
Heme Oxygenase-1 Exerts Antiviral Activity against Hepatitis A Virus In Vitro

 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Cell Lines, Viruses, and Chemicals
2.2. Cell Cytotoxicity Assay
2.3. Reverse Transcription-Quantitative Polymerase Chain Reaction
2.4. Plasmid Construction and Transfection
2.5. Western Blot Analysis
2.6. Statistical Analysis
3. Results
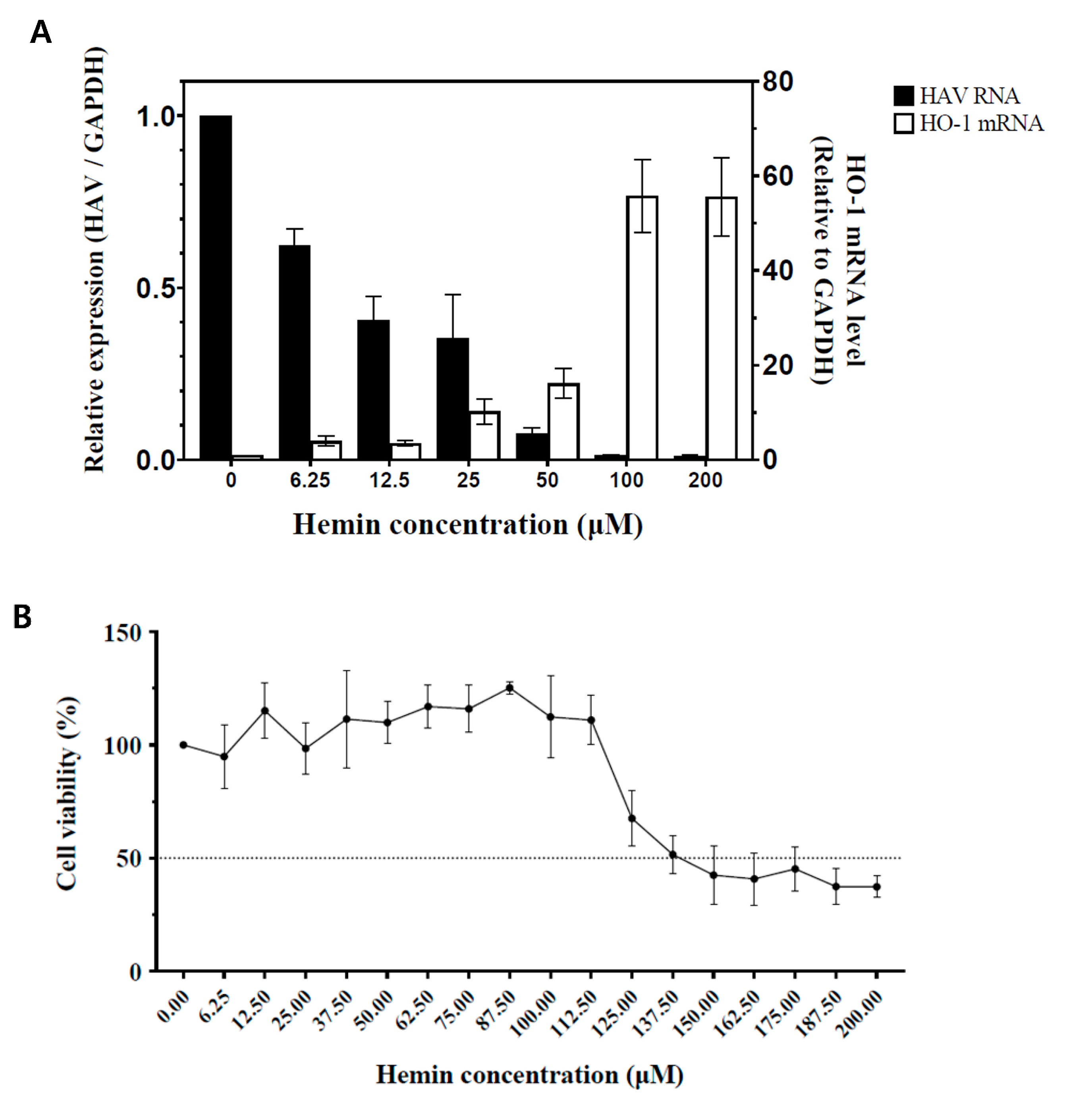
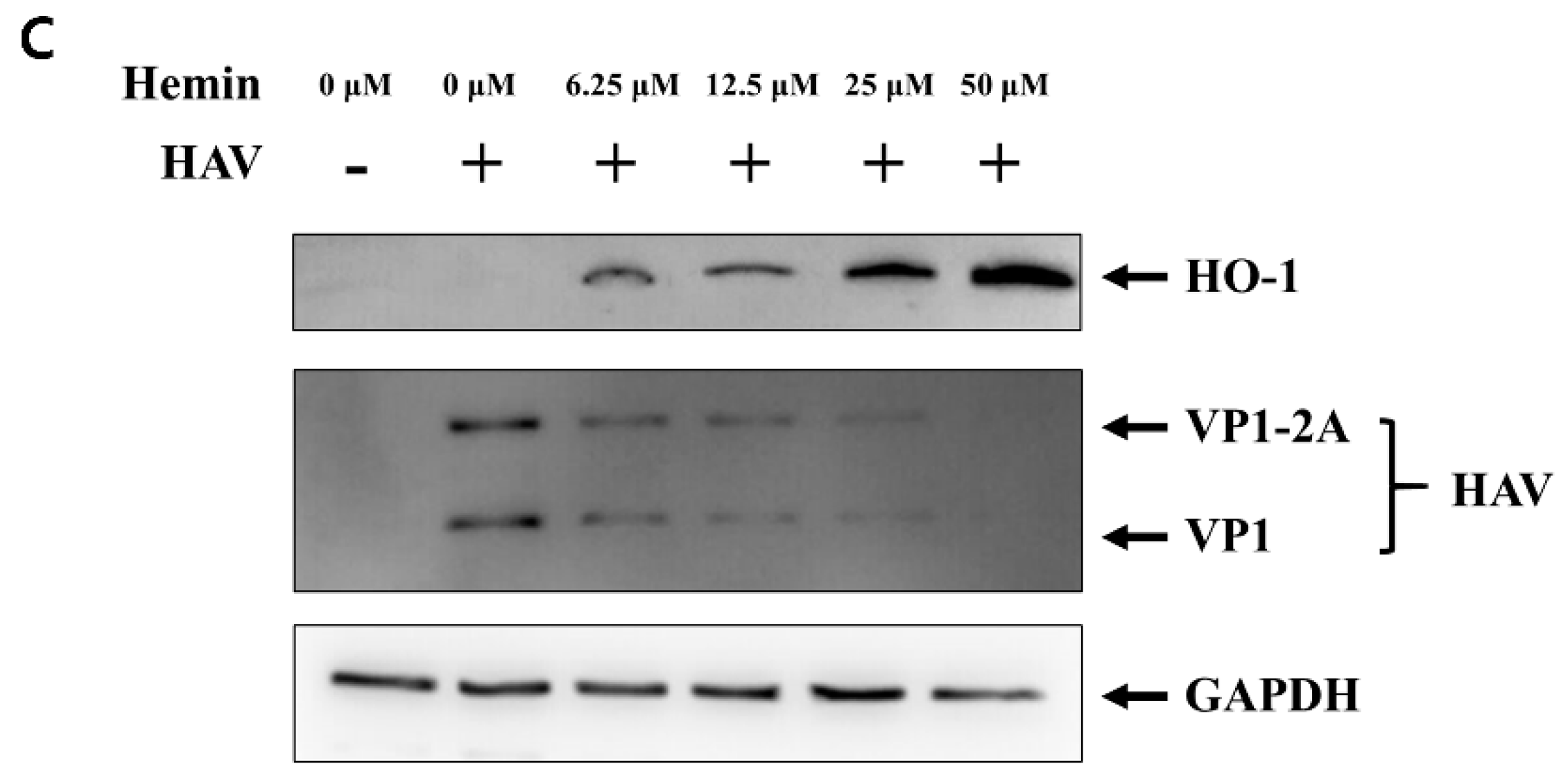
3.1. Inhibition of HAV Replication by Hemin-Mediated Upregulation of HO-1
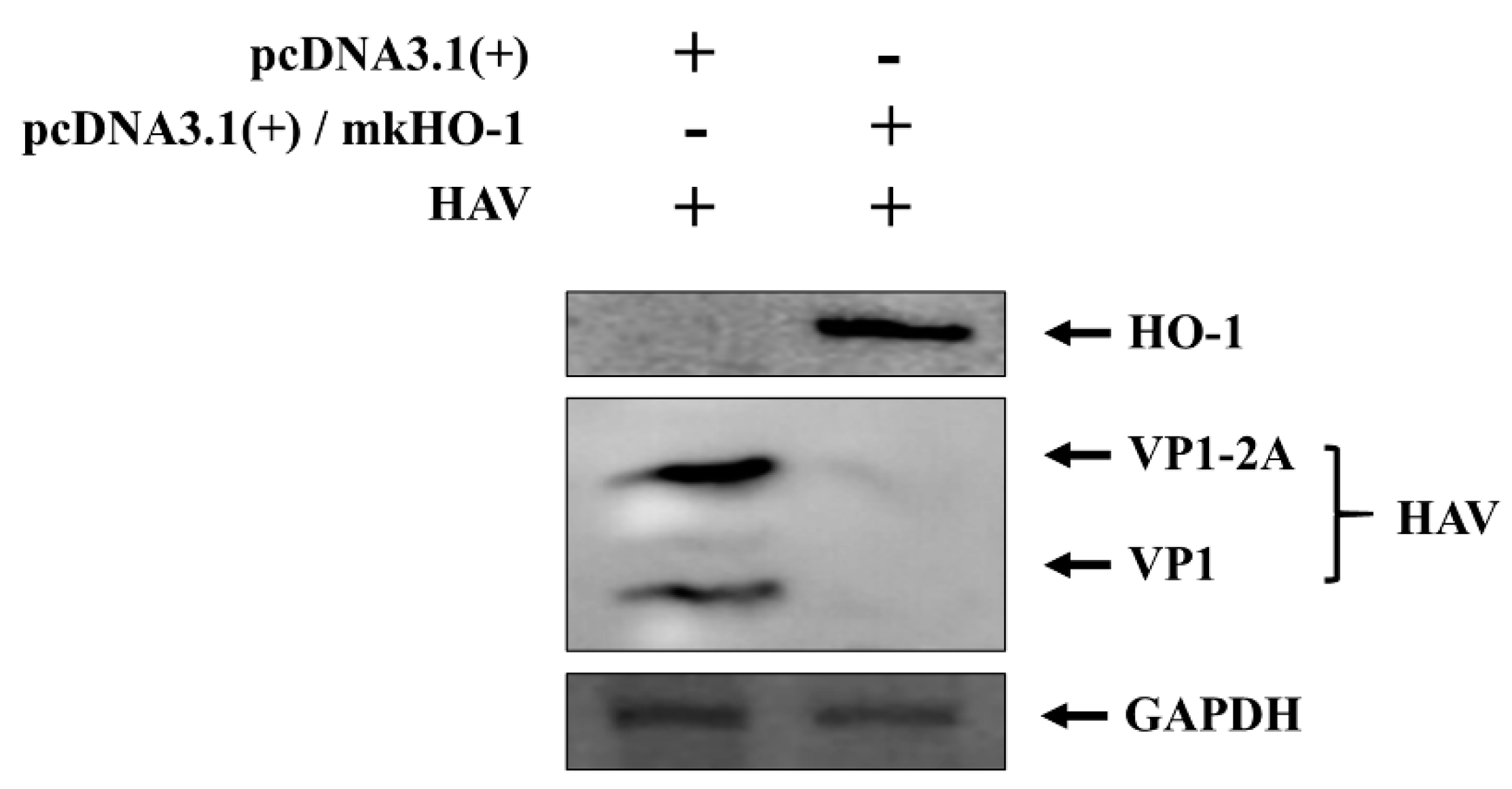
3.2. Reduction of the HAV Viral Protein through HO-1 Overexpression
3.3. Inhibition of HAV Caused by Enzymatic Activity of HO-1
3.4. Suppression of HAV Replication by Metabolites of Heme Produced through HO-1 Catalysis
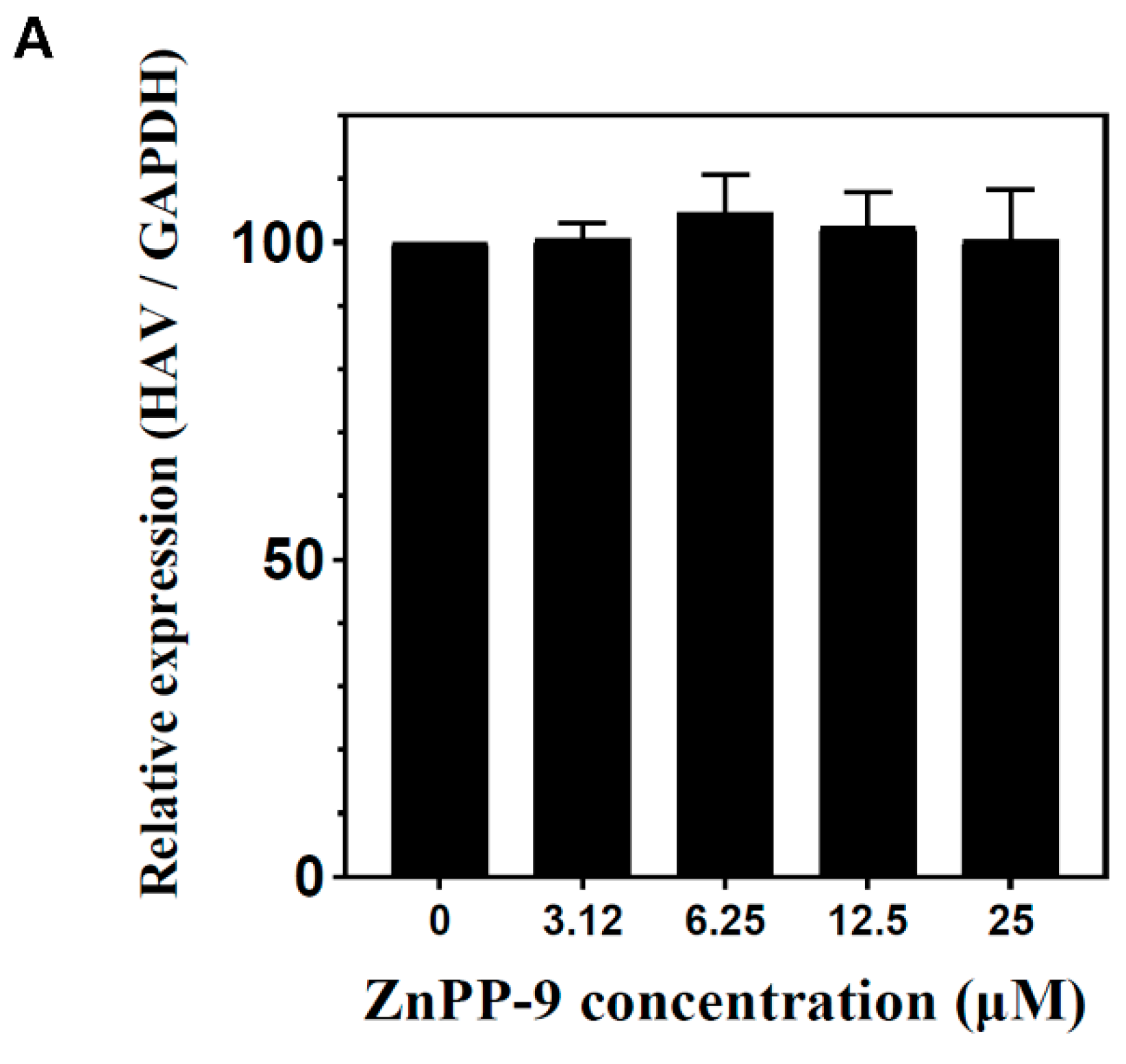
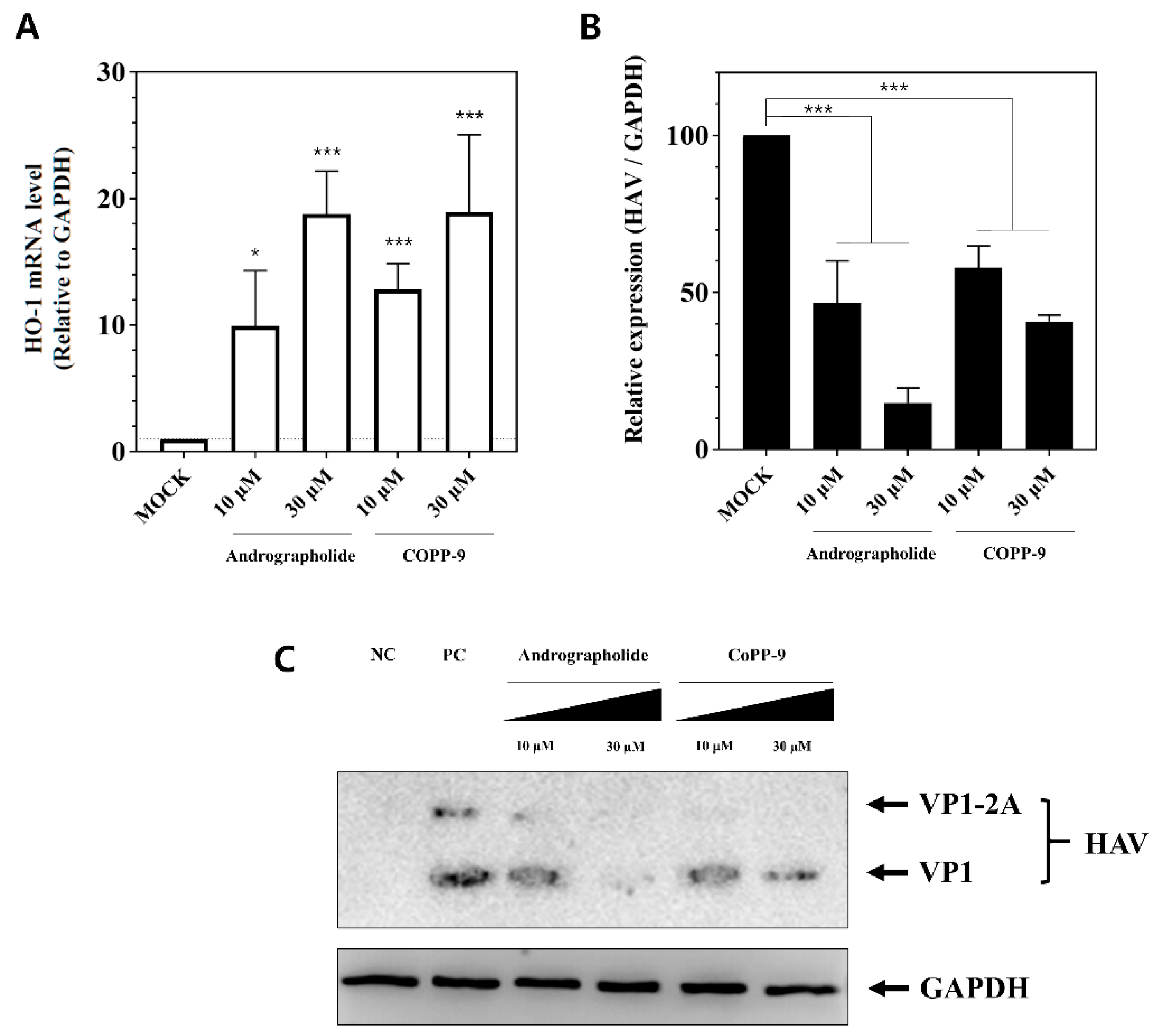
3.5. Suppression of HAV Replication by Other HO-1 Inducers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cohen, J.I.; Ticehurst, J.R.; Purcell, R.H.; Buckler-White, A.; Baroudy, B.M. Complete nucleotide sequence of wild-type hepatitis A virus: Comparison with different strains of hepatitis A virus and other picornaviruses. J. Virol. 1987, 61, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Hensley, L.; McKnight, K.L.; Hu, F.; Madden, V.; Ping, L.; Jeong, S.H.; Walker, C.; Lanford, R.E.; Lemon, S.M. A pathogenic picornavirus acquires an envelope by hijacking cellular membranes. Nature 2013, 496, 367–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nainan, O.V.; Xia, G.; Vaughan, G.; Margolis, H.S. Diagnosis of hepatitis A virus infection: A molecular approach. Clin. Microbiol. Rev. 2006, 19, 63–79. [Google Scholar] [CrossRef] [Green Version]
- Daniels, D.; Grytdal, S.; Wasley, A. Surveillance for Acute Viral Hepatitis—United States, 2007. Morb. Mortal. Wkly. Rep. Surveill. Summ. 2009, 58, 1–27. [Google Scholar]
- Feng, Z.; Lemon, S.M. Peek-a-boo: Membrane hijacking and the pathogenesis of viral hepatitis. Trends Microbiol. 2014, 22, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughan, G.; Goncalves Rossi, L.M.; Forbi, J.C.; de Paula, V.S.; Purdy, M.A.; Xia, G.; Khudyakov, Y.E. Hepatitis A virus: Host interactions, molecular epidemiology and evolution. Infect. Genet. Evol. 2014, 21, 227–243. [Google Scholar] [CrossRef]
- Wilson, T.R. The abcs of hepatitis. Nurse Pract. 2005, 30, 2000. [Google Scholar] [CrossRef]
- FitzSimons, D.; Hendrickx, G.; Vorsters, A.; Van Damme, P. Hepatitis A and E: Update on prevention and epidemiology. Vaccine 2010, 28, 583–588. [Google Scholar] [CrossRef]
- Collier, M.G.; Khudyakov, Y.E.; Selvage, D.; Adams-Cameron, M.; Epson, E.; Cronquist, A.; Jervis, R.H.; Lamba, K.; Kimura, A.C.; Sowadsky, R.; et al. Outbreak of hepatitis A in the USA associated with frozen pomegranate arils imported from Turkey: An epidemiological case study. Lancet Infect. Dis. 2014, 14, 976–981. [Google Scholar] [CrossRef]
- States, M.; Strategic, W.H.O.; Group, A.; Technical, N.; Groups, A.; Assembly, W.H. WHO position paper on hepatitis A vaccines—June 2012 = Note de synthèse: Position de l’OMS concernant les vaccins contre l’hépatite A—Juin 2012. Wkly. Epidemiol. Rec. 2012, 87, 28–29. [Google Scholar]
- da Silva, K.M.; Ferreira, J.D.S.; Carvalho, A.; Gomes, D.D.S.; Cavalcanti, M.D.S.; Ferreira-Júnior, G.C.; Matos-Rocha, T.J. Epidemiological profile of viral hepatitis infection in the population treated at a reference hospital in Alagoas. Braz. J. Biol. 2021, 82, e238431. [Google Scholar] [CrossRef]
- Vilaplana, T.G.; Leeman, D.; Balogun, K.; Ngui, S.L.; Phipps, E.; Khan, W.M.; Team, I.; Balasegaram, S.; Burns, A.; Sims, B.; et al. Hepatitis A outbreak associated with consumption of dates, England and Wales, January 2021 to April 2021. Eurosurveillance 2021, 26, 2100432. [Google Scholar] [CrossRef]
- Elbahrawy, A.; Ibrahim, M.K.; Eliwa, A.; Alboraie, M.; Madian, A.; Aly, H.H. Current situation of viral hepatitis in Egypt. Microbiol. Immunol. 2021. [Google Scholar] [CrossRef]
- Ramachandran, S.; Xia, G.L.; Dimitrova, Z.; Lin, Y.; Montgomery, M.; Augustine, R.; Kamili, S.; Khudyakov, Y. Changing molecular epidemiology of Hepatitis A virus infection, United States, 1996–2019. Emerg. Infect. Dis. 2021, 27, 1742–1745. [Google Scholar] [CrossRef] [PubMed]
- Global Hepatitis Report, 2017; World Health Organization: Geneva, Switzerland, 2017; ISBN 9789241565455.
- Otterbein, L.E.; Soares, M.P.; Yamashita, K.; Bach, F.H. Heme oxygenase-1: Unleashing the protective properties of heme. Trends Immunol. 2003, 24, 449–455. [Google Scholar] [CrossRef]
- Waza, A.A.; Hamid, Z.; Ali, S.; Bhat, S.A.; Bhat, M.A. A review on heme oxygenase-1 induction: Is it a necessary evil. Inflamm. Res. 2018, 67, 579–588. [Google Scholar] [CrossRef]
- Zhang, A.; Wan, B.; Jiang, D.; Wu, Y.; Ji, P.; Du, Y.; Zhang, G. The Cytoprotective Enzyme Heme Oxygenase-1 Suppresses Pseudorabies Virus Replication in vitro. Front. Microbiol. 2020, 11, 412. [Google Scholar] [CrossRef]
- Zhu, Z.; Wilson, A.T.; Mathahs, M.; Wen, F.; Brown, K.E.; Luxon, B.A.; Schmidt, W.N. Heme oxygenase-1 suppresses hepatitis C virus replication and increases resistance of hepatocytes to oxidant injury. Hepatology 2008, 48, 1430–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryter, S.W.; Alam, J.; Choi, A.M.K. Heme oxygenase-1/carbon monoxide: From basic science to therapeutic applications. Physiol. Rev. 2006, 86, 583–650. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.A.; González, P.A.; Kalergis, A.M. Modulation of Antiviral Immunity by Heme Oxygenase-1. Am. J. Pathol. 2017, 187, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Hill-Batorski, L.; Halfmann, P.; Neumann, G.; Kawaoka, Y. The Cytoprotective Enzyme Heme Oxygenase-1 Suppresses Ebola Virus Replication. J. Virol. 2013, 87, 13795–13802. [Google Scholar] [CrossRef] [Green Version]
- Reichard, J.F.; Motz, G.T.; Puga, A. Heme oxygenase-1 induction by NRF2 requires inactivation of the transcriptional repressor BACH1. Nucleic Acids Res. 2007, 35, 7074–7086. [Google Scholar] [CrossRef] [Green Version]
- Gharbi-Khelifi, H.; Ferre, V.; Sdiri, K.; Berthome, M.; Fki, L.; Harrath, R.; Billaudel, S.; Aouni, M. Hepatitis A in Tunisia: Phylogenetic analysis of hepatitis A virus from 2001 to 2004. J. Virol. Methods 2006, 138, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Zhang, A.; Zhang, C.; Ni, H.; Gao, J.; Wang, C.; Zhao, Q.; Wang, X.; Wang, X.; Ma, C.; et al. Heme oxygenase-1 acts as an antiviral factor for porcine reproductive and respiratory syndrome virus infection and over-expression inhibits virus replication in vitro. Antiviral Res. 2014, 110, 60–69. [Google Scholar] [CrossRef]
- Lei, A.H.; Zhang, G.H.; Tian, R.R.; Zhu, J.W.; Zheng, H.Y.; Pang, W.; Zheng, Y.T. Replication potentials of HIV-1/HSIV in PBMCs from northern pig-tailed macaque (Macaca leonina). Dongwuxue Yanjiu 2014, 35, 186–195. [Google Scholar] [CrossRef]
- Mittal, S.P.K.; Khole, S.; Jagadish, N.; Ghosh, D.; Gadgil, V.; Sinkar, V.; Ghaskadbi, S.S. Andrographolide protects liver cells from H2O2 induced cell death by upregulation of Nrf-2/HO-1 mediated via adenosine A2a receptor signalling. Biochim. Biophys. Acta-Gen. Subj. 2016, 1860, 2377–2390. [Google Scholar] [CrossRef]
- Bharucha, A.E.; Kulkarni, A.; Choi, K.M.; Camilleri, M.; Lempke, M.; Brunn, G.J.; Gibbons, S.J.; Zinsmeister, A.R.; Farrugia, G. First-in-Human Study Demonstrating Pharmacological Activation of Heme Oxygenase-1 in Humans. Clin. Pharmacol. Ther. 2010, 87, 187–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touret, F.; Gilles, M.; Barral, K.; Nougairède, A.; van Helden, J.; Decroly, E.; de Lamballerie, X.; Coutard, B. In vitro screening of a FDA approved chemical library reveals potential inhibitors of SARS-CoV-2 replication. Sci. Rep. 2020, 10, 13093. [Google Scholar] [CrossRef] [PubMed]
- Vzorov, A.N.; Dixon, D.W.; Trommel, J.S.; Marzilli, L.G.; Compans, R.W. Inactivation of human immunodeficiency virus type 1 by porphyrins. Antimicrob. Agents Chemother. 2002, 46, 3917–3925. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Nguyen, X.; Ou, J.; Rekulapelli, P.; Stevenson, D.K.; Dennery, P.A. Unique effects of zinc protoporphyrin on HO-1 induction and apoptosis. Blood 2001, 97, 1306–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; He, Z.; Wu, L.; Fang, Y. Effects of induction/inhibition of endogenous heme oxygenase-1 on lipid metabolism, endothelial function, and atherosclerosis in rabbits on a high fat diet. J. Pharmacol. Sci. 2012, 118, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez, F.J.; Farías, M.A.; Retamal-Díaz, A.; Espinoza, J.A.; Kalergis, A.M.; González, P.A. Pharmacological induction of heme oxygenase-1 impairs nuclear accumulation of herpes simplex virus capsids upon infection. Front. Microbiol. 2017, 8, 2108. [Google Scholar] [CrossRef] [PubMed]
- Fillebeen, C.; Rivas-Estilla, A.M.; Bisaillon, M.; Ponka, P.; Muckenthaler, M.; Hentze, M.W.; Koromilas, A.E.; Pantopoulos, K. Iron Inactivates the RNA Polymerase NS5B and Suppresses Subgenomic Replication of Hepatitis C Virus. J. Biol. Chem. 2005, 280, 9049–9057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sass, G.; Barikbin, R.; Tiegs, G. The multiple functions of heme oxygenase-1 in the liver. Z. Gastroenterol. 2012, 50, 34–40. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | FRhK-4 CC50:159.47 μM | |
|---|---|---|
| EC50 | SI 1 | |
| HAV (HM-175/18f) | 10.12 μM | 15.76 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.-H.; Ahn, H.-S.; Go, H.-J.; Kim, D.-Y.; Kim, J.-H.; Lee, J.-B.; Park, S.-Y.; Song, C.-S.; Lee, S.-W.; Choi, I.-S. Heme Oxygenase-1 Exerts Antiviral Activity against Hepatitis A Virus In Vitro. Pharmaceutics 2021, 13, 1229. https://doi.org/10.3390/pharmaceutics13081229
Kim D-H, Ahn H-S, Go H-J, Kim D-Y, Kim J-H, Lee J-B, Park S-Y, Song C-S, Lee S-W, Choi I-S. Heme Oxygenase-1 Exerts Antiviral Activity against Hepatitis A Virus In Vitro. Pharmaceutics. 2021; 13(8):1229. https://doi.org/10.3390/pharmaceutics13081229
Chicago/Turabian StyleKim, Dong-Hwi, Hee-Seop Ahn, Hyeon-Jeong Go, Da-Yoon Kim, Jae-Hyeong Kim, Joong-Bok Lee, Seung-Yong Park, Chang-Seon Song, Sang-Won Lee, and In-Soo Choi. 2021. "Heme Oxygenase-1 Exerts Antiviral Activity against Hepatitis A Virus In Vitro" Pharmaceutics 13, no. 8: 1229. https://doi.org/10.3390/pharmaceutics13081229
APA StyleKim, D.-H., Ahn, H.-S., Go, H.-J., Kim, D.-Y., Kim, J.-H., Lee, J.-B., Park, S.-Y., Song, C.-S., Lee, S.-W., & Choi, I.-S. (2021). Heme Oxygenase-1 Exerts Antiviral Activity against Hepatitis A Virus In Vitro. Pharmaceutics, 13(8), 1229. https://doi.org/10.3390/pharmaceutics13081229