
Timing of Novel Drug 1A-116 to Circadian Rhythms Improves Therapeutic Effects against Glioblastoma


 , and
, and 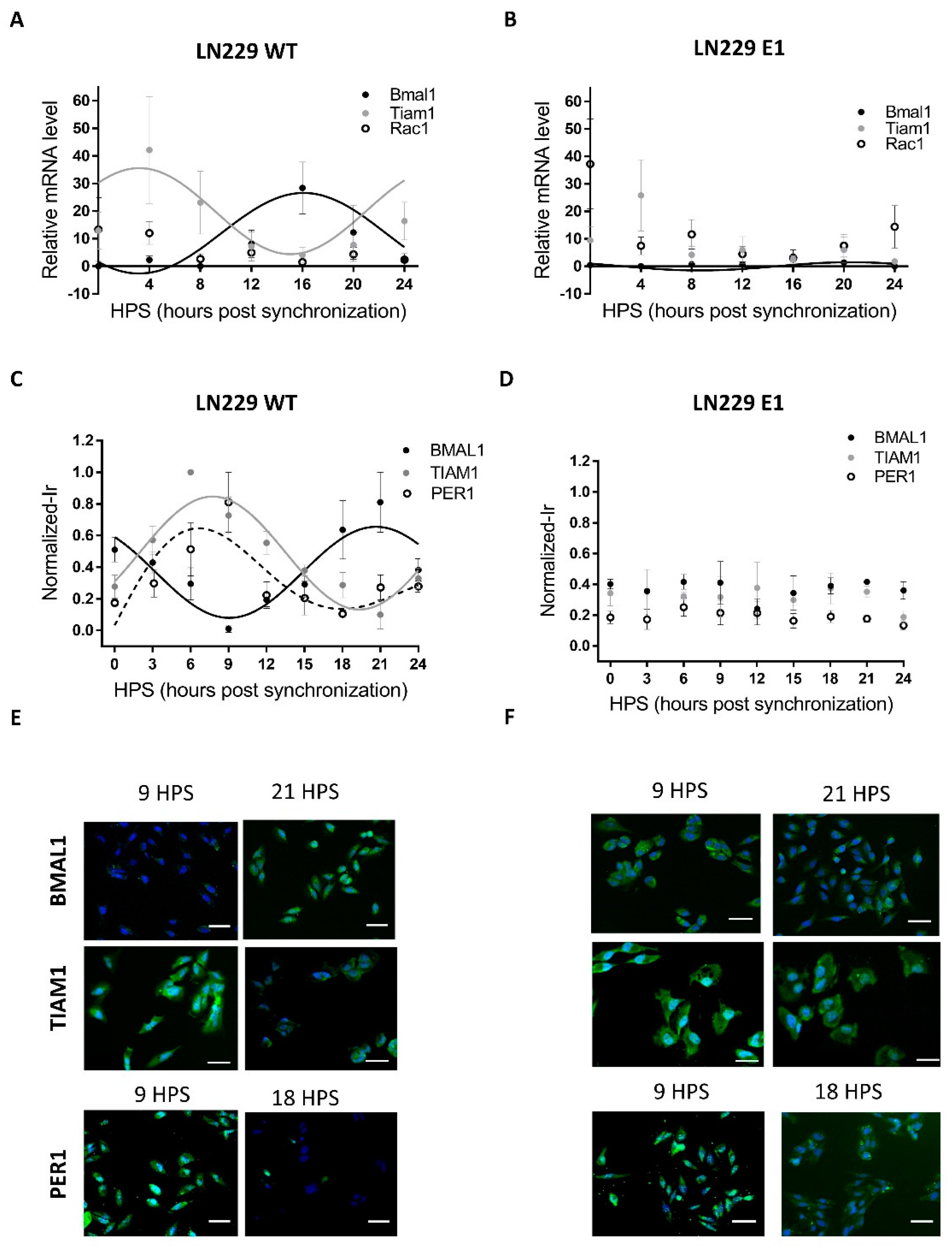{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Drugs and Reagents
2.3. CRISPR/cas9 Knock-Down of Bmal1 Expression
2.4. Bioluminescent Recordings
2.5. Cell Proliferation Assay
2.6. Cell Viability Assay
2.7. Cell Migration Assays
2.8. Early Apoptosis Assay
2.9. Quantitative Real-Time RT-PCRs
2.10. In-Cell Western Assays
2.11. Image Acquisition and Processing
2.12. Animals
2.13. Intracranial Surgery and Xenograft Implants
2.14. Chronopharmacological Drug Administration
2.15. Statistical Analysis
3. Results
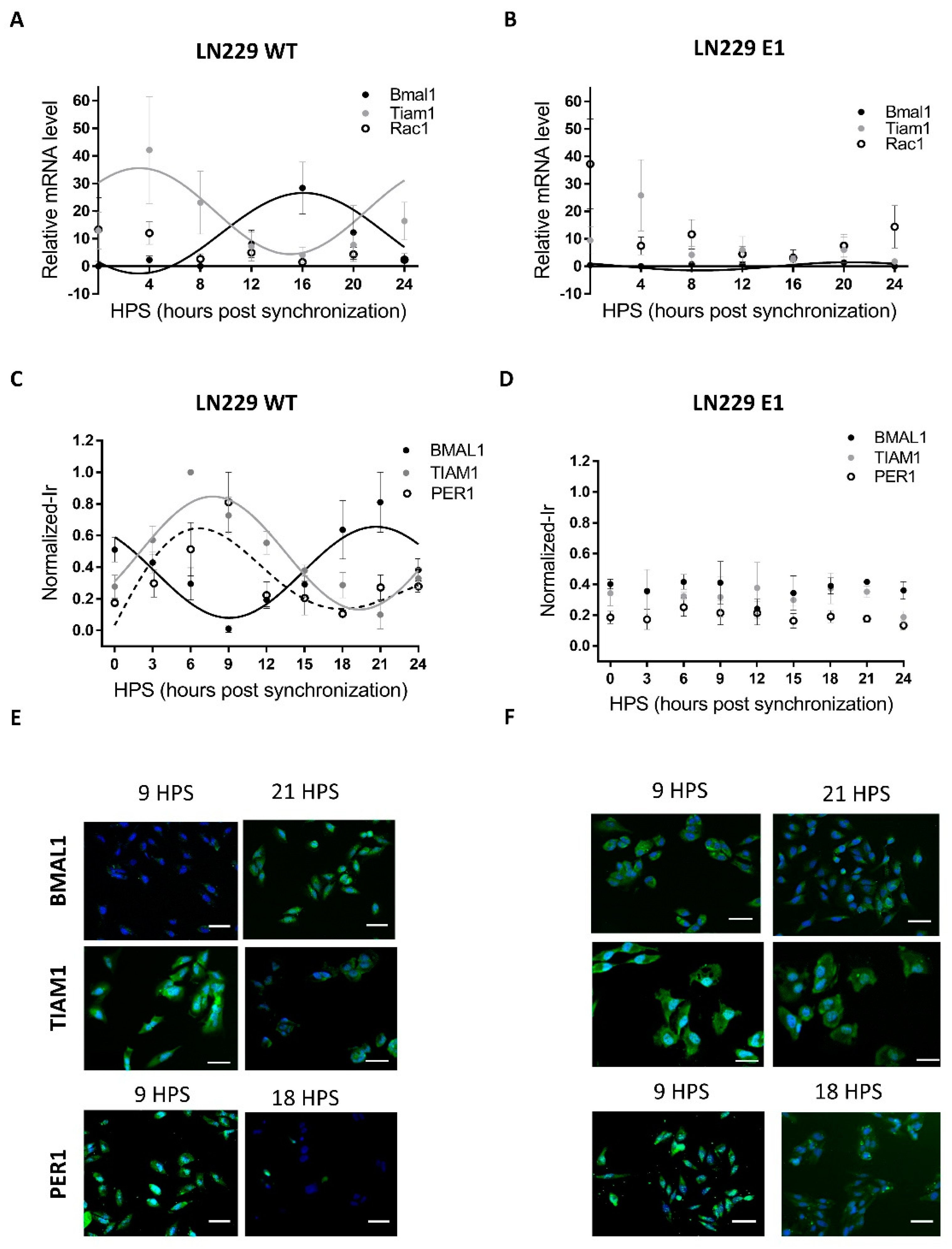
3.1. LN229 Cells Exhibit a Functional Circadian Clock That Modulates TIAM1 Expression
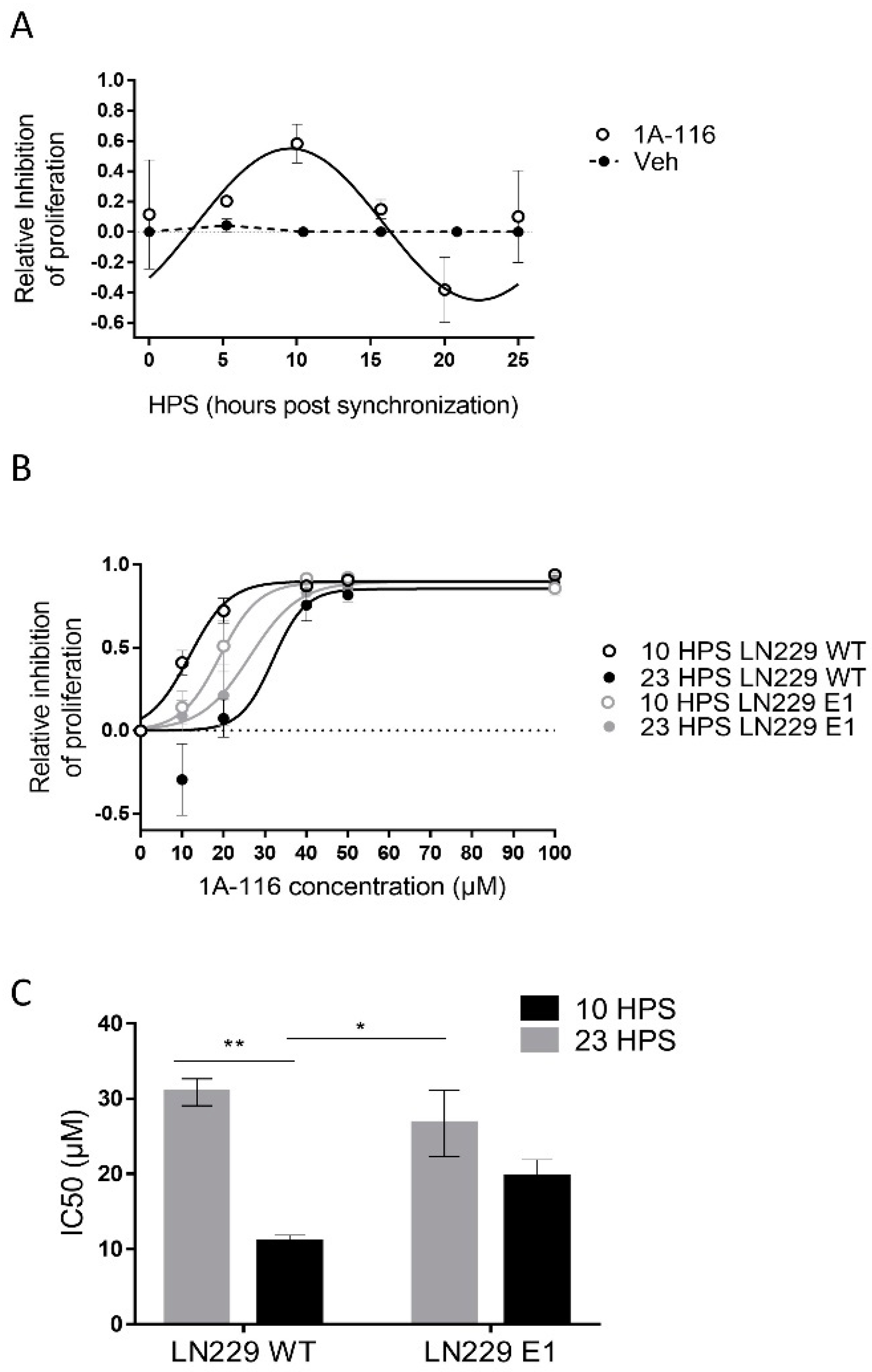
3.2. 1A-116 Inhibits GBM Cell Proliferation in a Circadian Manner
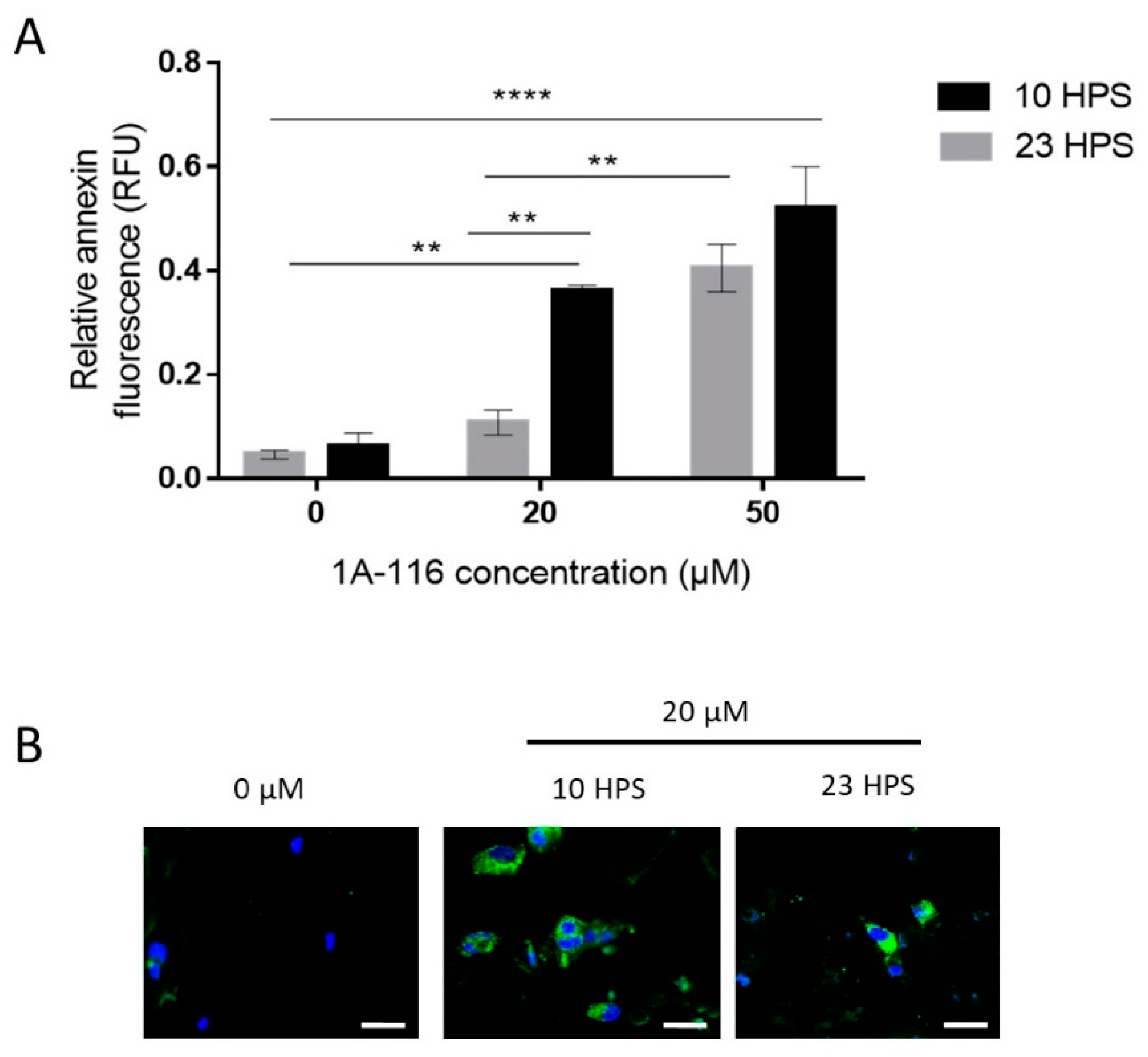
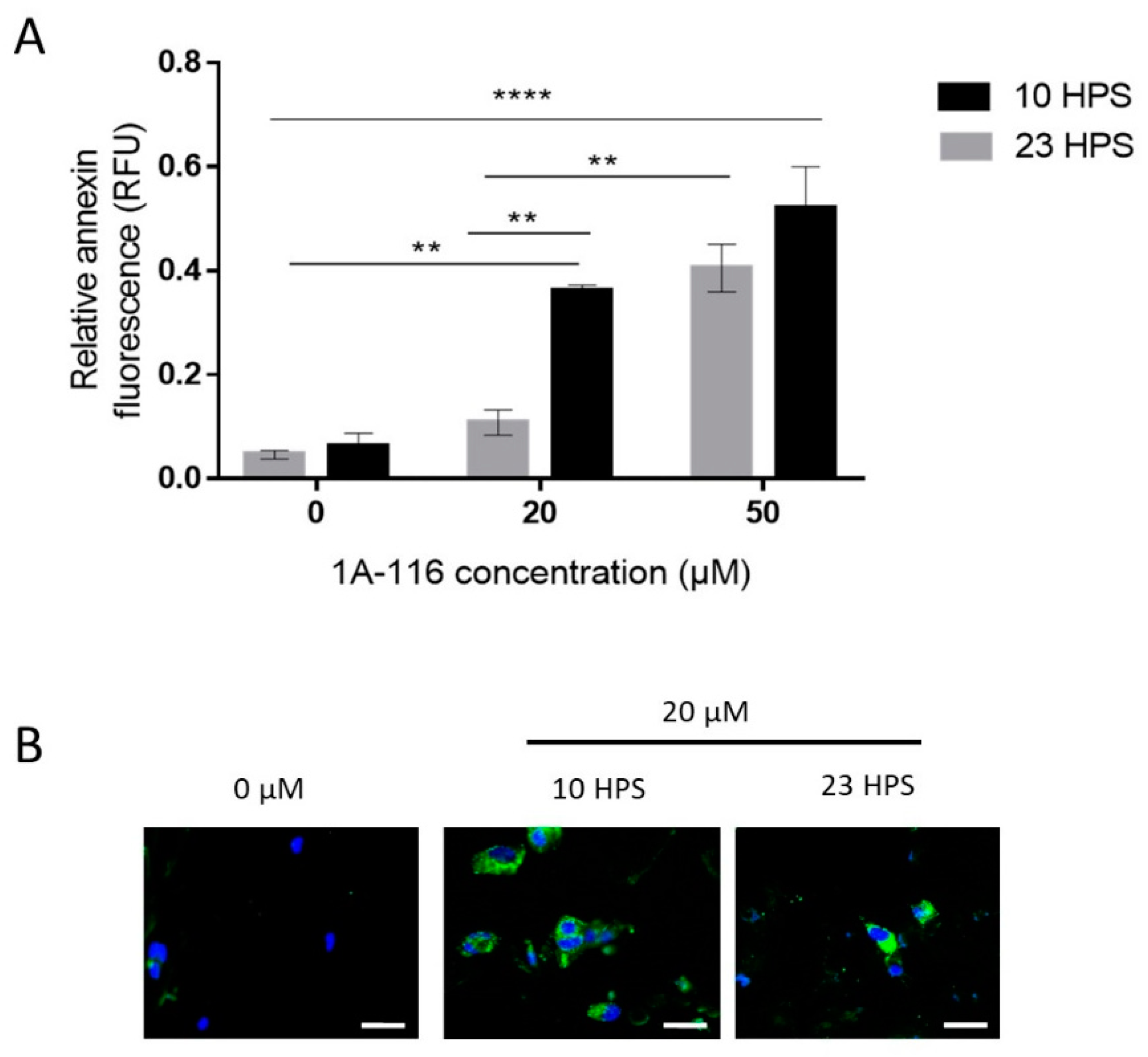
3.3. Pro-Apoptotic Effects of 1A-116 Are Circadian-Dependent
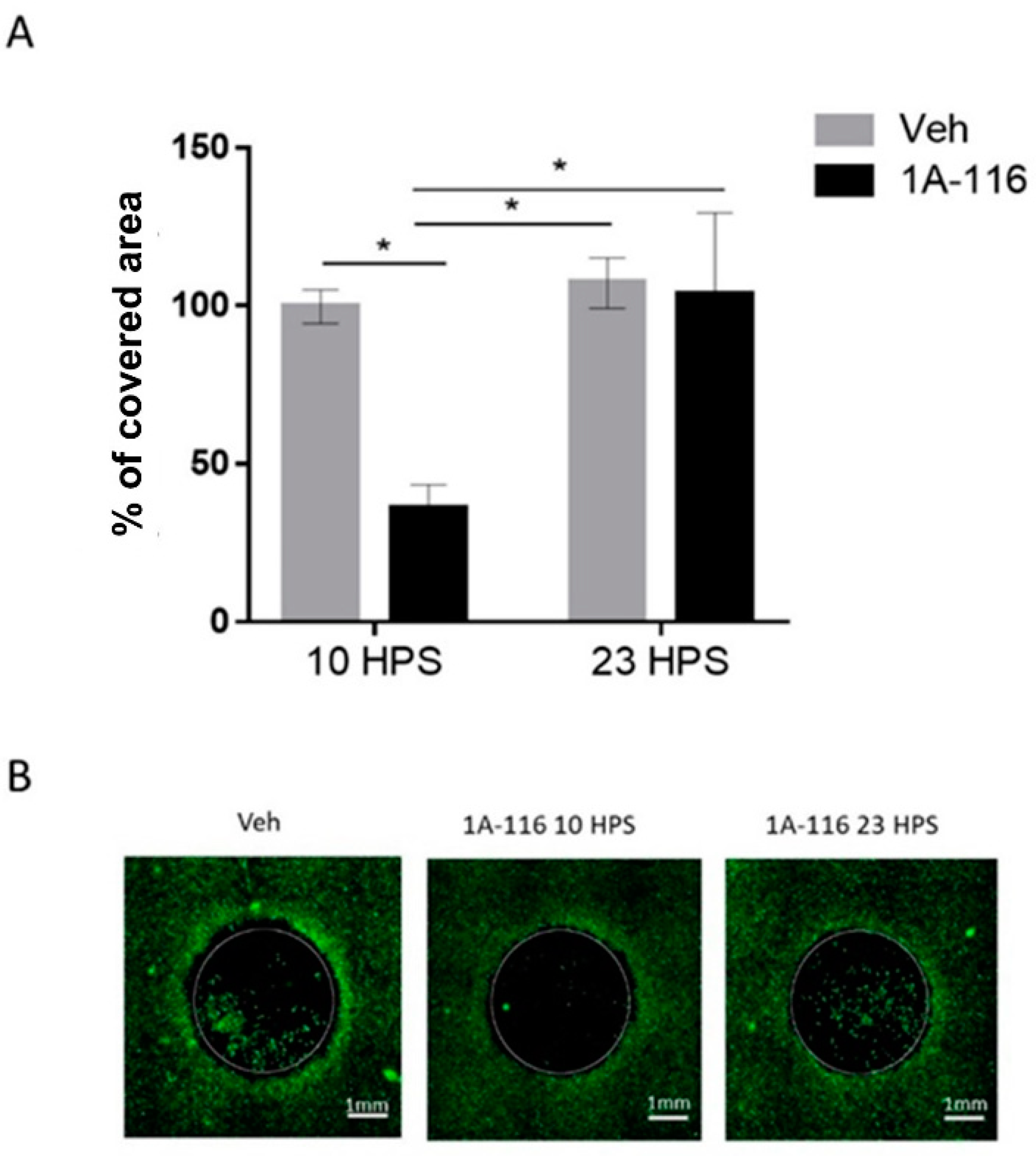
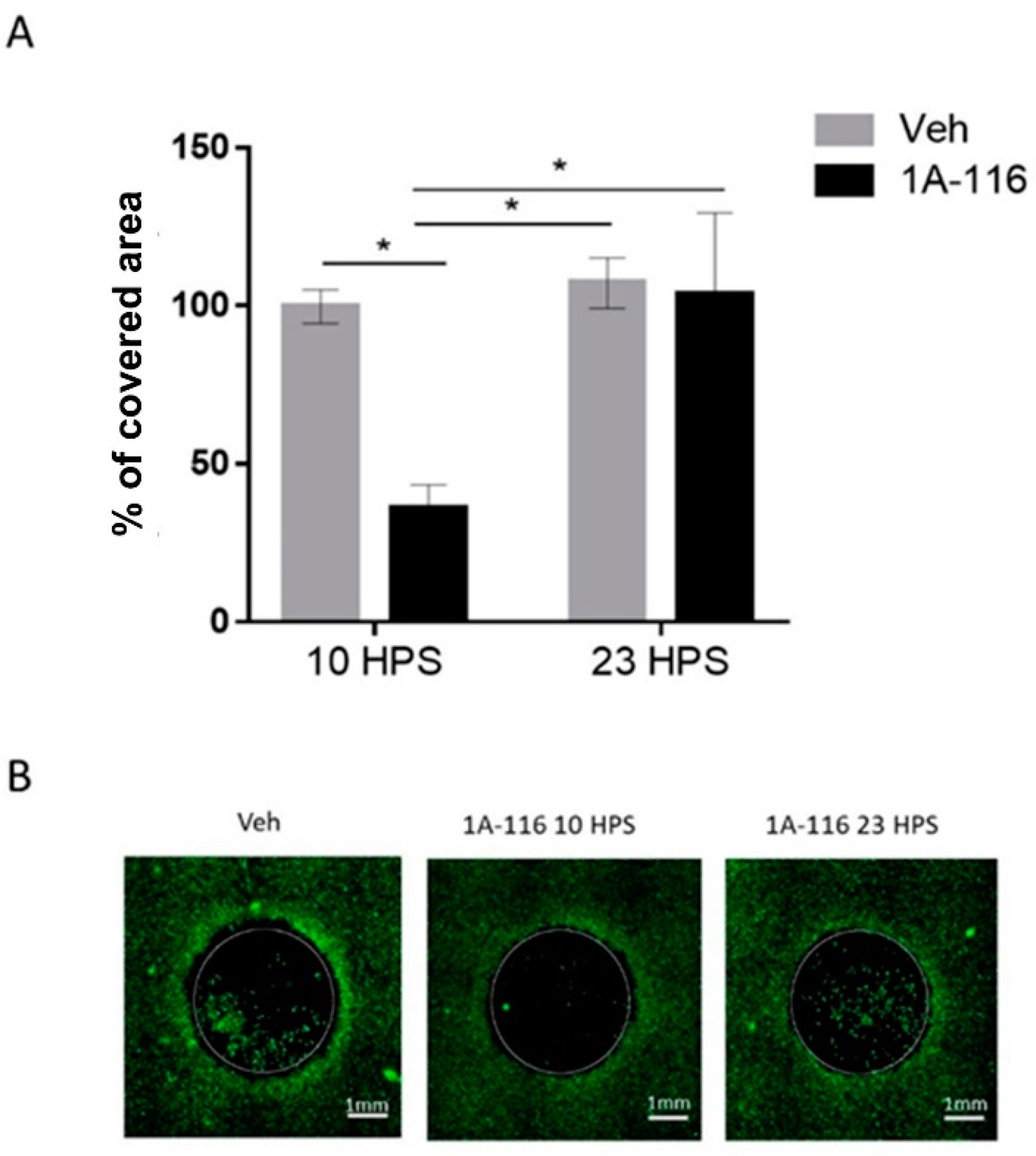
3.4. 1A-116 Inhibits LN229 Cell Migration Only at 10 HPS
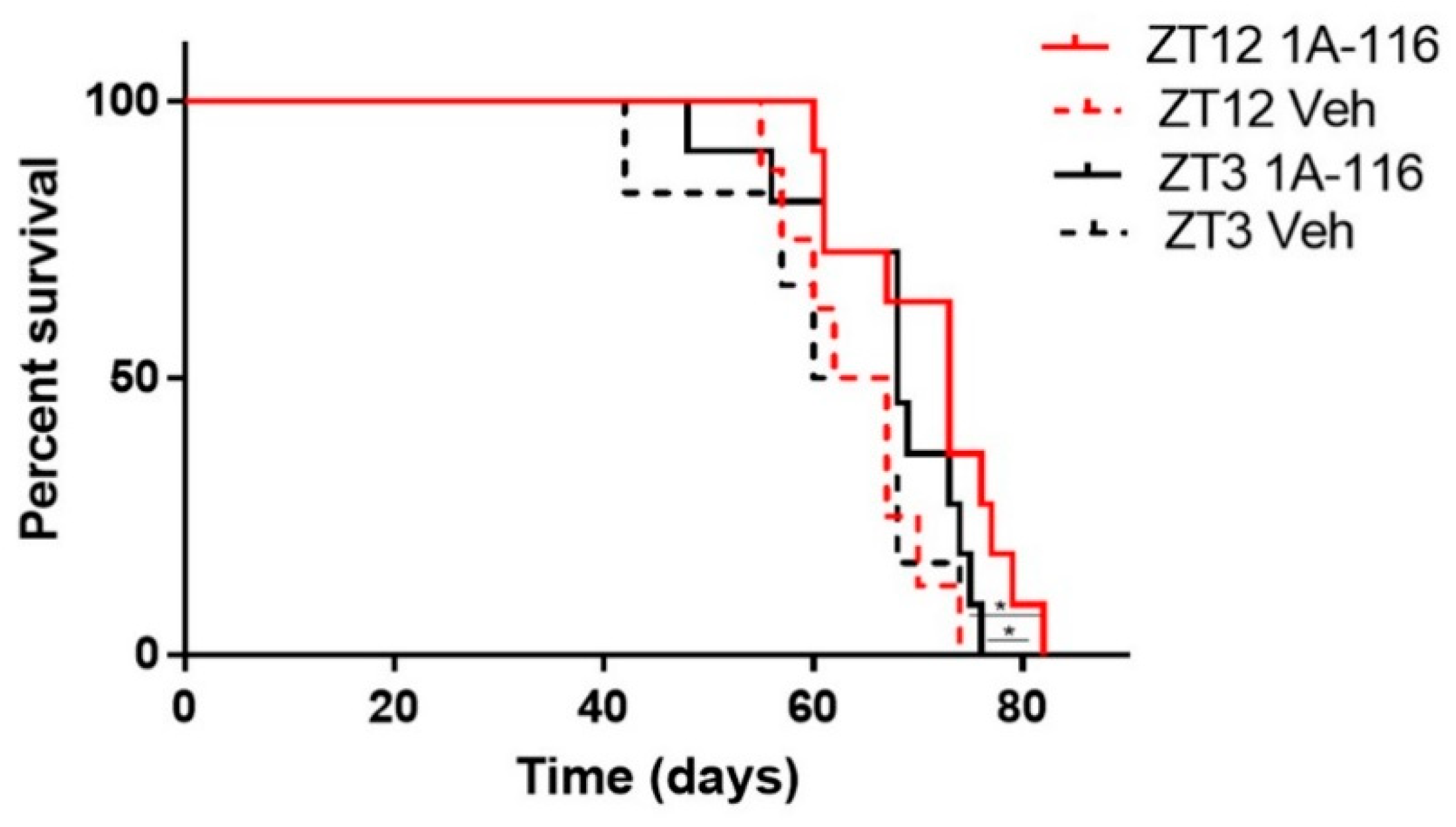
3.5. Daytime Dependent Effects of 1A-116 Treatment in Survival of GBM Nude Mice Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Golombek, D.A.; Rosenstein, R.E. Physiology of Circadian Entrainment. Physiol. Rev. 2010, 90, 1063–1102. [Google Scholar] [CrossRef] [Green Version]
- Gillette, M.U.; Medanic, M.; McArthur, A.J.; Liu, C.; Ding, J.M.; Faiman, L.E.; Weber, E.T.; Tcheng, T.K.; Gallman, E.A. Intrinsic Neuronal Rhythms in the Suprachiasmatic Nuclei and their Adjustment. Novartis Found. Symp. 2007, 183, 134–153. [Google Scholar] [CrossRef]
- Bjarnason, G.A. Chronobiology Implications for cancer chemotherapy. Acta Oncol. 1995, 34, 615–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahara, Y.; Shibata, S. Chronobiology and nutrition. Neuroscience 2013, 253, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Lee, C.C. The circadian clock: Pacemaker and tumour suppressor. Nat. Rev. Cancer 2003, 3, 350–361. [Google Scholar] [CrossRef] [PubMed]
- Omuro, A.; De Angelis, L.M. Glioblastoma and Other Malignant Gliomas: A Clinical Review. JAMA 2013, 310, 1842–1850. [Google Scholar] [CrossRef] [PubMed]
- Phillips, H.S.; Kharbanda, S.; Chen, R.; Forrest, W.F.; Soriano, R.H.; Wu, T.D.; Misra, A.; Nigro, J.M.; Colman, H.; Soroceanu, L.; et al. Molecular subclasses of high-grade glioma predict prognosis, delineate a pattern of disease progression, and resemble stages in neurogenesis. Cancer Cell 2006, 9, 157–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Watkins, S.; Sontheimer, H. Unique biology of gliomas: Challenges and opportunities. Trends Neurosci. 2012, 35, 546–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, A.L.; Colman, H. Glioma Biology and Molecular Markers. Cancer Treat. Res. 2015, 163, 15–30. [Google Scholar] [CrossRef]
- Davis, F.G.; McCarthy, B.J. Current epidemiological trends and surveillance issues in brain tumors. Expert Rev. Anticancer Ther. 2001, 1, 395–401. [Google Scholar] [CrossRef]
- Westphal, M.; Lamszus, K. The neurobiology of gliomas: From cell biology to the development of therapeutic approaches. Nat. Rev. Neurosci. 2011, 12, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Gomez, D.; Cardama, G.; Gonzalez, N.; Ciarlantini, M.; Comin, M.J.; Alonso, D.; Menna, P.L.; Donadio, L.G. Proapoptotic and antiinvasive activity of Rac1 small molecule inhibitors on malignant glioma cells. OncoTargets Ther. 2014, 7, 2021–2033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera, M.; Echeverria, E.; Lenicov, F.R.; Cardama, G.; Gonzalez, N.; Davio, C.; Fernández, N.; Menna, P.L. Pharmacological Rac1 inhibitors with selective apoptotic activity in human acute leukemic cell lines. Oncotarget 2017, 8, 98509–98523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parri, M.; Chiarugi, P. Rac and Rho GTPases in cancer cell motility control. Cell Commun. Signal. 2010, 8, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridley, A.J. Rho GTPase signalling in cell migration. Curr. Opin. Cell Biol. 2015, 36, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Scheidel, B.; Lemmer, B. Chronopharmacology of Oral Nitrates in Healthy Subjects. Chrono Int. 1991, 8, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Müller, F.O.; Van Dyk, M.; Hundt, H.K.L.; Joubert, A.L.; Luus, H.G.; Groenewoud, G.; Dunbar, G.C. Pharmacokinetics of temazepam after day-time and night-time oral administration. Eur. J. Clin. Pharmacol. 1987, 33, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Hollister, L.E. Chronopharmacology of amitriptyline. Clin. Pharmacol. Ther. 1983, 33, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Hermida, R.C.; Crespo, J.J.; Domínguez-Sardiña, M.; Otero, A.; Moyá, A.; Ríos, M.T.; Sineiro, E.; Castiñeira, M.C.; Callejas, P.A.; Pousa, L.; et al. Bedtime hypertension treatment improves cardiovascular risk reduction: The Hygia Chronotherapy Trial. Eur. Hear. J. 2020, 41, 4565–4576. [Google Scholar] [CrossRef]
- Shuboni-Mulligan, D.D.; Breton, G.; Smart, D.; Gilbert, M.; Armstrong, T.S. Radiation chronotherapy—clinical impact of treatment time-of-day: A systematic review. J. Neuro-Oncol. 2019, 145, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Lévi, F.; Karaboue, A.; Ettiene-Grimaldi, M.C.; Pantaud, G.; Focan, C.; Innominato, P.; Bouchahda, M.; Milano, G.; Chatelut, E. Pharmacokinetics of Irinotecan, Oxaliplatin and 5-fluorouracil during hepatic artery chronomodulated infusion: A translational European OPTILIV study. Clin. Pharmacok. 2017. [Google Scholar] [CrossRef] [PubMed]
- Dallmann, R.; Brown, S.A.; Gachon, F. Chronopharmacology: New Insights and Therapeutic Implications. Annu. Rev. Pharmacol. Toxicol. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballesta, A.; Innominato, P.F.; Dallmann, R.; Rand, D.A.; Lévi, F.A. Systems Chronotherapeutics. Pharmacol. Rev. 2017, 69, 161–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lévi, F.; Zidani, R.; Misset, J.-L. Randomised multicentre trial of chronotherapy with oxaliplatin, fluorouracil, and folinic acid in metastatic colorectal cancer. Lancet 1997, 350, 681–686. [Google Scholar] [CrossRef]
- Damato, A.R.; Luo, J.; Katumba, R.G.N.; Talcott, G.R.; Rubin, J.B.; Herzog, E.D.; Campian, J.L. Temozolomide chronotherapy in patients with glioblastoma: A retrospective single-institute study. Neuro-Oncol. Adv. 2021, 3, vdab041. [Google Scholar] [CrossRef] [PubMed]
- Balsalobre, A.; Damiola, F.; Schibler, U. A Serum Shock Induces Circadian Gene Expression in Mammalian Tissue Culture Cells. Cell 1998, 93, 929–937. [Google Scholar] [CrossRef] [Green Version]
- Wagner, P.M.; Alderete, L.G.S.; Gorné, L.D.; Gaveglio, V.; Salvador, G.; Pasquaré, S.; Guido, M.E. Proliferative Glioblastoma Cancer Cells Exhibit Persisting Temporal Control of Metabolism and Display Differential Temporal Drug Susceptibility in Chemotherapy. Mol. Neurobiol. 2019, 56, 1276–1292. [Google Scholar] [CrossRef] [Green Version]
- McInerney, M.P.; Pan, Y.; Short, J.L.; Nicolazzo, J.A. Development and Validation of an In-Cell Western for Quantifying P-Glycoprotein Expression in Human Brain Microvascular Endothelial (hCMEC/D3) Cells. J. Pharm. Sci. 2017, 106, 2614–2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, A.C.; Tran, H.G.; Zhang, E.; Priest, A.A.; Welsh, D.K.; Kay, S.A. Redundant Function of REV-ERBα and β and Non-Essential Role for Bmal1 Cycling in Transcriptional Regulation of Intracellular Circadian Rhythms. PLoS Genet. 2008, 4, e1000023. [Google Scholar] [CrossRef]
- Maier, B.; Lorenzen, S.; Finger, A.; Herzel, H.; Kramer, A. Searching novel clock genes using RNAi-based screening. In Circadian Clocks, Methods and Protocols; Brown, S.A., Ed.; Humana: New York, NY, USA, 2021. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Meth. 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Díez-Noguera, A. Methods for serial analysis of long time series in the study of biological rhythms. J. Circadian Rhythm. 2013, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Anafi, R.C.; Hughes, M.E.; Kornacker, K.; HogenEsch, J.B. MetaCycle: An integrated R package to evaluate periodicity in large scale data. Bioinformatics 2016, 32, 3351–3353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robbe-Sermesant, K.; Otto-Bruc, A.; Chardin, P.; Antonny, B.; Lee, E.B.; Skovronsky, D.M.; Abtahian, F.; Doms, R.W.; Lee, V.M.-Y. Dissociation of GDP Dissociation Inhibitor and Membrane Translocation Are Required for Efficient Activation of Rac by the Dbl Homology-Pleckstrin Homology Region of Tiam. J. Biol. Chem. 2003, 278, 4756–4762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotter, T.; Lennon, S.V.; Glynn, J.G.; Martin, S. Cell death via apoptosis and its relationship to growth, development and differentiation of both tumour and normal cells. Anticancer. Res. 1990, 10, 1153–1159. [Google Scholar]
- Andree, H.A.M.; Reutelingsperger, C.P.M.; Hauptmann, R.; Hemker, H.C.; Hermens, W.T.; Willems, G.M. Binding of vascular anticoagulant α (VACα) to planar phospholipid bilayers. J. Biol Chem. 1990, 265, 4923–4928. [Google Scholar] [CrossRef]
- Koopman, G.; Reutelingsperger, C.P.M.; Kuijten, G.A.M.; Keehnen, R.M.J.; Pals, S.T.; Van Oers, M.H.J. Annexin V for flow cytometric detection of phosphatidylserine expression on B cells undergoing apoptosis. Blood 1994, 84, 1415–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, D.R.; Omuro, A.M.P.; Ravelo, A.; Sommer, N.; Guerin, A.; Ionescu-Ittu, R.; Shi, S.; Macalalad, A.; Uhm, J.H. Overall survival in patients with glioblastoma before and after bevacizumab approval. Curr. Med. Res. Opin. 2017, 34, 813–820. [Google Scholar] [CrossRef]
- Benna, C.; Helfrich-Förster, C.; Rajendran, S.; Monticelli, H.; Pilati, P.; Nitti, D.; Mocellin, S. Genetic variation of clock genes and cancer risk: A field synopsis and meta-analysis. Oncotarget 2017, 8, 23978–23995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Relógio, A.; Thomas, P.; Medina-Pérez, P.; Reischl, S.; Bervoets, S.; Gloc, E.; Riemer, P.; Mang-Fatehi, S.; Maier, B.; Schäfer, R.; et al. Ras-Mediated Deregulation of the Circadian Clock in Cancer. PLoS Genet. 2014, 10, e1004338. [Google Scholar] [CrossRef] [PubMed]
- Madden, M.H.; Anic, G.M.; Thompson, R.C.; Nabors, L.B.; Olson, J.J.; Browning, J.E.; Monteiro, A.N.; Egan, K.M. Circadian pathway genes in relation to glioma risk and outcome. Cancer Causes Control 2013, 25, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Liu, Y.; Siddique, R.; Nabi, G.; Xue, M.; Hou, H. Impact of chronically alternating light-dark cycles on circadian clock mediated expression of cancer (glioma)-related genes in the brain. Int. J. Biol. Sci. 2019, 15, 1816–1834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varghese, R.T.; Young, S.; Pham, L.; Liang, Y.; Pridham, K.; Guo, S.; Murphy, S.; Kelly, D.F.; Sheng, Z. Casein Kinase 1 Epsilon Regulates Glioblastoma Cell Survival. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hainich, E.C.; Pizzio, G.A.; Golombek, D.A. Constitutive activation of the ERK-MAPK pathway in the suprachiasmatic nuclei inhibits circadian resetting. FEBS Lett. 2006, 580, 6665–6668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldsmith, C.S.; Kim, S.M.; Karunarathna, N.; Neuendorff, N.; Toussaint, L.G.; Earnest, D.J.; Bell-Pedersen, D. Inhibition of p38 MAPK activity leads to cell type-specific effects on the molecular circadian clock and time-dependent reduction of glioma cell invasiveness. BMC Cancer 2018, 18, 43. [Google Scholar] [CrossRef]
- Foldynova-Trantirkova, S.; Sekyrová, P.; Tmejová, K.; Brumovská, E.; Bernatík, O.; Blankenfeldt, W.; Krejčí, P.; Kozubík, A.; Doležal, T.; Trantírek, L.; et al. Breast cancer-specific mutations in CK1ε inhibit Wnt/β-catenin and activate the Wnt/Rac1/JNK and NFAT pathways to decrease cell adhesion and promote cell migration. Breast Cancer Res. 2010, 12, R30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruben, M.D.; Smith, D.F.; FitzGerald, G.A.; Hogenesch, J.B. Dosing time matters. Science 2019, 365, 547–549. [Google Scholar] [CrossRef]
- Focan, C. Chronobiology and biochemical markers of human cancer. Pathol. Biol. 1987, 35, 951–959. [Google Scholar] [PubMed]
- Slat, E.A.; Sponagel, J.; Marpegan, L.; Simon, T.; Kfoury, N.; Kim, A.; Binz, A.; Herzog, E.D.; Rubin, J.B. Cell-intrinsic, Bmal1-dependent Circadian Regulation of Temozolomide Sensitivity in Glioblastoma. J. Biol. Rhythm. 2017, 32, 121–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pevet, P.; Challet, E.; Felder-Schmittbuhl, M.-P. Melatonin and the circadian system: Keys for health with a focus on sleep. Handb. Clin. Neurol. 2021, 179, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Fong, S.Y.; Shon, J.; Zhang, S.L.; Brooks, R.; Lahens, N.F.; Chen, D.; Van Dang, C.; Field, J.M.; Sehgal, A. Time-of-day specificity of anticancer drugs may be mediated by circadian regulation of the cell cycle. Sci. Adv. 2021, 7, eabd2645. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, A.; Hayer, K.; Lahens, N.F.; HogenEsch, J.B. CircaDB: A database of mammalian circadian gene expression profiles. Nucleic Acids Res. 2012, 41, D1009–D1013. [Google Scholar] [CrossRef]
- Matsuo, T.; Yamaguchi, S.; Mitsui, S.; Emi, A.; Shimoda, F.; Okamura, H. Control mechanism of the circadian clock for timing of cell division in vivo. Science 2003, 302, 255–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farshadi, E.; Yan, J.; Leclere, P.; Goldbeter, A.; Chaves, I.; Van Der Horst, G.T.J. The positive circadian regulators CLOCK and BMAL1 control G2/M cell cycle transition through Cyclin B1. Cell Cycle 2019, 18, 16–33. [Google Scholar] [CrossRef] [PubMed]
- Gréchez-Cassiau, A.; Rayet, B.; Guillaumond, F.; Teboul, M.; Delaunay, F. The circadian clock component BMAL1 is a critical regulator of p21WAF1/CIP1 expression and hepatocyte proliferation. J. Biol. Chem. 2008, 283, 4535–4542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjarnason, G.A.; Jordan, R.C.; Wood, P.A.; Li, Q.; Lincoln, D.W.; Sothern, R.B.; Hrushesky, W.J.; Ben-David, Y. Circadian Expression of Clock Genes in Human Oral Mucosa and Skin: Association with Specific Cell-Cycle Phases. Am. J. Pathol. 2001, 158, 1793–1801. [Google Scholar] [CrossRef]
- Farshadi, E.; van der Horst, G.; Chaves, I. Molecular Links between the Circadian Clock and the Cell Cycle. J. Mol. Biol. 2020, 432, 3515–3524. [Google Scholar] [CrossRef]
- Kiessling, S.; Beaulieu-Laroche, L.; Blum, I.D.; Landgraf, D.; Welsh, D.K.; Storch, K.-F.; Labrecque, N.; Cermakian, N. Enhancing circadian clock function in cancer cells inhibits tumor growth. BMC Biol. 2017, 15, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korkmaz, T.; Aygenli, F.; Emisoglu, H.; Ozcelik, G.; Canturk, A.; Yilmaz, S.; Ozturk, N. Opposite Carcinogenic Effects of Circadian Clock Gene BMAL1. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kobuchi, S.; Yazaki, Y.; Ito, Y.; Sakaeda, T. Circadian variations in the pharmacokinetics of capecitabine and its metabolites in rats. Eur. J. Pharm. Sci. 2018, 112, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Sallam, H.; El-Serafi, A.T.; Filipski, E.; Terelius, Y.; Levi, F.; Hassan, M. The effect of circadian rhythm on pharmacokinetics and metabolism of the Cdk inhibitor, roscovitine, in tumor mice model. Chrono Int. 2015, 32, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Lahens, N.F.; Ballance, H.I.; Hughes, M.E.; HogenEsch, J.B. A circadian gene expression atlas in mammals: Implications for biology and medicine. Proc. Natl. Acad. Sci. USA 2014, 111, 16219–16224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermida, R.C.; Mojón, A.; Hermida-Ayala, R.G.; Smolensky, M.H.; Fernández, J.R. Extent of asleep blood pressure reduction by hypertension medications is ingestion-time dependent: Systematic review and meta-analysis of published human trials. Sleep Med. Rev. 2021, 59, 101454. [Google Scholar] [CrossRef] [PubMed]
- Muller, P.Y.; Milton, M.N. The determination and interpretation of the therapeutic index in drug development. Nat. Rev. Drug Discov. 2012, 11, 751–761. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trebucq, L.L.; Cardama, G.A.; Lorenzano Menna, P.; Golombek, D.A.; Chiesa, J.J.; Marpegan, L. Timing of Novel Drug 1A-116 to Circadian Rhythms Improves Therapeutic Effects against Glioblastoma. Pharmaceutics 2021, 13, 1091. https://doi.org/10.3390/pharmaceutics13071091
Trebucq LL, Cardama GA, Lorenzano Menna P, Golombek DA, Chiesa JJ, Marpegan L. Timing of Novel Drug 1A-116 to Circadian Rhythms Improves Therapeutic Effects against Glioblastoma. Pharmaceutics. 2021; 13(7):1091. https://doi.org/10.3390/pharmaceutics13071091
Chicago/Turabian StyleTrebucq, Laura Lucía, Georgina Alexandra Cardama, Pablo Lorenzano Menna, Diego Andrés Golombek, Juan José Chiesa, and Luciano Marpegan. 2021. "Timing of Novel Drug 1A-116 to Circadian Rhythms Improves Therapeutic Effects against Glioblastoma" Pharmaceutics 13, no. 7: 1091. https://doi.org/10.3390/pharmaceutics13071091
APA StyleTrebucq, L. L., Cardama, G. A., Lorenzano Menna, P., Golombek, D. A., Chiesa, J. J., & Marpegan, L. (2021). Timing of Novel Drug 1A-116 to Circadian Rhythms Improves Therapeutic Effects against Glioblastoma. Pharmaceutics, 13(7), 1091. https://doi.org/10.3390/pharmaceutics13071091