


The Rate of Cisplatin Dosing Affects the Resistance and Metastatic Potential of Triple Negative Breast Cancer Cells, Independent of Hypoxia



Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. NanoParticle (NP) Preparation and CDDP Loading
2.3. Characterization of the Lipid NP: ζ-Potential
2.4. Characterization of the Lipid NP: Release of CDDP from the NP
2.5. Determination of the CDDP IC50 Values on the Cell Monolayers
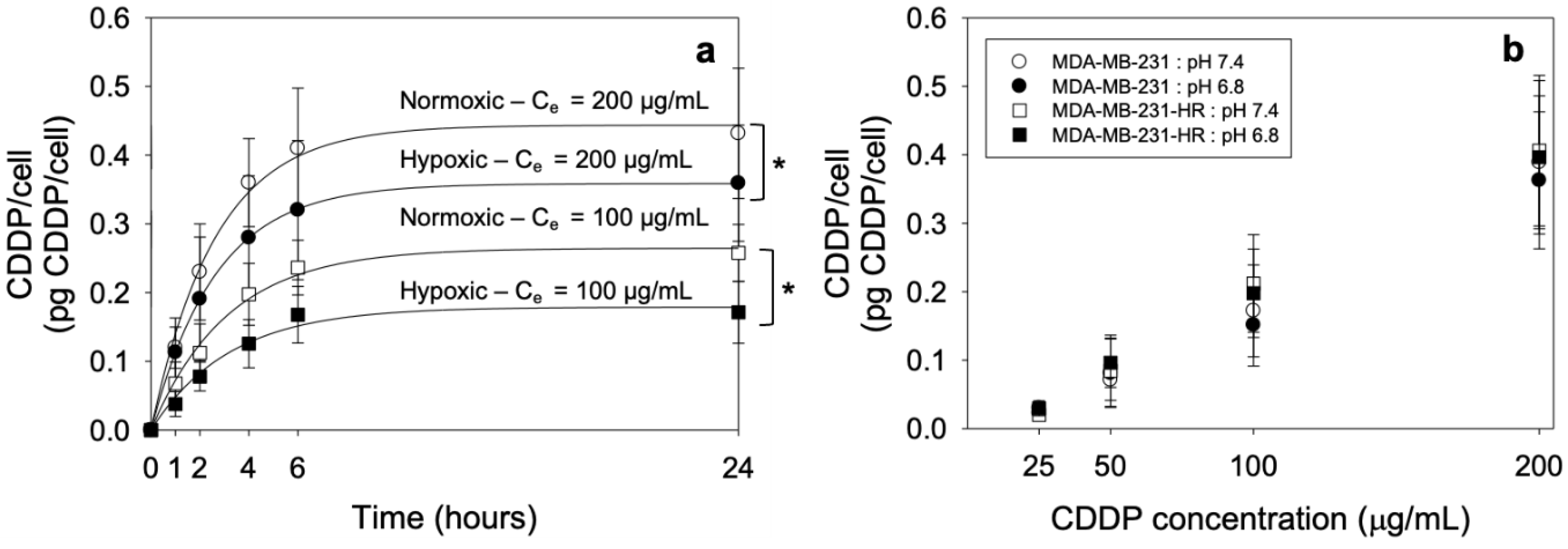
2.6. Measurement of the CDDP Cell Uptake
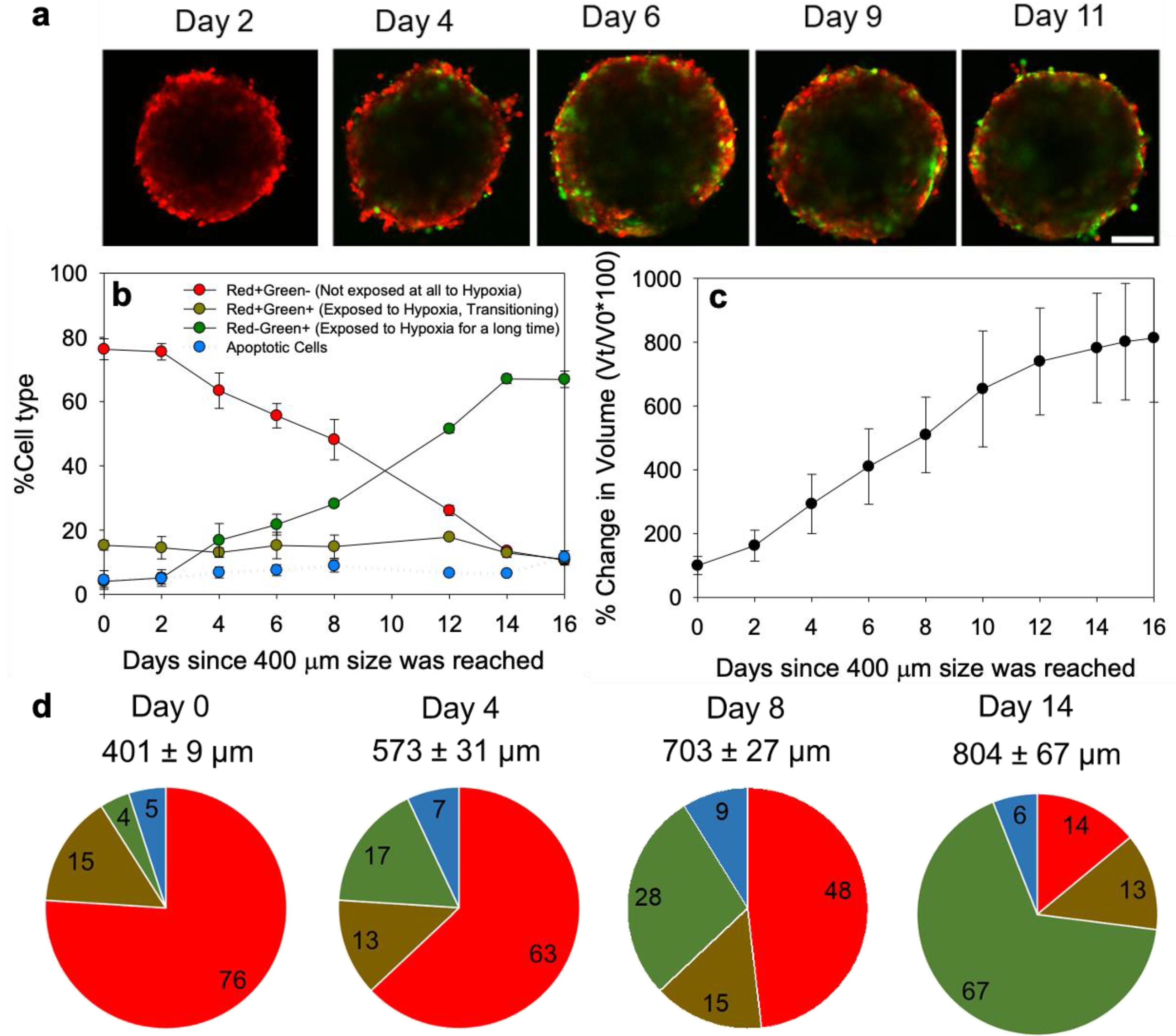
2.7. Spheroid Formation and the Cell Population Characterization
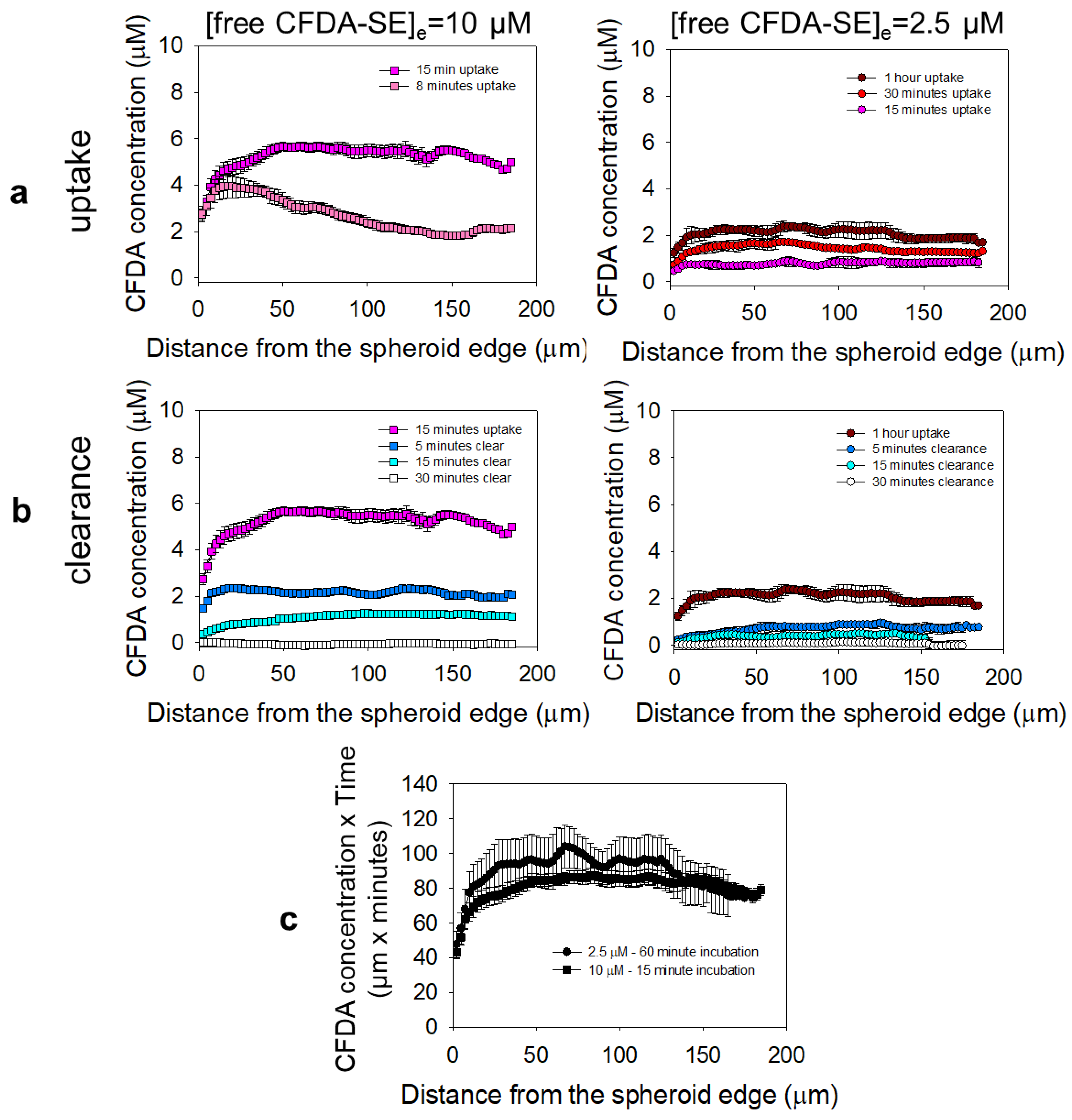
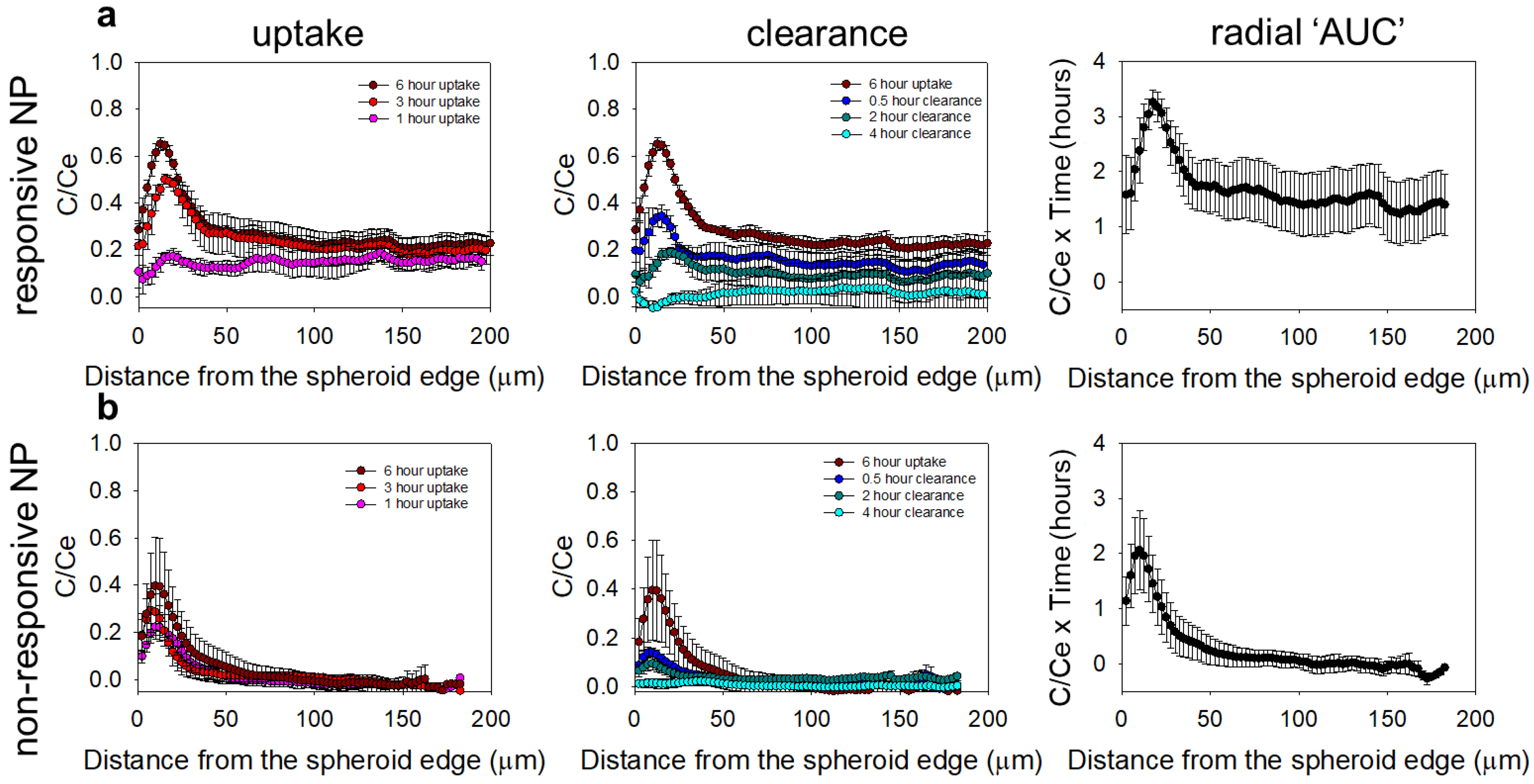
2.8. Spatiotemporal Profiles of the NPs and the Drug Surrogate in the Spheroids
2.9. Spheroid Treatment
2.10. Cell Migration from the Spheroids Embedded in a Matrix
2.11. Statistical Analysis
3. Results
3.1. Cell Characterization
3.2. NP Characterization
3.3. Ic50 Values and the Drug Efficacy on the Cell Monolayers
3.4. Spheroid Characterization
3.5. Microdistributions in the Spheroids of a Drug Surrogate for the Different Delivery Approaches
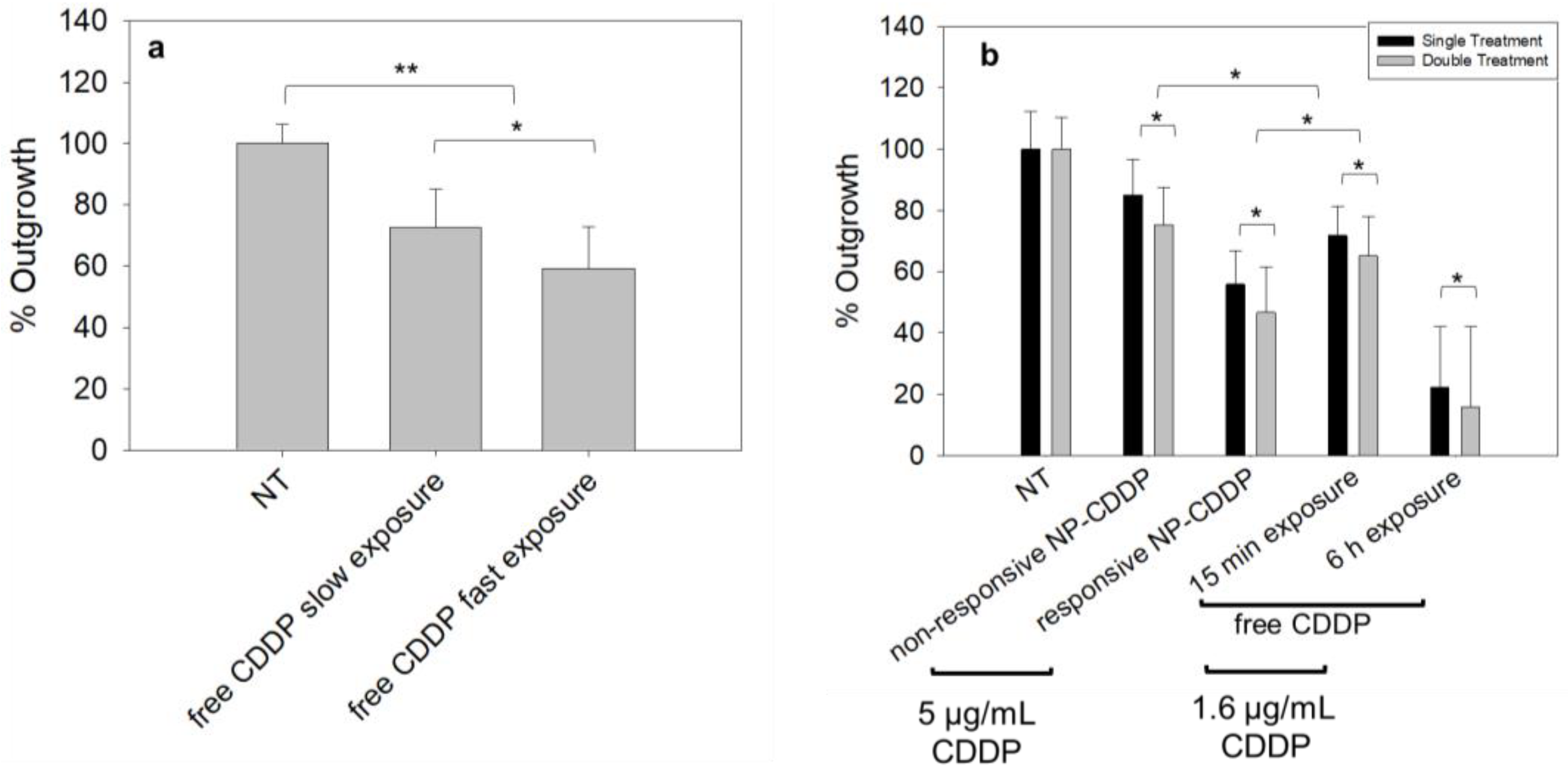
3.6. Effect of the Rate of the Free CDDP Treatment on the Spheroids: Cell Survival
3.7. Effect of the CDDP Delivery Regimens on the Spheroids: Cell Survival
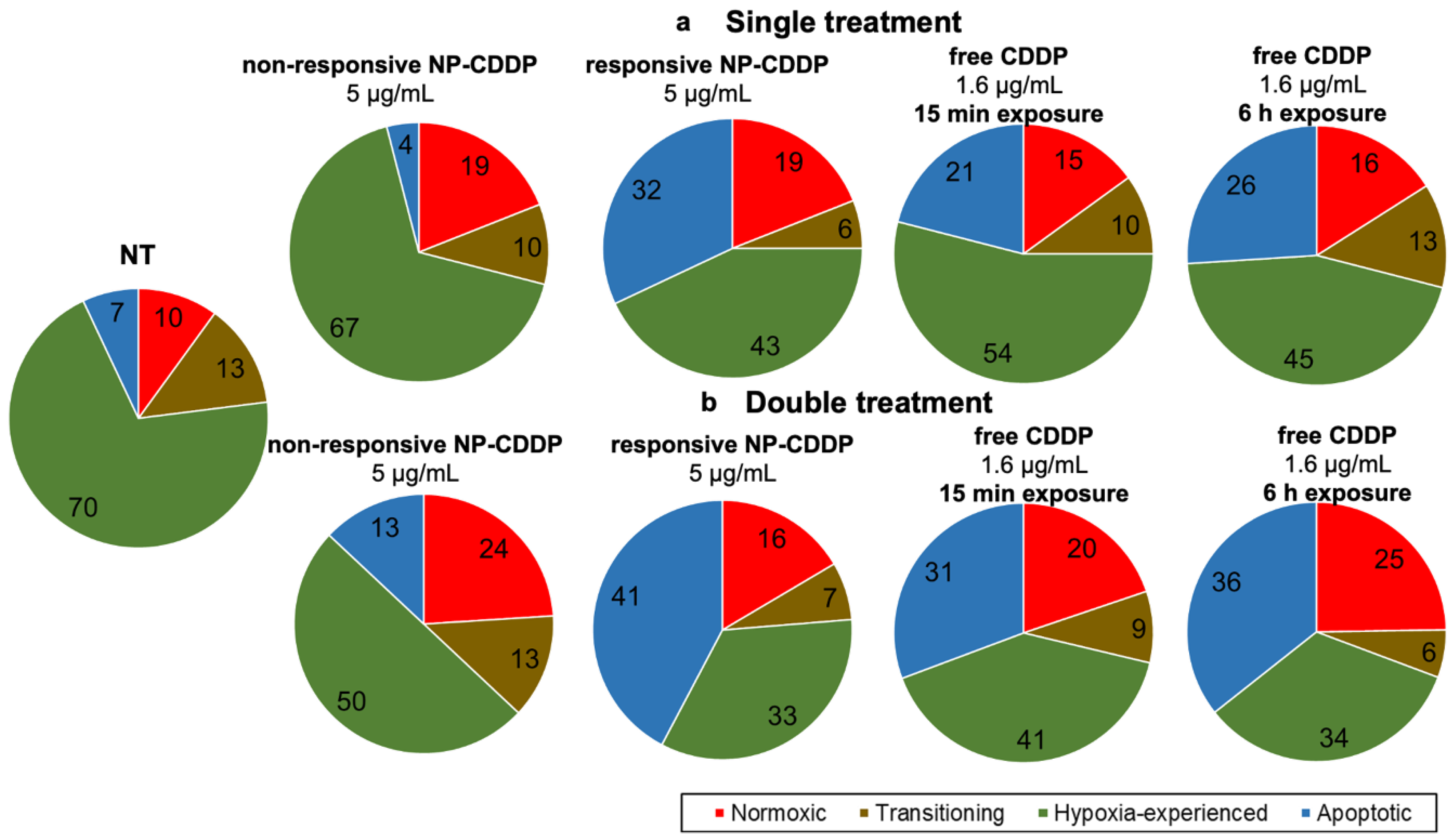
3.8. Effect of the CDDP Delivery Regimens on the Spheroids: Cell Population Progression
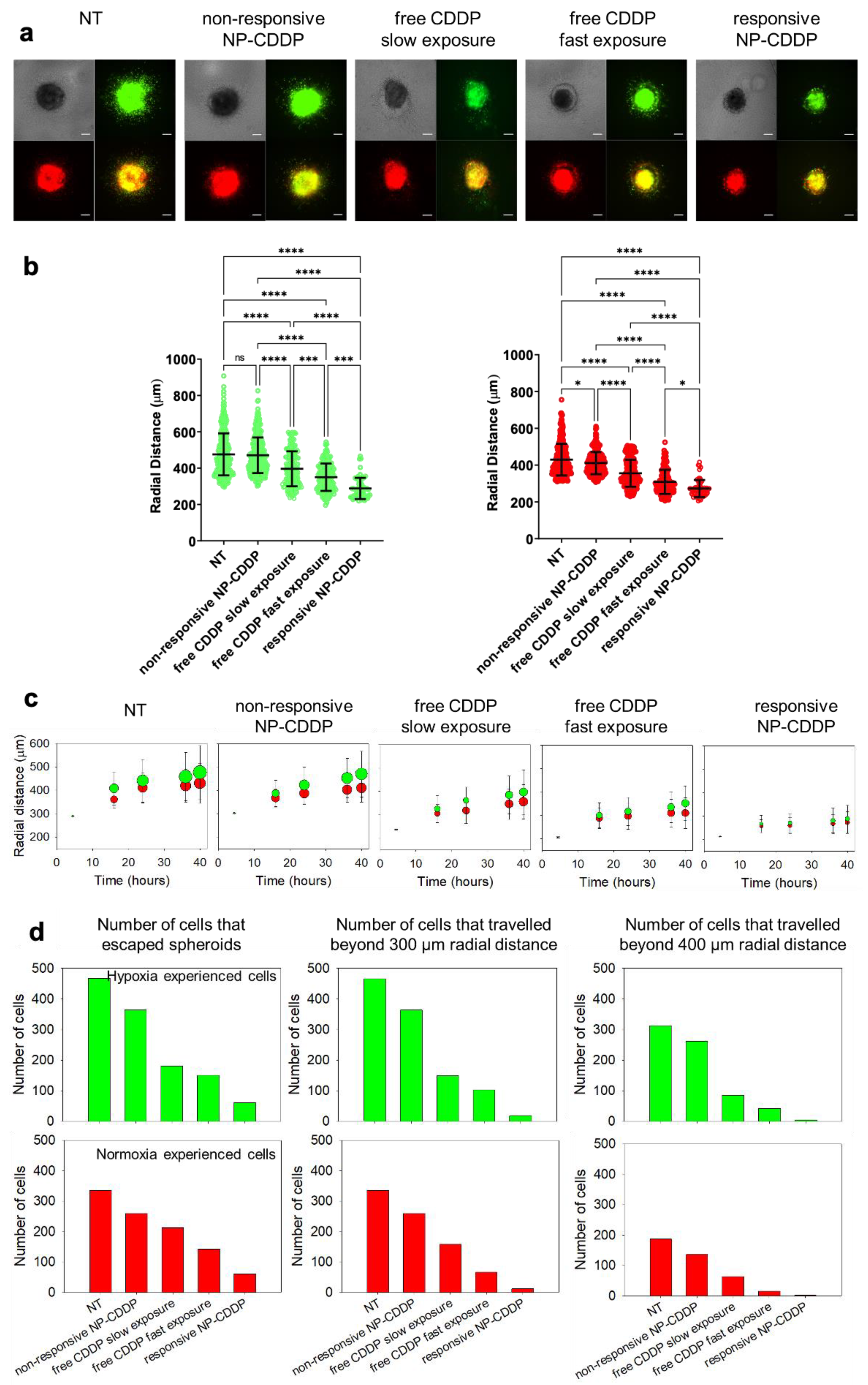
3.9. Effect of the CDDP Delivery Regimens on the Spheroids: Cell Migration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hutchinson, L. Breast cancer: TNBC: Can we treat the untargetable? Nat. Rev. Clin. Oncol. 2014, 11, 379. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.K.; Stylianopoulos, T. Delivering nanomedicine to solid tumors. Nat. Rev. Clin. Oncol. 2010, 7, 653–664. [Google Scholar] [CrossRef]
- Vagia, E.; Mahalingam, D.; Cristofanilli, M. The Landscape of Targeted Therapies in TNBC. Cancers 2020, 12, 916. [Google Scholar] [CrossRef]
- Kwapisz, D. Pembrolizumab and atezolizumab in triple-negative breast cancer. Cancer Immunol Immun. 2021, 70, 607–617. [Google Scholar] [CrossRef] [PubMed]
- He, M.Y.; Rancoule, C.; Rehailia-Blanchard, A.; Espenel, S.; Trone, J.C.; Bernichon, E.; Guillaume, E.; Vallard, A.; Magne, N. Radiotherapy in triple-negative breast cancer: Current situation and upcoming strategies. Crit. Rev. Oncol. Hematol. 2018, 131, 96–101. [Google Scholar] [CrossRef]
- Gupta, G.K.; Collier, A.L.; Lee, D.; Hoefer, R.A.; Zheleva, V.; Siewertsz van Reesema, L.L.; Tang-Tan, A.M.; Guye, M.L.; Chang, D.Z.; Winston, J.S.; et al. Perspectives on Triple-Negative Breast Cancer: Current Treatment Strategies, Unmet Needs, and Potential Targets for Future Therapies. Cancers 2020, 12, 2392. [Google Scholar] [CrossRef]
- Tannock, I.F. Tumor physiology and drug resistance. Cancer Metastasis Rev. 2001, 20, 123–132. [Google Scholar] [CrossRef]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-negative breast cancer: Clinical features and patterns of recurrence. Clin. Cancer Res. 2007, 13, 4429–4434. [Google Scholar] [CrossRef]
- Cohen, S.M.; Lippard, S.J. Cisplatin: From DNA damage to cancer chemotherapy. Prog. Nucleic Acid Res. Mol. Biol. 2001, 67, 93–130. [Google Scholar] [CrossRef]
- Sempkowski, M.; Locke, T.; Stras, S.; Zhu, C.; Sofou, S. Liposome-Based Approaches for Delivery of Mainstream Chemotherapeutics: Preparation Methods, Liposome Designs, Therapeutic Efficacy. Crit. Rev. Oncog. 2014, 19, 177–221. [Google Scholar] [CrossRef]
- Rottenberg, S.; Disler, C.; Perego, P. The rediscovery of platinum-based cancer therapy. Nat. Rev. Cancer 2021, 21, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Vianello, C.; Cocetta, V.; Catanzaro, D.; Dorn, G.W.; De Milito, A.; Rizzolio, F.; Canzonieri, V.; Cecchin, E.; Roncato, R.; Toffoli, G.; et al. Cisplatin resistance can be curtailed by blunting Bnip3-mediated mitochondrial autophagy. Cell Death Dis. 2022, 13, 398. [Google Scholar] [CrossRef]
- Harmers, F.P.; Gispen, W.H.; Neijt, J.P. Neurotoxic side-effects of cisplatin. Eur. J. Cancer 1991, 27, 372–376. [Google Scholar] [CrossRef]
- Duffy, E.A.; Fitzgerald, W.; Boyle, K.; Rohatgi, R. Nephrotoxicity: Evidence in Patients Receiving Cisplatin Therapy. Clin. J. Oncol. Nurs. 2018, 22, 175–183. [Google Scholar] [CrossRef]
- Santana-Davila, R.; Szabo, A.; Arce-Lara, C.; Williams, C.D.; Kelley, M.J.; Whittle, J. Cisplatin versus carboplatin-based regimens for the treatment of patients with metastatic lung cancer. An analysis of Veterans Health Administration data. J. Thorac. Oncol. 2014, 9, 702–709. [Google Scholar] [CrossRef]
- Gilkes, D.M.; Semenza, G.L. Role of hypoxia-inducible factors in breast cancer metastasis. Future Oncol. 2013, 9, 1623–1636. [Google Scholar] [CrossRef]
- Ebright, R.Y.; Zachariah, M.A.; Micalizzi, D.S.; Wittner, B.S.; Niederhoffer, K.L.; Nieman, L.T.; Chirn, B.; Wiley, D.F.; Wesley, B.; Shaw, B.; et al. HIF1A signaling selectively supports proliferation of breast cancer in the brain. Nat. Commun. 2020, 11, 6311. [Google Scholar] [CrossRef]
- Semenza, G.L. Targeting HIF-1 for cancer therapy. Nat. Rev. Cancer 2003, 3, 721–732. [Google Scholar] [CrossRef]
- Damgaci, S.; Ibrahim-Hashim, A.; Enriquez-Navas, P.M.; Pilon-Thomas, S.; Guvenis, A.; Gillies, R.J. Hypoxia and acidosis: Immune suppressors and therapeutic targets. Immunology 2018, 154, 354–362. [Google Scholar] [CrossRef]
- Voss, N.C.S.; Dreyer, T.; Henningsen, M.B.; Vahl, P.; Honoré, B.; Boedtkjer, E. Targeting the Acidic Tumor Microenvironment: Unexpected Pro-Neoplastic Effects of Oral NaHCO3 Therapy in Murine Breast Tissue. Cancers 2020, 12, 891. [Google Scholar] [CrossRef]
- Godet, I.; Shin, Y.J.; Ju, J.A.; Ye, I.C.; Wang, G.; Gilkes, D.M. Fate-mapping post-hypoxic tumor cells reveals a ROS-resistant phenotype that promotes metastasis. Nat. Commun. 2019, 10, 4862. [Google Scholar] [CrossRef] [PubMed]
- Godet, I.; Mamo, M.; Thurnheer, A.; Rosen, D.M.; Gilkes, D.M. Post-Hypoxic Cells Promote Metastatic Recurrence after Chemotherapy Treatment in TNBC. Cancers 2021, 13, 5509. [Google Scholar] [CrossRef] [PubMed]
- Stras, S.; Holleran, T.; Howe, A.; Sofou, S. Interstitial Release of Cisplatin from Triggerable Liposomes Enhances Efficacy against Triple Negative Breast Cancer Solid Tumor Analogues. Mol. Pharm. 2016, 13, 3224–3233. [Google Scholar] [CrossRef] [PubMed]
- Stras, S.; Howe, A.; Prasad, A.; Salerno, D.; Bhatavdekar, O.; Sofou, S. Growth of Metastatic Triple-Negative Breast Cancer Is Inhibited by Deep Tumor-Penetrating and Slow Tumor-Clearing Chemotherapy: The Case of Tumor-Adhering Liposomes with Interstitial Drug Release. Mol. Pharm. 2020, 17, 118–131. [Google Scholar] [CrossRef]
- Bandekar, A.; Karve, S.; Chang, M.Y.; Mu, Q.; Rotolo, J.; Sofou, S. Antitumor efficacy following the intracellular and interstitial release of liposomal doxorubicin. Biomaterials 2012, 33, 4345–4352. [Google Scholar] [CrossRef]
- Karve, S.; Bajagur Kempegowda, G.; Sofou, S. Heterogeneous domains and membrane permeability in phosphatidylcholine-phosphatidic acid rigid vesicles as a function of pH and lipid chain mismatch. Langmuir 2008, 24, 5679–5688. [Google Scholar] [CrossRef]
- Bailey, A.L.; Cullis, P.R. Modulation of membrane fusion by asymmetric transbilayer distributions of amino lipids. Biochemistry 1994, 33, 12573–12580. [Google Scholar] [CrossRef]
- Helmlinger, G.; Yuan, F.; Dellian, M.; Jain, R.K. Interstitial pH and pO2 gradients in solid tumors in vivo: High-resolution measurements reveal a lack of correlation. Nat. Med. 1997, 3, 177–182. [Google Scholar] [CrossRef]
- Vaupel, P.; Kallinowski, F.; Okunieff, P. Blood flow, oxygen and nutrient supply, and metabolic microenvironment of human tumors: A review. Cancer Res. 1989, 49, 6449–6465. [Google Scholar]
- Van Hennik, M.B.; van der Vijgh, W.J.; Klein, I.; Elferink, F.; Vermorken, J.B.; Winograd, B.; Pinedo, H.M. Comparative pharmacokinetics of cisplatin and three analogues in mice and humans. Cancer Res. 1987, 47, 6297–6301. [Google Scholar]
- Newman, M.S.; Colbern, G.T.; Working, P.K.; Engbers, C.; Amantea, M.A. Comparative pharmacokinetics, tissue distribution, and therapeutic effectiveness of cisplatin encapsulated in long-circulating, pegylated liposomes (SPI-077) in tumor-bearing mice. Cancer Chemother. Pharmacol. 1999, 43, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Valencia, A.M.J.; Wu, P.H.; Yogurtcu, O.N.; Rao, P.; DiGiacomo, J.; Godet, I.; He, L.; Lee, M.H.; Gilkes, D.; Sun, S.X.; et al. Collective cancer cell invasion induced by coordinated contractile stresses. Oncotarget 2015, 6, 43438–43451. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Nair, R.; Bhatavdekar, O.; Howe, A.; Salerno, D.; Sempkowski, M.; Josefsson, A.; Pacheco-Torres, J.; Bhujwalla, Z.M.; Gabrielson, K.L.; et al. Transport-driven engineering of liposomes for delivery of alpha-particle radiotherapy to solid tumors: Effect on inhibition of tumor progression and onset delay of spontaneous metastases. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 4246–4258. [Google Scholar] [CrossRef]
- Li, J.Q.; Wu, X.; Gan, L.; Yang, X.L.; Miao, Z.H. Hypoxia induces universal but differential drug resistance and impairs anticancer mechanisms of 5-fluorouracil in hepatoma cells. Acta Pharmacol. Sin. 2017, 38, 1642–1654. [Google Scholar] [CrossRef] [PubMed]
- Leite, E.A.; Lana, A.M.; Junior, A.D.; Coelho, L.G.; De Oliveira, M.C. Acute toxicity study of cisplatin loaded long-circulating and pH-sensitive liposomes administered in mice. J. Biomed. Nanotechnol. 2012, 8, 229–239. [Google Scholar] [CrossRef]
- Rocha, H.L.; Godet, I.; Kurtoglu, F.; Metzcar, J.; Konstantinopoulos, K.; Bhoyar, S.; Gilkes, D.M.; Macklin, P. A persistent invasive phenotype in post-hypoxic tumor cells is revealed by fate mapping and computational modeling. iScience 2021, 24, 102935. [Google Scholar] [CrossRef]
- Kelland, L. The resurgence of platinum-based cancer chemotherapy. Nat. Rev. Cancer 2007, 7, 573–584. [Google Scholar] [CrossRef]
- Pandy, J.G.P.; Balolong-Garcia, J.C.; Cruz-Ordinario, M.V.B.; Que, F.V.F. Triple negative breast cancer and platinum-based systemic treatment: A meta-analysis and systematic review. BMC Cancer 2019, 19, 1065. [Google Scholar] [CrossRef]
- Berry, D.A.; Ueno, N.T.; Johnson, M.M.; Lei, X.; Caputo, J.; Smith, D.A.; Yancey, L.J.; Crump, M.; Stadtmauer, E.A.; Biron, P.; et al. High-dose chemotherapy with autologous hematopoietic stem-cell transplantation in metastatic breast cancer: Overview of six randomized trials. J. Clin. Oncol. 2011, 29, 3224–3231. [Google Scholar] [CrossRef][Green Version]
- Stathopoulos, G.P.; Boulikas, T. Lipoplatin formulation review article. J. Drug Deliv. 2012, 2012, 581363. [Google Scholar] [CrossRef]
- Boulikas, T. Clinical overview on Lipoplatin: A successful liposomal formulation of cisplatin. Expert Opin. Investig. Drugs 2009, 18, 1197–1218. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; He, C.; Kron, S.J.; Lin, W. Nanoparticle formulations of cisplatin for cancer therapy. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2016, 8, 776–791. [Google Scholar] [CrossRef]
- De Luca, A.; Parker, L.J.; Ang, W.H.; Rodolfo, C.; Gabbarini, V.; Hancock, N.C.; Palone, F.; Mazzetti, A.P.; Menin, L.; Morton, C.J.; et al. A structure-based mechanism of cisplatin resistance mediated by glutathione transferase P1-1. Proc. Natl. Acad. Sci. USA 2019, 116, 13943–13951. [Google Scholar] [CrossRef] [PubMed]
- Szturz, P.; Wouters, K.; Kiyota, N.; Tahara, M.; Prabhash, K.; Noronha, V.; Adelstein, D.; Van Gestel, D.; Vermorken, J.B. Low-Dose vs. High-Dose Cisplatin: Lessons Learned From 59 Chemoradiotherapy Trials in Head and Neck Cancer. Front. Oncol. 2019, 9, 86. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normoxic; pH 7.4 (h) (n = 3) | Normoxic; pH 6.8 (h) (n = 3) | Hypoxic; pH 6.8 (h) (n = 3) | |
|---|---|---|---|
| MDA-MB-231 | 36 ± 1 | 42 ± 2 | 41 ± 1 |
| MDA-MB-231-HR (Hypoxia Reporter) | 36 ± 1 * | 41 ± 2 * | 41 ± 2 |
| Size (nm) (n = 6) | PDI (n = 6) | % CDDP Loading (n = 6) | % Contents Retained at pH 7.4 after 6 h (n = 6) | % Contents Retained at pH 6.8 after 6 h (n = 6) | ζ-Potential (mV) (n = 3) | |||
|---|---|---|---|---|---|---|---|---|
| pH 7.4 | pH 6.5 | pH 6 | ||||||
| non-pH-responsive NP (R-A-) (non-responsive NP-CDDP) | 115.5 ± 7.1 | 0.06 ± 0.04 | 3.7 ± 0.3 | 93.0 ± 5.1 | 90.1 ± 8.1 | −3.78 ± 2.63 | −4.28 ± 2.68 | −4.42 ± 2.43 |
| pH-responsive NP (R+A+) (responsive NP-CDDP) | 118.6 ± 3.3 | 0.08 ± 0.06 | 3.5 ± 0.4 | 94.0 ± 7.9 * | 75.1 ± 6.9 * | −2.66 ± 0.40 ** | −0.88 ± 1.03 | −0.70 ± 1.20 ** |
| IC50 Normoxic Conditions pH 7.4 (a) (n = 3) | IC50 Normoxic Conditions pH 6.8 (b) (n = 3) | IC50 Hypoxic Conditions pH 6.8 (c) (n = 3) | Resistance Factor (IC50 Hypoxic/IC50 Normoxic at pH 6.8) (c)/(b) | |
|---|---|---|---|---|
| free CDDP | 9.2 ± 1.2 (10.5 ± 1.4 for HR cells) | 10.1 ± 1.6 | 26.8 ± 2.4 | 2.6 ± 0.5 |
| non-responsive NP-CDDP | NA | NA | NA | NA |
| responsive NP-CDDP | 230.9 ± 8.2 ** | 44.9 ± 7.8 ** | 116.7 ± 9.8 ** | 2.6 ± 0.5 |
| Ce (μg/mL) | Condition | a (μg/mL) | b (h−1) | ln2/b (h) |
|---|---|---|---|---|
| 200 | Normoxia | 0.44 ± 0.01 * | 0.38 ± 0.03 | 1.82 ± 0.13 |
| Hypoxia | 0.36 ± 0.03 * | 0.38 ± 0.04 | 1.85 ± 0.21 | |
| 100 | Normoxia | 0.26 ± 0.01 *# | 0.32 ± 0.02 | 2.16 ± 0.11 |
| Hypoxia | 0.18 ± 0.01 *# | 0.31 ± 0.04 | 2.21 ± 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhatavdekar, O.; Godet, I.; Gilkes, D.; Sofou, S. The Rate of Cisplatin Dosing Affects the Resistance and Metastatic Potential of Triple Negative Breast Cancer Cells, Independent of Hypoxia. Pharmaceutics 2022, 14, 2184. https://doi.org/10.3390/pharmaceutics14102184
Bhatavdekar O, Godet I, Gilkes D, Sofou S. The Rate of Cisplatin Dosing Affects the Resistance and Metastatic Potential of Triple Negative Breast Cancer Cells, Independent of Hypoxia. Pharmaceutics. 2022; 14(10):2184. https://doi.org/10.3390/pharmaceutics14102184
Chicago/Turabian StyleBhatavdekar, Omkar, Inês Godet, Daniele Gilkes, and Stavroula Sofou. 2022. "The Rate of Cisplatin Dosing Affects the Resistance and Metastatic Potential of Triple Negative Breast Cancer Cells, Independent of Hypoxia" Pharmaceutics 14, no. 10: 2184. https://doi.org/10.3390/pharmaceutics14102184
APA StyleBhatavdekar, O., Godet, I., Gilkes, D., & Sofou, S. (2022). The Rate of Cisplatin Dosing Affects the Resistance and Metastatic Potential of Triple Negative Breast Cancer Cells, Independent of Hypoxia. Pharmaceutics, 14(10), 2184. https://doi.org/10.3390/pharmaceutics14102184