug Deliv. Rev. 2002, 54, 1627–1656. [Google Scholar] [CrossRef]
- Michaud, M.; Balardy, L.; Moulis, G.; Gaudin, C.; Peyrot, C.; Vellas, B.; Cesari, M.; Nourhashemi, F. Proinflammatory cytokines, aging, and age-related diseases. J. Am. Med. Dir. Assoc. 2013, 14, 877–882. [Google Scholar] [CrossRef]
- Pluvinage, J.V.; Haney, M.S.; Smith, B.A.H.; Sun, J.; Iram, T.; Bonanno, L.; Li, L.; Lee, D.P.; Morgens, D.W.; Yang, A.C.; et al. CD22 blockade restores homeostatic microglial phagocytosis in ageing brains. Nature 2019, 568, 187–192. [Google Scholar] [CrossRef]
- Wang, L.; Yin, Y.L.; Liu, X.Z.; Shen, P.; Zheng, Y.G.; Lan, X.R.; Lu, C.B.; Wang, J.Z. Current understanding of metal ions in the pathogenesis of Alzheimer’s disease. Transl. Neurodegener. 2020, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Das, N.; Raymick, J.; Sarkar, S. Role of metals in Alzheimer’s disease. Metab. Brain Dis. 2021, 36, 1627–1639. [Google Scholar] [CrossRef]
- Turab Naqvi, A.A.; Hasan, G.M.; Hassan, M.I. Targeting Tau Hyperphosphorylation via Kinase Inhibition: Strategy to Address Alzheimer’s Disease. Curr. Top. Med. Chem. 2020, 20, 1059–1073. [Google Scholar] [CrossRef] [PubMed]
- House, E.; Collingwood, J.; Khan, A.; Korchazkina, O.; Berthon, G.; Exley, C. Aluminium, iron, zinc and copper influence the In vitro formation of amyloid fibrils of Aβ 42 in a manner which may have consequences for metal chelation therapy in Alzheimer’s disease. J. Alzheimer’s Dis. 2004, 6, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Matheou, C.J.; Younan, N.D.; Viles, J.H. Cu2⁺ accentuates distinct misfolding of Aβ₁₋₄₀ and Aβ₁₋₄₂ peptides, and potentiates membrane disruption. Biochem. J. 2015, 466, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Van Duijn, S.; Bulk, M.; van Duinen, S.G.; Nabuurs, R.J.A.; van Buchem, M.A.; van der Weerd, L.; Natté, R. Cortical Iron Reflects Severity of Alzheimer’s Disease. J. Alzheimers Dis. 2017, 60, 1533–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breuer, W.; Ghoti, H.; Shattat, A.; Goldfarb, A.; Koren, A.; Levin, C.; Rachmilewitz, E.; Cabantchik, Z.I. Non-transferrin bound iron in Thalassemia: Differential detection of redox active forms in children and older patients. Am. J. Hematol. 2012, 87, 55–61. [Google Scholar] [CrossRef]
- Bush, A.I. The metallobiology of Alzheimer’s disease. Trends Neurosci. 2003, 26, 207–214. [Google Scholar] [CrossRef]
- Mezzaroba, L.; Alfieri, D.F.; Colado Simão, A.N.; Vissoci Reiche, E.M. The role of zinc, copper, manganese and iron in neurodegenerative diseases. Neurotoxicology 2019, 74, 230–241. [Google Scholar] [CrossRef]
- Lakey-Beitia, J.; Burillo, A.M.; La Penna, G.; Hegde, M.L.; Rao, K.S. Polyphenols as Potential Metal Chelation Compounds Against Alzheimer’s Disease. J. Alzheimers Dis. 2021, 82, S335–S357. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.Y.L.; Mak, S.; Kong, X.; Xia, Y.; Kwan, K.K.L.; Xu, M.L.; Tsim, K.W.K. Tacrine Induces Endoplasmic Reticulum-Stressed Apoptosis via Disrupting the Proper Assembly of Oligomeric Acetylcholinesterase in Cultured Neuronal Cells. Mol. Pharm. 2021, 100, 456–469. [Google Scholar] [CrossRef]
- Palmer, A.M. The role of the blood brain barrier in neurodegenerative disorders and their treatment. J. Alzheimers Dis. 2011, 24, 643–656. [Google Scholar] [CrossRef] [PubMed]
- Duwa, R.; Jeong, J.-H.; Yook, S. Development of immunotherapy and nanoparticles-based strategies for the treatment of Parkinson’s disease. J. Pharm. Investig. 2021, 51, 465–481. [Google Scholar] [CrossRef]
- Gonzalez-Carter, D.; Liu, X.; Tockary, T.A.; Dirisala, A.; Toh, K.; Anraku, Y.; Kataoka, K. Targeting nanoparticles to the brain by exploiting the blood-brain barrier impermeability to selectively label the brain endothelium. Proc. Natl. Acad. Sci. USA 2020, 117, 19141–19150. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.S.; Leong, K.W. Advances in microphysiological blood-brain barrier (BBB) models towards drug delivery. Curr. Opin. Biotechnol. 2020, 66, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Nguyen, T.T.D.; Nguyen, T.K.O.; Vo, T.K.; Vo, V.G. Advances in developing therapeutic strategies for Alzheimer’s disease. Biomed. Pharmacother. 2021, 139, 111623. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Vo, T.K.; Vo, G.V. Therapeutic Strategies and Nano-Drug Delivery Applications in Management of Aging Alzheimer’s Disease. Adv. Exp. Med. Biol. 2021, 1286, 183–198. [Google Scholar]
- Béduneau, A.; Saulnier, P.; Benoit, J.P. Active targeting of brain tumors using nanocarriers. Biomaterials 2007, 28, 4947–4967. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Jiang, C. Evolution of blood-brain barrier in brain diseases and related systemic nanoscale brain-targeting drug delivery strategies. Acta Pharm. Sin. B 2021, 11, 2306–2325. [Google Scholar] [CrossRef]
- Zhou, Y.; Peng, Z.; Seven, E.S.; Leblanc, R.M. Crossing the blood-brain barrier with nanoparticles. J. Control. Release 2018, 270, 290–303. [Google Scholar] [CrossRef]
- Kumagai, A.; Eisenberg, J.B.; Pardridge, W. Absorptive-mediated endocytosis of cationized albumin and a beta-endorphin-cationized albumin chimeric peptide by isolated brain capillaries. Model system of blood-brain barrier transport. J. Biol. Chem. 1987, 262, 15214–15219. [Google Scholar] [CrossRef]
- Hervé, F.; Ghinea, N.; Scherrmann, J.M. CNS delivery via adsorptive transcytosis. Aaps J. 2008, 10, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Lu, C.; Leszek, J.; Zhang, J. Design and Development of Nanomaterial-Based Drug Carriers to Overcome the Blood-Brain Barrier by Using Different Transport Mechanisms. Int. J. Mol. Sci. 2021, 22, 10118. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.P.; Romme, E.; van der Spek, P.J.; Dirven, C.M.; Willemsen, R.; Kros, J.M. Glut1/SLC2A1 is crucial for the development of the blood-brain barrier in vivo. Ann. Neurol. 2010, 68, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Huttunen, J.; Peltokangas, S.; Gynther, M.; Natunen, T.; Hiltunen, M.; Auriola, S.; Ruponen, M.; Vellonen, K.S.; Huttunen, K.M. L-Type Amino Acid Transporter 1 (LAT1/Lat1)-Utilizing Prodrugs Can Improve the Delivery of Drugs into Neurons, Astrocytes and Microglia. Sci. Rep. 2019, 9, 12860. [Google Scholar] [CrossRef] [Green Version]
- Akanuma, S.I. [Membrane Transporters and Their Regulatory Mechanisms at the Brain and Retinal Barriers to Establish Therapies for Refractory Central Nervous System Diseases]. Yakugaku Zasshi 2020, 140, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.R.; Shusta, E.V. Blood-brain barrier transport of therapeutics via receptor-mediation. Pharm. Res. 2007, 24, 1759–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasnjevic, I.; Steinbusch, H.W.; Schmitz, C.; Martinez-Martinez, P. Delivery of peptide and protein drugs over the blood-brain barrier. Prog. Neurobiol. 2009, 87, 212–251. [Google Scholar] [CrossRef] [PubMed]
- Scheld, W.M. Drug delivery to the central nervous system: General principles and relevance to therapy for infections of the central nervous system. Rev. Infect. Dis. 1989, 11 (Suppl. 7), S1669–S1690. [Google Scholar] [CrossRef]
- Zhu, J.; Zhang, Y.; Chen, X.; Zhang, Y.; Zhang, K.; Zheng, H.; Wei, Y.; Zheng, H.; Zhu, J.; Wu, F.; et al. Angiopep-2 modified lipid-coated mesoporous silica nanoparticles for glioma targeting therapy overcoming BBB. Biochem. Biophys. Res. Commun. 2021, 534, 902–907. [Google Scholar] [CrossRef]
- Yurek, D.M.; Hasselrot, U.; Cass, W.A.; Sesenoglu-Laird, O.; Padegimas, L.; Cooper, M.J. Age and lesion-induced increases of GDNF transgene expression in brain following intracerebral injections of DNA nanoparticles. Neuroscience 2015, 284, 500–512. [Google Scholar] [CrossRef] [Green Version]
- Noor, N.A.; Hosny, E.N.; Khadrawy, Y.A.; Mourad, I.M.; Othman, A.I.; Aboul Ezz, H.S.; Mohammed, H.S. Effect of curcumin nanoparticles on streptozotocin-induced male Wistar rat model of Alzheimer’s disease. Metab. Brain Dis. 2022, 37, 343–357. [Google Scholar] [CrossRef]
- Heydari, S.; Hedayati Ch, M.; Saadat, F.; Abedinzade, M.; Nikokar, I.; Aboutaleb, E.; Khafri, A.; Mokarram, A.R. Diphtheria toxoid nanoparticles improve learning and memory impairment in animal model of Alzheimer’s disease. Pharm. Rep. 2020, 72, 814–826. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, O.; Baharuldin, M.T.H.; Nor, N.H.M.; Chiroma, S.M.; Jagadeesan, S.; Moklas, M.A.M. Chemicals used for the induction of Alzheimer’s disease-like cognitive dysfunctions in rodents. Biomed. Res. Ther. 2019, 6, 3460–3484. [Google Scholar] [CrossRef] [Green Version]
- Householder, K.T.; Dharmaraj, S.; Sandberg, D.I.; Wechsler-Reya, R.J.; Sirianni, R.W. Fate of nanoparticles in the central nervous system after intrathecal injection in healthy mice. Sci. Rep. 2019, 9, 12587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, H.; Navath, R.S.; Balakrishnan, B.; Guru, B.R.; Mishra, M.K.; Romero, R.; Kannan, R.M.; Kannan, S. Intrinsic targeting of inflammatory cells in the brain by polyamidoamine dendrimers upon subarachnoid administration. Nanomedicine 2010, 5, 1317–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Formicola, B.; Cox, A.; Dal Magro, R.; Masserini, M.; Re, F. Nanomedicine for the Treatment of Alzheimer’s Disease. J. Biomed. Nanotechnol. 2019, 15, 1997–2024. [Google Scholar] [CrossRef]
- Islam, S.U.; Shehzad, A.; Ahmed, M.B.; Lee, Y.S. Intranasal Delivery of Nanoformulations: A Potential Way of Treatment for Neurological Disorders. Molecules 2020, 25, 1929. [Google Scholar] [CrossRef] [Green Version]
- Prabakaran, A.; Agrawal, M.; Dethe, M.R.; Ahmed, H.; Yadav, A.; Gupta, U.; Alexander, A. Nose-to-brain drug delivery for the treatment of Alzheimer’s disease: Current advancements and challenges. Expert. Opin. Drug Deliv. 2022, 19, 87–102. [Google Scholar]
- Fonseca, L.C.; Lopes, J.A.; Vieira, J.; Viegas, C.; Oliveira, C.S.; Hartmann, R.P.; Fonte, P. Intranasal drug delivery for treatment of Alzheimer’s disease. Drug Deliv. Transl. Res. 2021, 11, 411–425. [Google Scholar] [CrossRef]
- Bhatt, P.C.; Srivastava, P.; Pandey, P.; Khan, W.; Panda, B.P. Nose to brain delivery of astaxanthin-loaded solid lipid nanoparticles: Fabrication, radio labeling, optimization and biological studies. RSC Adv. 2016, 6, 10001–10010. [Google Scholar] [CrossRef]
- Lungare, S.; Hallam, K.; Badhan, R.K.S. Phytochemical-loaded mesoporous silica nanoparticles for nose-to-brain olfactory drug delivery. Int. J. Pharm. 2016, 513, 280–293. [Google Scholar] [CrossRef] [Green Version]
- Muntimadugu, E.; Dhommati, R.; Jain, A.; Challa, V.G.; Shaheen, M.; Khan, W. Intranasal delivery of nanoparticle encapsulated tarenflurbil: A potential brain targeting strategy for Alzheimer’s disease. Eur. J. Pharm. Sci. 2016, 92, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.K.; Tsai, C.L.; Huang, Y.; Hynynen, K. Focused Ultrasound and Microbubbles-Mediated Drug Delivery to Brain Tumor. Pharmaceutics 2020, 13, 15. [Google Scholar] [CrossRef] [PubMed]
- Burgess, A.; Shah, K.; Hough, O.; Hynynen, K. Focused ultrasound-mediated drug delivery through the blood-brain barrier. Expert. Rev. Neurother. 2015, 15, 477–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Gong, Y.; Xie, W.; Huang, A.; Yuan, X.; Zhou, H.; Zhu, X.; Chen, X.; Liu, J.; Liu, J.; et al. Microbubbles in combination with focused ultrasound for the delivery of quercetin-modified sulfur nanoparticles through the blood brain barrier into the brain parenchyma and relief of endoplasmic reticulum stress to treat Alzheimer’s disease. Nanoscale 2020, 12, 6498–6511. [Google Scholar] [CrossRef]
- Kofoed, R.H.; Heinen, S.; Silburt, J.; Dubey, S.; Dibia, C.L.; Maes, M.; Simpson, E.M.; Hynynen, K.; Aubert, I. Transgene distribution and immune response after ultrasound delivery of rAAV9 and PHP.B to the brain in a mouse model of amyloidosis. Mol. Methods Clin. Dev. 2021, 23, 390–405. [Google Scholar] [CrossRef]
- Cheng, C.S.; Liu, T.P.; Chien, F.C.; Mou, C.Y.; Wu, S.H.; Chen, Y.P. Codelivery of Plasmid and Curcumin with Mesoporous Silica Nanoparticles for Promoting Neurite Outgrowth. ACS Appl. Mater. Interfaces 2019, 11, 15322–15331. [Google Scholar] [CrossRef]
- Raymond, J.J.; Robertson, D.M.; Dinsdale, H.B. Pharmacological modification of bradykinin induced breakdown of the blood-brain barrier. Can. J. Neurol. Sci. 1986, 13, 214–220. [Google Scholar] [CrossRef] [Green Version]
- Praça, C.; Rai, A.; Santos, T.; Cristovão, A.C.; Pinho, S.L.; Cecchelli, R.; Dehouck, M.P.; Bernardino, L.; Ferreira, L.S. A nanoformulation for the preferential accumulation in adult neurogenic niches. J. Control. Release 2018, 284, 57–72. [Google Scholar] [CrossRef]
- Picone, P.; Ditta, L.A.; Sabatino, M.A.; Militello, V.; San Biagio, P.L.; Di Giacinto, M.L.; Cristaldi, L.; Nuzzo, D.; Dispenza, C.; Giacomazza, D.; et al. Ionizing radiation-engineered nanogels as insulin nanocarriers for the development of a new strategy for the treatment of Alzheimer’s disease. Biomaterials 2016, 80, 179–194. [Google Scholar] [CrossRef]
- Zhou, H.; Gong, Y.; Liu, Y.; Huang, A.; Zhu, X.; Liu, J.; Yuan, G.; Zhang, L.; Wei, J.A.; Liu, J. Intelligently thermoresponsive flower-like hollow nano-ruthenium system for sustained release of nerve growth factor to inhibit hyperphosphorylation of tau and neuronal damage for the treatment of Alzheimer’s disease. Biomaterials 2020, 237, 119822. [Google Scholar] [CrossRef]
- Liu, W.; Wang, W.; Dong, X.; Sun, Y. Near-Infrared Light-Powered Janus Nanomotor Significantly Facilitates Inhibition of Amyloid-β Fibrillogenesis. ACS Appl. Mater. Interfaces 2020, 12, 12618–12628. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Kanekiyo, T. Blood-Brain Barrier Dysfunction and the Pathogenesis of Alzheimer’s Disease. Int. J. Mol. Sci. 2017, 18, 1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honary, S.; Zahir, F. Effect of zeta potential on the properties of nano-drug delivery systems-a review (Part 1). Trop. J. Pharm. Res. 2013, 12, 255–264. [Google Scholar]
- Zhang, S.; Gao, H.; Bao, G. Physical Principles of Nanoparticle Cellular Endocytosis. ACS Nano 2015, 9, 8655–8671. [Google Scholar] [CrossRef] [Green Version]
- Sukhanova, A.; Bozrova, S.; Sokolov, P.; Berestovoy, M.; Karaulov, A.; Nabiev, I. Dependence of nanoparticle toxicity on their physical and chemical properties. Nanoscale Res. Lett. 2018, 13, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Zhou, Y.; Zhao, N.; Hao, B.; Wang, X.; Kong, P. Pharmacokinetic behavior and efficiency of acetylcholinesterase inhibition in rat brain after intranasal administration of galanthamine hydrobromide loaded flexible liposomes. Environ. Toxicol. Pharm. 2012, 34, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.Z.; Zhang, Y.Q.; Wang, Z.Z.; Wu, K.; Lou, J.N.; Qi, X.R. Enhanced brain distribution and pharmacodynamics of rivastigmine by liposomes following intranasal administration. Int. J. Pharm. 2013, 452, 344–354. [Google Scholar] [CrossRef]
- Al Asmari, A.K.; Ullah, Z.; Tariq, M.; Fatani, A. Preparation, characterization, and in vivo evaluation of intranasally administered liposomal formulation of donepezil. Drug Des. Devel. 2016, 10, 205–215. [Google Scholar]
- Ordóñez-Gutiérrez, L.; Wandosell, F. Nanoliposomes as a Therapeutic Tool for Alzheimer’s Disease. Front. Synaptic. Neurosci. 2020, 12, 20. [Google Scholar] [CrossRef]
- Arora, S.; Layek, B.; Singh, J. Design and Validation of Liposomal ApoE2 Gene Delivery System to Evade Blood-Brain Barrier for Effective Treatment of Alzheimer’s Disease. Mol. Pharm. 2021, 18, 714–725. [Google Scholar] [CrossRef]
- Saffari, P.M.; Alijanpour, S.; Takzaree, N.; Sahebgharani, M.; Etemad-Moghadam, S.; Noorbakhsh, F.; Partoazar, A. Metformin loaded phosphatidylserine nanoliposomes improve memory deficit and reduce neuroinflammation in streptozotocin-induced Alzheimer’s disease model. Life Sci. 2020, 255, 117861. [Google Scholar] [CrossRef] [PubMed]
- Rotman, M.; Welling, M.M.; Bunschoten, A.; de Backer, M.E.; Rip, J.; Nabuurs, R.J.; Gaillard, P.J.; van Buchem, M.A.; van der Maarel, S.M.; van der Weerd, L. Enhanced glutathione PEGylated liposomal brain delivery of an anti-amyloid single domain antibody fragment in a mouse model for Alzheimer’s disease. J. Control. Release 2015, 203, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Li, X.T.; Ni, Y.N.; Xiao, H.H.; Yao, Y.J.; Wang, Y.Y.; Ju, R.J.; Li, H.Y.; Liu, J.J.; Fu, M.; et al. Transferrin-Modified Osthole PEGylated Liposomes Travel the Blood-Brain Barrier and Mitigate Alzheimer’s Disease-Related Pathology in APP/PS-1 Mice. Int. J. Nanomed. 2020, 15, 2841–2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardridge, W.M. Brain Delivery of Nanomedicines: Trojan Horse Liposomes for Plasmid DNA Gene Therapy of the Brain. Front. Med. Technol. 2020, 2, 8. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. Preparation of Trojan horse liposomes (THLs) for gene transfer across the blood-brain barrier. Cold Spring Harb. Protoc. 2010, 4, pdb.prot5407. [Google Scholar] [CrossRef]
- Kocsis, I.; Sanna, E.; Hunter, C.A. Liposome Enhanced Detection of Amyloid Protein Aggregates. Org. Lett. 2021, 23, 647–650. [Google Scholar] [CrossRef]
- Yang, P.; Sheng, D.; Guo, Q.; Wang, P.; Xu, S.; Qian, K.; Li, Y.; Cheng, Y.; Wang, L.; Lu, W.; et al. Neuronal mitochondria-targeted micelles relieving oxidative stress for delayed progression of Alzheimer’s disease. Biomaterials 2020, 238, 119844. [Google Scholar] [CrossRef]
- Candela, P.; Gosselet, F.; Saint-Pol, J.; Sevin, E.; Boucau, M.C.; Boulanger, E.; Cecchelli, R.; Fenart, L. Apical-to-basolateral transport of amyloid-β peptides through blood-brain barrier cells is mediated by the receptor for advanced glycation end-products and is restricted by P-glycoprotein. J. Alzheimers Dis. 2010, 22, 849–859. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Guo, Z.; Zhang, Y.; Li, C.; Zhang, Y.; Guo, Q.; Chen, Q.; Chen, X.; He, X.; Liu, L.; et al. Microenvironment Remodeling Micelles for Alzheimer’s Disease Therapy by Early Modulation of Activated Microglia. Adv. Sci. 2019, 6, 1801586. [Google Scholar] [CrossRef] [Green Version]
- Agwa, M.M.; Abdelmonsif, D.A.; Khattab, S.N.; Sabra, S. Self- assembled lactoferrin-conjugated linoleic acid micelles as an orally active targeted nanoplatform for Alzheimer’s disease. Int. J. Biol. Macromol. 2020, 162, 246–261. [Google Scholar] [CrossRef]
- Geng, H.; Yuan, H.; Qiu, L.; Gao, D.; Cheng, Y.; Xing, C. Inhibition and disaggregation of amyloid β protein fibrils through conjugated polymer-core thermoresponsive micelles. J. Mater. Chem. B 2020, 8, 10126–10135. [Google Scholar] [CrossRef] [PubMed]
- Misra, S.; Chopra, K.; Sinha, V.R.; Medhi, B. Galantamine-loaded solid-lipid nanoparticles for enhanced brain delivery: Preparation, characterization, In vitro and in vivo evaluations. Drug Deliv. 2016, 23, 1434–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, B.; Khunt, D.; Bhatt, H.; Misra, M.; Padh, H. Application of quality by design approach for intranasal delivery of rivastigmine loaded solid lipid nanoparticles: Effect on formulation and characterization parameters. Eur. J. Pharm. Sci. 2015, 78, 54–66. [Google Scholar] [CrossRef]
- Chauhan, M.K.; Sharma, P.K. Optimization and characterization of rivastigmine nanolipid carrier loaded transdermal patches for the treatment of dementia. Chem. Phys. Lipids 2019, 224, 104794. [Google Scholar] [CrossRef] [PubMed]
- Mendes, I.T.; Ruela, A.L.M.; Carvalho, F.C.; Freitas, J.T.J.; Bonfilio, R.; Pereira, G.R. Development and characterization of nanostructured lipid carrier-based gels for the transdermal delivery of donepezil. Colloids Surf. B Biointerfaces 2019, 177, 274–281. [Google Scholar] [CrossRef]
- Dara, T.; Vatanara, A.; Sharifzadeh, M.; Khani, S.; Vakilinezhad, M.A.; Vakhshiteh, F.; Nabi Meybodi, M.; Sadegh Malvajerd, S.; Hassani, S.; Mosaddegh, M.H. Improvement of memory deficits in the rat model of Alzheimer’s disease by erythropoietin-loaded solid lipid nanoparticles. Neurobiol. Learn. Mem. 2019, 166, 107082. [Google Scholar] [CrossRef] [PubMed]
- Vakilinezhad, M.A.; Amini, A.; Akbari Javar, H.; Baha’addini Beigi Zarandi, B.F.; Montaseri, H.; Dinarvand, R. Nicotinamide loaded functionalized solid lipid nanoparticles improves cognition in Alzheimer’s disease animal model by reducing Tau hyperphosphorylation. Daru 2018, 26, 165–177. [Google Scholar] [CrossRef]
- Yadav, A.; Sunkaria, A.; Singhal, N.; Sandhir, R. Resveratrol loaded solid lipid nanoparticles attenuate mitochondrial oxidative stress in vascular dementia by activating Nrf2/HO-1 pathway. Neurochem. Int. 2018, 112, 239–254. [Google Scholar] [CrossRef]
- Loureiro, J.A.; Andrade, S.; Duarte, A.; Neves, A.R.; Queiroz, J.F.; Nunes, C.; Sevin, E.; Fenart, L.; Gosselet, F.; Coelho, M.A.N.; et al. Resveratrol and Grape Extract-loaded Solid Lipid Nanoparticles for the Treatment of Alzheimer’s Disease. Molecules 2017, 22, 277. [Google Scholar] [CrossRef]
- Pinheiro, R.G.R.; Granja, A.; Loureiro, J.A.; Pereira, M.C.; Pinheiro, M.; Neves, A.R.; Reis, S. Quercetin lipid nanoparticles functionalized with transferrin for Alzheimer’s disease. Eur. J. Pharm. Sci. 2020, 148, 105314. [Google Scholar] [CrossRef]
- Dhawan, S.; Kapil, R.; Singh, B. Formulation development and systematic optimization of solid lipid nanoparticles of quercetin for improved brain delivery. J. Pharm. Pharm. 2011, 63, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Asghar, S.; Gao, S.; Su, Z.; Song, J.; Huo, M.; Meng, W.; Ping, Q.; Xiao, Y. A novel LDL-mimic nanocarrier for the targeted delivery of curcumin into the brain to treat Alzheimer’s disease. Colloids Surf. B: Biointerfaces 2015, 134, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, A.; Carreiró, F.; Oliveira, A.M.; Neves, A.; Pires, B.; Venkatesh, D.N.; Durazzo, A.; Lucarini, M.; Eder, P.; Silva, A.M.; et al. Polymeric Nanoparticles: Production, Characterization, Toxicology and Ecotoxicology. Molecules 2020, 25, 3731. [Google Scholar] [CrossRef] [PubMed]
- Allahyari, M.; Mohit, E. Peptide/protein vaccine delivery system based on PLGA particles. Hum. Vaccin Immunother. 2016, 12, 806–828. [Google Scholar] [CrossRef] [Green Version]
- Baysal, I.; Ucar, G.; Gultekinoglu, M.; Ulubayram, K.; Yabanoglu-Ciftci, S. Donepezil loaded PLGA-b-PEG nanoparticles: Their ability to induce destabilization of amyloid fibrils and to cross blood brain barrier In vitro. J. Neural. Transm. 2017, 124, 33–45. [Google Scholar] [CrossRef]
- Nanaki, S.G.; Spyrou, K.; Bekiari, C.; Veneti, P.; Baroud, T.N.; Karouta, N.; Grivas, I.; Papadopoulos, G.C.; Gournis, D.; Bikiaris, D.N. Hierarchical Porous Carbon-PLLA and PLGA Hybrid Nanoparticles for Intranasal Delivery of Galantamine for Alzheimer’s Disease Therapy. Pharmaceutics 2020, 12, 227. [Google Scholar] [CrossRef] [Green Version]
- Pagar, K.; Vavia, P. Rivastigmine-loaded L-lactide-depsipeptide polymeric nanoparticles: Decisive formulation variable optimization. Sci. Pharm. 2013, 81, 865–885. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Si, J.; Xu, H.; Dong, J.; Zheng, D.; Lu, X.; Li, X. Resveratrol-loaded nanoparticles reduce oxidative stress induced by radiation or amyloid-beta in transgenic Caenorhabditis elegans. J. Biomed. Nanotechnol. 2014, 10, 1536–1544. [Google Scholar] [CrossRef]
- Fan, S.; Zheng, Y.; Liu, X.; Fang, W.; Chen, X.; Liao, W.; Jing, X.; Lei, M.; Tao, E.; Ma, Q.; et al. Curcumin-loaded PLGA-PEG nanoparticles conjugated with B6 peptide for potential use in Alzheimer’s disease. Drug Deliv. 2018, 25, 1091–1102. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Gao, X.; Kang, T.; Jiang, M.; Miao, D.; Gu, G.; Hu, Q.; Song, Q.; Yao, L.; Tu, Y.; et al. B6 peptide-modified PEG-PLA nanoparticles for enhanced brain delivery of neuroprotective peptide. Bioconjug. Chem. 2013, 24, 997–1007. [Google Scholar] [CrossRef]
- Cano, A.; Ettcheto, M.; Chang, J.H.; Barroso, E.; Espina, M.; Kühne, B.A.; Barenys, M.; Auladell, C.; Folch, J.; Souto, E.B.; et al. Dual-drug loaded nanoparticles of Epigallocatechin-3-gallate (EGCG)/Ascorbic acid enhance therapeutic efficacy of EGCG in a APPswe/PS1dE9 Alzheimer’s disease mice model. J. Control. Release 2019, 301, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Silva-Abreu, M.; Calpena, A.C.; Andrés-Benito, P.; Aso, E.; Romero, I.A.; Roig-Carles, D.; Gromnicova, R.; Espina, M.; Ferrer, I.; García, M.L.; et al. PPARγ agonist-loaded PLGA-PEG nanocarriers as a potential treatment for Alzheimer’s disease: In vitro and in vivo studies. Int. J. Nanomed. 2018, 13, 5577–5590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Li, N.; Zhang, W.; Zhao, Z.; Mou, Z.; Huang, D.; Liu, J.; Wang, W. Design of PLGA-functionalized quercetin nanoparticles for potential use in Alzheimer’s disease. Colloids Surf. B Biointerfaces 2016, 148, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Manek, E.; Darvas, F.; Petroianu, G.A. Use of Biodegradable, Chitosan-Based Nanoparticles in the Treatment of Alzheimer’s Disease. Molecules 2020, 25, 4866. [Google Scholar] [CrossRef]
- Singh, S.K.; Mishra, D.N. Nose to Brain Delivery of Galantamine Loaded Nanoparticles: In-vivo Pharmacodynamic and Biochemical Study in Mice. Curr. Drug Deliv. 2019, 16, 51–58. [Google Scholar]
- Mahl, C.R.A.; Taketa, T.B.; Rocha-Neto, J.B.M.; Almeida, W.P.; Beppu, M.M. Copper Ion Uptake by Chitosan in the Presence of Amyloid-β and Histidine. Appl. Biochem. Biotechnol. 2020, 190, 949–965. [Google Scholar] [CrossRef]
- Gothwal, A.; Kumar, H.; Nakhate, K.T.; Ajazuddin; Dutta, A.; Borah, A.; Gupta, U. Lactoferrin Coupled Lower Generation PAMAM Dendrimers for Brain Targeted Delivery of Memantine in Aluminum-Chloride-Induced Alzheimer’s Disease in Mice. Bioconjug Chem. 2019, 30, 2573–2583. [Google Scholar] [CrossRef]
- DeRidder, L.; Sharma, A.; Liaw, K.; Sharma, R.; John, J.; Kannan, S.; Kannan, R.M. Dendrimer-tesaglitazar conjugate induces a phenotype shift of microglia and enhances β-amyloid phagocytosis. Nanoscale 2021, 13, 939–952. [Google Scholar] [CrossRef]
- Klajnert, B.; Wasiak, T.; Ionov, M.; Fernandez-Villamarin, M.; Sousa-Herves, A.; Correa, J.; Riguera, R.; Fernandez-Megia, E. Dendrimers reduce toxicity of Aβ 1-28 peptide during aggregation and accelerate fibril formation. Nanomedicine 2012, 8, 1372–1378. [Google Scholar] [CrossRef]
- Gupta, A. Chapter 21—Nanoemulsions. In Nanoparticles for Biomedical Applications; Chung, E.J., Leon, L., Rinaldi, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 371–384. [Google Scholar]
- Altinoglu, G.; Adali, T. Alzheimer’s Disease Targeted Nano-Based Drug Delivery Systems. Curr. Drug Targets 2020, 21, 628–646. [Google Scholar] [CrossRef]
- Nirale, P.; Paul, A.; Yadav, K.S. Nanoemulsions for targeting the neurodegenerative diseases: Alzheimer’s, Parkinson’s and Prion’s. Life Sci. 2020, 245, 117394. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Nigam, K.; Srivastava, S.; Tyagi, A.; Dang, S. Memantine nanoemulsion: A new approach to treat Alzheimer’s disease. J. Microencapsul. 2020, 37, 355–365. [Google Scholar] [CrossRef]
- Kaur, A.; Nigam, K.; Bhatnagar, I.; Sukhpal, H.; Awasthy, S.; Shankar, S.; Tyagi, A.; Dang, S. Treatment of Alzheimer’s diseases using donepezil nanoemulsion: An intranasal approach. Drug Deliv. Transl. Res. 2020, 10, 1862–1875. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Liu, C.; Zhai, W.; Zhuang, N.; Han, T.; Ding, Z. The Optimization Design Of Lactoferrin Loaded HupA Nanoemulsion For Targeted Drug Transport Via Intranasal Route. Int. J. Nanomed. 2019, 14, 9217–9234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Tsibouklis, J.; Weng, T.; Zhang, B.; Yin, G.; Feng, G.; Cui, Y.; Savina, I.N.; Mikhalovska, L.I.; Sandeman, S.R.; et al. Nano carriers for drug transport across the blood-brain barrier. J. Drug Target 2017, 25, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tak, K.; Sharma, R.; Dave, V.; Jain, S.; Sharma, S. Clitoria ternatea Mediated Synthesis of Graphene Quantum Dots for the Treatment of Alzheimer’s Disease. ACS Chem. Neurosci. 2020, 11, 3741–3748. [Google Scholar] [CrossRef]
- Wang, S.; Li, C.; Xia, Y.; Chen, S.; Robert, J.; Banquy, X.; Huang, R.; Qi, W.; He, Z.; Su, R. Nontoxic Black Phosphorus Quantum Dots Inhibit Insulin Amyloid Fibrillation at an Ultralow Concentration. iScience 2020, 23, 101044. [Google Scholar] [CrossRef]
- Zhou, X.; Hu, S.; Wang, S.; Pang, Y.; Lin, Y.; Li, M. Large Amino Acid Mimicking Selenium-Doped Carbon Quantum Dots for Multi-Target Therapy of Alzheimer’s Disease. Front. Pharm. 2021, 12, 778613. [Google Scholar] [CrossRef]
- Mars, A.; Hamami, M.; Bechnak, L.; Patra, D.; Raouafi, N. Curcumin-graphene quantum dots for dual mode sensing platform: Electrochemical and fluorescence detection of APOe4, responsible of Alzheimer’s disease. Anal. Chim. Acta 2018, 1036, 141–146. [Google Scholar] [CrossRef]
- Hou, K.; Zhao, J.; Wang, H.; Li, B.; Li, K.; Shi, X.; Wan, K.; Ai, J.; Lv, J.; Wang, D.; et al. Chiral gold nanoparticles enantioselectively rescue memory deficits in a mouse model of Alzheimer’s disease. Nat. Commun. 2020, 11, 4790. [Google Scholar] [CrossRef]
- Dos Santos Tramontin, N.; da Silva, S.; Arruda, R.; Ugioni, K.S.; Canteiro, P.B.; de Bem Silveira, G.; Mendes, C.; Silveira, P.C.L.; Muller, A.P. Gold Nanoparticles Treatment Reverses Brain Damage in Alzheimer’s Disease Model. Mol. Neurobiol. 2020, 57, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Sanati, M.; Khodagholi, F.; Aminyavari, S.; Ghasemi, F.; Gholami, M.; Kebriaeezadeh, A.; Sabzevari, O.; Hajipour, M.J.; Imani, M.; Mahmoudi, M.; et al. Impact of Gold Nanoparticles on Amyloid β-Induced Alzheimer’s Disease in a Rat Animal Model: Involvement of STIM Proteins. ACS Chem. Neurosci. 2019, 10, 2299–2309. [Google Scholar] [CrossRef] [PubMed]
- Razzino, C.A.; Serafín, V.; Gamella, M.; Pedrero, M.; Montero-Calle, A.; Barderas, R.; Calero, M.; Lobo, A.O.; Yáñez-Sedeño, P.; Campuzano, S.; et al. An electrochemical immunosensor using gold nanoparticles-PAMAM-nanostructured screen-printed carbon electrodes for tau protein determination in plasma and brain tissues from Alzheimer patients. Biosens. Bioelectron. 2020, 163, 112238. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Wang, Y.; Tan, R.; Li, H.; Tu, Y. Determination of β-amyloid oligomer using electrochemiluminescent aptasensor with signal enhancement by AuNP/MOF nanocomposite. Mikrochim. Acta 2021, 188, 53. [Google Scholar] [CrossRef]
- Enteshari Najafabadi, R.; Kazemipour, N.; Esmaeili, A.; Beheshti, S.; Nazifi, S. Using superparamagnetic iron oxide nanoparticles to enhance bioavailability of quercetin in the intact rat brain. BMC Pharm. Toxicol. 2018, 19, 59. [Google Scholar] [CrossRef]
- Li, Y.; Lim, E.; Fields, T.; Wu, H.; Xu, Y.; Wang, Y.A.; Mao, H. Improving Sensitivity and Specificity of Amyloid-β Peptides and Tau Protein Detection with Antibiofouling Magnetic Nanoparticles for Liquid Biopsy of Alzheimer’s Disease. ACS Biomater. Sci. Eng. 2019, 5, 3595–3605. [Google Scholar] [CrossRef] [PubMed]
- Antonoglou, O.; Giannousi, K.; Mourdikoudis, S.; Dendrinou-Samara, C. Magnetic nanoemulsions as candidates for Alzheimer’s disease dual imaging theranostics. Nanotechnology 2020, 31, 465702. [Google Scholar] [CrossRef]
- Yin, T.; Yang, L.; Liu, Y.; Zhou, X.; Sun, J.; Liu, J. Sialic acid (SA)-modified selenium nanoparticles coated with a high blood-brain barrier permeability peptide-B6 peptide for potential use in Alzheimer’s disease. Acta Biomater. 2015, 25, 172–183. [Google Scholar] [CrossRef]
- Lopez-Barbosa, N.; Garcia, J.G.; Cifuentes, J.; Castro, L.M.; Vargas, F.; Ostos, C.; Cardona-Gomez, G.P.; Hernandez, A.M.; Cruz, J.C. Multifunctional magnetite nanoparticles to enable delivery of siRNA for the potential treatment of Alzheimer’s. Drug Deliv. 2020, 27, 864–875. [Google Scholar] [CrossRef]
- Morgan, D. Immunotherapy for Alzheimer’s disease. J. Intern. Med. 2011, 269, 54–63. [Google Scholar] [CrossRef]
- Schenk, D. Amyloid-β immunotherapy for Alzheimer’s disease: The end of the beginning. Nat. Rev. Neurosci. 2002, 3, 824–828. [Google Scholar] [CrossRef] [PubMed]
- Sarlus, H.; Heneka, M.T. Microglia in Alzheimer’s disease. J. Clin. Investig. 2017, 127, 3240–3249. [Google Scholar] [CrossRef]
- Hansen, D.V.; Hanson, J.E.; Sheng, M. Microglia in Alzheimer’s disease. J. Cell Biol. 2018, 217, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Fassbender, K.; Walter, S.; Kühl, S.; Landmann, R.; Ishii, K.; Bertsch, T.; Stalder, A.; Muehlhauser, F.; Liu, Y.; Ulmer, A. The LPS receptor (CD14) links innate immunity with Alzheimer’s disease. FASEB J. 2004, 18, 203–205. [Google Scholar] [CrossRef]
- Hardy, J.; Duff, K.; Hardy, K.G.; Perez-Tur, J.; Hutton, M. Genetic dissection of Alzheimer’s disease and related dementias: Amyloid and its relationship to tau. Nat. Neurosci. 1998, 1, 355–358. [Google Scholar] [CrossRef] [PubMed]
- Ittner, A.; Bertz, J.; Suh, L.S.; Stevens, C.H.; Götz, J.; Ittner, L.M. Tau-targeting passive immunization modulates aspects of pathology in tau transgenic mice. J. Neurochem. 2015, 132, 135–145. [Google Scholar] [CrossRef]
- Baruch, K.; Deczkowska, A.; Rosenzweig, N.; Tsitsou-Kampeli, A.; Sharif, A.M.; Matcovitch-Natan, O.; Kertser, A.; David, E.; Amit, I.; Schwartz, M. PD-1 immune checkpoint blockade reduces pathology and improves memory in mouse models of Alzheimer’s disease. Nat. Med. 2016, 22, 135–137. [Google Scholar] [CrossRef]
- Schwartz, M. Can immunotherapy treat neurodegeneration? Science 2017, 357, 254–255. [Google Scholar] [CrossRef]
- Rosenzweig, N.; Dvir-Szternfeld, R.; Tsitsou-Kampeli, A.; Keren-Shaul, H.; Ben-Yehuda, H.; Weill-Raynal, P.; Cahalon, L.; Kertser, A.; Baruch, K.; Amit, I.; et al. PD-1/PD-L1 checkpoint blockade harnesses monocyte-derived macrophages to combat cognitive impairment in a tauopathy mouse model. Nat. Commun. 2019, 10, 465. [Google Scholar] [CrossRef]
- Latta-Mahieu, M.; Elmer, B.; Bretteville, A.; Wang, Y.; Lopez-Grancha, M.; Goniot, P.; Moindrot, N.; Ferrari, P.; Blanc, V.; Schussler, N.; et al. Systemic immune-checkpoint blockade with anti-PD1 antibodies does not alter cerebral amyloid-β burden in several amyloid transgenic mouse models. Glia 2018, 66, 492–504. [Google Scholar] [CrossRef]
- Obst, J.; Mancuso, R.; Simon, E.; Gomez-Nicola, D. PD-1 deficiency is not sufficient to induce myeloid mobilization to the brain or alter the inflammatory profile during chronic neurodegeneration. Brain Behav. Immun. 2018, 73, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Liu, N.; Wang, C.; Qin, B.; Zhou, Y.; Xiao, M.; Chang, L.; Yan, L.J.; Zhao, B. Role of RAGE in Alzheimer’s Disease. Cell Mol. Neurobiol. 2016, 36, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Lue, L.F.; Yan, S.; Xu, H.; Luddy, J.S.; Chen, D.; Walker, D.G.; Stern, D.M.; Yan, S.; Schmidt, A.M.; et al. RAGE-dependent signaling in microglia contributes to neuroinflammation, Abeta accumulation, and impaired learning/memory in a mouse model of Alzheimer’s disease. FASEB J. 2010, 24, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Liu, C.; Zhou, Y.; Qi, J.; Zhang, C.; Sun, B.; Wang, J.; Guan, Y. Progress of RAGE Molecular Imaging in Alzheimer’s Disease. Front. Aging Neurosci. 2020, 12, 227. [Google Scholar] [CrossRef]
- Loureiro, J.A.; Gomes, B.; Fricker, G.; Cardoso, I.; Ribeiro, C.A.; Gaiteiro, C.; Coelho, M.A.; Pereira Mdo, C.; Rocha, S. Dual ligand immunoliposomes for drug delivery to the brain. Colloids Surf. B Biointerfaces 2015, 134, 213–219. [Google Scholar] [CrossRef]
- Hao, S.; Li, X.; Han, A.; Yang, Y.; Fang, G.; Liu, J.; Wang, S. CLVFFA-Functionalized Gold Nanoclusters Inhibit Aβ40 Fibrillation, Fibrils’ Prolongation, and Mature Fibrils’ Disaggregation. ACS Chem. Neurosci. 2019, 10, 4633–4642. [Google Scholar] [CrossRef]
- Skaat, H.; Corem-Slakmon, E.; Grinberg, I.; Last, D.; Goez, D.; Mardor, Y.; Margel, S. Antibody-conjugated, dual-modal, near-infrared fluorescent iron oxide nanoparticles for antiamyloidgenic activity and specific detection of amyloid-β fibrils. Int. J. Nanomed. 2013, 8, 4063–4076. [Google Scholar]
- Chen, L.; Lin, J.; Yi, J.; Weng, Q.; Zhou, Y.; Han, Z.; Li, C.; Chen, J.; Zhang, Q. A tyrosinase-induced fluorescence immunoassay for detection of tau protein using dopamine-functionalized CuInS(2)/ZnS quantum dots. Anal. Bioanal. Chem. 2019, 411, 5277–5285. [Google Scholar] [CrossRef]
- Lo Buono, N.; Parrotta, R.; Morone, S.; Bovino, P.; Nacci, G.; Ortolan, E.; Horenstein, A.L.; Inzhutova, A.; Ferrero, E.; Funaro, A. The CD157-integrin partnership controls transendothelial migration and adhesion of human monocytes. J. Biol. Chem. 2011, 286, 18681–18691. [Google Scholar] [CrossRef] [Green Version]
- Wimmer, I.; Tietz, S.; Nishihara, H.; Deutsch, U.; Sallusto, F.; Gosselet, F.; Lyck, R.; Muller, W.A.; Lassmann, H.; Engelhardt, B. PECAM-1 Stabilizes Blood-Brain Barrier Integrity and Favors Paracellular T-Cell Diapedesis Across the Blood-Brain Barrier During Neuroinflammation. Front. Immunol. 2019, 10, 711. [Google Scholar] [CrossRef] [Green Version]
- Chang, X.; Li, J.; Niu, S.; Xue, Y.; Tang, M. Neurotoxicity of metal-containing nanoparticles and implications in glial cells. J. Appl. Toxicol. 2021, 41, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.L.; Gao, J.Q. Potential neurotoxicity of nanoparticles. Int. J. Pharm. 2010, 394, 115–121. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Drugs | Trade Name (Company) | Action Mechanism | Dosage Form | Dose | LogP (Permeability) Molecular Weight (M.W) (g/mol) | K (Partition Coefficient) |
|---|---|---|---|---|---|---|
| Tacrine | Cognex (Sciele, Atlanta, GA, USA) | Reversible inhibition of acetylcholinesterase (AChE) | Capsule | Initial dose: 10 mg orally q.i.d (between meals if possible) for 6 weeks. Maintenance dose: may increase to 20 mg orally q.i.d. Further increases to 120 mg and 160 mg/day may be done in 6-week intervals. | 2.71 M.W = 234.72 | |
| Donepezil | Aricept (Pfizer, New York City, NY, USA) | Reversible inhibitor of acetylcholinesterase which prevents the hydrolysis of acetylcholine. | Tablet | 5 mg q.d., may increase to 10 mg/day after 4–6 weeks if tolerated, then to 23 mg/day after at least 3 months | 4.7 M.W = 379.5 | log Kow = 4.86 (est) |
| Rivastigmine | Exelon (Novartis, Basel, Switzerland) | Inhibits the hydrolytic activity of AChE and BChE and binds to catalytic sites. | Solution Capsule | Oral Solution and capsules 6 mg to 12 mg per day, administered b.i.d. (daily doses of 3 mg to 6 mg b.i.d.) | 2.3 M.W = 250.34 | |
| Galantamine | Razadyne (Janssen, Beerse, Belgium) | Binds reversibly to acetylcholine esterase and enhances the intrinsic action of acetylcholine on nicotinic receptors. | Tablet Capsule Oral solution | Tablets contain 4 mg, 8 mg, and 12 mg galantamine. Capsules contain 8 mg, 16 mg, and 24 mg galantamine. Oral solution contains 4 mg galantamine (as 5.13 mg galantamine hydrobromide) per mL. | 1.8 M.W = 287.35 | |
| Memantine | Namenda (Allergan, Dublin, Ireland) | N-methyl D-aspartate (NMDA) antagonist | Tablet Oral solution | Tablets: 5 mg q.d., may increase to 10 mg/day, 15 mg/day, and 20 mg/day at 1-week intervals if tolerated Oral solution: same as above | 3.28 M.W = 179.3 | log Kow = 3.28 |
| Memantine and Donepezil | Namzaric (Actavis, Parsippany, NJ, USA) | N-methyl D-aspartate (NMDA) antagonist plus cholinesterase inhibitor | Capsule (ER) | 7 mg memantine/10 mg donepezil q.d., may increase to 28 mg memantine/10 mg donepezil in 7 mg increments at 1-week intervals if tolerated | M.W = 215.76 | |
| Aducanumab | Aduhelm (Biogen, Cambridge, MA, USA) | Targets accumulated Aβ plaques | Intravenous infusion | Doses 1 and 2 (weeks 0 and 4)—1 mg/kg IV over one hour Doses 3 and 4 (weeks 8 and 16—3 mg/kg IV over one hour | M.W = 145,912.34 |
| Drug | Mechanism of Action | Therapeutic Purpose | Stage of AD | Target | Phase | Sponsor | Start Estimated End Dates | |
|---|---|---|---|---|---|---|---|---|
| Disease-Modifying Biologics | ||||||||
| New Therapeutics | Aducanumab | Disrupts Aβ plaques and oligomers | DMT | Mild to moderate | Amyloid | 3 | Biogen, Cambridge, MA, USA | Mar 2020 Oct 2023 |
| Gantenerumab | Monoclonal antibody acts at Aβ plaques and oligomers | DMT | Mild to moderate | Amyloid | 2 | Roche, Basel, Switzerland | Dec 2020 Feb 2024 | |
| Pepinemab | Monoclonal antibody directed at semaphoring 4D to reduce inflammation | DMT | Mild | Inflammation | 1 | Vaccinex. Inc., Rochester, NY, USA | Jul 2021 Jan 2023 | |
| LY3372689 | Tau protein aggregation inhibitor | DMT | Moderate | Tau | 2 | Eli Lilly & Company, Indianapolis, IN, USA | Sep 2021 Jun 2024 | |
| TRx0237 | Inhibition of tau protein aggregation | DMT | Mild to moderate | Tau | 3 | TauRx Therapeutics, Aberdeen, Scotland | Jan 2018 Jun 2023 | |
| Disease-Modifying Small Molecules | ||||||||
| Sumifilam | Alters conformation of filamin A | DMT | Mild to moderate | Filamin A | 2 | Cassava Sciences, Inc, Austin, TX, USA | Nov 2021 Oct 2023 | |
| Tricaprilin (contains caprylic acid triglyceride) | Induces ketosis and improves mitochondrial and neuronal function | DMT | Mild to moderate | Metabolism and bioenergetics | 3 | Cerecin, Anson Road, Singapore | Jan 2021 Feb 2023 | |
| Symptoms-Reducing Small Molecules | ||||||||
| AD-35 | Acetylcholinesterase inhibitor | Cognitive enhancer | Mild to moderate | Neurotransmitter receptors | 2 | Zhejiang Hisun Pharmaceutical, Jiaojiang District, Taizhou, China | Dec 2018 July 2021 | |
| Disease-Modifying Biologics | ||||||||
| Repositioned Drugs | BCG vaccine | Vaccination against tuberculosis infection; immunomodulator | DMT | - | Inflammation/immunity | 2 | Mindful Diagnostics and Therapeutics, Eau Claire, WI, USA | Nov 2020 Dec 2021 |
| IVIG (NewGam 10%) | Polyclonal antibody | DMT | Mild | Amyloid | 2 | Sutter Health, Sacramento, CA, USA | Jan 2011 Dec 2019 | |
| Disease-Modifying Small Molecules | ||||||||
| Losartan & amlodipine & atorvastatin + exercise | Angiotensin II receptor blocker (losartan), calcium channel blocker (amlodipine), cholesterol agent (atorvastatin) | DMT | - | Vasculature | 3 | University of Texas Southwestern, Houston, TX, USA | Feb 2017 Mar 2022 | |
| Metformin | Sensitize insulin to improve metabolism of CNS glucose | DMT | Mild | Metabolism and bioenergetics | 3 | Columbia University, NIA, NY, USA | Jan 2021 Apr 2024 | |
| Montelukast | Leukotriene receptor antagonist | DMT | Mild to moderate | Inflammation | 2 | Emory University, Atlanta, GA, USA | Sep 2019 Oct 2022 | |
| Hydralazine | Antioxidant | DMT | Mild to moderate | Oxidative stress | 3 | Shahid Sadoughi University of Medical Sciences and Health services, Yazd, Iran | Jun 2021 Jun 2023 | |
| Suvorexant | Dual orexin receptor antagonist; improves sleep with effects on CSF Aβ | DMT | - | Neurotransmitter receptors | 2 | Washington University School of Medicine, St. Louis, MO, USA | Jan 2021 Jan 2025 | |
| Trehalose | Induces autophagy and promotes clearance of aggregated proteins | DMT | Mild to moderate | Cell death | 1 | Mashhad University of Medical Sciences, Mashhad, Razavi Khorasan, Iran | Aug 2020 Aug 2022 | |
| Vorinostat | Histone deacetylase (HDAC) inhibitor; enhanced synaptic plasticity | DMT | Mild | Epigenetics | 1 | German Center of Neurodegenrative Disease (DZNE), Bonn, Germany | Sep 2017 Mar 2022 | |
| Symptoms-Reducing Small Molecules | ||||||||
| Caffeine | Pleiotropic effect on CNS function | Cognitive Enhancer | Mild | Metabolism and bioenergetics | 3 | University Hospital, Lille, France | Nov 2021 Nov 2024 | |
| Escitalopram | Selective serotonin reuptake inhibitor | Neuropsychiatric symptoms | Mild | Neurotransmitter receptors | 3 | Johns Hopkins University, NIA, Baltimore, MD, USA | Jan 2018 Aug 2022 | |
| Nabilone | Synthetic cannabinoid; antiemetic | Neuropsychiatric symptoms (agitation) | Mild to moderate | Neurotransmitter receptors | 3 | Sunnybrook Health Sciences Center, ADDF, Torinto, ON, Canada | Feb 2021 Oct 2025 | |
| Nicotine | Nicotinic acetylcholine receptor agonist | Cognitive enhancer | Mild | Neurotransmitter receptors | 2 | University of Southern, California, NIA, ATRI, Vanderbilt University, Nashville, TN, USA | Jan 2017 July 2023 | |
| Prazosin | α-1 adrenoreceptor antagonist | Neuropsychiatric symptoms (agitation) | Mild to moderate | Neurotransmitter receptors | 2 | ADCS, NIA, Maitland, FL, USA | Oct 2018 Dec 2022 | |
| Riluzole | Glutamate modulator agent | Cognitive enhancer | Mild | Neurotransmitter receptors | 2 | Icahn School of Medicine at Mount Sinai, NY city, NY, USA | Completed | |
| Sargramostim | Recombinant human GM-CSF | Cognitive enhancer | Mild to moderate | Metabolism and bioenergetics | 2 | University of Colorado, Denver, USA | Dec 2021 Jul 2024 | |
| THC-free CBD oil | Cannabinoid with effects on cannabinoid receptors | Neuropsychiatric symptoms agents (agitation) | Severe | Neurotransmitter receptors | 2 | Eastern Virgina Medical School, Norfolk, VA, USA | Feb 2021 Jun 2022 | |
| Influx Transporters | Efflux Transporters | Receptor Transporters |
|---|---|---|
| Choline transporter (ChT) | Peptide transport system-6 (PTs-6) | Insulin receptors (IR) |
| Sodium-coupled glucose transporters (SGLTs) | Breast cancer resistant protein (BCRP) | Insulin-like growth factor receptor (IGFR) |
| Cationic amino acid transporter (CAT1) | P-glycoprotein (P-gp) | Transferrin receptors (TfR) |
| 1-type large amino-acid transporter (LAT1) Excitatory amino-acid transporters (EAATs) | Leptin receptor (LepR) | |
| Glucose transporter (GLUT1) (GLUT3) | Low-density lipoprotein receptor (LDLR) | |
| Monocarboxylate lactate transporter (MCT1) Receptor for glycation end products (RAGE) | Neonatal Fc receptor (FcRN) | |
| Lactoferrin receptor (LR) |
| Drug | Carrier Material | Ligand | Particle Size (nm) | Zeta Potential (mV) | Route of Administration, Dose | In Vitro/In Vivo Model | Outcome | Reference |
|---|---|---|---|---|---|---|---|---|
| Liposome | ||||||||
| Metformin | Phosphatidyl serine | - | 145 | −41 | Intraperitoneal, 50 mg/kg | Adult male Wistar rats | in vivo: Decreased neuroinflammation. Enhanced cognition restoration. | [112] |
| - | Chitosan | pApoE2 | 167.8 ± 2.47 | 19.8 ± 3.6 | Intravenous, 1 mg/kg | bEnd.3 cells C57BL/6 mice | in vitro: Decreased cell viability in all cell lines. Increased concentration of phospholipid. in vivo: Approximately 2 times higher ApoE protein expression than endogenous ApoE levels. | [111] |
| Osthole | - | Transferrin | 104.28 ± 3.76 | −6.95 ± 0.56 | Tail vein intravenous, 10 mg/kg | hCMEC/D3 cells and APP-SH-SY5Y cells APP/PS-transgenic mice | in vitro/in vivo: pH-controlled release up to 72 h. Enhanced penetration and drug accumulation. | [114] |
| GSH-PEG | VHH-pa2H Glutathione (GSH) | 108 | IV bolus injection, 5 mg/kg | APPswe/PS1dE9 transgenic mice | in vivo: Increased standard uptake values (SUV) of VHH-pa2H in the blood. | [113] | ||
| Galantamine HBr | Soya Phosphatidylcholine | - | 112 ± 8 | −49.2 ± 0.7 | Oral and intranasal, 3 mg/kg | PC-12 cell, male SD rats | in vitro: Reduced cytotoxicity Enhanced BBB penetration. in vivo: More bioavailability in brain. Enhanced efficacy. | [107] |
| Donepezil | 1,2-distearyl-sn-glycero-3-phospholine (DSPC) | - | 102 ± 3.3 | −28.31 ± 0.85 | Oral and intranasal, 1 mg/kg | Male Wistar rats | in vitro: 75.5% drug release in 8 h. in vivo: Enhanced penetration. Enhanced bioavailability. | [109] |
| Rivastigmine | EPC, Cholesterol, DSPE-PEG-CPP | CPP | 178.9 | −8.6 ± 2.4 | Intranasal and intravenous, 1 mg/kg | Endothelial cells, male SD rats | in vitro High encapsulation efficiency. Sustained-release behavior. in vivo IN route shows more activity | [108] |
| Micelles | ||||||||
| Resveratrol | PEG-PLA | C3 peptide | 43.85 ± 0.94 | 12.9 ± 0.17 | Intravenous, 10 mg/kg | HT22 cells, APP/PS1 transgenic mice | in vitro: Improvement on brain accumulation and increased targeting. in vivo: Enhance cognitive performance. Inhibited Aβ aggregation. Enhanced activity. | [118] |
| Curcumin | PEG | Aβ peptide | 65 | Intravenous | SH-SY5Y, APPswe/PS1dE9 transgenic mice | in vitro/in vivo: Increased efficacy. Inhibited Aβ aggregation. Improved memory behavior. | [120] | |
| - | Linoleic acid | Lactoferrin | 120 ± 12.4 | −32.8 ± 3.66 | Oral, 4 gm/mL | Adult male Wistar rats | in vitro/in vivo Enhanced cognition Reduced oxidative stress. Reduced Aβ aggregation. | [121] |
| PMO-b-PBM, POEG-b-PBM and PF | - | 70 | - | PC-12 cells | in vitro: Reduced Aβ aggregation. Reduced Aβ fibrillation-induced cytotoxicity. | [122] | ||
| Solid–lipid NPs | ||||||||
| galantamine HBr | Glyceryl behnate, pluronic F-127, tween 80 | - | 88 ± 1.89 | −18.75 ± 1.7 | Oral route, 2.5 mg/kg | Adult Wistar rats | in vitro: Maximum drug entrapment. Drug release > 90% for a period of 24 h. in vivo: Enhanced bioavailability 2-fold Memory restoration. | [123] |
| Rivastigmine | Campritol 888 ATO | - | 82.5 ± 4.07 | 3.20 ± 1.44 | - | Franz diffusion cell, goat nasal mucosa | in vitro: Higher diffusion with SLN. Ex-vivo: Maximum diffusion was seen with up to 8 h. | [124] |
| Donepezil | Stearic acid, oleic acid, lecithin, sodium taurodeoxytaurocholate | - | 177.05 ± 2.12 | −55.35 | Transdermal | - | in vitro: Increased drug skin permeation. Enhanced drug delivery. | [126] |
| Rivastigmine | GMS, castor oil | - | 134.5 ± 15.1 | −11.8 ± 2.24 | Transdermal | Albino Wistar rats | in vitro/in vivo: Non-irritant. Enhanced bioavailability. | [125] |
| Erythropoietin | GMS, span 60, span 80, tween 80 | - | 219.9 ± 15.6 | −22.4 ± 0.8 | Intraperitoneal, 1250 IU/kg and 2500 IU/kg | Albino male Wistar rats | in vitro: 39% of EPO released in first 3 h followed by approximately 90% for 72 h. in vivo: Improved memory dysfunction. Enhanced efficacy. | [127] |
| Nicotinamide | Stearic acid, phospholipon 90G, sodium taurocholate | Phosphatidylserine | 124 ± 0.8 | −46.1 ± 0.65 | Intravenous or intraperitoneal | BCES, SH-SY5Y, adult male Sprague-Dawley rats | in vitro: Enhanced BBB penetration. in vivo: Neuroprotective potential Improved cognition impairment. | [128] |
| Resveratrol | Lecithin | - | 286 ± 1.47 | −17.5 ± 0.23 | Oral, 10 mg/kg | Male Sprague-Dawley rats | in vitro: 91% sustained release after 24 h. in vivo: 4.5 times bioavailability in brain. Effective antioxidant activity. | [129] |
| Resveratrol and grape extract | Cetyl palmitate, tween 80, tween 20 | Anti-transferrin receptor mAb (OX26 mAb) | 254 ± 17 | −4.0 ± 0.1 | - | HBEC | in vitro: Efficient cellular uptake. Inhibited Aβ aggregation. | [130] |
| Quercetin | - | - | 152 | −20.7 | Intravenous, 4.41 mg/kg | Male Wistar rats | in vitro/in vivo: 85.73% Drug entrapment efficiency. Effective in crossing BBB. Improvement in memory. | [132] |
| Lipid NPs | ||||||||
| Quercetin | - | Transferrin | 219 ± 13 | −28 ± 2 | - | hCMEC/D3 cells | in vitro: 80–90% of entrapment efficiency Minimum cytotoxicity at BBB cell line. | [131] |
| Curcumin | PC, cholesterol oleate, glycerol trioleate | Lactoferrin | 103.8 ± 0.6 | −5.80 ± 0.73 | Intravenous, 10 mg/kg | BCECs, SD rats | in vitro/in vivo: Effective BBB penetration. Long-term leads to enhanced activity. | [133] |
| Polymeric-NPs | ||||||||
| Galantamine | PLA-PLGA | - | 198.00 ± 0.02 | −27.42 ± 0.03 | Intranasal, 3 mg/kg | Wistar rats | in vitro/in vivo: Good stability between NP and drug. More bioavailability in the brain. | [137] |
| Donepezil | PEG-PLGA | - | 174 ± 12 | −20.45 | - | HBMEC and HA cell | in vitro: High destabilizing effect on fibril formation. Decreased neuroinflammation. | [136] |
| Rivastigmine | L-Lactide-depsipeptide | - | 142.2 ± 21.3 | - | - | in vitro: Entrapment efficiency of 60.72 ± 3.72% was obtained. Showed sustained release with 90% of drug for up to 72 h. | [138] | |
| Resveratrol | Methoxy PEG, -caprolactone | - | 80.70 ± 7.12 | ~0 mV | - | Caenorhabditis elegans, N2, CF1553, CL4176, and CL1175 | in vitro/in vivo: Enhanced antioxidant properties. Enhanced radical scavenging. Enhanced anti-lipid peroxidation. | [139] |
| Curcumin | PLGA-PEG | B6 peptide | ~100 | 3.83 ± 0.89 | Intraperitoneal, 25 mg/kg | HT22 cells/APP/PS1 transgenic mice | in vitro: Enhanced cellular uptake. Possessed good blood compatibility in vivo: Improvement in spatial learning and memory. Ex-vivo: Reduced Aβ aggregation. | [140] |
| ECG | PLGA, PEG, ascorbic acid, tween 80 | - | 124.8 ± 5.2 | −15 | Oral, 40 mg/kg | BMVECs, APP/PS1, C57BL/6 mice | in vitro: Effective in crossing BBB. in vivo: Reduced neuroinflammation. Enhance spatial learning and memory. | [142] |
| Pioglitazone | PLGA-PEG, tween 80 | Anti-Aβ antibody | 155.0 ± 1.8 | −13.0 ± 0.5 | Oral, 10 mg/kg | HBEC,hCMEC/D3 cell line, APP/PS1 transgenic mice | in vitro: Better BBB penetration. in vivo: Reduced Aβ peptide in brain. Improved memory defect. | [143] |
| Quercetin | PLGA, PVA | - | 150 | Intravenous, 20 mg/kg | SH-SY5Y cells, APP/PS1 mice, BALB/c nude mice | in vitro/in vivo: Low cellular toxicity. High cell viability. Inhibited neurotoxicity. Enhanced cognition and memory. | [144] | |
| - | PEG-PLA | B6 peptide | 118.3 ± 7.8 | −22.65 ± 0.85 | Intravenous, 1 mg/kg | bEnd.3 cells, male ICR mice | in vitro: Enhanced brain penetration. in vivo: Enhanced cognition. | [141] |
| Galantamine | Thiolated-chitosan NPs | - | 149.3 | 27.2 | Intranasal 4 mg/kg | Swiss albino mice | in vivo: Effective acetylcholinesterase activity. | [146] |
| Memantine | PAMAM (dendrimer) | Lactoferrin | 131.72 ± 4.73 | 20.13 ± 0.94 | Intraperitoneally, 2 mg/kg | Swiss albino mice | in vitro: PAMAM-MEM maximum release concentration was 77.14 ± 6.0% after 6 h. in vivo: Improvement in behavioral responses. Enhanced acetylcholinesterase (AChE) activity. | [148] |
| Nanoemulsions | ||||||||
| Memantine | - | - | ~11 | −19.6 | Intranasal, 5 mg/kg | Neuro 2a, Sprague-Dawley rats | in vitro/in vivo: 98% cell viability and sustained its antioxidative potential. Increased bioavailability. | [154] |
| Donepezil | Labrasol (10%) as oil, CPC (1%) as surfactant in water (80%), glycerol (10%) as co-surfactant | 65.36 | −10.7 | Intranasal, 0.45 mg/kg | Neuro 2a, Sprague-Dawley rats | in vitro/in vivo: Maximum drug release of 99.22% in 4 h in PBS. Non-toxic Effective drug delivery. | [155] | |
| Huperazine A | Capryol 90 (oil phase), cremophor EL & labrasol (surfactant & co-surfactant) & lactoferrin (targeting ligand) | Lactoferrin | 16.75 ± 0.4 | 5.67 ± 0.39 | Intranasal | hCMEC/D3 cells, adult Wistar rats | in vitro/in vivo: No nasal mucosal toxicity. Effective BBB penetration. Enhanced activity. | [156] |
| Quantum Dots | ||||||||
| - | Graphene QDs | - | 10 ± 1.3 | −40 ± 0.4 | - | Adult male Wistar rats | in vitro/in vivo: Enhanced penetration across BBB. Improved learning and memory. Reduced level of lipid peroxide and nitric oxide. | [158] |
| - | Black phosphorous QDs | - | ~3 | - | PC12 cells | in vitro: Low cell toxicity. Inhibited insulin and Aβ aggregation. | [159] | |
| - | Selenium-doped carbon QD | - | ~25 | Intravenous | PC12 cells, adult male Wistar rats | in vitro/in vivo: High cell viability. Inhibited Aβ aggregation. Improved memory and cognitive function of an AD rat model. | [160] | |
| Curcumin | Graphene QD & indium-tin-oxide Electrode | - | ~8 | - | - | in vitro: High sensitivity on detection of ApoE4 DNA. Enhanced efficacy. | [161] | |
| Gold nanoparticles | ||||||||
| - | - | L and D glutathione | 4 | Intravenous, 25 mg/kg | SH-SY5Y cells, C57BL/6 mice | in vitro/in vivo: Effective BBB penetration. Improved behavioral performance. | [162] | |
| AuNP | - | - | 20 | Intraperitoneal, 2.5 mg/kg | Male Wistar rats | in vivo: Normalize Tau phosphorylation. Prevented oxidative stress and neuroinflammation. | [163] | |
| AuNP | - | Bucladesine | 5 | −47.7 ± 10.9 | Intrahippocampal, intraperitoneal | Male Wistar rats | in vivo: Better acquisition and retention of spatial learning and memory. Improved neuron survival. | [164] |
| - | 3D-Au-PAMAM, electro grafted PABA | CAb-GA conjugate | - | - | Detection: Effective detection of tau protein. LOD value of 1.7 pg/mL. | [165] | ||
| Magnetic Nanoparticles | ||||||||
| Quercetin | SPIONs | - | 50 | Oral, 50 and 100 mg/kg | Male Wistar rats | in vivo: Increased penetration. Enhanced Bioavailability. | [167] | |
| - | Sialic acid (SA)-modified selenium (Se) NPs | B6 peptide | 95 | −14.4 | - | PC12 cells and bEnd.3 cells | in vitro: Effective in crossing BBB. Inhibitory effects on Aβ peptide. | [170] |
| siRNA | PEGylated magnetite NPs | OmpA | 10 | - | HFF-1 cells and SH-SY5Y cells | in vitro: Reduced cell toxicity. Enhanced activity. Silencing of BACE1 gene in HFF-1 cells. | [171] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poudel, P.; Park, S. Recent Advances in the Treatment of Alzheimer’s Disease Using Nanoparticle-Based Drug Delivery Systems. Pharmaceutics 2022, 14, 835. https://doi.org/10.3390/pharmaceutics14040835
Poudel P, Park S. Recent Advances in the Treatment of Alzheimer’s Disease Using Nanoparticle-Based Drug Delivery Systems. Pharmaceutics. 2022; 14(4):835. https://doi.org/10.3390/pharmaceutics14040835
Chicago/Turabian StylePoudel, Prashant, and Soyeun Park. 2022. "Recent Advances in the Treatment of Alzheimer’s Disease Using Nanoparticle-Based Drug Delivery Systems" Pharmaceutics 14, no. 4: 835. https://doi.org/10.3390/pharmaceutics14040835
APA StylePoudel, P., & Park, S. (2022). Recent Advances in the Treatment of Alzheimer’s Disease Using Nanoparticle-Based Drug Delivery Systems. Pharmaceutics, 14(4), 835. https://doi.org/10.3390/pharmaceutics14040835