1. Introduction
Targeted radionuclide therapy (TRT) is an attractive approach to treat cancer because it allows the specific irradiation of tumour cells. As, theoretically, TRT spares healthy tissues that do not express high levels of the targeted receptor, it represents a method of choice for treating diffuse and metastatic disease [
1,
2]. Historically, radiopharmaceuticals administered to treat patients have been based on beta-particle emitters, such as iodine-131 (
131I) in thyroid carcinoma and phosphorus-32 (
32P) in ovarian cancer. Later, [
90Y]Y-ibritumomab tiuxetan (Zevalin) and [
131I]I-tositumomab (Bexxar) were approved for radioimmunotherapy of non-Hodgkin B lymphoma. More recently, [
177Lu]Lu-DOTATATE was also approved for therapy of neuroendocrine tumours [
3], [
177Lu]Lu-PSMA-617 for metastatic castration-resistant prostate cancer [
4], and many clinical trials are currently assessing this radiopharmaceutical. Beta-particle emitters are largely available, at reasonable costs, and they are easy to chelate. However, although their relatively long range (≥1 mm) can counterbalance heterogeneity in radiopharmaceutical tumour distribution, it can also cause non-specific irradiation of healthy cells/tissues, thus limiting their interest for TRT. Moreover, beta particles are low linear energy transfer (LET) particles (like X and gamma rays). This means that they have a low ionising power and produce simple lesions in cells that can be repaired. Therefore, they can show limited efficacy against radioresistant solid tumours. This can be overcome by using high LET alpha particles, such as those emitted by radium-223 or by actinium-225, which have been tested for the management of prostate-cancer bone metastases. In addition, alpha-particle emitters decay according to a chain of disintegration, leading to several daughters that enhance therapeutic efficacy. Some questions remain, such as the choice of radionuclides, the stability of chelation, and the fate of daughters. Moreover, due to their range in matter (50–100 µm), alpha particles can also cause some non-specific irradiation. Auger electron emitters (AEE
S), another class of therapeutic radionuclides, also have generated much interest and could mitigate the non-specific irradiation issue. Auger electrons are emitted in cascades by atoms decaying by electron capture and/or conversion processes that usually produce vacancies in K shells. These vacancies are filled by electrons dropping from the outer shells. These transitions are accompanied by a release of energy that can be transferred to electrons of external layers emitted in cascades (Auger electron cascades) or emitted as fluorescent X-rays.
The energy of Auger electrons can reach tens of keV, but most of them have a very low energy (<1 keV) that is released in a sphere of few cubic nanometres. Therefore, they are considered medium to high linear energy transfer (LET) particles (4–26 keV/µm). These high energy deposits produce locally dense ionisations that are very destructive when the radionuclide is incorporated in nuclear DNA, [
5,
6,
7,
8] the cell nucleus [
9,
10,
11,
12,
13], or the cell membrane [
14,
15,
16,
17,
18,
19]. In a context where specific nuclear DNA targeting is still challenging, our group previously showed that AEE-based TRT with
125I-labelled non-internalising monoclonal antibodies (mAbs) against carcinoembryonic antigen (CEA) leads to significant tumour growth delay in preclinical mouse models harbouring small tumours [
18]. While nuclear targeting directly mediates the DNA damage response (DDR) leading to cell death, Auger electron cascades produced at the cell membrane induce the local formation of ceramide-enriched large domains (lipid rafts) that activate signalling pathways involving NF-kB and ultimately, leading to reactive oxygen species (ROS) production, nuclear damage, and DDR activation [
15,
19]. We also demonstrated that bystander effects induced by nuclear (5-[
125I]iodo-2′-deoxyuridine, [
125I]I-UdR) and cell membrane (non-internalising
125I-mAbs) irradiation contribute significantly to cell death [
13,
15]. Notably, we showed that, after incubation with the DNA base analogue [
125I]I-UdR, 30–35% of cells were killed by bystander effects and 50% of cells by the direct effect of Auger electrons.
AEEs could be a precious asset for TRT, but there has not been, so far, any convincing candidate, although about 50% of radionuclides emit AEs. Indeed, a major advantage of AEE-based TRT is to decrease the non-specific irradiation of healthy tissues, while benefiting from Auger electrons’ high LET features. However, due to their short range and lack of cross-fire irradiation in tumours, AAE-based TRT might require the administration of high activities or repeated administrations to counterbalance the radionuclide distribution heterogeneity that would not allow reaching all tumour cells after one single administration. Therefore, ideal AEE should not emit (or may emit only in very low proportion) other radiation types (γ, β, CE), responsible for non-specific irradiations, as these would overwhelm AEE-TRT’s benefits. AEE also need to locally release high amount of energy, a parameter that is influenced by the number of Auger electrons released per decay and their average energy. Iodine-125, which emits about 20 Auger electrons/decay and releases low amount of energy through the emission of conversion electrons (CE) or photons, is a very attractive AEE candidate. Iodine-125 radiopharmaceuticals have been developed ([
125I]I-UdR or mAbs conjugates) [
20,
21,
22,
23], but their clinical use is limited by
125I’s long physical half-life (t
1/2(Phys) = 59.4 days). A comprehensive list of AEE candidates has recently been reviewed in [
24]. Platinum-193 m (t
1/2(phys) = 4.3 days) and platinum-195 m (t
1/2(phys) = 4.0 days) also are very attractive AEEs because they emit a very high number of Auger electrons (between 20 and 30) per decay and photons of intermediate energy (66 and 76 keV) than can be used for imaging. However, they are still not routinely available, and the radiolabelling of biological vectors is still challenging. Many studies have used indium-111 because it has a short physical half-life (2.8 days), is easily conjugated to macromolecules, and is easily produced [
25]. However,
111In is a poor AEE with about 7–8 Auger electrons per decay, emission of energetic CEs (0.16 per decay; average energy = 176 keV) and γ-rays (1.85 energetic photons of 171.28 and 245.35 keV per decay) that can cause non-specific irradiation and raise radiation protection concerns, respectively.
Another difficulty with AEEs is the need to build radioimmunoconjugates (RIC) that can specifically target cancer cells and also drive activity at least into the nucleus, if not into DNA. Indeed, as Auger electrons with high LET have a range of few nm, nuclear localisation without incorporation into DNA would lead to a significant decrease in efficacy, although the results might still be acceptable.
Herein, we assessed two new
111In-RICs based on the anti-HER2 antibody trastuzumab functionalised with two cationic peptides (NLS and TAT) harbouring a nuclear localising sequence. Pioneering groups have shown that these peptides allow the nuclear transportation of the functionalised antibodies in AEE-based TRT settings [
11,
26,
27]. Although, other authors highlighted the need for alternative approaches to improve their delivery [
28,
29,
30], none of these approaches has been evaluated beyond the preclinical stage. Moreover, only one study investigated [
111In]In-DTPA-human epidermal growth factor in patients [
31] but did not confirm the promising results obtained in animal models [
28]. In this study, we evaluated whether NLS-/TAT-immunoconjugates are suitable vectors to specifically deliver AEE-based TRT to the cancer cell nucleus. We hypothesised that the
111In-trastuzumab RIC specifically targets HER2-positive cancer cells before internalization in the cell cytoplasm. When conjugated to NLS or TAT peptides, the nuclear localisation signal should drive this RIC through the nuclear pore complex to enhance Auger electron cytotoxicity. To test this hypothesis, we thoroughly investigated the subcellular localisation, activity uptake, and cytotoxic effects of the developed
111In-RICs.
2. Materials and Methods
2.1. Cell Lines
A-431 vulvar squamous carcinoma and SK-OV-3 ovarian carcinoma cells were obtained from the American Type Culture Collection (ATCC, Rockville, MD, USA). HER1- and HER2-expressing A-431 cells were transfected with the genes encoding CEA and luciferase to obtain the A-431
CEA-luc cell line, as previously described [
14]. HER1- and HER2-expressing SK-OV-3 cells were transfected with the gene encoding CEA to obtain the SK-OV-3
1B9 cell line. All cell lines were grown in DMEM supplemented with 10% heat-inactivated foetal bovine serum, 1% penicillin/streptomycin, and 200 µg/mL geneticin. Hygromycin (100 µg/mL) was added to the medium for A-431
CEA-luc cells. Cell lines were routinely tested for mycoplasma contamination using the MycoTect assay from Life technologies (Thermo Fisher Scientific, Waltham, MA, USA).
2.2. Antibodies and Peptides
Trastuzumab (
Herceptin, Roche, Basel, Switzerland) is a humanised IgG1k mAb against the HER2 receptor. A humanised anti-AMHRII IgG1 mAb, 3C23K, was used as an irrelevant mAb, derived from the murine anti-AMHRII mAb 12G4 [
32].
The 16-mer TAT peptide [GRKKRRQRRRPPQGYG] harbouring the nuclear localisation sequence (underlined) of the HIV-1 tat protein was synthesised and provided by Genscript Biotech (Leiden, The Netherlands) with a purity ≥ 90%. The 13-mer Simian Virus 40 T-Ag-derived NLS peptide [CGYGPKKKRKVGG] (the nuclear localisation sequence is underlined) was synthesised and provided by Proteogenix (Schiltigheim, France) with a purity of 90.40%. Both peptides were redissolved in chelexed PBS (pH 7.2) at a concentration of 7 mg/mL and kept at −80 °C until use.
2.3. Bioconjugation, Radiolabelling, Purification and Quality Control
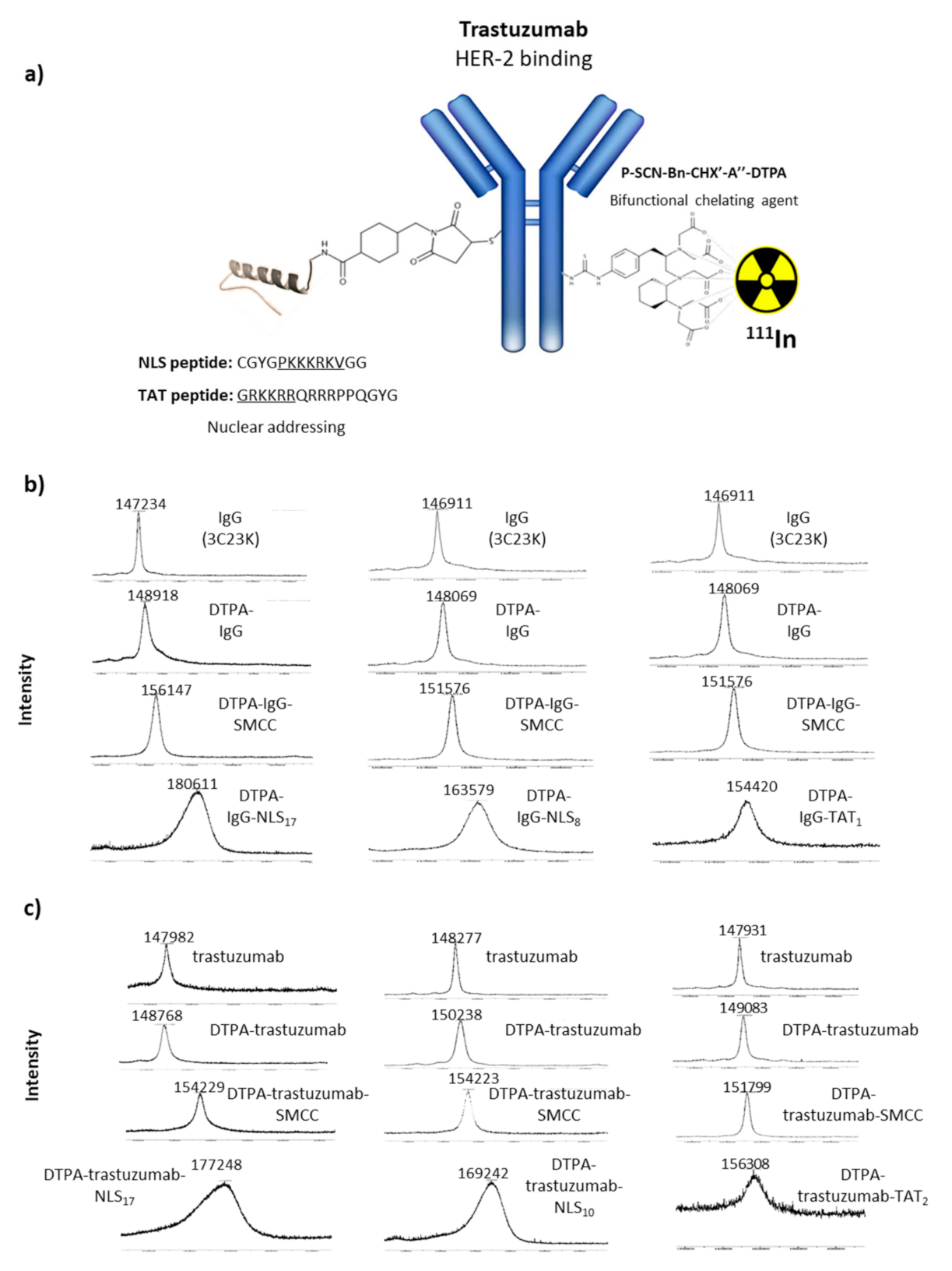
Trastuzumab and the irrelevant humanised IgG (3C23K) were conjugated to p-SCN-Bn-CHX-A″-DTPA ([(R)-2-amino-3-(4-isothiocyanatophenyl)propyl]-trans-(S,S)-cyclohexane-1,2-diamine-pentaacetic acid), provided by Macrocyclics (Plano, TX, USA), a chelating agent allowing further radiolabelling with 111In. Briefly, the pH of the trastuzumab and IgG solutions (2–8 mg in chelexed PBS; 8–12 mg/mL) was adjusted to 8.4 using a 0.2 M chelexed Na2CO3 solution (pH 10). A 15-fold molar excess of p-SCN-Bn-CHX-A″-DTPA (25 mg/mL in DMSO) was added drop by drop (2 µL) to prevent precipitation. Bioconjugation was performed at 37 °C for 90 min, followed by purification on an Amicon 30 kDa (Merck Millipore, Molsheim, France) and chelexed PBS washes (pH 7.2).
The NLS and TAT peptides were conjugated to DTPA-modified trastuzumab or IgG using the heterobifunctional cross-linking agent sulphosuccinimidyl-4-(N-maleimidomethyl)cyclohexane-1-carboxylate (sulpho-SMCC; Thermo Fisher Scientific, Rockford, IL, USA). Briefly, DTPA-trastuzumab or DTPA-IgG (chelexed PBS, pH 7.2) was mixed with 25- to 50-fold molar excess of sulpho-SMCC dissolved to 2.34 mg/mL in Ultra Trace Elemental Analysis Grade water (Thermo Fisher Scientific, Illkirch-Graffenstaden, France) at room temperature (RT) for 60 min. The obtained activated-ester DTPA-trastuzumab or IgG conjugates were transferred to Amicon 30 kDa (Merck Millipore, Burlington, MA, USA), purified, and concentrated to 3–7 mg/mL with chelexed PBS (pH 7.5). Then, conjugates were mixed with 40- to 60-fold molar excess of NLS or TAT peptides (7 mg/mL in chelexed PBS [pH 7.2]) at 37 °C for 90 min. The final conjugates were transferred to Amicon 30 kDa (Merck Millipore), and the PBS buffer was exchanged to ammonium acetate (0.25 M, pH 5.5) with two consecutive washes.
Matrix-assisted laser desorption ionisation–time of flight (MALDI–TOF) was used to determine the number of DTPA, SMCC, NLS, or TAT moieties per antibody. The m/z difference between the native mAbs and their conjugates was divided by the molecular weight of the added functional group (DTPA, sulpho-SMCC, or peptide) to find the chelate/peptide-to-antibody ratio.
HER2-specific immunoconjugates (DTPA-trastuzumab, DTPA-trastuzumab-NLS, and DTPA-trastuzumab-TAT) (
Figure 1a) and irrelevant immunoconjugates (DTPA-IgG, DTPA-IgG-NLS, and DTPA-IgG-TAT) were labelled with indium-111 ([
111In]InCl
3) to obtain a specific activity of 370 MBq/mg. Typically, 119 µL (100 MBq) of [
111In]InCl
3 (in HCl 0.05 N; Curium, Paris, France) were mixed with 119 µL 0.25 M ammonium acetate (pH 5.5) to stabilise the pH at 5.5. Then, 270 µg (volume (µL), depending on the concentration) of immunoconjugate were added at 37 °C for 90 min. Radiolabelling yields and radiochemical purity were determined by instant thin-layer silica-gel chromatography (iTLC, Montpellier, France) with 50 mM EDTA (pH 5.5) as eluant. RICs were purified using a desalting PD-10 column and eluted with PBS. All RICs were successfully obtained, with radiochemical yields between 80 and 98% and specific activities ranging from 310 to 380 MBq/mg.
2.4. Immunoconjugates Cell Binding and HER2 Cell Expression
Immunoconjugate cell binding was assessed by flow cytometry. Cells were fixed with 4% (v/v) paraformaldehyde in PBS at RT for 15 min, permeabilised with Triton X-100 (0.5% in PBS) at RT for 15 min, and blocked with PBS/bovine serum albumin (BSA) blocking solution (10 mg/mL) at 37 °C for 1 h. Then, cells were incubated with each immunoconjugate (10 µg/mL) at RT for 1 h, followed by incubation with an AlexaFluor-488-conjugated anti-human secondary antibody (1:200; (F(ab′)2-goat anti-human IgG (H+L) Cross-Adsorbed, Thermo Fisher Scientific) at RT in the dark for 1 h. Cells were then washed three times and suspended in PBS for analysis using a Gallios cytometer (Beckman Coulter, Villepinte, France).
2.5. RIC Stability
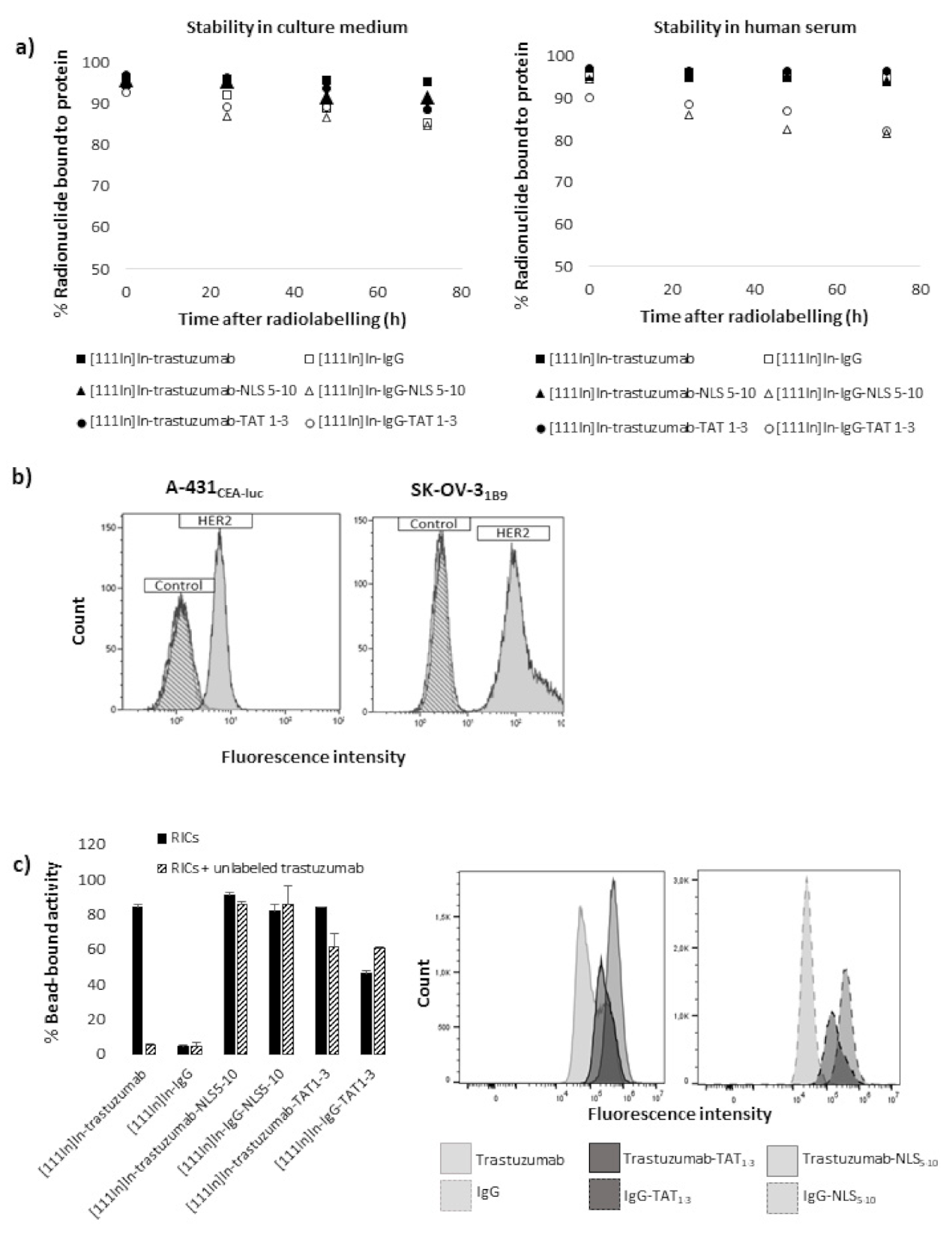
A 100 µL aliquot of each radiopharmaceutical was added to 900 µL of culture medium or human serum. Then, samples were incubated on a shaker at 37 °C. At the desired time points, the radiopharmaceutical stability was evaluated by iTLC using a 50 mM EDTA (pH 5.5) solution as eluant. The retention factor (Rf) of the RIC was 0 and that of free indium was 1. Each experiment was repeated twice and the percentage of radionuclides bound to proteins for each RIC was calculated after measurement with a γ-ray counter.
2.6. Determination of the RIC Immunoreactive Fraction
RIC immunoreactive fractions were assessed using a bead-based assay method described by Sharma et al. [
33]. Briefly, 20 µL of the magnetic HisPur Ni-NTA bead slurry (Thermo Scientific) was incubated with 1 µg His-Tag human recombinant HER2 protein (Sino Biological). Coated beads were then incubated with 1 ng of RIC in the presence (or not) of unlabelled trastuzumab (blocked condition; ×1000-fold excess) on a shaker at RT for 30 min. Beads were isolated, and the supernatant containing unbound RICs was collected. Next, beads were washed twice with PBS-Tween (0.05%) to remove non-specifically bound RICs. Finally, the activity in each fraction (beads alone, supernatant, and wash) was measured using a γ-ray counter, and the immunoreactive fractions were calculated.
2.7. Clonogenic Assays
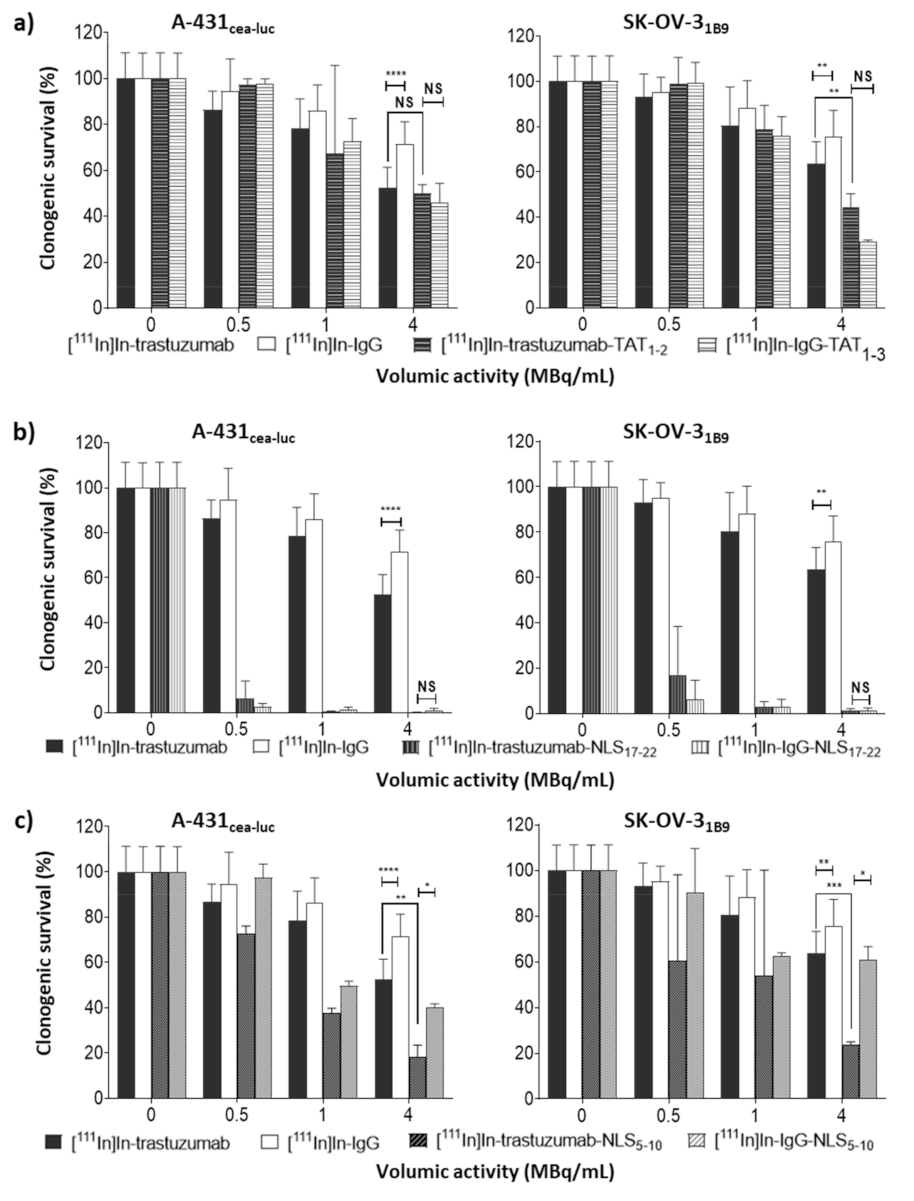
Cell survival after incubation with 111In-RICs or unlabelled immunoconjugates was assessed using a standard clonogenic assay. Briefly, a defined number of cells, from 100 to 800, were plated in 6-well plates in 2 mL of fresh culture medium. The next day, cells were incubated with an increasing amount of activities of 111In-RICs (0 to 4 MBq/mL) or 10.8 µg/mL unlabelled immunoconjugates at 37 °C/5% CO2 for 48 h, followed by three washes with 2 mL PBS and the addition of fresh medium (2 mL). Then, cells were cultured for 12 days, and colonies were fixed (acetic acid:methanol [1:3]) and stained with Giemsa diluted in H2O (1:5; Sigma-Aldrich, Saint-Quentin Fallavier, France). Colonies were manually counted, and the surviving fraction was calculated. All experiments were repeated at least three times in triplicate for each condition.
2.8. In Vitro Measurement of DNA Damage
Cells were seeded on coverslips in 6-well plates. The next day, cells were incubated with 4 MBq/mL of HER2-specific or irrelevant
111In-RICs for 48 h. After RIC removal, cells were washed with PBS three times and fixed with 4% (
v/v) paraformaldehyde in PBS at RT for 15 min, permeabilised with Triton X-100 (0.5% in PBS) at RT for 15 min, and blocked with PBS/BSA blocking solution (10 mg/mL) at 37 °C for 1 h. Next, cells were incubated with an anti-phosphorylated histone H2AX (γH2AX) antibody diluted in PBS (1:200; 05-636, Sigma Aldrich, Saint-Quentin Fallavier, France) at 37 °C for 1 h, followed by washing and incubation with a goat anti-mouse IgG (H+L) FITC-conjugated antibody diluted in PBS/BSA (1:400; 12-506, Sigma-Aldrich, Saint-Quentin Fallavier, France) at 37 °C for 1 h. Finally, cells were washed with PBS/Tween-20 (0.1%) and then with PBS (three times/each). Coverslips were mounted on slides with Vectashield antifade mounting medium and DAPI (Vector laboratories, Les Ulis, France) and imaged using a ×63 or ×40 objective on a Leica fluorescence inverted microscope (Leica Microsystems, Wetzlar, Germany), as described previously [
15].
2.9. Radioactivity Uptake and Cell Fractionation
To isolate nuclei, the chaotropic method described by Pouget et al. [
34] was used. Briefly, 1 × 10
6 cells grown in T25 flasks overnight were incubated with 0.1 MBq/mL (3 mL)
111In-RICs for 48 h. At various time-points (1, 2, 3, and 6 days), cells were trypsinised and rinsed with PBS three times to remove unbound RICs. The activity of isolated whole cells was determined with a γ-ray counter (Hidex Automatic gamma counter, Villebon-sur-Yvette, France). Next, cells were resuspended in 1 mL lysis buffer (320 mM sucrose, 5 mM MgCl
2, 10 mM Tris-HCl, 0.1 mM deferoxamine, and 1% Triton X-100 in H
2O) and centrifuged at 1500×
g for 10 min. The supernatant (cytoplasm + membrane) was collected. The pellet was again resuspended in 1 mL lysis buffer, centrifuged at 1500×
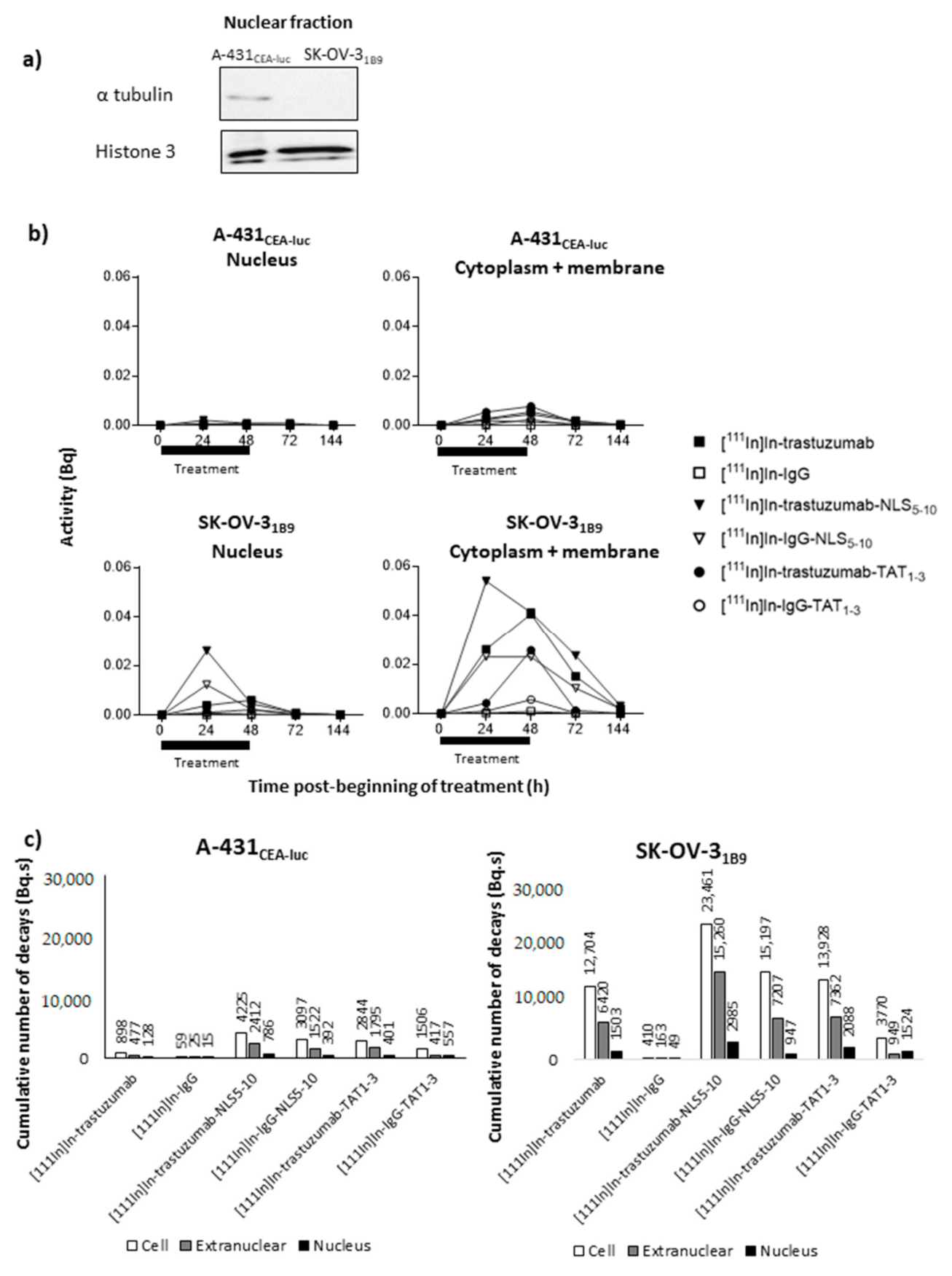
g for 10 min, and the supernatant was collected. The pellet (cell nuclei) was rinsed with 1 mL PBS three times to remove all traces of cytoplasm or membrane. Finally, the activity present in both fractions (nuclei and cytoplasm + membrane) was measured using a γ-ray counter. The cumulative number of decays (Bq.s) per cell over 6 days was then calculated as well as the cumulative number of decays (Bq.s) per fraction (nucleus and cytoplasm + membrane).
2.10. Evaluation of Nuclear Fraction Contamination by Western Blotting
Nuclei were rinsed twice with PBS and lysed in RIPA buffer (Santa Cruz; Santa Cruz, CA, USA) at 4 °C for 30 min. The total protein concentration in the nuclear fraction was determined using the BCA Protein Assay Reagent (BCA Assay Kit, Thermo Fisher scientific, Illkirch, France). Then, 6 µg of nuclear proteins were separated by SDS-PAGE (12% poly-acrylamide gel) and electrotransferred onto nitrocellulose membranes (Bio-Rad, Grabels, France). Membranes were incubated with anti-α-tubulin (11H10) (1/1000; Cell Signaling Technology, Leiden, The Netherlands) and anti-phosphorylated histone 3 (Ser10) antibodies (1/1000; Cell Signaling Technology, Leiden, The Netherlands) diluted in Tris-buffered saline/BSA (5%). A horseradish peroxidase-conjugated anti-rabbit IgG (Cell Signaling Technology, Danvers, MA, USA) was used as a secondary antibody. Proteins were revealed using an enhanced chemiluminescence system according to the manufacturer’s instructions (Clarity, Bio-Rad, Marnes-La-Coquette, France). Protein expression was imaged with a PXi analyser (Ozyme, St Quentin en Yvelines, France).
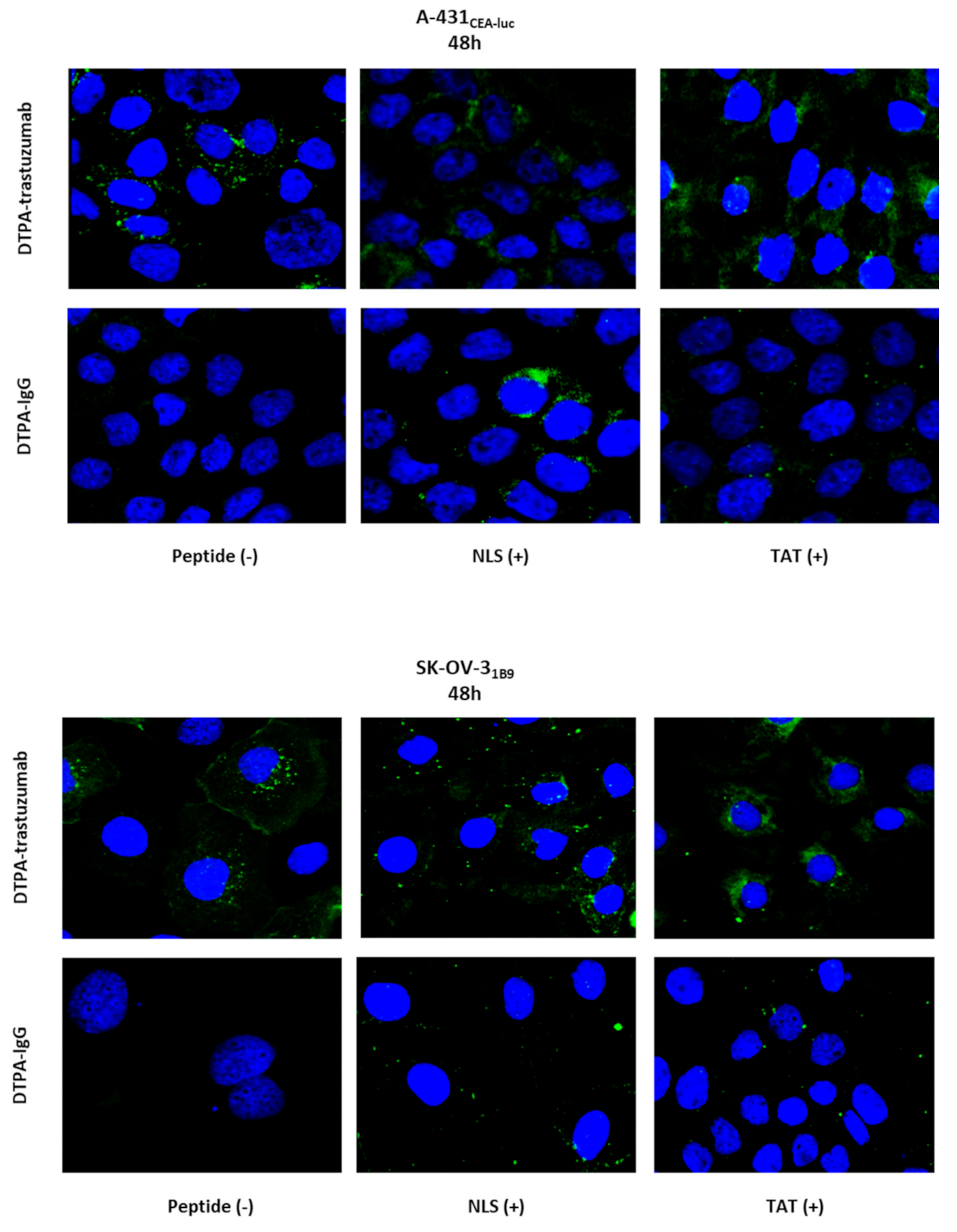
2.11. Immunofluorescence Analysis of the Immunoconjugate Subcellular Localisation
As described in
Section 2.8, cells were plated on coverslips in 6-well plates, and, the day after, they were incubated with different unlabelled immunoconjugates for 48 h. Then, cells were fixed and permeabilised (see
Section 2.8). After blocking with PBS/BSA, an AlexaFluor-488-conjugated anti-human secondary antibody (1:200; (F(ab′)
2-goat anti-human IgG (H+L) Cross-Adsorbed, Thermo Fisher Scientific) was added at 37 °C in the dark for 1 h. Coverslips were then mounted with Vectashield with DAPI (see
Section 2.8) and imaged using a ×63 or ×40 objective and a Leica fluorescence inverted microscope.
2.12. Energy Deposition Calculations
The energy deposition of Auger electrons, CEs, gamma- and X-rays in spheres was evaluated using the PENELOPE code system [
35] that simulates the interactions of radiations as they pass through the medium, in this case water. The detailed energy spectra of the radiations were generated with the BrIccEmis [
36,
37] code and typically using 5 million decay events. All simulations were done using the Monte Carlo approach.
2.13. Statistical Analysis
Data were analysed using GraphPad Prism version 8.4.3 (686). Data were described using means and standard deviation (SD). For gamma-H2AX, data were described using means and the standard error of the mean (SEM). Data were compared using the Student’s
t test. A
p value < 0.05 was considered significant. The Bliss independence model was used to determine the percentage of specific irradiation. In this method, the global efficacy of [
111In]In-trastuzumab was dissociated in specific and non-specific efficacy using the following formula, wherein the non-specific efficacy was determined as [
111In]In-IgG efficacy:
4. Discussion
This study assessed the ability of two highly cationic peptides, the synthetic 13-mer NLS peptide [CGYGPKKKRKVGG] from the simian virus 40 large-T antigen and the synthetic 16-mer TAT peptide [GRKKRRQRRRPPQGYG] from the HIV-1 TAT protein, to drive the
111In-anti-HER2 (trastuzumab) RIC into the cell nucleus of two HER2-expressing cancer cell lines, A-431
CEA-luc (HER2
+) and SK-OV-3
1B9 (HER2
++). According to the literature, after HER2-mediated internalisation in the cytoplasm, NLS and TAT peptide sequences are recognised by importin-α and importin-β1 to form a nuclear-pore-targeting complex [
38,
39]. This complex facilitates RIC passage through the nuclear pore and their translocation into the nucleus where they will be dissociated to release NLS-/TAT-conjugates [
11]. In total, eight RICs were obtained and characterised: [
111In]In-trastuzumab, [
111In]In-trastuzumab-NLS
5-10, [
111In]In-trastuzumab-NLS
17-22, [
111In]In-trastuzumab-TAT
1-3 as a specific anti-HER2 RIC, and [
111In]In-IgG, [
111In]In-IgG-NLS
5-10, [
111In]In-IgG-NLS
17-22, and [
111In]In-IgG-TAT
1-3 as non-specific RICs.
First, functionalisation of trastuzumab with the TAT or NLS peptides slightly increased its cell binding, but this binding was at least predominantly mediated by non-specific mechanisms. Moreover, non-radioactive immunoconjugates showed very low cytotoxicity, even when used at the highest concentration (10.8 µg/mL; equivalent to tested volumic activity of 4 MBq/mL for
111In-RIC) (
Supplementary Figure S1).
Then, comparison of the different RICs showed that [111In]In-IgG was significantly less cytotoxic than [111In]In-trastuzumab, but more than what was reported in previous studies. The origin of 111In non-specific cytotoxic effect in vitro could be explained by CE emission. It must be noted that, in several studies showing the absence of non-specific cytotoxicity, clonogenic assays were performed using a protocol different from the one followed in the present study. This previous protocol included two consecutive steps. First, cells were exposed to the radionuclide for several hours before centrifugation and washes. Then, cells were seeded at low concentration for clonogenic assay. It is likely that the first step might have led to the loss of the most damaged cells, and therefore, cell survival was only measured for the most viable cells that had not been eliminated by washing and that could adhere to the flask bottom. Moreover, the non-specific irradiation contribution could be different in vivo, particularly considering that, in the present in vitro experiments, cells were incubated with RICs at high activity for 48 h, a situation that might not reflect the in vivo pharmacokinetics of 111In-RICs.
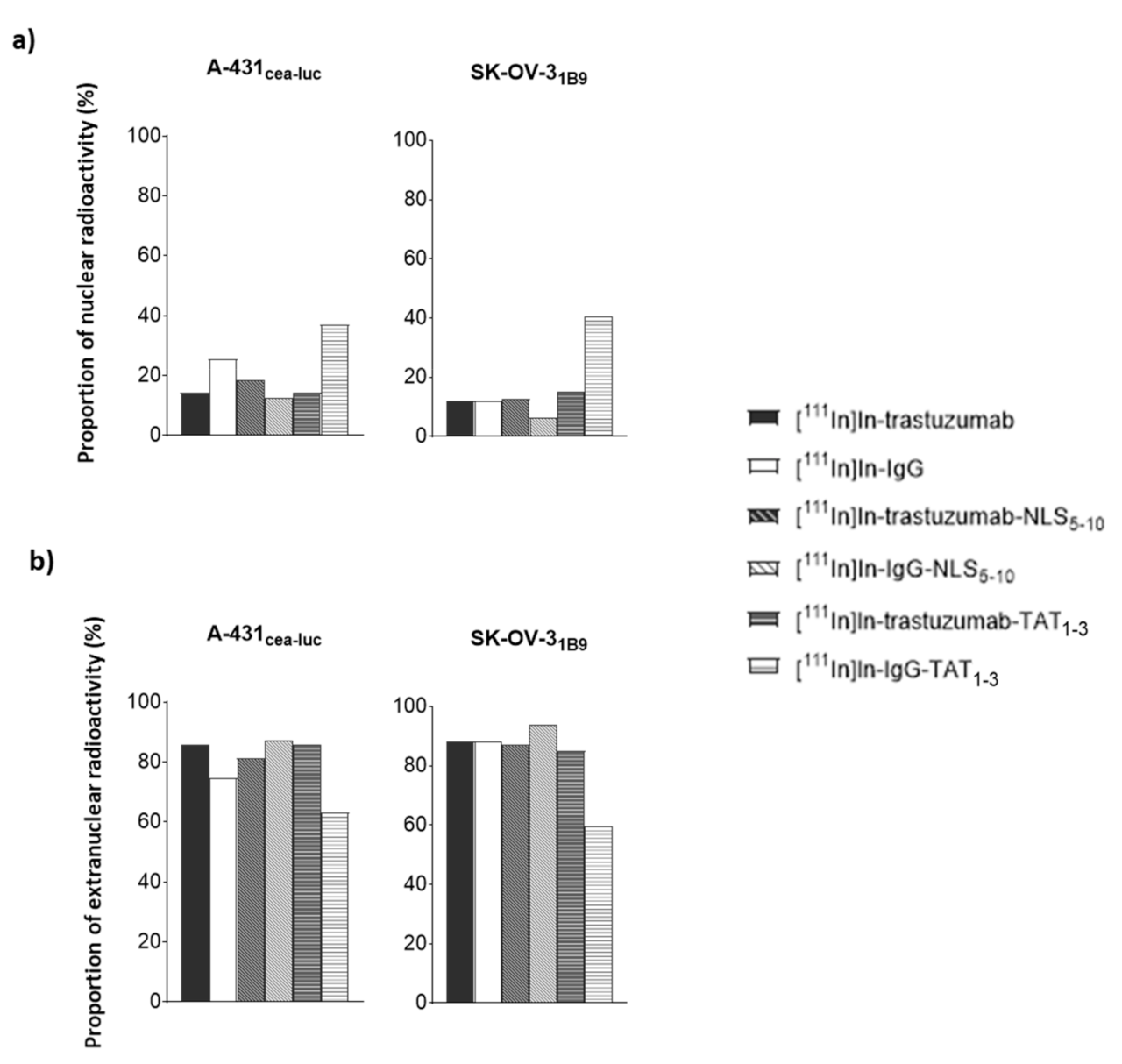
Functionalisation with TAT1-3 peptides did not significantly modify [111In]In-trastuzumab cytotoxicity in A-431CEA-luc cells and slightly increased it in SK-OV-31B9 cells (p < 0.01). The observation that the non-specific [111In]In-IgG-TAT1-3 was as cytotoxic as [111In]In-trastuzumab-TAT1-3indicates that TAT conjugation is associated with the loss of mAb specificity. The lack of or limited additional cytotoxicity provided by functionalisation with TAT1-3 could be explained by the low increase of activity localisation in the nucleus (Ãnuclear) after cell exposure to [111In]In-trastuzumab-TAT1-3. Ãcellular increase in A-431CEA-luc and in SK-OV-31B9 cells exposed to [111In]In-trastuzumab-TAT1-3 was associated with non-specificity because it also increased in cells incubated with [111In]In-IgG- TAT1-3. However, the Ãcellular increase in SK-OV-31B9 cells was lower than in A-431CEA-luc cells, suggesting that the higher HER2 expression level of this cell line prevented the passive internalisation of the TAT-RIC.
Functionalisation of radiolabelled mAbs with 17 to 22 NLS/mAb (NLS17-22) resulted in a drastic reduction of the surviving fractions for both [111In]In-trastuzumab-NLS17-22 and [111In]In-IgG-NLS17-22. Again the addition of NLS peptides was associated with a loss of specificity. Reducing the number of NLS to 5–10 (NLS5-10) increased RIC specificity and maintained high cytotoxicity. Nevertheless, even functionalisation with NLS5-10 was associated with non-specific cytotoxicity in A-431CEA-luc cells because [111In]In-IgG-NLS5-10 was more cytotoxic than [111In]In-trastuzumab (p = 0.001), but the difference between the specific and non-specific [111In]In-mAb-NLS5-10 effects was more pronounced than for [111In]In-mAb-TAT1-3. Cell fractionation experiments showed that functionalisation with NLS5-10 was accompanied by an increase of the Ãnuclear values for both [111In]In-trastuzumab-NLS5-10 and [111In]In-IgG-NLS5-10 in A-431CEA-luc cells and, to a lower extent, in SK-OV-31B9 cells. Nevertheless, the most striking observation was that the functionalisation of [111In]In-trastuzumab with NLS5-10 significantly increased Ãcellular in both A-431CEA-luc (× 4.7) and in SK-OV-31B9 (× 1.8) cells, with a similar trend also for [111In]In-IgG (×52.5 in A-431CEA-luc cells; ×37.2 in SK-OV-31B9 cells), highlighting again the lack of specificity of the NLS-mediated uptake. It has been proven that NLS or TAT peptides pass through plasma membranes in a non-receptor-dependent manner thanks to their highly cationic properties. In the context of our study, NLS or TAT peptides’ addition to mAbs will generate a highly cationic macromolecule that will passively penetrate cells or accumulate onto negatively charged cell-surface proteins. Our results showed an increase of cell uptake when the mAb is conjugated with NLS or TAT peptides suggesting a correlation with the addition of NLS or TAT.
Moreover, the Ã
nuclear versus Ã
extranuc ratio with [
111In]In-trastuzumab-NLS
5-10 was similar to that obtained with [
111In]In-trastuzumab, suggesting that the NLS peptide did not improve the nuclear translocation of
111In-RIC once internalised in the cytoplasm (
Figure 4).
These results were supported by the fluorescent detection of immunoconjugate foci, mainly in the cytoplasm, and of very few NLS- and TAT-functionalised immunoconjugate foci in the nucleus. This low nuclear localisation might be due to endosomal–lysosomal entrapment, leading to RIC hydrolysis before they can reach the nucleus and might constitutes a key limiting factor for bringing AEEs into the nucleus. Several groups have developed attractive methods to promote endosomal escape. For instance, in hetero-functional constructs, a target-specific antibody is conjugated to cholic acid for endosomal escape and an NLS peptide is added for nuclear targeting [
30,
40]. Moreover, modular nanotransporters consist of a target-specific module, a diphtheria toxin translocation domain as endosomolytic module, an NLS peptide for transport to the nucleus, and the
Escherichia coli haemoglobin-like protein as a carrier module [
28,
40] [
28,
41]. These approaches should be further investigated to bypass Auger electron-carrier endosomal entrapment.
At this stage it was not possible to thoroughly investigate the relationship between clonogenic survival and activity uptake in the different cell compartments. Such study would require a comprehensive dosimetric approach (out of the scope of this study) and an assessment of the possible contribution of bystander effects. Moreover, it is important to keep in mind that the cell fractionation experiments were done using activities of 0.1 MBq/mL, and therefore, the only relationship that could be investigated would be with the clonogenic survival data measured at 0.5 MBq/mL.
Interestingly, in a previous study [
15], we showed that, in HCT116 cells exposed to [
125I]I-UdR, Ã
nuclear was about 441 Bq.s and Ã
cellular was 455 Bq.s. Using the Bliss model and considering that cytotoxicity was only due to radioactivity in the nucleus, we calculated that 80% of cell killing was due to
125I located in the DNA [
15], while here it was 20% (at 0.5 MBq/mL, a test activity close to the one used for determining the à values and for which non-specific irradiation can be neglected) but with about 1.8 (786/441) more decays. This would mean that one decay of
125I is 7.2 times more efficient than one decay of
111In. If we consider that the energy deposited per decay and due to Auger electrons is about 8.7 keV (
125I) and 5.4 keV (
111In) in a 5 µm diameter sphere, then
125I deposits about 1.6 times more “high LET” energy than
111In. Moreover, a correction factor also needs to be introduced to counterbalance the localisation differences (nuclear but not bound to DNA versus nuclear and bound to DNA). As a relative biological effectiveness value of seven has been proposed when
125I is bound to DNA [
42,
43] and of four [
7] when unbound, then the factor that needs to be introduced here is 1.7. Additionally, HCT116 cells are about 1.8 times more radiosensitive than A-431 cells at a dose of 2 Gy [
44]. In conclusion,
111In incorporated in our RICs should be 8.8 times (2.1 × 1.6 × 1.7 × 1.8) less efficient than [
125I]I-UdR, a value that can be compared to the value of 7.2 found above.
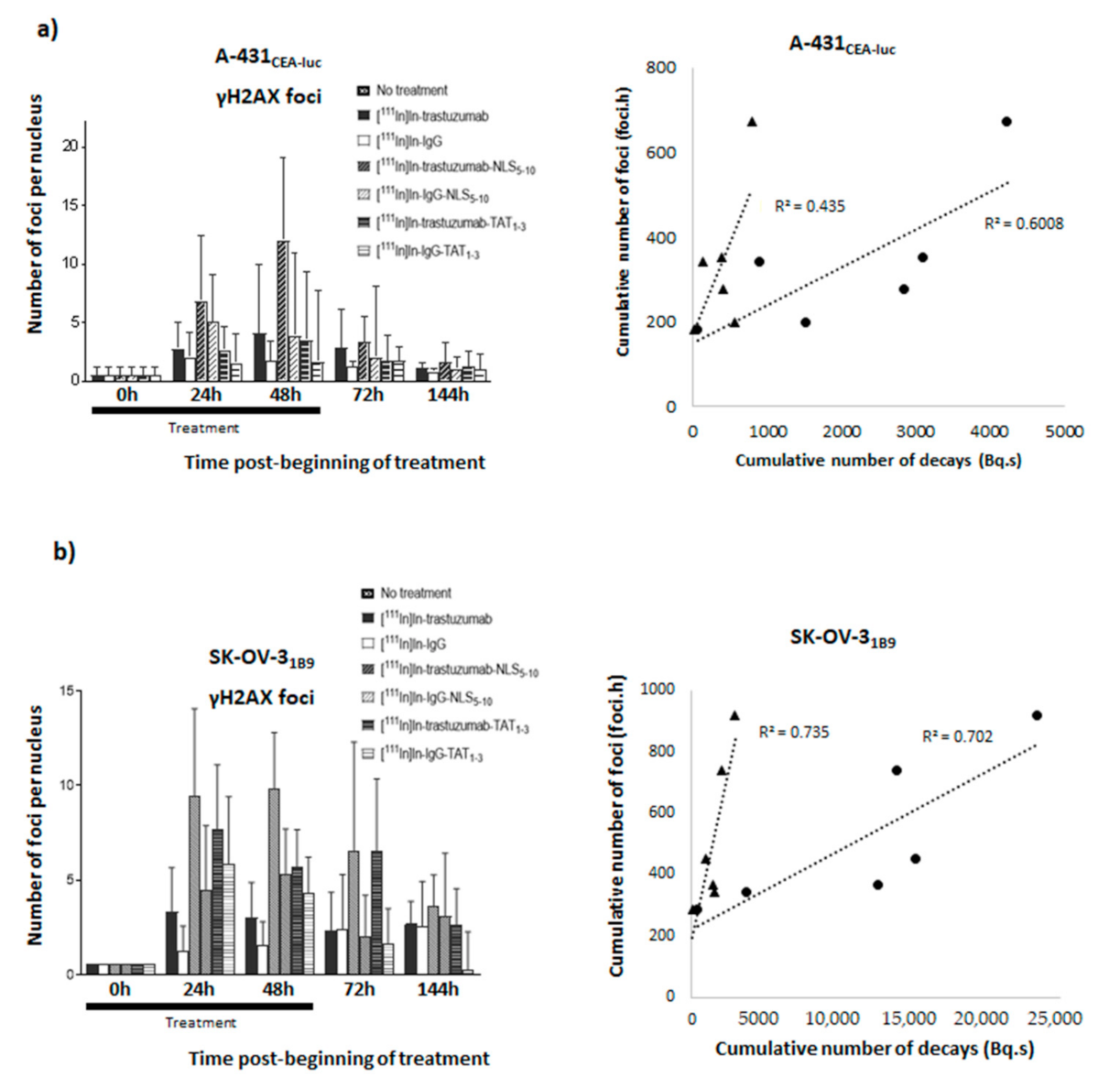
The preliminary analysis of DNA damage yield (γH2AX foci) in A-431
CEA-luc cells exposed to 4 MBq/mL RIC showed that the highest levels of γH2AX foci were observed with [
111In]In-trastuzumab-NLS
5-10 and [
111In]In-IgG-NLS
5-10, which are associated with the highest cytotoxicity and the highest Ã
nuclear and Ã
cellular values, although the transient DNA DSB yield cannot be strictly correlated with cell survival. As DNA breaks can be considered to be produced mostly by radiation, we investigated the relationship between the cumulative number of γH2AX foci measured over 144 h and the Ã
nuclear and Ã
cellular values. However, it must be kept in mind that DNA DSBs can also be produced by radioactivity contained in the culture medium or in neighbouring cells, two parameters not considered here. Moreover, the efficacy of DNA repair occurring at the same time as irradiation [
16] could be influenced by the nature of DNA DSBs induced by radioactivity localised in or outside the nucleus. Despite these approximations, we found a relationship between the cumulative number of DSBs and the Ã
nuclear and Ã
cellular values, although none of these parameters seemed to fit the data best. It must be also noted that some DNA DSBs can be produced by a delayed mechanism involving cell membrane irradiation. We showed previously that Auger electron-mediated irradiation of the cell membrane induces lipid raft formation and the subsequent activation of signalling pathways, leading to the formation of ROS via NF-kB and DNA damage [
15,
16,
17]. We could hypothesise that the high Ã
extranuc value of [
111In]In-trastuzumab-NLS
5-10 (2412 Bq.s) is related to HER2 binding at the cell membrane, which is not possible with [
111In]In-IgG. However, this hypothesis needs to be further assessed because we do not know whether enough energy is deposited at the cell membrane.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






