
Modulation of Hair Growth Promoting Effect by Natural Products


Abstract
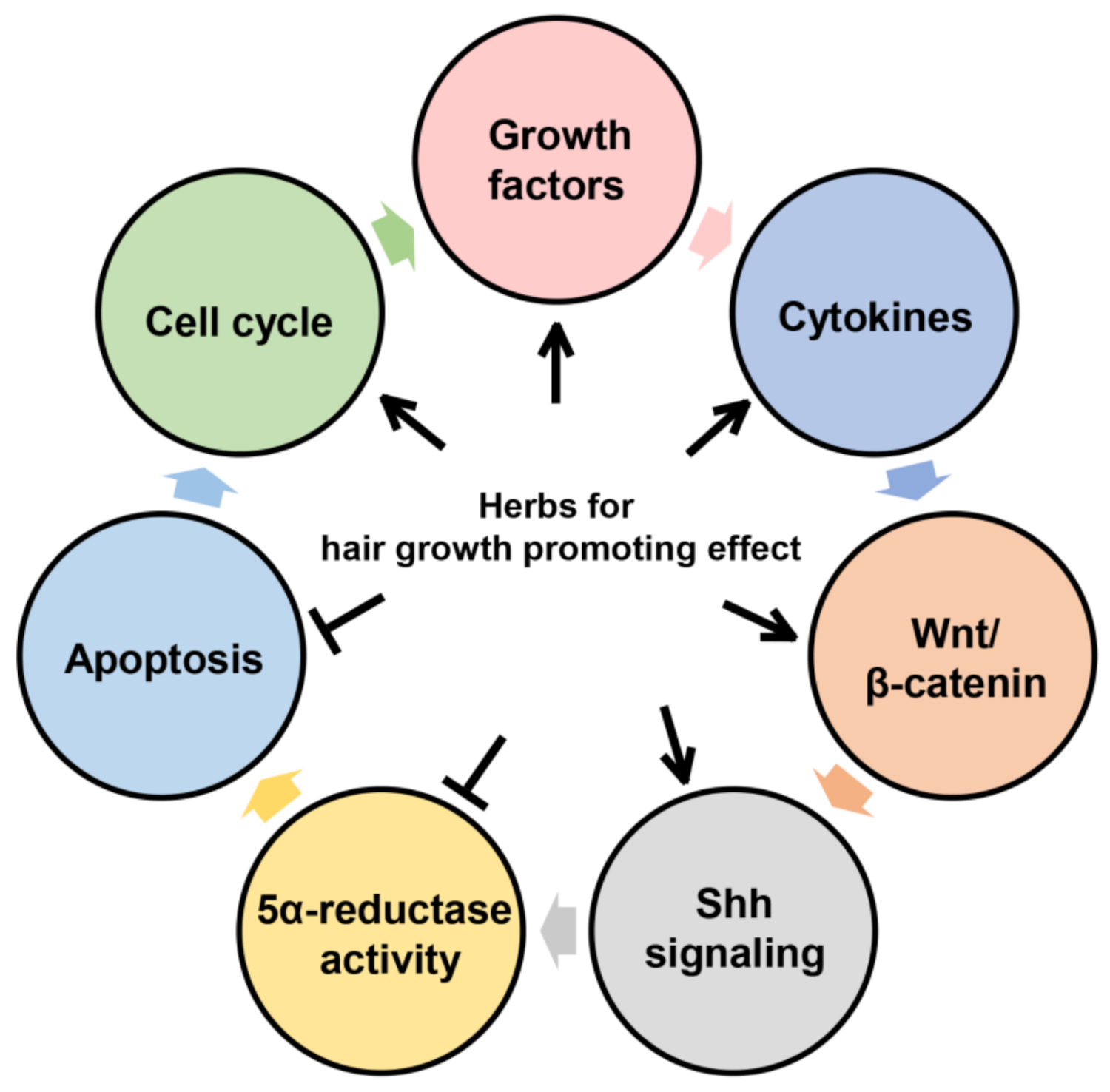
1. Introduction
2. Biochemical Action of Herbs and Their Extracts
2.1. Growth Factors
2.2. Cytokines
2.3. Wnt/β-Catenin
2.4. 5α-Reductase Inhibitory Effect
2.5. Sonic Hedgehog (Shh) Signaling
2.6. Apoptosis
2.7. Cell Cycle
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, X.M.; Li, Z.X.; Zhang, D.Y.; Yang, Y.C.; Fu, S.A.; Zhang, Z.Q.; Yang, R.H.; Xiong, K. A systematic summary of survival and death signalling during the life of hair follicle stem cells. Stem Cell Res. Ther. 2021, 12, 453. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.R.; Schmidt-Ullrich, R.; Paus, R. The hair follicle as a dynamic miniorgan. Curr. Biol. 2009, 19, R132–R142. [Google Scholar] [CrossRef]
- Blume-Peytavi, U.; Whiting, D.A.; Trüeb, R.M. Hair Growth and Disorders; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Wells, P.A.; Willmoth, T.; Russell, R.J. Does fortune favour the bald? Psychological correlates of hair loss in males. Br. J. Psychol. 1995, 86, 337–344. [Google Scholar] [CrossRef]
- Paus, R.; Cotsarelis, G. The biology of hair follicles. N. Engl. J. Med. 1999, 341, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Danilenko, D.M.; Ring, B.D.; Pierce, G.F. Growth factors and cytokines in hair follicle development and cycling: Recent insights from animal models and the potentials for clinical therapy. Mol. Med. Today 1996, 2, 460–467. [Google Scholar] [CrossRef]
- Kos, L.; Conlon, J. An update on alopecia areata. Curr. Opin. Pediatr. 2009, 21, 475–480. [Google Scholar] [CrossRef]
- Wasserman, D.; Guzman-Sanchez, D.A.; Scott, K.; McMichael, A. Alopecia areata. Int. J. Derm. 2007, 46, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.; Sinclair, R. Telogen effluvium. Clin. Exp. Derm. 2002, 27, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Tosti, A.; Piraccini, B.M.; Sisti, A.; Duque-Estrada, B. Hair loss in women. Minerva Ginecol. 2009, 61, 445–452. [Google Scholar]
- Harries, M.J.; Paus, R. The pathogenesis of primary cicatricial alopecias. Am. J. Pathol. 2010, 177, 2152–2162. [Google Scholar] [CrossRef] [PubMed]
- Messenger, A.G.; Rundegren, J. Minoxidil: Mechanisms of action on hair growth. Br. J. Derm. 2004, 150, 186–194. [Google Scholar] [CrossRef]
- Kaufman, K.D.; Olsen, E.A.; Whiting, D.; Savin, R.; DeVillez, R.; Bergfeld, W.; Price, V.H.; Van Neste, D.; Roberts, J.L.; Hordinsky, M. Finasteride in the treatment of men with androgenetic alopecia. J. Am. Acad. Dermatol. 1998, 39, 578–589. [Google Scholar] [CrossRef]
- Jain, P.K.; Das, D.; Singhai, A. Alternative herbal drugs used for treating hair disease. Asian J. Pharm. Clin. Res. 2016, 9, 75–77. [Google Scholar]
- Działo, M.; Mierziak, J.; Korzun, U.; Preisner, M.; Szopa, J.; Kulma, A. The potential of plant phenolics in prevention and therapy of skin disorders. Int. J. Mol. Sci. 2016, 17, 160. [Google Scholar] [CrossRef] [PubMed]
- Woo, H.; Lee, S.; Kim, S.; Park, D.; Jung, E. Effect of sinapic acid on hair growth promoting in human hair follicle dermal papilla cells via Akt activation. Arch. Derm. Res. 2017, 309, 381–388. [Google Scholar] [CrossRef]
- Su, Y.S.; Fan, Z.X.; Xiao, S.E.; Lin, B.J.; Miao, Y.; Hu, Z.Q.; Liu, H. Icariin promotes mouse hair follicle growth by increasing insulin-like growth factor 1 expression in dermal papillary cells. Clin. Exp. Derm. 2017, 42, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Boisvert, W.A.; Yu, M.; Choi, Y.; Jeong, G.H.; Zhang, Y.L.; Cho, S.; Choi, C.; Lee, S.; Lee, B.H. Hair growth-promoting effect of Geranium sibiricum extract in human dermal papilla cells and C57BL/6 mice. BMC Complement. Altern. Med. 2017, 17, 109. [Google Scholar] [CrossRef]
- Tong, T.; Kim, N.; Park, T. Topical Application of Oleuropein Induces Anagen Hair Growth in Telogen Mouse Skin. PLoS ONE 2015, 10, e0129578. [Google Scholar] [CrossRef]
- Fischer, T.W.; Herczeg-Lisztes, E.; Funk, W.; Zillikens, D.; Biro, T.; Paus, R. Differential effects of caffeine on hair shaft elongation, matrix and outer root sheath keratinocyte proliferation, and transforming growth factor-beta2/insulin-like growth factor-1-mediated regulation of the hair cycle in male and female human hair follicles in vitro. Br. J. Derm. 2014, 171, 1031–1043. [Google Scholar] [CrossRef]
- Junlatat, J.; Sripanidkulchai, B. Hair growth-promoting effect of Carthamus tinctorius floret extract. Phytother. Res. 2014, 28, 1030–1036. [Google Scholar] [CrossRef]
- Truong, V.L.; Bak, M.J.; Lee, C.; Jun, M.; Jeong, W.S. Hair Regenerative Mechanisms of Red Ginseng Oil and Its Major Components in the Testosterone-Induced Delay of Anagen Entry in C57BL/6 Mice. Molecules 2017, 22. [Google Scholar] [CrossRef]
- Kim, J.; Kim, S.R.; Choi, Y.H.; Shin, J.Y.; Kim, C.D.; Kang, N.G.; Park, B.C.; Lee, S. Quercitrin Stimulates Hair Growth with Enhanced Expression of Growth Factors via Activation of MAPK/CREB Signaling Pathway. Molecules 2020, 25, 4004. [Google Scholar] [CrossRef] [PubMed]
- Roh, S.S.; Kim, C.D.; Lee, M.H.; Hwang, S.L.; Rang, M.J.; Yoon, Y.K. The hair growth promoting effect of Sophora flavescens extract and its molecular regulation. J. Derm. Sci. 2002, 30, 43–49. [Google Scholar] [CrossRef]
- Choi, M.; Choi, S.J.; Jang, S.; Choi, H.I.; Kang, B.M.; Hwang, S.T.; Kwon, O. Shikimic acid, a mannose bioisostere, promotes hair growth with the induction of anagen hair cycle. Sci. Rep. 2019, 9, 17008. [Google Scholar] [CrossRef]
- Lim, W.; Mudge, K.W.; Vermeylen, F. Effects of population, age, and cultivation methods on ginsenoside content of wild American ginseng (Panax quinquefolium). J. Agric. Food Chem. 2005, 53, 8498–8505. [Google Scholar] [CrossRef]
- Zhang, H.; Su, Y.; Wang, J.; Gao, Y.; Yang, F.; Li, G.; Shi, Q. Ginsenoside Rb1 promotes the growth of mink hair follicle via PI3K/AKT/GSK-3beta signaling pathway. Life Sci. 2019, 229, 210–218. [Google Scholar] [CrossRef]
- Park, H.J.; Jin, G.R.; Jung, J.H.; Hwang, S.B.; Lee, S.H.; Lee, B.H. Hair Growth Promotion Effect of Nelumbinis Semen Extract with High Antioxidant Activity. Evid. Based Complement. Altern. Med. 2021, 2021, 6661373. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.O.; Kim, S.E.; Kim, Y.C. Action Mechanism of Chamaecyparis obtusa Oil on Hair Growth. Toxicol. Res. 2013, 29, 241–247. [Google Scholar] [CrossRef]
- Shin, J.Y.; Choi, Y.H.; Kim, J.; Park, S.Y.; Nam, Y.J.; Lee, S.Y.; Jeon, J.H.; Jin, M.H.; Lee, S. Polygonum multiflorum extract support hair growth by elongating anagen phase and abrogating the effect of androgen in cultured human dermal papilla cells. BMC Complement. Med. Ther. 2020, 20, 144. [Google Scholar] [CrossRef]
- Ha, E.J.; Yun, J.H.; Si, C.; Bae, Y.S.; Jeong, Y.H.; Park, K.H.; Choi, S.E. Application of Ethanol Extracts From Alnus sibirica Fisch. ex Turcz in Hair Growth Promotion. Front. Bioeng. Biotechnol. 2021, 9, 673314. [Google Scholar] [CrossRef]
- Lee, E.Y.; Choi, E.J.; Kim, J.A.; Hwang, Y.L.; Kim, C.D.; Lee, M.H.; Roh, S.S.; Kim, Y.H.; Han, I.; Kang, S. Malva verticillata seed extracts upregulate the Wnt pathway in human dermal papilla cells. Int. J. Cosmet. Sci. 2016, 38, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H.S.; Jeong, J.; Lee, C.M.; Lee, K.S.; Lee, J.N.; Park, S.M.; Lee, Y.M. Activation of Hair Cell Growth Factors by Linoleic Acid in Malva verticillata Seed. Molecules 2021, 26. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, J.; Chen, F.; Wang, C.; Guo, X.; Wang, C.; Fan, Y.; Wang, Y.; Peng, Y.; Li, W. Liposomal honokiol promotes hair growth via activating Wnt3a/beta-catenin signaling pathway and down regulating TGF-beta1 in C57BL/6N mice. Biomed. Pharm. 2021, 141, 111793. [Google Scholar] [CrossRef]
- Lee, T.K.; Kim, B.; Kim, D.W.; Ahn, J.H.; Sim, H.; Lee, J.C.; Yang, G.E.; Her, Y.; Park, J.H.; Kim, H.S.; et al. Effects of Decursin and Angelica gigas Nakai Root Extract on Hair Growth in Mouse Dorsal Skin via Regulating Inflammatory Cytokines. Molecules 2020, 25, 3697. [Google Scholar] [CrossRef]
- Kim, Y.E.; Choi, H.C.; Lee, I.C.; Yuk, D.Y.; Lee, H.; Choi, B.Y. 3-Deoxysappanchalcone Promotes Proliferation of Human Hair Follicle Dermal Papilla Cells and Hair Growth in C57BL/6 Mice by Modulating WNT/beta-Catenin and STAT Signaling. Biomol. Ther. 2016, 24, 572–580. [Google Scholar] [CrossRef]
- Lee, Y.H.; Nam, G.; Kim, M.-K.; Cho, S.-C.; Choi, B.Y. Broussonetia papyrifera Promotes Hair Growth Through the Regulation of β-Catenin and STAT6 Target Proteins: A Phototrichogram Analysis of Clinical Samples. Cosmetics 2020, 7, 40. [Google Scholar] [CrossRef]
- Truong, V.L.; Keum, Y.S.; Jeong, W.S. Red ginseng oil promotes hair growth and protects skin against UVC radiation. J. Ginseng Res. 2021, 45, 498–509. [Google Scholar] [CrossRef]
- Zhang, N.N.; Park, D.K.; Park, H.J. Hair growth-promoting activity of hot water extract of Thuja orientalis. BMC Complement. Altern. Med. 2013, 13, 9. [Google Scholar] [CrossRef]
- Bejaoui, M.; Villareal, M.O.; Isoda, H. β-catenin-mediated hair growth induction effect of 3, 4, 5-tri-O-caffeoylquinic acid. Aging 2019, 11, 4216. [Google Scholar] [CrossRef]
- Kang, J.I.; Kim, E.J.; Kim, M.K.; Jeon, Y.J.; Kang, S.M.; Koh, Y.S.; Yoo, E.S.; Kang, H.K. The promoting effect of Ishige sinicola on hair growth. Mar. Drugs 2013, 11, 1783–1799. [Google Scholar] [CrossRef]
- Zhou, Y.; Tang, G.; Li, X.; Sun, W.; Liang, Y.; Gan, D.; Liu, G.; Song, W.; Wang, Z. Study on the chemical constituents of nut oil from Prunus mira Koehne and the mechanism of promoting hair growth. J. Ethnopharmacol. 2020, 258, 112831. [Google Scholar] [CrossRef]
- Kim, Y.E.; Choi, H.C.; Nam, G.; Choi, B.Y. Costunolide promotes the proliferation of human hair follicle dermal papilla cells and induces hair growth in C57BL/6 mice. J. Cosmet. Derm. 2019, 18, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, H.; Jing, J.; Yu, L.; Wu, X.; Lu, Z. Morroniside regulates hair growth and cycle transition via activation of the Wnt/β-catenin signaling pathway. Sci. Rep. 2018, 8, 13785. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Zhang, X.; Chen, Z.; Li, J.; Li, B.; Li, L. Molecular Pathways Involved in Promoting Activity of Timosaponin BII on Hair Growth in C57BL/6 Mice. Biomed. Res. Int. 2020, 2020, 9451596. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.R.; Zhang, Y.L.; Yap, J.; Boisvert, W.A.; Lee, B.H. Hair growth potential of Salvia plebeia extract and its associated mechanisms. Pharm. Biol. 2020, 58, 400–409. [Google Scholar] [CrossRef]
- Kang, J.-I.; Kim, M.-K.; Lee, J.-H.; Jeon, Y.-J.; Hwang, E.-K.; Koh, Y.-S.; Hyun, J.-W.; Kwon, S.-Y.; Yoo, E.-S.; Kang, H.-K. Undariopsis peterseniana promotes hair growth by the activation of Wnt/β-catenin and ERK pathways. Mar. Drugs 2017, 15, 130. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Noguchi, K.; Kondo, M.; Onishi, M.; Watanabe, N.; Okamura, K.; Matsuda, H. Inhibitory activities of Puerariae Flos against testosterone 5alpha-reductase and its hair growth promotion activities. J. Nat. Med. 2012, 66, 158–165. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, R.; Yin, X.; Lao, Z.; Zhang, Z.; Wu, Q.; Yu, L.; Lai, X.; Wan, Y.; Li, G. Inhibitory activities of some traditional Chinese herbs against testosterone 5α-reductase and effects of Cacumen platycladi on hair re-growth in testosterone-treated mice. J. Ethnopharmacol. 2016, 177, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Takeshita, F.; Samukawa, K.; Tani, T.; Matsuda, H. Effects of ginseng rhizome and ginsenoside Ro on testosterone 5alpha-reductase and hair re-growth in testosterone-treated mice. Phytother. Res. 2012, 26, 48–53. [Google Scholar] [CrossRef]
- Lao, Z.; Fan, Y.; Huo, Y.; Liao, F.; Zhang, R.; Zhang, B.; Kong, Z.; Long, H.; Xie, J.; Sang, C.; et al. Physcion, a novel inhibitor of 5α-reductase that promotes hair growth in vitro and in vivo. Arch. Derm. Res. 2021. [Google Scholar] [CrossRef]
- Murata, K.; Noguchi, K.; Kondo, M.; Onishi, M.; Watanabe, N.; Okamura, K.; Matsuda, H. Promotion of hair growth by Rosmarinus officinalis leaf extract. Phytother. Res. 2013, 27, 212–217. [Google Scholar] [CrossRef]
- Jain, R.; Monthakantirat, O.; Tengamnuay, P.; De-Eknamkul, W. Avicequinone C isolated from Avicennia marina exhibits 5α-reductase-type 1 inhibitory activity using an androgenic alopecia relevant cell-based assay system. Molecules 2014, 19, 6809–6821. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Yoo, H.G.; Inui, S.; Itami, S.; Kim, I.G.; Cho, A.-R.; Lee, D.H.; Park, W.S.; Kwon, O.; Cho, K.H. Induction of transforming growth factor-beta 1 by androgen is mediated by reactive oxygen species in hair follicle dermal papilla cells. BMB Rep. 2013, 46, 460. [Google Scholar] [CrossRef]
- Zhang, H.; Nan, W.; Wang, S.; Song, X.; Si, H.; Li, T.; Li, G. Epigallocatechin-3-Gallate promotes the growth of mink hair follicles through sonic hedgehog and protein kinase B signaling pathways. Front. Pharmacol. 2018, 9, 674. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, S.N.; Hong, Y.D.; Park, B.C.; Na, Y. Panax ginseng extract antagonizes the effect of DKK1-induced catagen-like changes of hair follicles. Int. J. Mol. Med. 2017, 40, 1194–1200. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.L.; Gao, Y.H.; Yang, J.Q.; Li, J.B.; Gao, J. Serenoa repens extracts promote hair regeneration and repair of hair loss mouse models by activating TGF-beta and mitochondrial signaling pathway. Eur. Rev. Med. Pharm. Sci. 2018, 22, 4000–4008. [Google Scholar] [CrossRef]
- Miele, C.; Rochford, J.J.; Filippa, N.; Giorgetti-Peraldi, S.; Van Obberghen, E. Insulin and insulin-like growth factor-I induce vascular endothelial growth factor mRNA expression via different signaling pathways. J. Biol. Chem. 2000, 275, 21695–21702. [Google Scholar] [CrossRef] [PubMed]
- Ozeki, M.; Tabata, Y. In vivo promoted growth of mice hair follicles by the controlled release of growth factors. Biomaterials 2003, 24, 2387–2394. [Google Scholar] [CrossRef]
- Peus, D.; Pittelkow, M.R. Growth factors in hair organ development and the hair growth cycle. Derm. Clin. 1996, 14, 559–572. [Google Scholar] [CrossRef]
- Philpott, M.P.; Sanders, D.A.; Kealey, T. Effects of insulin and insulin-like growth factors on cultured human hair follicles: IGF-I at physiologic concentrations is an important regulator of hair follicle growth in vitro. J. Investig. Dermatol. 1994, 102, 857–861. [Google Scholar] [CrossRef]
- Castela, M.; Linay, F.; Roy, E.; Moguelet, P.; Xu, J.; Holzenberger, M.; Khosrotehrani, K.; Aractingi, S. Igf1r signalling acts on the anagen-to-catagen transition in the hair cycle. Exp. Dermatol. 2017, 26, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.X.; Lin, C.; Lin, B.B.; Wang, Z.G.; Zhang, H.Y.; Wu, F.Z.; Cheng, Y.; Xiang, L.J.; Guo, D.J.; Luo, X.; et al. The anti-scar effects of basic fibroblast growth factor on the wound repair in vitro and in vivo. PLoS ONE 2013, 8, e59966. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H. Stimulation of human hair growth by the recombinant human keratinocyte growth factor-2 (KGF-2). Biotechnol. Lett. 2005, 27, 749–752. [Google Scholar] [CrossRef] [PubMed]
- Jindo, T.; Tsuboi, R.; Imai, R.; Takamori, K.; Rubin, J.S.; Ogawa, H. Hepatocyte growth factor/scatter factor stimulates hair growth of mouse vibrissae in organ culture. J. Investig. Dermatol. 1994, 103, 306–309. [Google Scholar] [CrossRef][Green Version]
- Jindo, T.; Tsuboi, R.; Takamori, K.; Ogawa, H. Local injection of hepatocyte growth factor/scatter factor (HGF/SF) alters cyclic growth of murine hair follicles. J. Investig. Dermatol. 1998, 110, 338–342. [Google Scholar] [CrossRef]
- Soma, T.; Dohrmann, C.E.; Hibino, T.; Raftery, L.A. Profile of transforming growth factor-beta responses during the murine hair cycle. J. Investig. Dermatol. 2003, 121, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Chen, C. Sinapic Acid and Its Derivatives as Medicine in Oxidative Stress-Induced Diseases and Aging. Oxid. Med. Cell Longev. 2016, 2016, 3571614. [Google Scholar] [CrossRef]
- Huang, X.; Pan, Q.; Mao, Z.; Zhang, R.; Ma, X.; Xi, Y.; You, H. Sinapic Acid Inhibits the IL-1beta-Induced Inflammation via MAPK Downregulation in Rat Chondrocytes. Inflammation 2018, 41, 562–568. [Google Scholar] [CrossRef]
- Yun, K.J.; Koh, D.J.; Kim, S.H.; Park, S.J.; Ryu, J.H.; Kim, D.G.; Lee, J.Y.; Lee, K.T. Anti-inflammatory effects of sinapic acid through the suppression of inducible nitric oxide synthase, cyclooxygase-2, and proinflammatory cytokines expressions via nuclear factor-kappaB inactivation. J. Agric. Food Chem. 2008, 56, 10265–10272. [Google Scholar] [CrossRef]
- Eroglu, C.; Avci, E.; Vural, H.; Kurar, E. Anticancer mechanism of Sinapic acid in PC-3 and LNCaP human prostate cancer cell lines. Gene 2018, 671, 127–134. [Google Scholar] [CrossRef]
- Cherng, Y.G.; Tsai, C.C.; Chung, H.H.; Lai, Y.W.; Kuo, S.C.; Cheng, J.T. Antihyperglycemic action of sinapic acid in diabetic rats. J. Agric. Food Chem. 2013, 61, 12053–12059. [Google Scholar] [CrossRef]
- Lee, H.E.; Kim, D.H.; Park, S.J.; Kim, J.M.; Lee, Y.W.; Jung, J.M.; Lee, C.H.; Hong, J.G.; Liu, X.; Cai, M.; et al. Neuroprotective effect of sinapic acid in a mouse model of amyloid beta(1-42) protein-induced Alzheimer’s disease. Pharm. Biochem. Behav. 2012, 103, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.B.; Yang, Q.T. The testosterone mimetic properties of icariin. Asian J. Ther. 2006, 8, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Nian, H.; Ma, M.H.; Nian, S.S.; Xu, L.L. Antiosteoporotic activity of icariin in ovariectomized rats. Phytomedicine 2009, 16, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Kong, L.; Xia, X.; Zhang, W.; Xia, Z.; Jiang, F. Antidepressant-like effect of icariin and its possible mechanism in mice. Pharm. Biochem. Behav. 2005, 82, 686–694. [Google Scholar] [CrossRef]
- Shim, J.U.; Lim, K.T. Antioxidative activity of glycoprotein isolated from Geranium sibiricum Linne. Nat. Prod. Res. 2009, 23, 375–387. [Google Scholar] [CrossRef]
- Shim, J.U.; Oh, P.S.; Lim, K.T. Anti-inflammatory activity of ethanol extract from Geranium sibiricum Linne. J. Ethnopharmacol. 2009, 126, 90–95. [Google Scholar] [CrossRef]
- Wu, N.; Zu, Y.; Fu, Y.; Kong, Y.; Zhao, J.; Li, X.; Li, J.; Wink, M.; Efferth, T. Antioxidant activities and xanthine oxidase inhibitory effects of extracts and main polyphenolic compounds obtained from Geranium sibiricum L. J. Agric. Food Chem. 2010, 58, 4737–4743. [Google Scholar] [CrossRef] [PubMed]
- Barbaro, B.; Toietta, G.; Maggio, R.; Arciello, M.; Tarocchi, M.; Galli, A.; Balsano, C. Effects of the olive-derived polyphenol oleuropein on human health. Int. J. Mol. Sci. 2014, 15, 18508–18524. [Google Scholar] [CrossRef] [PubMed]
- Shamshoum, H.; Vlavcheski, F.; Tsiani, E. Anticancer effects of oleuropein. Biofactors 2017, 43, 517–528. [Google Scholar] [CrossRef]
- Omar, S.H. Cardioprotective and neuroprotective roles of oleuropein in olive. Saudi Pharm. J. 2010, 18, 111–121. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Battig, K.; Holmen, J.; Nehlig, A.; Zvartau, E.E. Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharm. Rev. 1999, 51, 83–133. [Google Scholar]
- Kolahdouzan, M.; Hamadeh, M.J. The neuroprotective effects of caffeine in neurodegenerative diseases. CNS Neurosci. Ther. 2017, 23, 272–290. [Google Scholar] [CrossRef] [PubMed]
- Hall, S.; Desbrow, B.; Anoopkumar-Dukie, S.; Davey, A.K.; Arora, D.; McDermott, C.; Schubert, M.M.; Perkins, A.V.; Kiefel, M.J.; Grant, G.D. A review of the bioactivity of coffee, caffeine and key coffee constituents on inflammatory responses linked to depression. Food Res. Int. 2015, 76, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Sabisz, M.; Skladanowski, A. Modulation of cellular response to anticancer treatment by caffeine: Inhibition of cell cycle checkpoints, DNA repair and more. Curr. Pharm. Biotechnol. 2008, 9, 325–336. [Google Scholar] [CrossRef]
- Zhou, X.; Tang, L.; Xu, Y.; Zhou, G.; Wang, Z. Towards a better understanding of medicinal uses of Carthamus tinctorius L. in traditional Chinese medicine: A phytochemical and pharmacological review. J. Ethnopharmacol. 2014, 151, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Delshad, E.; Yousefi, M.; Sasannezhad, P.; Rakhshandeh, H.; Ayati, Z. Medical uses of Carthamus tinctorius L. (Safflower): A comprehensive review from Traditional Medicine to Modern Medicine. Electron. Physician 2018, 10, 6672–6681. [Google Scholar] [CrossRef]
- Kumar, N.; Rungseevijitprapa, W.; Narkkhong, N.A.; Suttajit, M.; Chaiyasut, C. 5alpha-reductase inhibition and hair growth promotion of some Thai plants traditionally used for hair treatment. J. Ethnopharmacol. 2012, 139, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.J.; Jun, M.; Jeong, W.S. Antioxidant and hepatoprotective effects of the red ginseng essential oil in H(2)O(2)-treated hepG2 cells and CCl(4)-treated mice. Int. J. Mol. Sci. 2012, 13, 2314–2330. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.J.; Hong, S.G.; Lee, J.W.; Jeong, W.S. Red ginseng marc oil inhibits iNOS and COX-2 via NfkappaB and p38 pathways in LPS-stimulated RAW 264.7 macrophages. Molecules 2012, 17, 13769–13786. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Youn, K.; Jun, M. Major compounds of red ginseng oil attenuate Abeta25-35-induced neuronal apoptosis and inflammation by modulating MAPK/NF-kappaB pathway. Food Funct. 2018, 9, 4122–4134. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.J.; Truong, V.L.; Ko, S.Y.; Nguyen, X.N.; Jun, M.; Hong, S.G.; Lee, J.W.; Jeong, W.S. Induction of Nrf2/ARE-mediated cytoprotective genes by red ginseng oil through ASK1-MKK4/7-JNK and p38 MAPK signaling pathways in HepG2 cells. J. Ginseng Res. 2016, 40, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Oh, K.H.; Chung, A.Y.; Park, H.C.; Lee, S.H.; Kwon, S.Y.; Choi, J. Protective role of quercetin against cisplatin-induced hair cell damage in zebrafish embryos. Hum. Exp. Toxicol. 2015, 34, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.H.; Kim, J.S.; Kang, S.S.; Son, K.H.; Chang, H.W.; Kim, H.P. Anti-inflammatory and anti-arthritic activity of total flavonoids of the roots of Sophora flavescens. J. Ethnopharmacol. 2010, 127, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Piao, X.L.; Piao, X.S.; Kim, S.W.; Park, J.H.; Kim, H.Y.; Cai, S.Q. Identification and characterization of antioxidants from Sophora flavescens. Biol. Pharm. Bull. 2006, 29, 1911–1915. [Google Scholar] [CrossRef] [PubMed]
- Batory, M.; Rotsztejn, H. Shikimic acid in the light of current knowledge. J. Cosmet. Dermatol. 2021. [Google Scholar] [CrossRef]
- Quinde-Axtell, Z.; Baik, B.K. Phenolic compounds of barley grain and their implication in food product discoloration. J. Agric. Food Chem. 2006, 54, 9978–9984. [Google Scholar] [CrossRef]
- Kamimura, A.; Takahashi, T. Procyanidin B-3, isolated from barley and identified as a hair-growth stimulant, has the potential to counteract inhibitory regulation by TGF-β1. Exp. Dermatol. 2002, 11, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.Y. Hair-Growth Potential of Ginseng and Its Major Metabolites: A Review on Its Molecular Mechanisms. Int. J. Mol. Sci. 2018, 19, 2703. [Google Scholar] [CrossRef]
- Huang, B.; Ban, X.; He, J.; Tong, J.; Tian, J.; Wang, Y. Comparative analysis of essential oil components and antioxidant activity of extracts of Nelumbo nucifera from various areas of China. J. Agric. Food Chem. 2010, 58, 441–448. [Google Scholar] [CrossRef]
- Arooj, M.; Imran, S.; Inam-Ur-Raheem, M.; Rajoka, M.S.R.; Sameen, A.; Siddique, R.; Sahar, A.; Tariq, S.; Riaz, A.; Hussain, A.; et al. Lotus seeds (Nelumbinis semen) as an emerging therapeutic seed: A comprehensive review. Food Sci. Nutr. 2021, 9, 3971–3987. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, X.; Zeng, S.; Huang, X.; Guo, Z.; Zheng, Y.; Tian, Y.; Zheng, B. Nutritional composition, physiological functions and processing of lotus (Nelumbo nucifera Gaertn.) seeds: A review. Phytochem. Rev. 2015, 14, 321–334. [Google Scholar] [CrossRef]
- Yang, J.K.; Choi, M.S.; Seo, W.T.; Rinker, D.L.; Han, S.W.; Cheong, G.W. Chemical composition and antimicrobial activity of Chamaecyparis obtusa leaf essential oil. Fitoterapia 2007, 78, 149–152. [Google Scholar] [CrossRef]
- Bae, M.S.; Park, D.H.; Choi, C.Y.; Kim, G.Y.; Yoo, J.C.; Cho, S.S. Essential Oils and Non-volatile Compounds Derived from Chamaecyparis obtusa: Broad Spectrum Antimicrobial Activity against Infectious Bacteria and MDR (multidrug resistant) Strains. Nat. Prod. Commun. 2016, 11, 693–694. [Google Scholar] [CrossRef]
- An, B.S.; Kang, J.H.; Yang, H.; Jung, E.M.; Kang, H.S.; Choi, I.G.; Park, M.J.; Jeung, E.B. Anti-inflammatory effects of essential oils from Chamaecyparis obtusa via the cyclooxygenase-2 pathway in rats. Mol. Med. Rep. 2013, 8, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Jung, S.M.; Yoo, S.A.; Kim, W.U.; Cho, C.S.; Park, B.J.; Woo, J.M.; Yoon, C.H. Antinociceptive and anti-inflammatory effects of essential oil extracted from Chamaecyparis obtusa in mice. Int. Immunopharmacol. 2015, 29, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Bounda, G.A.; Feng, Y.U. Review of clinical studies of Polygonum multiflorum Thunb. and its isolated bioactive compounds. Pharmacogn. Res. 2015, 7, 225–236. [Google Scholar] [CrossRef]
- Lv, L.; Gu, X.H.; Tang, J.; Ho, C.T. Antioxidant activity of stilbene glycoside from Polygonum multiflorum Thunb in vivo. Food Chem. 2007, 104, 1678–1681. [Google Scholar] [CrossRef]
- Lin, H.W.; Sun, M.X.; Wang, Y.H.; Yang, L.M.; Yang, Y.R.; Huang, N.; Xuan, L.J.; Xu, Y.M.; Bai, D.L.; Zheng, Y.T.; et al. Anti-HIV activities of the compounds isolated from Polygonum cuspidatum and Polygonum multiflorum. Planta Med. 2010, 76, 889–892. [Google Scholar] [CrossRef]
- Li, X.; Matsumoto, K.; Murakami, Y.; Tezuka, Y.; Wu, Y.; Kadota, S. Neuroprotective effects of Polygonum multiflorum on nigrostriatal dopaminergic degeneration induced by paraquat and maneb in mice. Pharm. Biochem. Behav. 2005, 82, 345–352. [Google Scholar] [CrossRef]
- Um, M.Y.; Choi, W.H.; Aan, J.Y.; Kim, S.R.; Ha, T.Y. Protective effect of Polygonum multiflorum Thunb on amyloid beta-peptide 25-35 induced cognitive deficits in mice. J. Ethnopharmacol. 2006, 104, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Quan, Y.; Gong, L.; Gong, X.; Li, Y. A review of the processed Polygonum multiflorum (Thunb.) for hepatoprotection: Clinical use, pharmacology and toxicology. J. Ethnopharmacol. 2020, 261, 113121. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, Y.; Genjida, F.; Suga, T. Four new flavonoids isolated from Alnus sieboldiana. Bull. Chem. Soc. Jpn. 1971, 44, 297. [Google Scholar] [CrossRef]
- Ishimatsu, M.; Tanaka, T.; Nonaka, G.-i.; Nishioka, I. Alnusnins A and B from the leaves of Alnus sieboldiana. Phytochemistry 1989, 28, 3179–3184. [Google Scholar] [CrossRef]
- Yoshida, T.; Yazaki, K.; Memon, M.U.; Maruyama, I.; Kurokawa, K.; Shingu, T.; Okuda, T. Structures of alnusiin and bicornin, new hydrolyzable tannins having a monolactonized tergalloyl group. Chem. Pharm. Bull. 1989, 37, 2655–2660. [Google Scholar] [CrossRef]
- Kuroyanagi, M.; Shimomae, M.; Nagashima, Y.; Muto, N.; Okuda, T.; Kawahara, N.; Nakane, T.; Sano, T. New diarylheptanoids from Alnus japonica and their antioxidative activity. Chem. Pharm. Bull. 2005, 53, 1519–1523. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, L.P.; Bovini, M.G.; da Costa Bortoluzzi, R.L.; Boff, M.I.C.; Boff, P. Species of Malva L.(Malvaceae) cultivated in the western of santa Catarina state and conformity with species marketed as medicinal plants in southern Brazil. J. Agric. Sci. 2019, 11, 171–180. [Google Scholar] [CrossRef]
- Bao, L.; Bao, X.; Li, P.; Wang, X.; Ao, W. Chemical profiling of Malva verticillata L. by UPLC-Q-TOF-MSE and their antioxidant activity in vitro. J. Pharm. Biomed. Anal. 2018, 150, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Tomoda, M.; Shimizu, N.; Gonda, R.; Kanari, M.; Yamada, H.; Hikino, H. Anti-complementary and hypoglycemic activities of the glycans from the seeds of Malva verticillata. Planta Med. 1990, 56, 168–170. [Google Scholar] [CrossRef]
- Jeong, Y.T.; Song, C.H. Antidiabetic activities of extract from Malva verticillata seed via the activation of AMP-activated protein kinase. J. Microbiol. Biotechnol. 2011, 21, 921–929. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Q.; Yang, G.; Fan, L.; Tang, J.; Garrard, I.; Ignatova, S.; Fisher, D.; Sutherland, I.A. Rapid purification and scale-up of honokiol and magnolol using high-capacity high-speed counter-current chromatography. J. Chromatogr. A 2007, 1142, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.M.; Chen, C.C.; Ko, F.N.; Lee, L.G.; Huang, T.F.; Chen, Y.P.; Hsu, H.Y. Two antiplatelet agents from Magnolia officinalis. Thromb. Res. 1988, 50, 757–765. [Google Scholar] [CrossRef]
- Wang, J.; Liu, D.; Guan, S.; Zhu, W.; Fan, L.; Zhang, Q.; Cai, D. Hyaluronic acid-modified liposomal honokiol nanocarrier: Enhance anti-metastasis and antitumor efficacy against breast cancer. Carbohydr. Polym. 2020, 235, 115981. [Google Scholar] [CrossRef]
- Jiang, Q.Q.; Fan, L.Y.; Yang, G.L.; Guo, W.H.; Hou, W.L.; Chen, L.J.; Wei, Y.Q. Improved therapeutic effectiveness by combining liposomal honokiol with cisplatin in lung cancer model. BMC Cancer 2008, 8, 242. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, R.; Eicheler, W.; Huth, A.; Wenzel, E.; Happle, R. Cytokines and growth factors influence hair growth in vitro. Possible implications for the pathogenesis and treatment of alopecia areata. Arch. Derm. Res. 1996, 288, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Philpott, M.P.; Sanders, D.A.; Bowen, J.; Kealey, T. Effects of interleukins, colony-stimulating factor and tumour necrosis factor on human hair follicle growth in vitro: A possible role for interleukin-1 and tumour necrosis factor-alpha in alopecia areata. Br. J. Dermatol. 1996, 135, 942–948. [Google Scholar] [CrossRef]
- Hull, S.M.; Nutbrown, M.; Pepall, L.; Thornton, M.J.; Randall, V.A.; Cunliffe, W.J. Immunohistologic and ultrastructural comparison of the dermal papilla and hair follicle bulb from “active” and “normal” areas of alopecia areata. J. Investig. Dermatol. 1991, 96, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Groves, R.W.; Mizutani, H.; Kieffer, J.D.; Kupper, T.S. Inflammatory skin disease in transgenic mice that express high levels of interleukin 1 alpha in basal epidermis. Proc. Natl. Acad. Sci. USA 1995, 92, 11874–11878. [Google Scholar] [CrossRef] [PubMed]
- Ruckert, R.; Lindner, G.; Bulfone-Paus, S.; Paus, R. High-dose proinflammatory cytokines induce apoptosis of hair bulb keratinocytes in vivo. Br. J. Dermatol. 2000, 143, 1036–1039. [Google Scholar] [CrossRef] [PubMed]
- Teraki, Y.; Imanishi, K.; Shiohara, T. Cytokines in alopecia areata: Contrasting cytokine profiles in localized form and extensive form (alopecia universalis). Acta Dermatol. Venereol. 1996, 76, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, L.; Jiang, C.; Xing, C.; Kim, S.H.; Lu, J. Anti-cancer and other bioactivities of Korean Angelica gigas Nakai (AGN) and its major pyranocoumarin compounds. Anticancer Agents Med. Chem. 2012, 12, 1239–1254. [Google Scholar] [CrossRef]
- Choi, S.S.; Han, K.J.; Lee, H.K.; Han, E.J.; Suh, H.W. Antinociceptive profiles of crude extract from roots of Angelica gigas NAKAI in various pain models. Biol. Pharm. Bull. 2003, 26, 1283–1288. [Google Scholar] [CrossRef]
- Oh, T.W.; Park, K.H.; Jung, H.W.; Park, Y.K. Neuroprotective effect of the hairy root extract of Angelica gigas NAKAI on transient focal cerebral ischemia in rats through the regulation of angiogenesis. BMC Complement. Altern. Med. 2015, 15, 101. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Lim, C.; Cho, S. Angelica gigas root ameliorates ischaemic stroke-induced brain injury in mice by activating the PI3K/AKT/mTOR and MAPK pathways. Pharm. Biol. 2021, 59, 662–671. [Google Scholar] [CrossRef]
- Kim, J.H.; Choo, Y.Y.; Tae, N.; Min, B.S.; Lee, J.H. The anti-inflammatory effect of 3-deoxysappanchalcone is mediated by inducing heme oxygenase-1 via activating the AKT/mTOR pathway in murine macrophages. Int. Immunopharmacol. 2014, 22, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Huang, H.; Choi, B.Y.; Liu, X.; Zhang, M.; Zhou, S.; Song, M.; Yin, F.; Chen, H.; Shim, J.H.; et al. Cell growth inhibition by 3-deoxysappanchalcone is mediated by directly targeting the TOPK signaling pathway in colon cancer. Phytomedicine 2019, 61, 152813. [Google Scholar] [CrossRef]
- Yodsaoue, O.; Cheenpracha, S.; Karalai, C.; Ponglimanont, C.; Tewtrakul, S. Anti-allergic activity of principles from the roots and heartwood of Caesalpinia sappan on antigen-induced beta-hexosaminidase release. Phytother. Res. 2009, 23, 1028–1031. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.C.; Nam, G.W.; Jeong, N.H.; Choi, B.Y. Hair growth promotion by extracts of Inula Helenium and Caesalpinia Sappan Bark in patients with androgenetic alopecia: A pre-clinical study using phototrichogram analysis. Cosmetics 2019, 6, 66. [Google Scholar] [CrossRef]
- Lee, D.; Bhat, K.P.; Fong, H.H.; Farnsworth, N.R.; Pezzuto, J.M.; Kinghorn, A.D. Aromatase inhibitors from Broussonetia papyrifera. J. Nat. Prod. 2001, 64, 1286–1293. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Wu, Z.; Huang, B.; Sun, L.; Ding, C.; Yuan, S.; Zhang, Z.; Chen, Y.; Hu, C.; Zhou, L.; et al. Extraction, antioxidant and antibacterial activities of Broussonetia papyrifera fruits polysaccharides. Int. J. Biol. Macromol. 2016, 92, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H.W.; Park, M.H.; Kwon, O.K.; Kim, D.Y.; Hwang, J.Y.; Jo, Y.H.; Ahn, K.S.; Hwang, B.Y.; Oh, S.R. Anti-inflammatory flavonoids from root bark of Broussonetia papyrifera in LPS-stimulated RAW264.7 cells. Bioorg. Chem. 2019, 92, 103233. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Zhang, Y.; Xu, M.; Yang, Y.; Ito, M.; Peng, T.; Cui, Z.; Nagy, A.; Hadjantonakis, A.K.; Lang, R.A.; et al. Distinct functions for Wnt/beta-catenin in hair follicle stem cell proliferation and survival and interfollicular epidermal homeostasis. Cell Stem Cell 2013, 13, 720–733. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/beta-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef]
- Soma, T.; Fujiwara, S.; Shirakata, Y.; Hashimoto, K.; Kishimoto, J. Hair-inducing ability of human dermal papilla cells cultured under Wnt/beta-catenin signalling activation. Exp. Dermatol. 2012, 21, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Li, H.; Wu, Q.; Lee, H.J.; Ryu, J.H. A new labdane diterpenoid with anti-inflammatory activity from Thuja orientalis. J. Ethnopharmacol. 2013, 146, 760–767. [Google Scholar] [CrossRef] [PubMed]
- Miyamae, Y.; Han, J.; Sasaki, K.; Terakawa, M.; Isoda, H.; Shigemori, H. 3,4,5-tri-O-caffeoylquinic acid inhibits amyloid beta-mediated cellular toxicity on SH-SY5Y cells through the upregulation of PGAM1 and G3PDH. Cytotechnology 2011, 63, 191–200. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Miyamae, Y.; Kurisu, M.; Murakami, K.; Han, J.; Isoda, H.; Irie, K.; Shigemori, H. Protective effects of caffeoylquinic acids on the aggregation and neurotoxicity of the 42-residue amyloid beta-protein. Bioorg. Med. Chem. 2012, 20, 5844–5849. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kang, J.H.; Oh, G.H.; Park, M.H. Ishige sinicola extract stimulates osteoblast proliferation and differentiation via the bone morphogenetic protein 2/runt-related gene 2 signalling pathway. Z Nat. C J. Biosci. 2019, 74, 167–174. [Google Scholar] [CrossRef]
- Ali, I.; Manzoor, Z.; Koo, J.E.; Moon, S.R.; Byeon, S.H.; Yoo, E.S.; Kang, H.K.; Hyun, J.W.; Lee, N.H.; Koh, Y.S. Monoolein, isolated from Ishige sinicola, inhibits lipopolysaccharide-induced inflammatory response by attenuating mitogen-activated protein kinase and NF-kappaB pathways. Food Sci. Biotechnol. 2017, 26, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Ying, H.; Shi, J.; Zhang, S.; Pingcuo, G.; Wang, S.; Zhao, F.; Cui, Y.; Zeng, X. Transcriptomic and metabolomic profiling provide novel insights into fruit development and flesh coloration in Prunus mira Koehne, a special wild peach species. BMC Plant. Biol. 2019, 19, 463. [Google Scholar] [CrossRef]
- Peng, Z.X.; Wang, Y.; Gu, X.; Wen, Y.Y.; Yan, C. A platform for fast screening potential anti-breast cancer compounds in traditional Chinese medicines. Biomed. Chromatogr. 2013, 27, 1759–1766. [Google Scholar] [CrossRef]
- Lin, X.; Peng, Z.; Su, C. Potential anti-cancer activities and mechanisms of costunolide and dehydrocostuslactone. Int. J. Mol. Sci. 2015, 16, 10888–10906. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Choi, B.Y. Costunolide-A Bioactive Sesquiterpene Lactone with Diverse Therapeutic Potential. Int. J. Mol. Sci. 2019, 20, 2926. [Google Scholar] [CrossRef]
- Kim, S.H.; Lee, M.K.; Ahn, M.J.; Lee, K.Y.; Kim, Y.C.; Sung, S.H. Efficient Method for Extraction and Simultaneous Determination of Active Constituents in Cornus officinalis by Reflux Extraction and High Performance Liquid Chromatography with Diode Array Detection. J. Liq. Chromatogr. R T 2009, 32, 822–832. [Google Scholar] [CrossRef]
- Wang, W.; Xu, J.; Li, L.; Wang, P.; Ji, X.; Ai, H.; Zhang, L.; Li, L. Neuroprotective effect of morroniside on focal cerebral ischemia in rats. Brain Res. Bull. 2010, 83, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Huang, W.; Li, L.; Ai, H.; Sun, F.; Liu, C.; An, Y. Morroniside prevents peroxide-induced apoptosis by induction of endogenous glutathione in human neuroblastoma cells. Cell Mol. Neurobiol. 2008, 28, 293–305. [Google Scholar] [CrossRef]
- Wang, W.; Sun, F.; An, Y.; Ai, H.; Zhang, L.; Huang, W.; Li, L. Morroniside protects human neuroblastoma SH-SY5Y cells against hydrogen peroxide-induced cytotoxicity. Eur. J. Pharm. 2009, 613, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Zhao, H.; Li, N.; Su, L.; Xu, X.; Hong, Z. Protective effects of timosaponin BII on oxidative stress damage in PC12 cells based on metabolomics. Biomed. Chromatogr. 2018, 32, e4321. [Google Scholar] [CrossRef]
- Li, T.J.; Qiu, Y.; Yang, P.Y.; Rui, Y.C.; Chen, W.S. Timosaponin B-II improves memory and learning dysfunction induced by cerebral ischemia in rats. Neurosci. Lett. 2007, 421, 147–151. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, C.; Qi, Y.; Fang, L.; Luo, J.; Bi, K.; Jia, Y. Timosaponin B-II ameliorates scopolamine-induced cognition deficits by attenuating acetylcholinesterase activity and brain oxidative damage in mice. Metab. Brain Dis. 2016, 31, 1455–1461. [Google Scholar] [CrossRef] [PubMed]
- King, F.W.; Fong, S.; Griffin, C.; Shoemaker, M.; Staub, R.; Zhang, Y.L.; Cohen, I.; Shtivelman, E. Timosaponin AIII is preferentially cytotoxic to tumor cells through inhibition of mTOR and induction of ER stress. PLoS ONE 2009, 4, e7283. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Zhang, X.; Chen, Z.; Li, B.; Li, L. A Timosaponin B-II containing scalp care solution for improvement of scalp hydration, dandruff reduction, and hair loss prevention: A comparative study on healthy volunteers before and after application. J. Cosmet. Dermatol. 2021, 20, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Shim, K.S.; Lee, C.J.; Yim, N.H.; Ha, H.; Ma, J.Y. A water extract of Malva verticillata seeds suppresses osteoclastogenesis and bone resorption stimulated by RANK ligand. BMC Complement. Altern. Med. 2016, 16, 332. [Google Scholar] [CrossRef]
- Ko, J.H.; Castaneda, R.; Joo, S.W.; Kim, H.G.; Lee, Y.G.; Lee, Y.H.; Kang, T.H.; Baek, N.I. Glycerides isolated from the aerial parts of Malva verticillata cause immunomodulation effects via splenocyte function and NK anti-tumor activity. Food Sci. Biotechnol. 2018, 27, 1023–1030. [Google Scholar] [CrossRef]
- Hegazy, M.-E.F.; Mohamed, T.A.; ElShamy, A.I.; Abou-El-Hamd, H.M.; Mahalel, U.A.; Reda, E.H.; Shaheen, A.M.; Tawfik, W.A.; Shahat, A.A.; Shams, K.A. Microbial biotransformation as a tool for drug development based on natural products from mevalonic acid pathway: A review. J. Adv. Res. 2015, 6, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.; Lee, S.; Cha, J. Bacillus subtilis Fermentation of Malva verticillata Leaves Enhances Antioxidant Activity and Osteoblast Differentiation. Foods 2020, 9, 671. [Google Scholar] [CrossRef]
- Liang, Y.Y.; Wan, X.H.; Niu, F.J.; Xie, S.M.; Guo, H.; Yang, Y.Y.; Guo, L.Y.; Zhou, C.Z. Salvia plebeia R. Br.: An overview about its traditional uses, chemical constituents, pharmacology and modern applications. Biomed. Pharm. 2020, 121, 109589. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.H.; Zhao, L.; Xu, Y.K.; Bao, J.M.; Liu, X.; Zhang, J.S.; Li, W.; Ahmed, A.; Yin, S.; Tang, G.H. Anti-inflammatory sesquiterpenoids from the Traditional Chinese Medicine Salvia plebeia: Regulates pro-inflammatory mediators through inhibition of NF-kappaB and Erk1/2 signaling pathways in LPS-induced Raw264.7 cells. J. Ethnopharmacol. 2018, 210, 95–106. [Google Scholar] [CrossRef]
- Jang, H.H.; Cho, S.Y.; Kim, M.J.; Kim, J.B.; Lee, S.H.; Lee, M.Y.; Lee, Y.M. Anti-inflammatory effects of Salvia plebeia R. Br extract in vitro and in ovalbumin-induced mouse model. Biol. Res. 2016, 49, 41. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.; Li, W.; Ha, T.K.Q.; Lee, C.; Oh, W.K.; Shim, S.H. Anti-influenza effect of the major flavonoids from Salvia plebeia R.Br. via inhibition of influenza H1N1 virus neuraminidase. Nat. Prod. Res. 2018, 32, 1224–1228. [Google Scholar] [CrossRef]
- Hwang, E.K.; Gong, Y.G.; Park, C.S. Cultivation of a brown alga, Undariopsis peterseniana (Kjellman) Miyabe and Okamura, as a warm-temperature species by artificial seed production in Korea. J. Appl. Phycol. 2011, 23, 449–455. [Google Scholar] [CrossRef]
- Cho, M.; Yoon, S.J.; Kim, Y.-B. The nutritional composition and antioxidant activity from Undariopsis peterseniana. Ocean. Polar Res. 2013, 35, 273–280. [Google Scholar] [CrossRef]
- Lee, J.H.; Ko, J.Y.; Kim, E.A.; Hwang, E.K.; Park, C.S.; Lee, J.S.; Kim, C.Y.; Lee, H.S.; Kang, H.K.; Cha, S.H.; et al. Identification and large isolation of an anti-inflammatory compound from an edible brown seaweed, Undariopsis peterseniana, and evaluation on its anti-inflammatory effect in in vitro and in vivo zebrafish. J. Appl. Phycol. 2017, 29, 1587–1596. [Google Scholar] [CrossRef]
- Kang, M.C.; Ham, Y.M.; Heo, S.J.; Yoon, S.A.; Cho, S.H.; Kwon, S.H.; Jeong, M.S.; Jeon, Y.J.; Sanjeewa, K.; Yoon, W.J.; et al. Anti-inflammation effects of 8-oxo-9-octadecenoic acid isolated from Undaria peterseniana in lipopolysaccharide-stimulated macrophage cells. EXCLI J. 2018, 17, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Urysiak-Czubatka, I.; Kmiec, M.L.; Broniarczyk-Dyla, G. Assessment of the usefulness of dihydrotestosterone in the diagnostics of patients with androgenetic alopecia. Postepy Dermatol. Alergol. 2014, 31, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Adil, A.; Godwin, M. The effectiveness of treatments for androgenetic alopecia: A systematic review and meta-analysis. J. Am. Acad. Dermatol. 2017, 77, 136–141.e135. [Google Scholar] [CrossRef]
- Dinh, Q.Q.; Sinclair, R. Female pattern hair loss: Current treatment concepts. Clin. Interv. Aging 2007, 2, 189. [Google Scholar] [PubMed]
- Said, M.A.; Mehta, A. The Impact of 5α-Reductase inhibitor use for male pattern hair loss on men’s health. Curr. Urol. Rep. 2018, 19, 65. [Google Scholar] [CrossRef]
- Yu, W.; Zha, W.; Guo, S.; Cheng, H.; Wu, J.; Liu, C. Flos Puerariae extract prevents myocardial apoptosis via attenuation oxidative stress in streptozotocin-induced diabetic mice. PLoS ONE 2014, 9, e98044. [Google Scholar] [CrossRef] [PubMed]
- Lertpatipanpong, P.; Janpaijit, S.; Park, E.Y.; Kim, C.T.; Baek, S.J. Potential Anti-Diabetic Activity of Pueraria lobata Flower (Flos Puerariae) Extracts. Molecules 2020, 25, 3970. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.O.; Han, M.J.; Park, S.H.; Kim, D.H. Protective effects of kakkalide from Flos puerariae on ethanol-induced lethality and hepatic injury are dependent on its biotransformation by human intestinal microflora. J. Pharm. Sci. 2003, 93, 331–336. [Google Scholar] [CrossRef]
- Zhuang, B.; Bi, Z.M.; Wang, Z.Y.; Duan, L.; Lai, C.J.; Liu, E.H. Chemical profiling and quantitation of bioactive compounds in Platycladi Cacumen by UPLC-Q-TOF-MS/MS and UPLC-DAD. J. Pharm. Biomed. Anal. 2018, 154, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, S.; Qu, F.; Su, G.; Zhao, Y. In vivo and in vitro evaluation of hair growth potential of Cacumen Platycladi, and GC-MS analysis of the active constituents of volatile oil. J Ethnopharmacol. 2019, 238, 111835. [Google Scholar] [CrossRef]
- Pan, J.; Zheng, W.; Pang, X.; Zhang, J.; Chen, X.; Yuan, M.; Yu, K.; Guo, B.; Ma, B. Comprehensive Investigation on Ginsenosides in Different Parts of a Garden-Cultivated Ginseng Root and Rhizome. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Wang, H.P.; Zhang, Y.B.; Yang, X.W.; Zhao, D.Q.; Wang, Y.P. Rapid characterization of ginsenosides in the roots and rhizomes of Panax ginseng by UPLC-DAD-QTOF-MS/MS and simultaneous determination of 19 ginsenosides by HPLC-ESI-MS. J. Ginseng Res. 2016, 40, 382–394. [Google Scholar] [CrossRef]
- Kim, S.; Oh, M.H.; Kim, B.S.; Kim, W.I.; Cho, H.S.; Park, B.Y.; Park, C.; Shin, G.W.; Kwon, J. Upregulation of heme oxygenase-1 by ginsenoside Ro attenuates lipopolysaccharide-induced inflammation in macrophage cells. J. Ginseng Res. 2015, 39, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Samukawa, K.; Kubo, M. Anti-inflammatory activity of ginsenoside ro. Planta Med. 1990, 56, 19–23. [Google Scholar] [CrossRef]
- Kang, H.J.; Oh, Y.; Lee, S.; Ryu, I.W.; Kim, K.; Lim, C.J. Antioxidative properties of ginsenoside Ro against UV-B-induced oxidative stress in human dermal fibroblasts. Biosci. Biotechnol. Biochem. 2015, 79, 2018–2021. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.S.; Li, W.; Zhuang, T.X.; Yu, J.J.; Sun, S.; Ju, Z.C.; Wang, Z.T.; Ding, L.L.; Yang, L. Ginsenoside Ro Ameliorates High-Fat Diet-Induced Obesity and Insulin Resistance in Mice via Activation of the G Protein-Coupled Bile Acid Receptor 5 Pathway. J. Pharm. Exp. Ther. 2021, 377, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dong, R.; Wang, Y.; Wang, L.; Zhou, T.; Jia, D.; Meng, Z. The anti-breast cancer property of physcion via oxidative stress-mediated mitochondrial apoptosis and immune response. Pharm. Biol. 2021, 59, 303–310. [Google Scholar] [CrossRef]
- Trybus, W.; Krol, T.; Trybus, E.; Stachurska, A. Physcion Induces Potential Anticancer Effects in Cervical Cancer Cells. Cells 2021, 10, 2029. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wu, N.; Zu, Y.; Fu, Y. Antioxidative activity of Rosmarinus officinalis L. essential oil compared to its main components. Food Chem. 2008, 108, 1019–1022. [Google Scholar] [CrossRef] [PubMed]
- Oluwatuyi, M.; Kaatz, G.W.; Gibbons, S. Antibacterial and resistance modifying activity of Rosmarinus officinalis. Phytochemistry 2004, 65, 3249–3254. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Sánchez, A.; Barrajón-Catalán, E.; Ruiz-Torres, V.; Agulló-Chazarra, L.; Herranz-López, M.; Valdés, A.; Cifuentes, A.; Micol, V. Rosemary (Rosmarinus officinalis) extract causes ROS-induced necrotic cell death and inhibits tumor growth in vivo. Sci. Rep. 2019, 9, 808. [Google Scholar] [CrossRef] [PubMed]
- Sharaf, M.; El-Ansari, M.A.; Saleh, N.A. New flavonoids from Avicennia marina. Fitoterapia 2000, 71, 274–277. [Google Scholar] [CrossRef]
- Heo, S.-J.; Park, E.-J.; Lee, K.-W.; Jeon, Y.-J. Antioxidant activities of enzymatic extracts from brown seaweeds. Bioresour. Technol. 2005, 96, 1613–1623. [Google Scholar] [CrossRef]
- Wijesinghe, W.; Jeon, Y.-J. Exploiting biological activities of brown seaweed Ecklonia cava for potential industrial applications: A review. Int. J. Food Sci. Nutr. 2012, 63, 225–235. [Google Scholar] [CrossRef]
- Kang, J.-I.; Kim, S.-C.; Kim, M.-K.; Boo, H.-J.; Jeon, Y.-J.; Koh, Y.-S.; Yoo, E.-S.; Kang, S.-M.; Kang, H.-K. Effect of Dieckol, a component of Ecklonia cava, on the promotion of hair growth. Int. J. Mol. Sci. 2012, 13, 6407–6423. [Google Scholar] [CrossRef]
- Abe, Y.; Tanaka, N. Roles of the Hedgehog Signaling Pathway in Epidermal and Hair Follicle Development, Homeostasis, and Cancer. J. Dev. Biol. 2017, 5. [Google Scholar] [CrossRef]
- Bitgood, M.J.; McMahon, A.P. Hedgehog and Bmp genes are coexpressed at many diverse sites of cell-cell interaction in the mouse embryo. Dev. Biol. 1995, 172, 126–138. [Google Scholar] [CrossRef] [PubMed]
- St-Jacques, B.; Dassule, H.R.; Karavanova, I.; Botchkarev, V.A.; Li, J.; Danielian, P.S.; McMahon, J.A.; Lewis, P.M.; Paus, R.; McMahon, A.P. Sonic hedgehog signaling is essential for hair development. Curr. Biol. 1998, 8, 1058–1068. [Google Scholar] [CrossRef]
- Chakrawarti, L.; Agrawal, R.; Dang, S.; Gupta, S.; Gabrani, R. Therapeutic effects of EGCG: A patent review. Expert Opin. Pat. 2016, 26, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Sharma, V.; Chauhan, N.S.; Thakur, M.; Dixit, V.K. Hair growth: Focus on herbal therapeutic agent. Curr. Drug Discov. Technol. 2015, 12, 21–42. [Google Scholar] [CrossRef] [PubMed]
- Saraste, A.; Pulkki, K. Morphologic and biochemical hallmarks of apoptosis. Cardiovasc Res. 2000, 45, 528–537. [Google Scholar] [CrossRef]
- Fulda, S.; Debatin, K.M. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene 2006, 25, 4798–4811. [Google Scholar] [CrossRef] [PubMed]
- Knudson, C.M.; Korsmeyer, S.J. Bcl-2 and Bax function independently to regulate cell death. Nat. Genet. 1997, 16, 358–363. [Google Scholar] [CrossRef]
- Stenn, K.S.; Paus, R. Controls of hair follicle cycling. Physiol. Rev. 2001, 81, 449–494. [Google Scholar] [CrossRef]
- Seiberg, M.; Marthinuss, J.; Stenn, K.S. Changes in expression of apoptosis-associated genes in skin mark early catagen. J. Investig. Dermatol. 1995, 104, 78–82. [Google Scholar] [CrossRef][Green Version]
- Giacomini, F.; Starace, M.; Tosti, A. Short anagen syndrome. Pediatric Dermatol. 2011, 28, 133–134. [Google Scholar] [CrossRef]
- Murugusundram, S. Serenoa repens: Does it have any role in the management of androgenetic alopecia? J. Cutan. Aesthetic Surg. 2009, 2, 31. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lyle, S.; Liu, Y.; Solky, B.; Cotsarelis, G. Differential expression of cyclin D1 in the human hair follicle. Am. J. Pathol. 2003, 163, 969–978. [Google Scholar] [CrossRef]
- Purba, T.S.; Brunken, L.; Hawkshaw, N.J.; Peake, M.; Hardman, J.; Paus, R. A primer for studying cell cycle dynamics of the human hair follicle. Exp. Dermatol. 2016, 25, 663–668. [Google Scholar] [CrossRef]
- Whittaker, S.R.; Mallinger, A.; Workman, P.; Clarke, P.A. Inhibitors of cyclin-dependent kinases as cancer therapeutics. Pharm. Ther. 2017, 173, 83–105. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Botanical Name | Parts Used | Bioactive Components | Type of Extract | Ref |
|---|---|---|---|---|
| Citrus limon L., Fragaria ananassa L., Secale cereale L. | - | Sinapic acid | - | [16] |
| Epimedium spp. | Leaves | Icariin | - | [17] |
| Geranium sibiricum L. | Whole plant | Corilagin, gallic acid | Methanolic extract | [18] |
| Olea europaea L. | Unprocessed olive fruit and leaves | Oleuropein | - | [19] |
| Camellia sinensis, Coffea ara bica | - | Caffeine | - | [20] |
| Carthamus tinctorius L. | Floret | Hydroxysafflor yellow A | Ethanolic extract | [21] |
| Panax ginseng Mayer | Whole plant | Linoleic acid, β-sitosterol, bicyclo(10.1.0)tridec-1-ene | Supercritical fluid extraction system | [22] |
| Hottuynia cordata Thunb. | - | Quercitrin | - | [23] |
| Sophora flavescens Aiton | Roots | L-maackiain, medicarpin | Methanolic extract | [24] |
| Illicium anisatum L., Illicium verum Hook. f. | - | Shikimic acid | - | [25] |
| Hordeum vulgare L. var. distichon Alefeld | Seed | Procyanidin B-3 | Aceton extract | [26] |
| Panax ginseng Mayer | - | Ginsenoside Rb1 | - | [27] |
| Nelumbinis semen | Whole plant | Anthraquinone, flavonoids, tannin, saponins | Ethanolic extract | [28] |
| Chamaecyparis obtusa | - | α-Terpinyl acetate, sabinene, isobornyl acetate, limonene | Oil | [29] |
| Polygonum multiflorum | Roots | 2,3,5,4′-Tetrahydroxystilbene2-O-β-D-glucoside, emodin | Ethanolic extract | [30] |
| Alnus sibirica Fisch. ex Turcz | Whole plant | Oregonin | Ethanolic extract | [31] |
| Malva verticillate | Seed | Myristoleic acid | Ethanolic extract | [32] |
| Malva verticillate | Seed | Linoleic acid | Ethanolic extract | [33] |
| Magnolia officinalis | - | Liposomal honokiol | - | [34] |
| Angelica gigas Nakai | Roots | Decursin | Ethanolic extract | [35] |
| Caesalpinia sappan L. | - | 3-Deoxysappanchalcone | - | [36] |
| Broussonetia papyrifera | Whole plant | 7-hydroxycoumarin, protocatechuate acid, ferulic acid, protocatechuic acid and epicatechin | Ethanolic extract | [37] |
| Panax ginseng Mayer | Whole plant | Linoleic acid, β-sitosterol | Supercritical fluid extraction system | [38] |
| Thuja orientalis | Leaves | Kaempferol, isoquercetin | Hot water extract | [39] |
| Ipomoea batatas L. | - | 3,4,5-tri-O-caffeoylquinic acid | - | [40] |
| Ishige sinicola | Whole plant | Octaphlorethol A | Ethanolic extract | [41] |
| Prunus mira Koehne | Nut oil | α-tocopherol, vitamin E β-sitosterol, linoleic acid, oleic acid | Pressing the seeds | [42] |
| Saussurea lappa Clarke | - | Costunolide | - | [43] |
| Cornus officinalis | - | Morroniside | - | [44] |
| Anemarrhena asphodeloides | - | Timosaponin BII | - | [45] |
| Salvia plebeia R. Br. | Whole plant | Flavonoids, monoterpenoids, sesquiterpenoids, diterpenoids, triterpenes, phenolic acids | Methanolic extract | [46] |
| Undariopsis peterseniana | Whole plant | Apo-9′-fucoxanthinone | Ethanolic extract | [47] |
| Pueraria thomsonii | Whole plant | Soyasaponin I, kaikasaponin III | Ethanolic extract | [48] |
| Platycladus orientalis (L.) Franco | Leaves | Myricitrin, isoquercitrin, quercitrin, myricetin, afzelin, quercetin, kaempferol, amentoflavone, hinokiflavone | Ethanolic extract, aqueous extract | [49] |
| Panax ginseng | Rhizome | Ginsenoside Ro | Ethanolic extract | [50] |
| Polygonum multiforum Thunb. | Leaves | Physcion | - | [51] |
| Rosmarinus officinalis | Leaves | 12-methoxycarnosic acid | Ethanolic extract | [52] |
| Avicennia marina | Heartwood | Avicequinone C | Methanolic extract | [53] |
| Ecklonia cava | Whole plant | Dieckol | Enzymatic hydrolysis reaction | [54] |
| Camellia sinensis L. Ktze. | - | Epigallocatechin-3-Gallate | - | [55] |
| Panax ginseng Mayer | Roots | Ginsenosides | Ethanolic extract | [56] |
| Sabal serrulatum | - | Fatty acids, phytosterols | - | [57] |
| Natural Products | Experimental Model | Treatment | Effects | Ref |
|---|---|---|---|---|
| Sinapic acid | Human hair-follicle-derived papilla cells | 10, 50, and 100 μM | Induction of cell proliferation and cell cycle progression; activation of Akt and GSK-3β/β-catenin signaling; increased expression of VEGF and IGF-1 | [16] |
| Icariin | Cultured vibrissae follicles | 10 and 20 μM | Induction of hair shaft elongation and prolonged anagen phase; increase of IGF-1 production and expression | [17] |
| Cultured dermal papilla cells | 10 and 20 μM | |||
| C57BL/6 mice | 0.01 | |||
| Geranium sibiricum extract | Human dermal papilla cells | 9.8–156.3 ppm | Induction of cell proliferation and migration; induced expression of Ki-67 protein, HGF, and VEGF in vitro; reduced number of mast cells and the expression of TGF-β1 in mouse skin | [18] |
| C57BL/6 mice | 1000 ppm | |||
| Oleuropein | Human follicle dermal papilla cells | 10, 20, and 50 μM | Induction of cell proliferation; increase of LEF1 and cyclin D1 mRNA expression and β-catenin protein expression in vitro; induction of anagenic hair growth and Wnt/β-catenin pathway in vivo; upregulation of IGF-1, KGF, HGF, and VEGF gene expression in mice | [19] |
| C57BL/6 mice | 0.4 mg/mouse/day | |||
| Caffeine | Cultured hair follicles Human hair-follicle-derived outer root sheath keratinocytes (ORSKs) | 0.0005% 0.00001, 0.0001, 0.001% | Increase of hair shaft elongation, anagen duration; increase of hair matrix keratinocyte proliferation and IGF-1 expression in hair follicles; increase of cell proliferation and IGF-1 expression in RSKs; inhibited apoptosis/necrosis and TGF-β2 protein secretion in RSKs | [20] |
| Carthamus Tinctorius Floret extract | Human keratinocytes (HaCaT) | 0.005–1.250 mg/mL | Induction of cell proliferation in dermal papilla cells and HaCaT increase of VEGF, KGF; de crease of TGF-β1; increase of length of cultured hair follicles and stimulated the growth of hair in mice | [21] |
| Human hair follicle-derived papilla cells | 0.005-1.250 mg/mL | |||
| Cultured hair follicles | 50, 100 and 200 μg/mL | |||
| Red Ginseng Oil | C57BL/6 mice | 10% | Increase of hair growth; upregulated β-catenin, Lef-1, Sonic hedgehog, Smoothened, Gli-1, Cyclin D1, and Cyclin E expression; reduced the protein level of TGF-β; enhanced the expression of Bcl-2 | [22] |
| Quercitrin | Human dermal papilla cells Cultured hair follicles | 0.1, 1, 10, 100 nM and 1 μM 5 and 10 μM | Enhanced the cell viability and cellular energy metabolism; increase of expression of Bcl-2 and Ki67; upregulation of bFGF, KGF, PDGF-AA and VEGF; stimulated hair shaft growth in cultured hair follicles | [23] |
| Sophora flavescens extract | Cultured dermal papilla cells | 10−6, 10−5 and 10−4% | Induction of hair growth in vivo; increase of IGF-1 and KGF in vitro; decrease of 5a-reductase activity in vivo | [24] |
| Sprague-Dawley rats | 0.001, 0.01 and 0.01% | |||
| C57BL/6 mice | 1% | |||
| Shikimic acid | Human follicle dermal papilla cells Human outer root sheath keratinocytes C57BL/6 mice Cultured hair follicles | 0.1, 1, 10, 100 μM and 1 mM 1 and 10 μM 10 and 100 mM 1 and 10 μM | Induction of hair growth in vivo; increase of Cell proliferation in hDPCs and hORSCs; en hanced hair shaft elongation in cultured hair follicles; increased c-myc, HGF, KGF, VEGF, p38 MAPK and CREB | [25] |
| Procyanidin B-3 | Cultured hair epithelial cells from C3H/HeNCrj mice | 0.1–100 μg/mL | Increased hair-growing activity in vitro and anagen-inducing activity in vivo; potential inhibitory effect of TGF-β1 | [26] |
| C3H mice | 200 μL/day/mouse | |||
| Ginsenoside Rb1 | Cultured mink hair follicles | 5 and 10 μg/mL | Increase of the growth of hair follicles; upregulated the expression levels of VEGF-A and VEGF-R2, while attenuated the TGF-β1 expression; activation of PI3K/AKT/GSK-3β signaling pathway in hair follicles and DPCs. | [27] |
| Cultured dermal papilla cells | 10 μg/mL | |||
| Nelumbinis Semen extract | Human follicle dermal papilla cells | 15.63–125 ppm | Enhanced cell proliferation and migration; high mRNA expression of VEGF and IGF-1; low TGF- β1 mRNA expression | [28] |
| C57BL/6 mice | 1000 ppm | |||
| Chamaecyparisobtusa oil | C57BL/6 mice | 3% | Increase of ALP and γ-GT activities in the skin tissue; increase of IGF-1 mRNA expression; increase of VEGF and decrease of EGF expression in the skin tissue; increase of SCF expression | [29] |
| Polygonum multiflorum extract | Human follicle dermal papilla cells Cultured hair follicles | 10 and 100 μg/mL 2, 20, and 50 μg/mL | Increased cell viability and mitochondrial activity; increase of Bcl-2 and decrease of BAD and DKK-1; increase of IGFBP2, PDGF and VEGF; prolonged the anagen of human hair follicles | [30] |
| Alnus sibirica Fisch. ex Turcz | Human follicle dermal papilla cells | 22, 66 and 200 μg/mL | Inhibition of apoptosis; increased IGF-1 and decreased TGF-β1 expression; decreased DHT production | [31] |
| Malva verticillata seed extracts | Human follicle dermal papilla cells | 10–100 μg/mL | Increased Wnt reporter activity; increased β-catenin level; increased IGF-1, KGF, VEGF and HGF | [32] |
| Linoleic acid in Malva verticillate seed | Human follicle dermal papilla cells | 3, 10 and 30 μg/mL | Activated Wnt/β-catenin signaling; induced cell growth by increasing the expression of cyclin D1 and CDK2; increased VEGF, IGF-1, HGF and KGF; inhibited DKK-1 | [33] |
| Liposomal honokiol | C57BL/6 mice | 20 mg/mL | Promoted hair regrowth; accelerated the hair growth cycle by up regulating the Wnt3a/β- catenin signaling pathway; inhibited the TGF-β1/p-smad2 signaling pathway during the anagen stage | [34] |
| Decursin, Angelica Gigas Nakai root extract | C57BL/6 mice | 2% | Induction of hair growth; decrease of TNF-and IL-1β; increase of IL-4 and IL-13 | [35] |
| 3-Deoxysappanchalcone | Human follicle dermal papilla cells C57BL/6 mice | 0.1–10 µM | Increased cell proliferation; increase of β-catenin and Tcf; increase of IL-6-induced phosphorylation and subsequent transactivation of STAT3, thereby increasing the expression of Cdk4, FGF and VEGF; promoted the anagen phase of hair growth in C57BL/6 mice | [36] |
| 3 mM | ||||
| Broussonetia papyrifera | Human follicle dermal papilla cells NIH3T3 cells Clinical Study | 1.25–40 μg/mL 1–40 μg/mL Korean males and females | Promoted cell proliferation; enhanced TCF/LEF-luciferase activity and increased the level of β-catenin protein; inhibited IL4-induced STAT6 phosphorylation; increased hair count after using the hair tonic for 12 weeks | [37] |
| Red Ginseng Oil | C57BL/6 mice SKH-1 hairless mice | 50% 1% | Induction of hair growth; upregulated expression of β-catenin, phospho-GSK3β, Lef-1, Gli-1, Smoothened, Cyclin D1, Cyclin E, IGF-1 and VEGF; protective effect against UVC-induced skin damage in SKH-1 hairless mice by inhibiting inflammation and apoptosis | [38] |
| Thuja orientalis | C57BL/6 mice | 5.05 mg/cm2/day | Induction of hair growth including hair number and size of hair follicles; induction of β-catenin and Shh protein expression | [39] |
| 3,4,5-tri-O-caffeoylquinic acid | Human follicle dermal papilla cells C3H mice | 5, 10, 15 and 25 μM 1% | Increase of β-catenin in vitro and in vivo; upregulation in hair growth-associated genes using microarray | [40] |
| Ishige sinicola | Cultured rat vibrissa follicles C57BL/6 mice Sprague-Dawley rats | 1, 10, and 100 μg/mL 0.1, 1, and 10 μg/mL 0.1, 1, and 10 μg/mL | Induction of anagen progression of the hair shaft; inhibition of 5α-reductase activity; in crease of cell proliferation; increase of phos pho-GSK3β, β-catenin, Cyclin E, and CDK2, and decrease of p27kip1 | [41] |
| Prunus mira Koehne | C57BL/6 mice | 15.06, 30.13, and 60.26 mg/cm2/day | Increase of hair growth; increase of Wnt 10b, β-catenin, and GSK-3β | [42] |
| KM mice | 0.47–60.26 mg/cm2/day | |||
| Costunolide | Human follicle dermal papilla cells C57BL/6 mice | 0.1, 0.3, 1, and 3 μM 0.3 μM | Increase of cell proliferation in vitro; inhibited the 5a-reductase activity in hHFDPCs; in creased the level of β-catenin and Gli1 and de creased TGF-β1; increase of hair length in vivo | [43] |
| Morroniside | Cultured outer root sheath cells C57BL/6 mice | 1 and 10 µM 100 µM | Enhanced outer root sheath cell proliferation and migration in vitro; upregulation of Wnt10b, β-catenin, and LEF1; accelerated the onset of anagen and delayed hair follicle cat agen | [44] |
| Timosaponin BII | C57BL/6 mice | 0.5% | Induced hair growth; increased expression of β-catenin and Wnt10b | [45] |
| Salvia plebeian extract | Human follicle dermal papilla cells C57BL/6 mice | 7.8, 15.6, and 31.3 μg/mL 1,000 μg/mL | Increased cell proliferation; increased HGF and decreased TGF-β1 and SMAD2/3; inhibi tion of apoptosis by increasing the Bcl-2/Bax ratio; enhanced hair growth in mice | [46] |
| Undariopsis peterseniana | C57BL/6 mice Cultured rat vibrissa follicles Sprague-Dawley rats NIH3T3 fibroblasts Dermal papilla cells | 0.1, 1, and 10 μg/mL 1, 10, and 100 μg/mL 0.1, 1, 10, and 100 μg/mL 0.1, 1, 10, and 100 μg/mL 0.1, 1, and 10 μg/mL | Increase of the hair-fiber lengths and anagen initiation in vivo; decreased 5α-reductase ac tivity and increased cell proliferation in vitro; increased the levels of Cyclin D1, phos pho(ser780)-pRB, Cyclin E, phospho-CDK2, and CDK2; increase of phosphorylation of ERK and the levels of Wnt/β-catenin signaling proteins | [47] |
| Puerariae Flos | Wistar/ST rats C57BL/6NCrSlc mice C3H/He mice | 50, 200, and 500 μg/mL 2 and 5 mg/mouse/day 2 and 5 mg/mouse/day | Increase of hair re-growth effect in testos terone-treated C57BL/6NCrSlc and C3H/He mice; inhibitory activity of against testos terone 5a-reductase | [48] |
| Cacumen platycladi | Sprague-Dawley rats C57BL/6NCrSlc(C57) mice | 0.02–2.5 μmol/L 2 and 5 mg/mouse/day | Increase of hair growth; increase of Wnt 10b, β-catenin, and GSK-3β | [49] |
| Ginseng rhizome, Ginsenoside Ro | Wistar rats C57BL/6 mice | Ginseng rhizome (200, 500, and 1,000 μg/mL) Ginseng rhizomes (2 mg/mouse), and ginsenoside Ro (0.2 mg/mouse) | Induced hair re-growth in vivo; inhibitory effects against 5αR | [50] |
| Physcion | Sprague-Dawley ratsC57BL/6 mice | 12.5–100 µM5 mg/mouse/day | Inhibited the 5a-reductase activity; hair -growth-promoting activity | [51] |
| Rosmarinus officinalis leaf extract | Wistar rats C3H/He mice C57BL/6 mice LNCaP cells | 50, 200, and 500 μg/mL 2 mg/mouse/day 2 mg/mouse/day 1–5 µM | Improved hair regrowth in C57BL/6NCrSlc mice; increased hair growth in C3H/He mice that had their dorsal areas shaved; decreased 5a-reductase activity; inhibited androgen-de pendent proliferation of LNCaP cells | [52] |
| Avicequinone C | Human hair dermal papilla cells | 5 and 10 µg/mL | Decreased 5a-reductase activity | [53] |
| Ecklonia cava enzymatic extract, dieckol | Cultured rat vibrissa follicles Sprague-Dawley rats C57BL/6 miceRat vibrissa, immortalized dermal papilla cell, NIH3T3 cells | 0.01–10 µg/mL 0.5% 0.5% 0.001–10 µg/mL 0.05 and 10 µg/mL | Increased hair-fiber length and anagen pro gression of the hair-shaf after E. cava enzy matic extract; decreased 5a-reductase activity in the presence of dieckol | [54] |
| Epigallocatechin-3-Gallate | Cultured mink hair follicles Cultured dermal papilla cells Cultured outer root sheath cells | 0.1–5 µM 0.25–4 µM 0.25–4 µM | Promoted hair follicle growth in DPCs and ORSCs; activated Shh and Akt signaling; in creased the expression of cyclin D1 and cyclin E1 | [55] |
| Panax ginseng extract | Cultured outer root sheath keratinocytes | 20 ppm 20 ppm | Increased proliferation and inhibited apopto sis in ORS keratinocytes; abrogated DKK-1-in duced growth inhibition of cultured HFs ex vivo | [56] |
| Serenoa repens extracts | Human keratinocyte cells C57BL/6 mice | 1, 5, 25, and 100 μg/ mL 50% | Increased cell viabilities; stimulated hair folli cle growth; decreased inflammatory response; decreased TGF-β2 and cleaved caspase-3 ex pression of hair loss mouse; inhibited apopto sis | [57] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.; Lee, J. Modulation of Hair Growth Promoting Effect by Natural Products. Pharmaceutics 2021, 13, 2163. https://doi.org/10.3390/pharmaceutics13122163
Park S, Lee J. Modulation of Hair Growth Promoting Effect by Natural Products. Pharmaceutics. 2021; 13(12):2163. https://doi.org/10.3390/pharmaceutics13122163
Chicago/Turabian StylePark, Seyeon, and Joomin Lee. 2021. "Modulation of Hair Growth Promoting Effect by Natural Products" Pharmaceutics 13, no. 12: 2163. https://doi.org/10.3390/pharmaceutics13122163
APA StylePark, S., & Lee, J. (2021). Modulation of Hair Growth Promoting Effect by Natural Products. Pharmaceutics, 13(12), 2163. https://doi.org/10.3390/pharmaceutics13122163