
Investigating Structural Property Relationships to Enable Repurposing of Pharmaceuticals as Zinc Ionophores

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
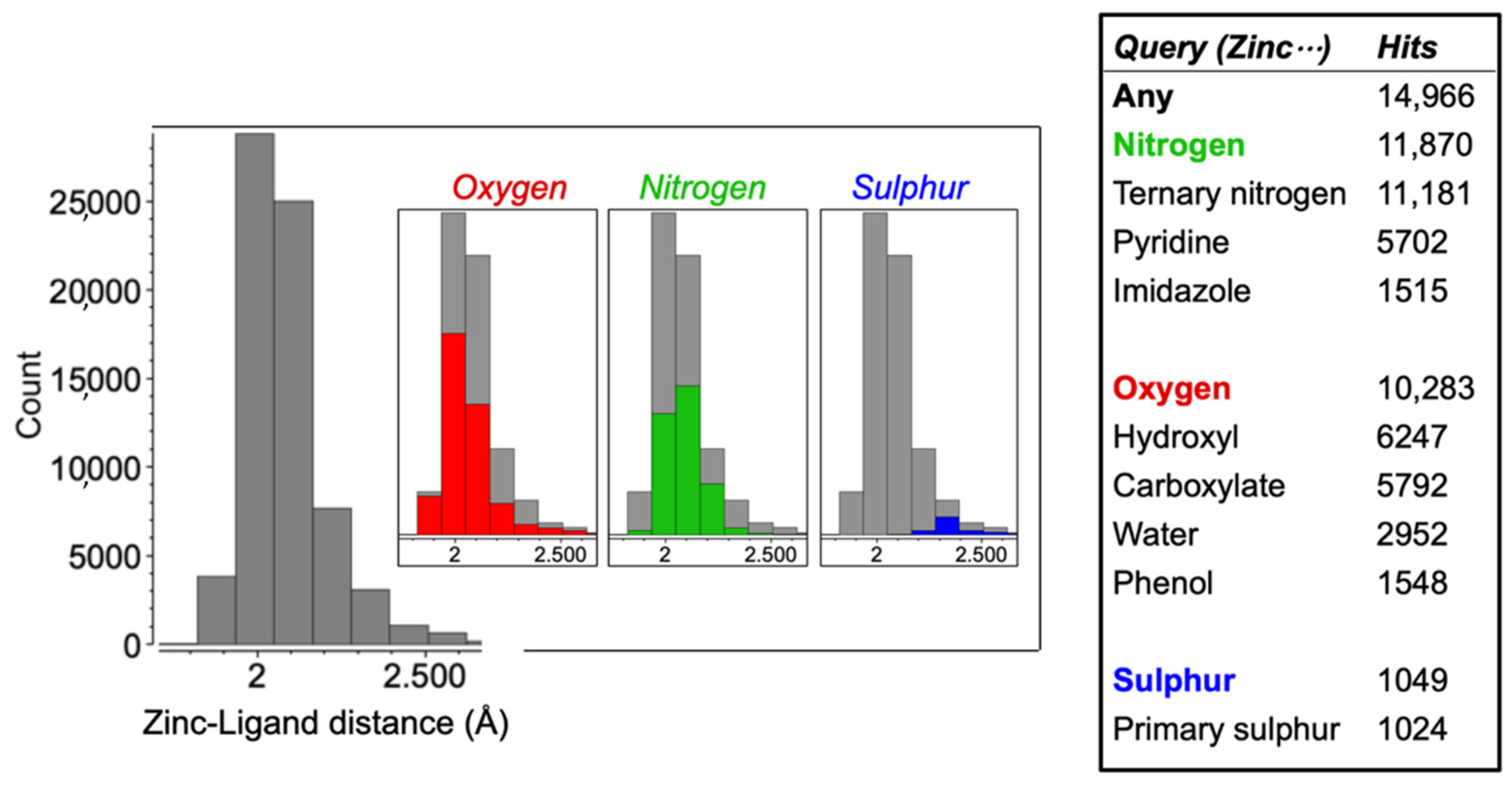
2.1. Cambridge Structural Database
2.2. Synthesis of FluoZin-3 Loaded Liposomes
2.3. Liposomal Penetration Assay
2.4. Cell Culture Conditions
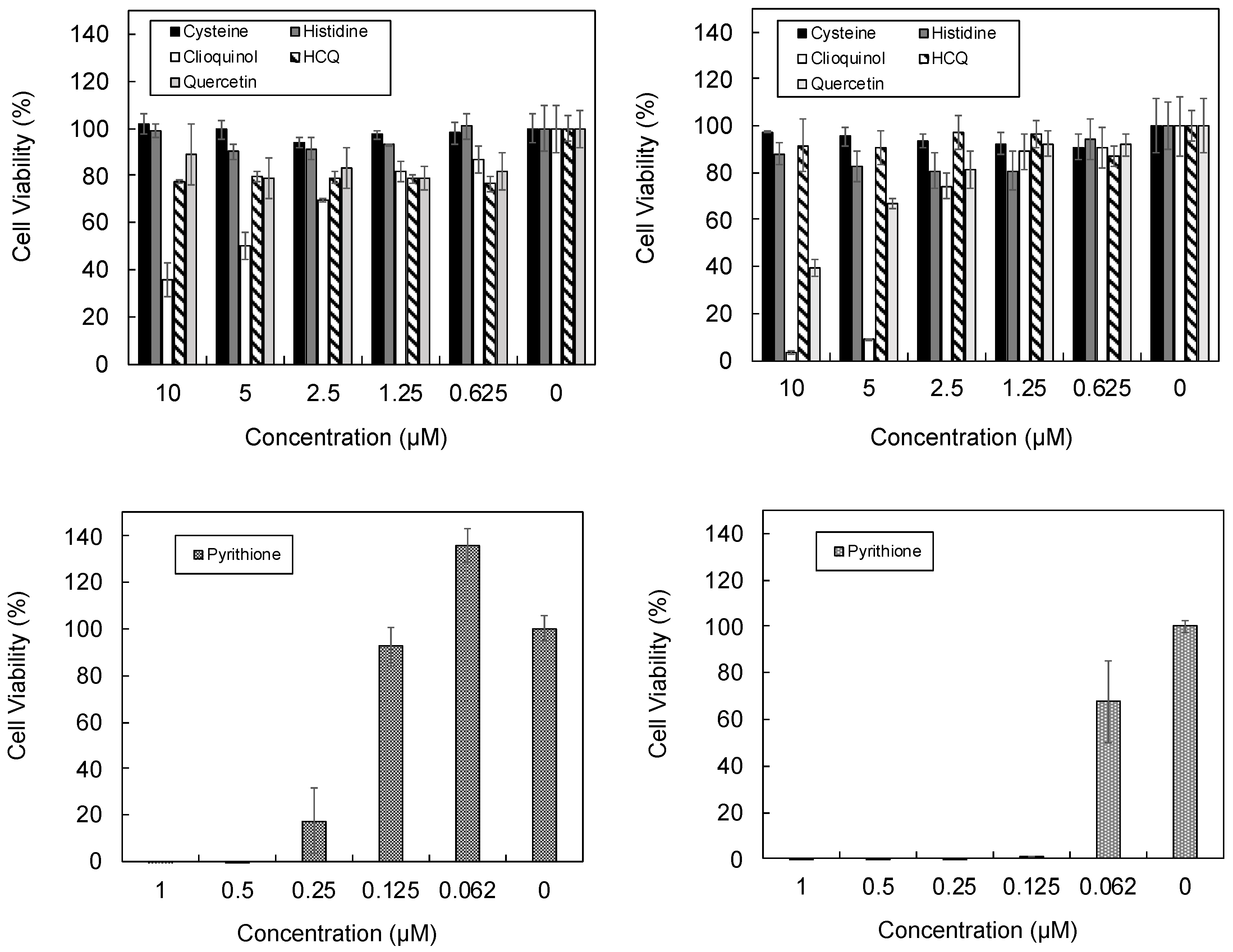
2.5. Cell Toxicity Assay
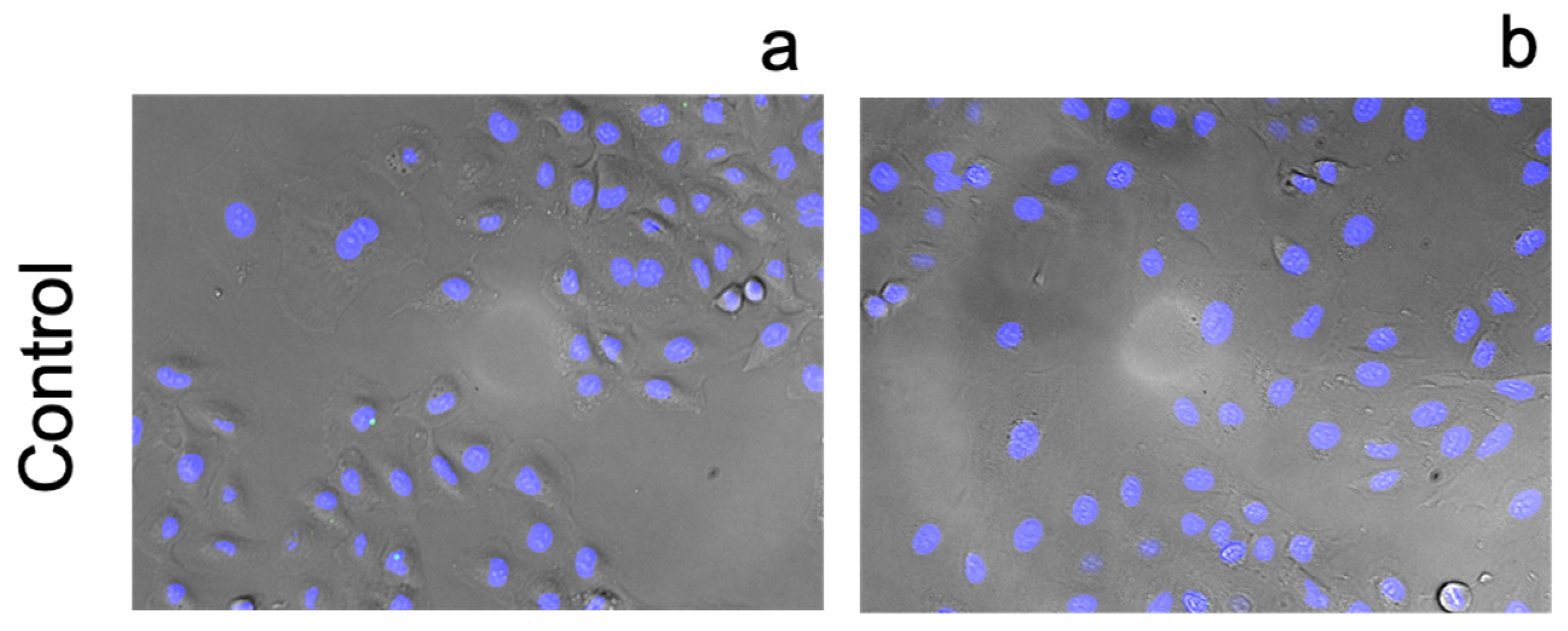
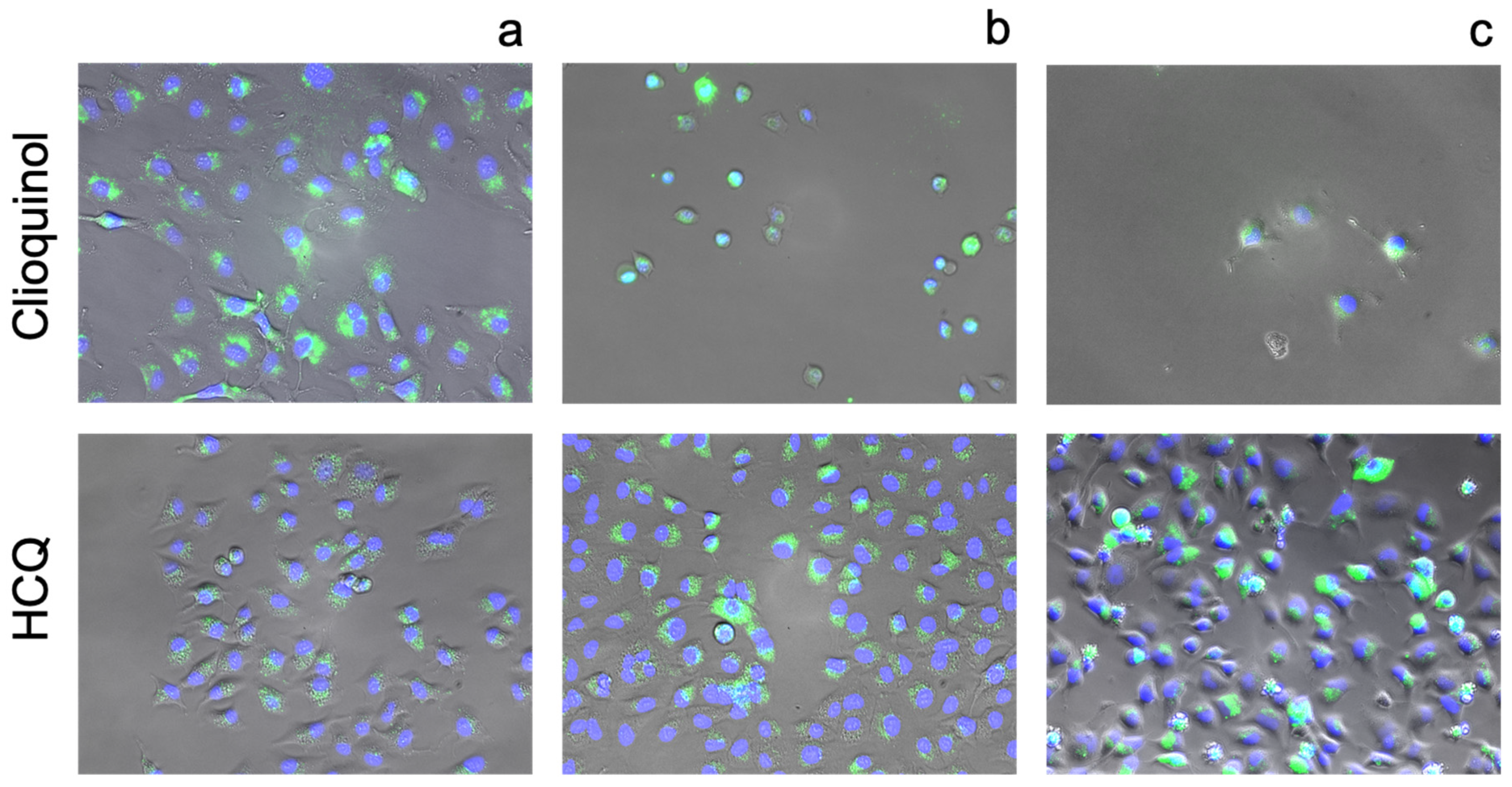
2.6. Fluorescence Imaging
3. Results
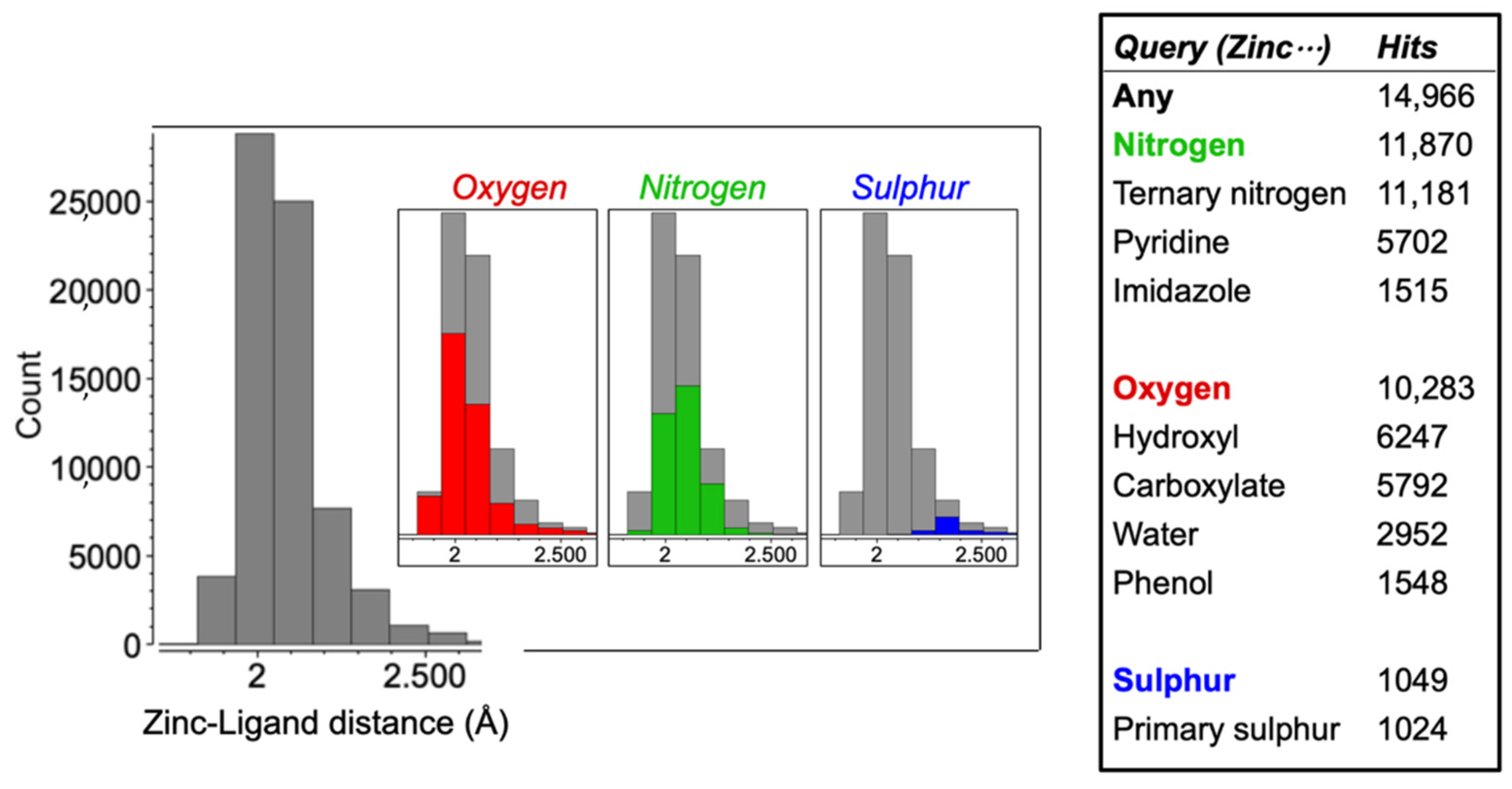
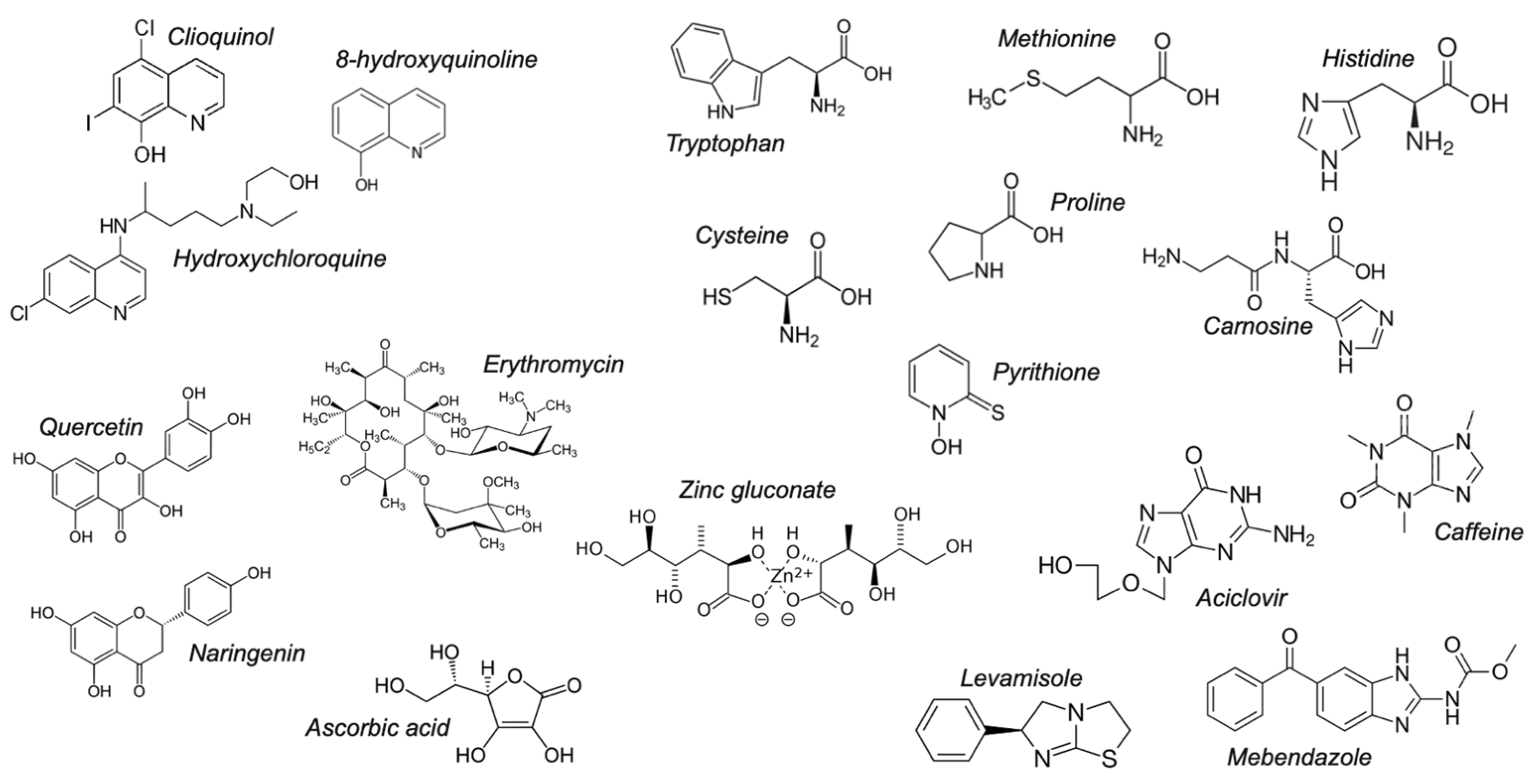
3.1. Identification of Zinc Chelators with CSD Analysis
3.2. Liposomal Penetration Behaviour of Selected Compounds
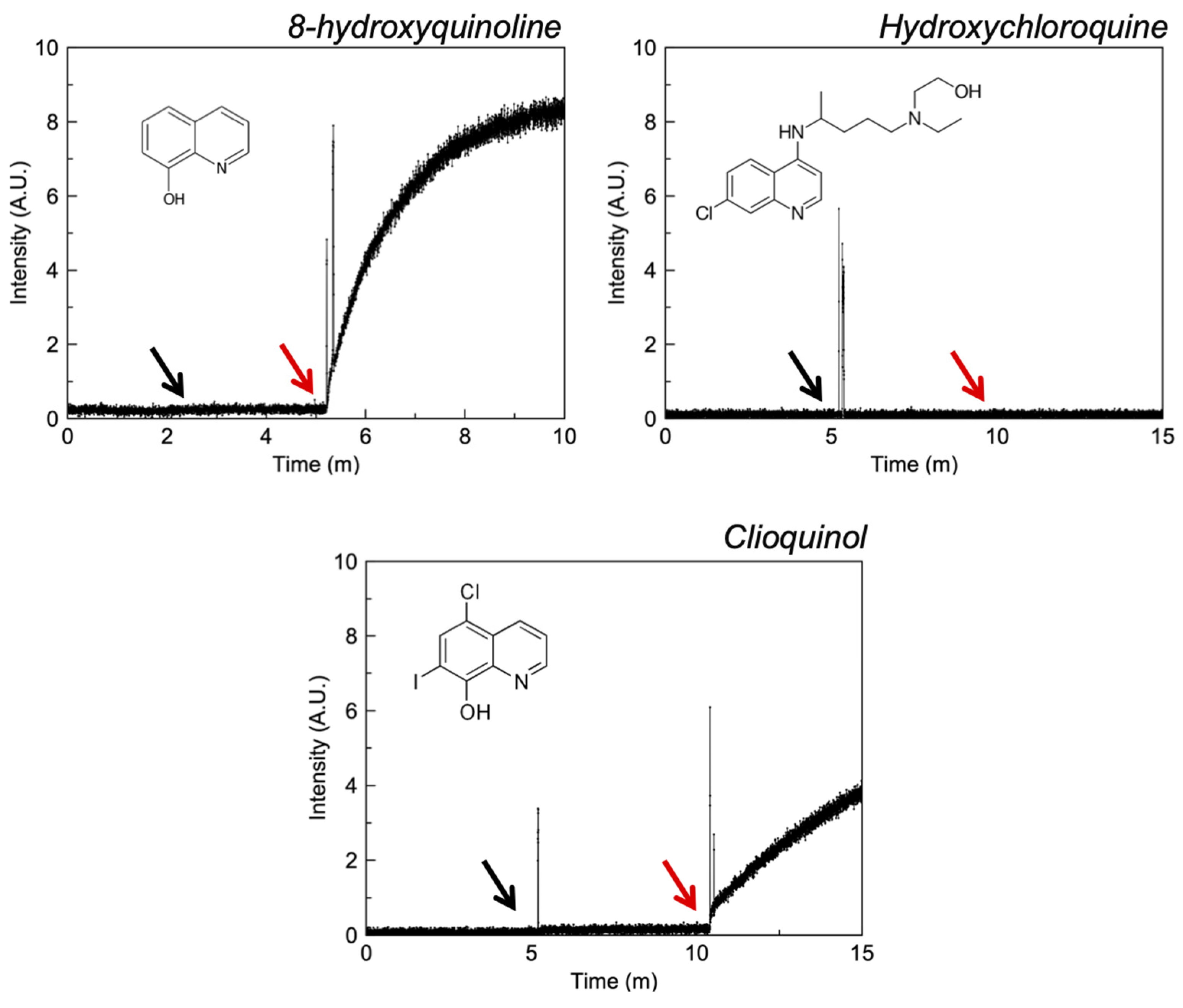
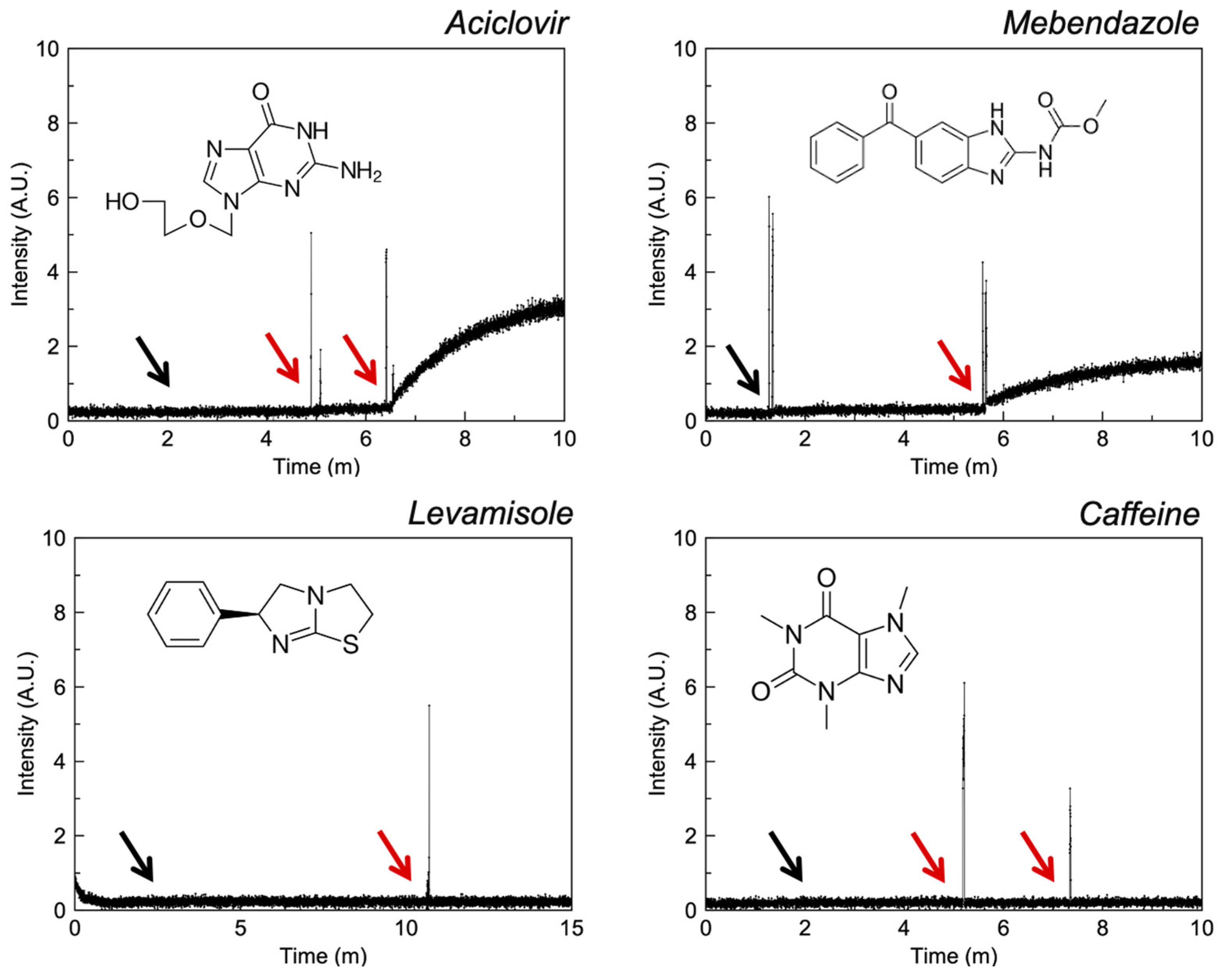
3.2.1. Quinolines
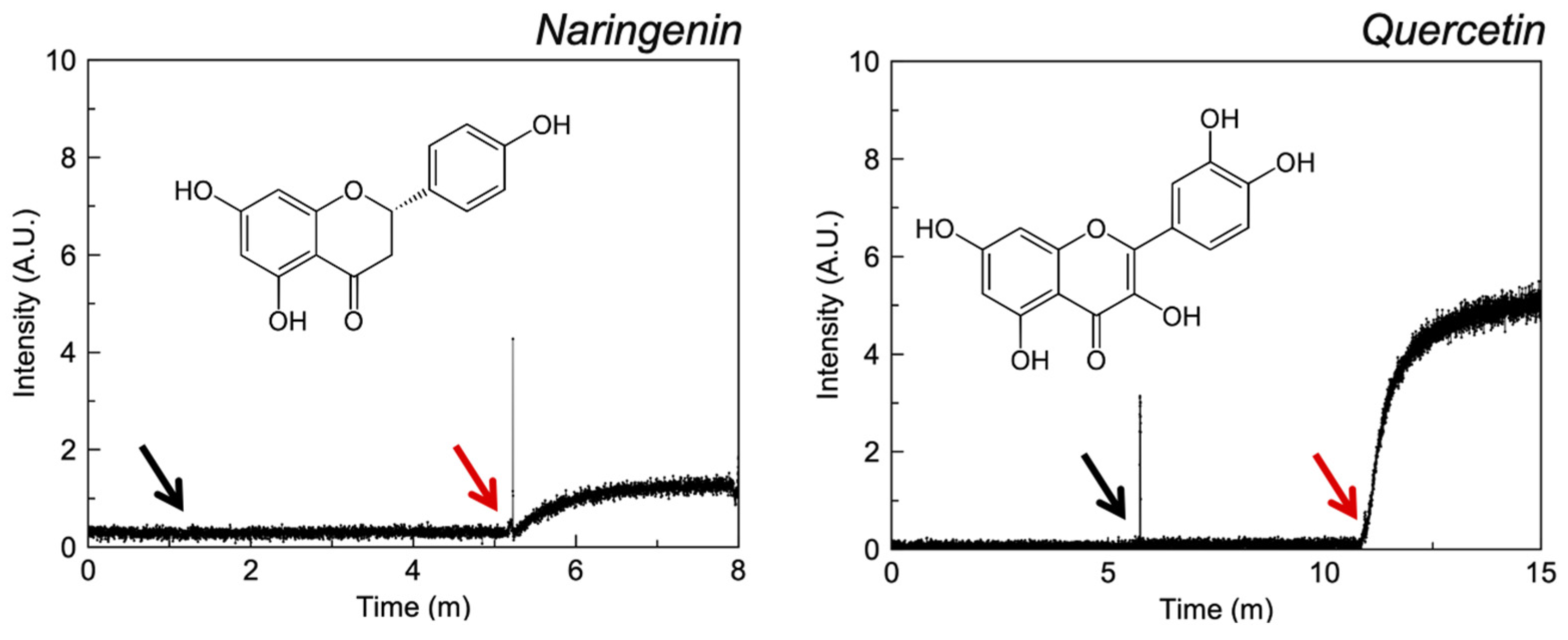
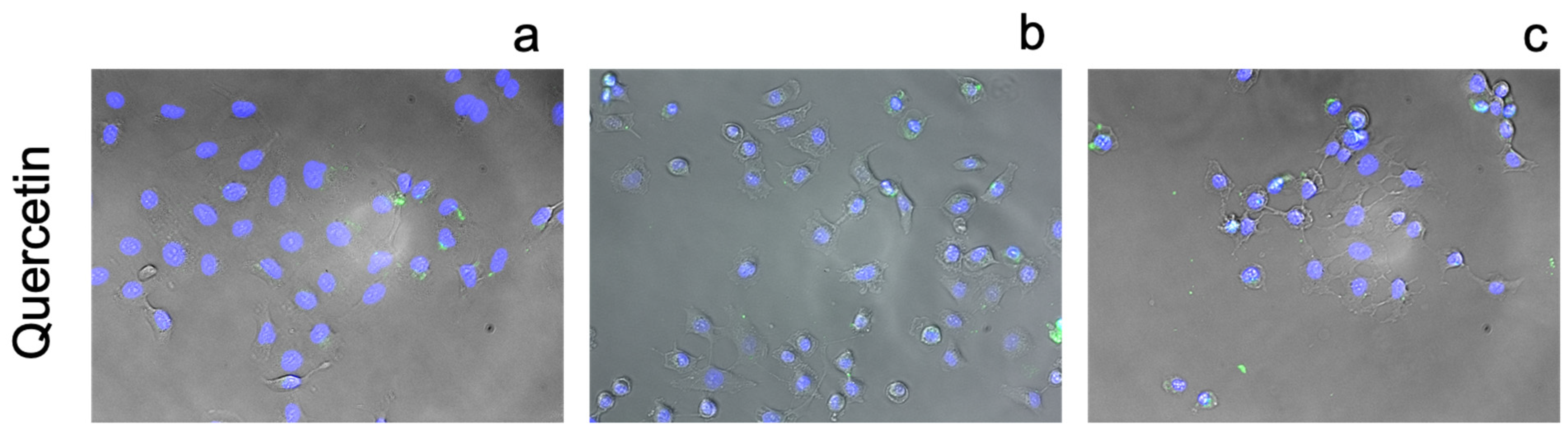
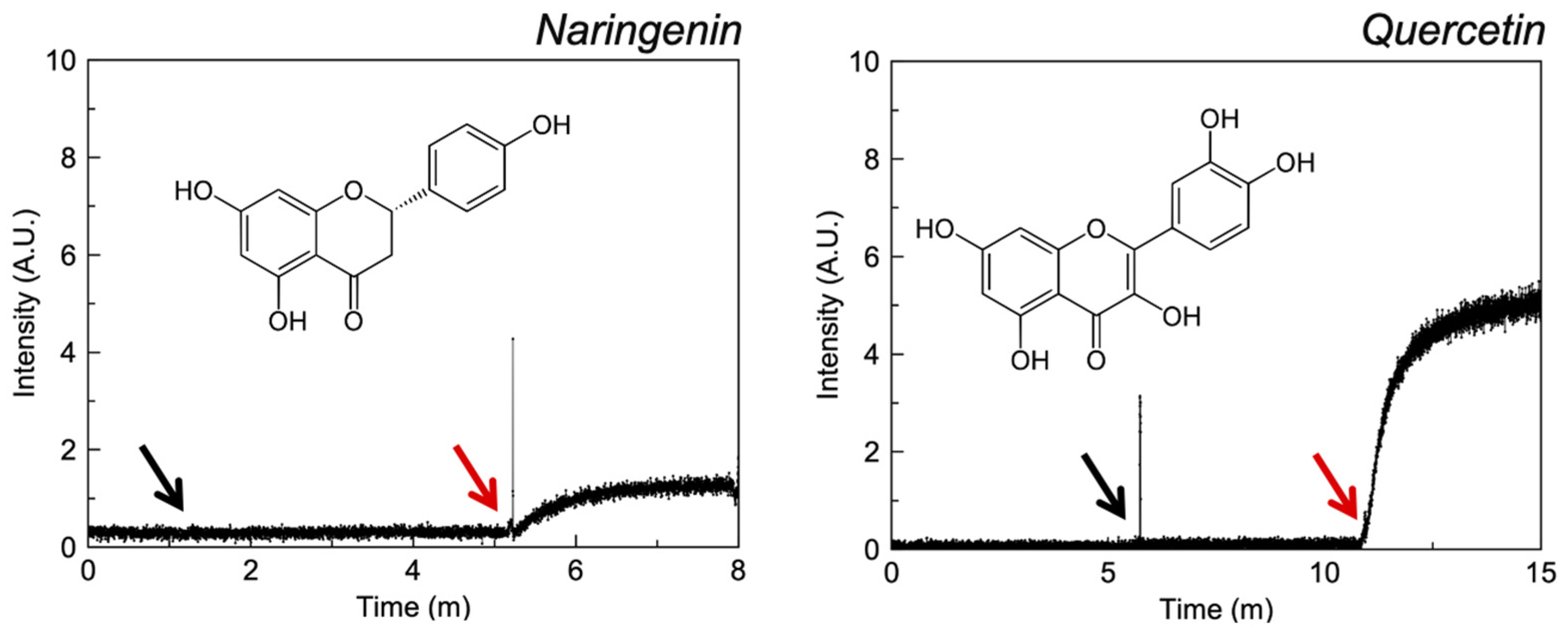
3.2.2. Flavonoids
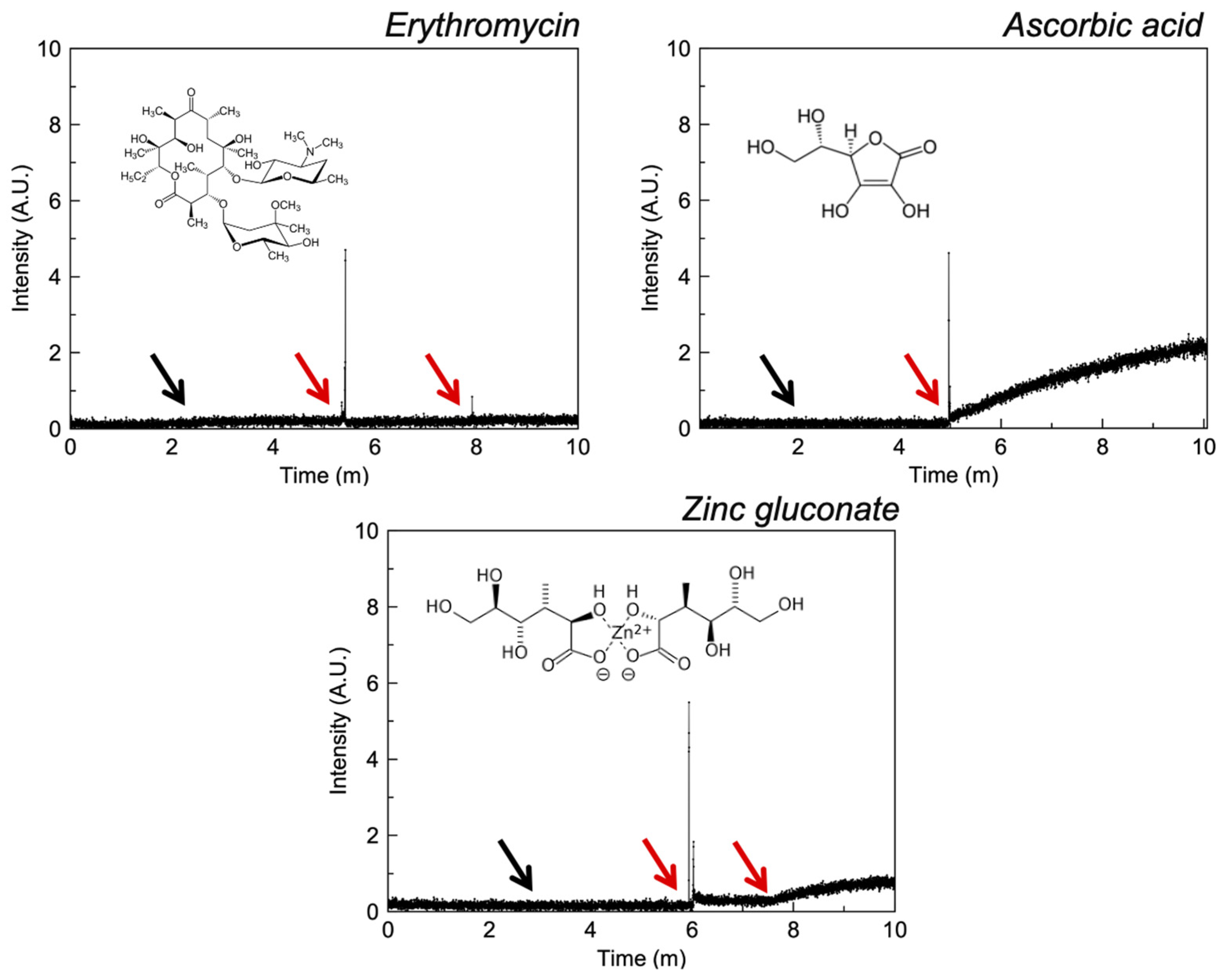
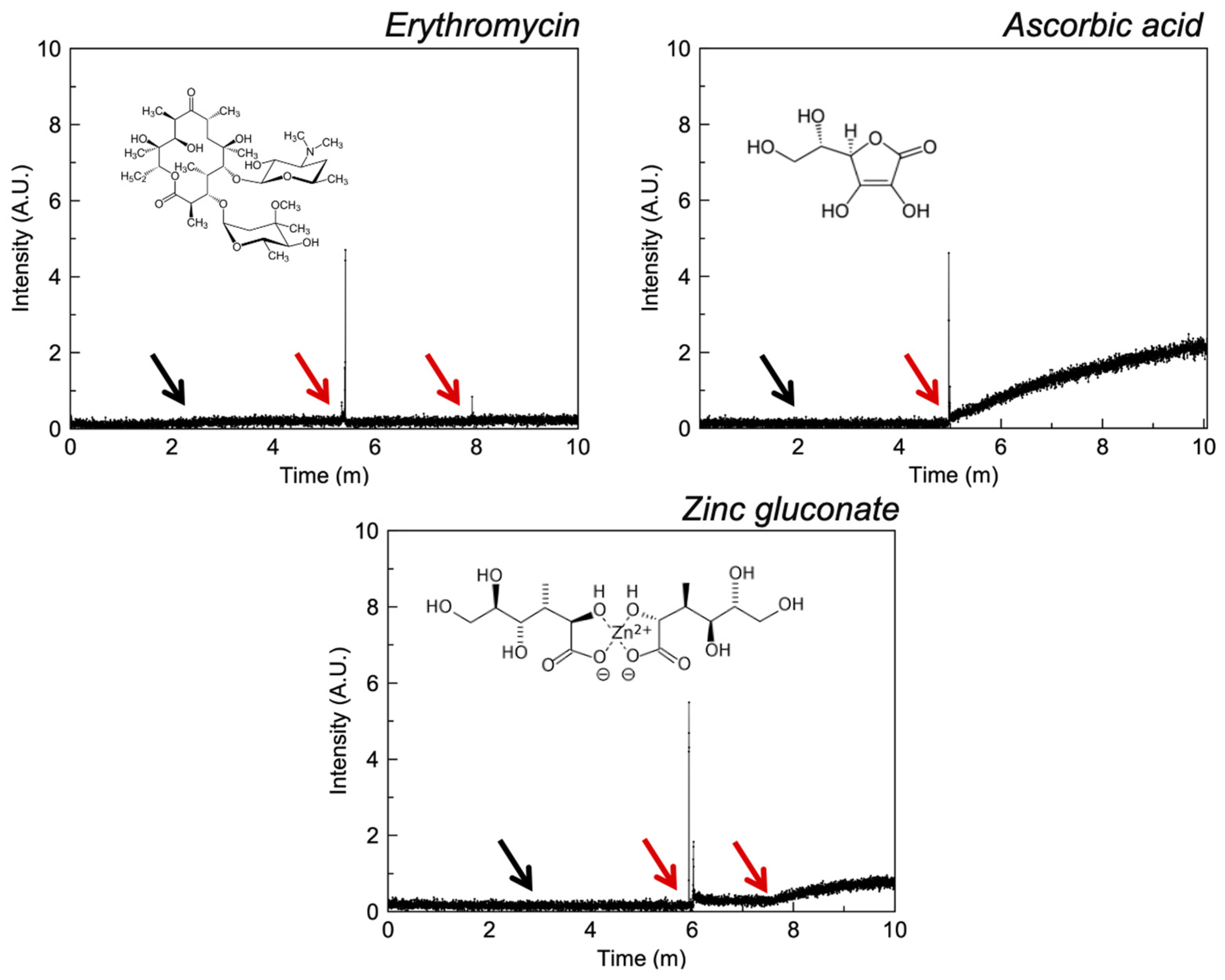
3.2.3. Polyols
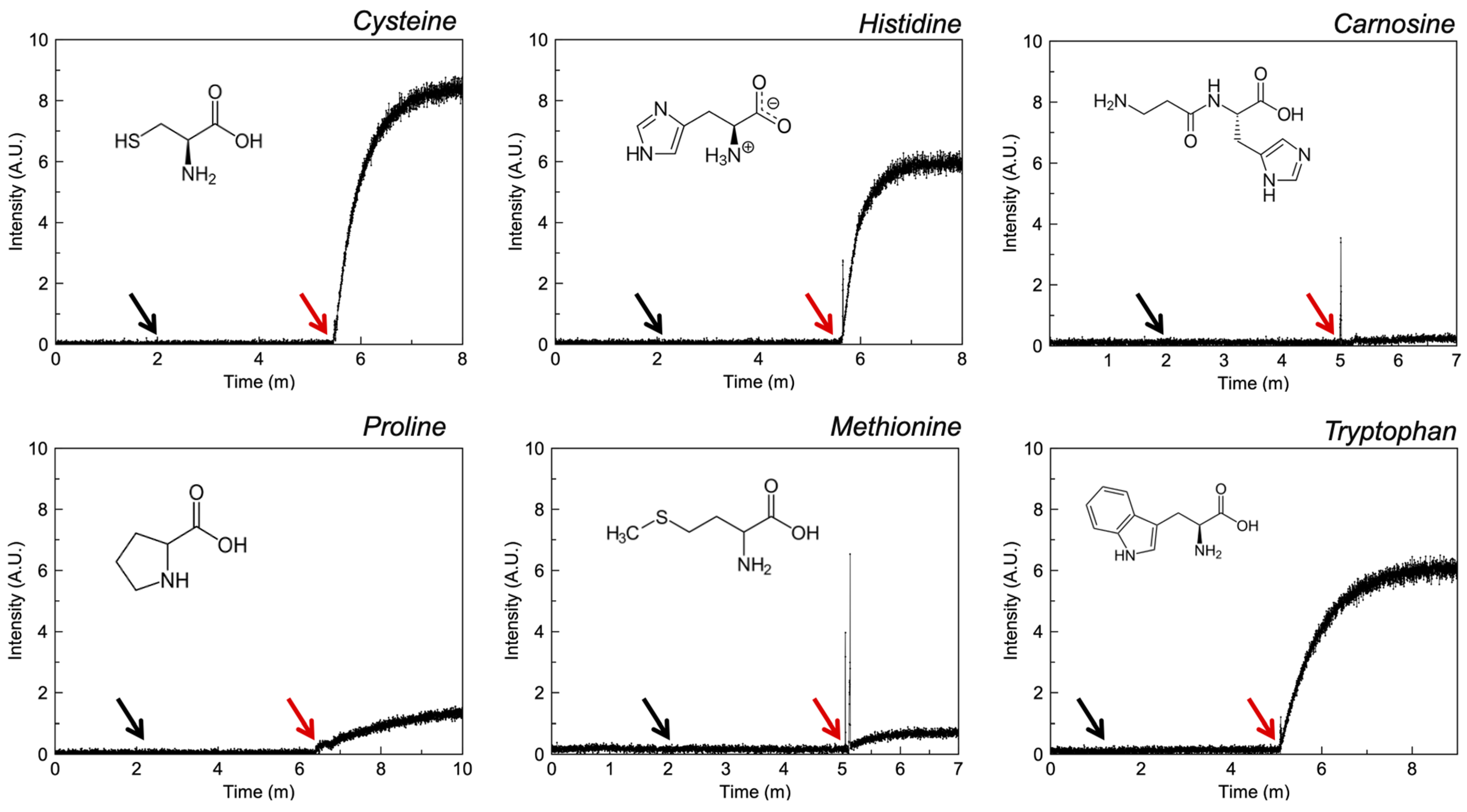
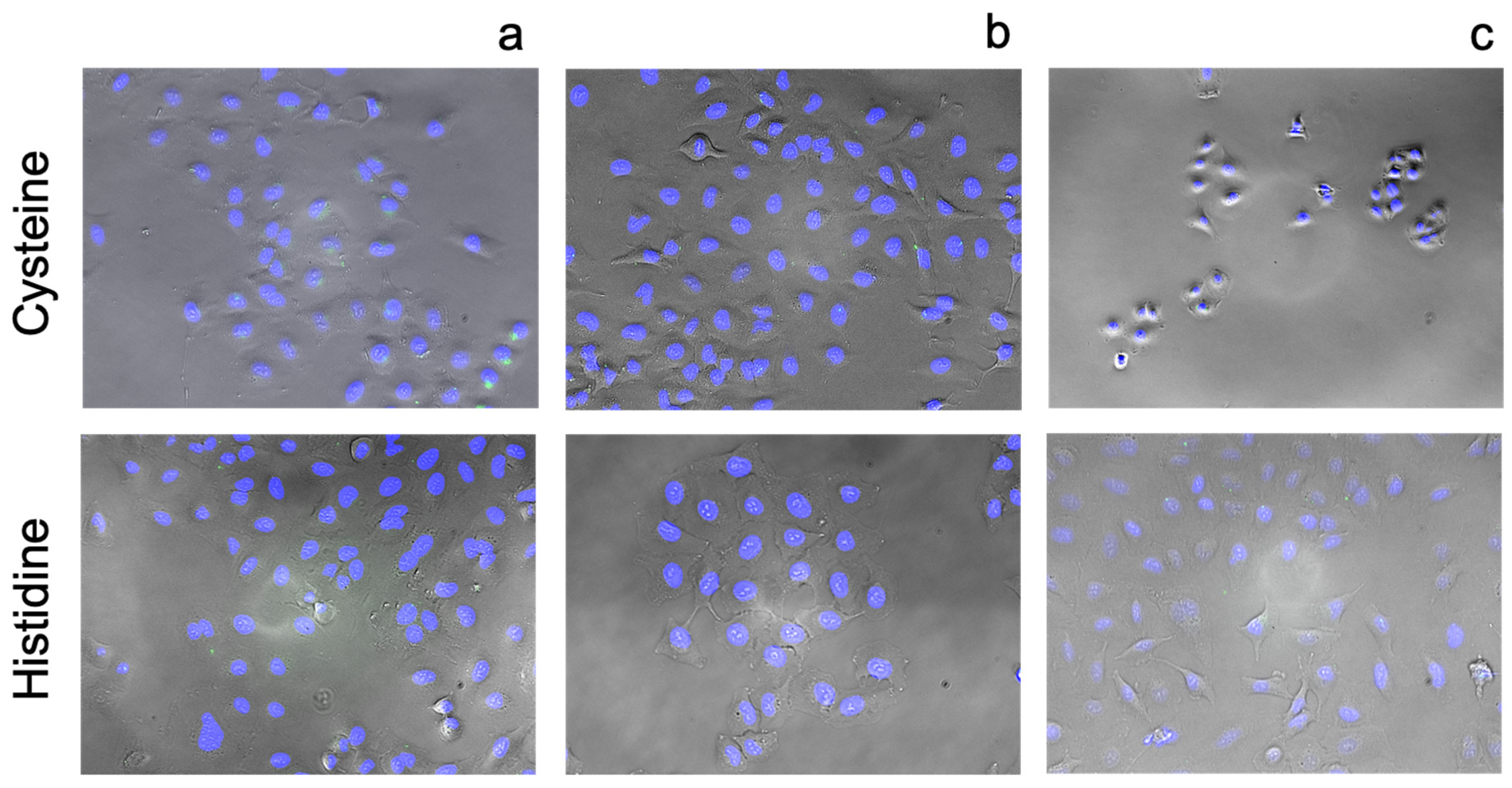
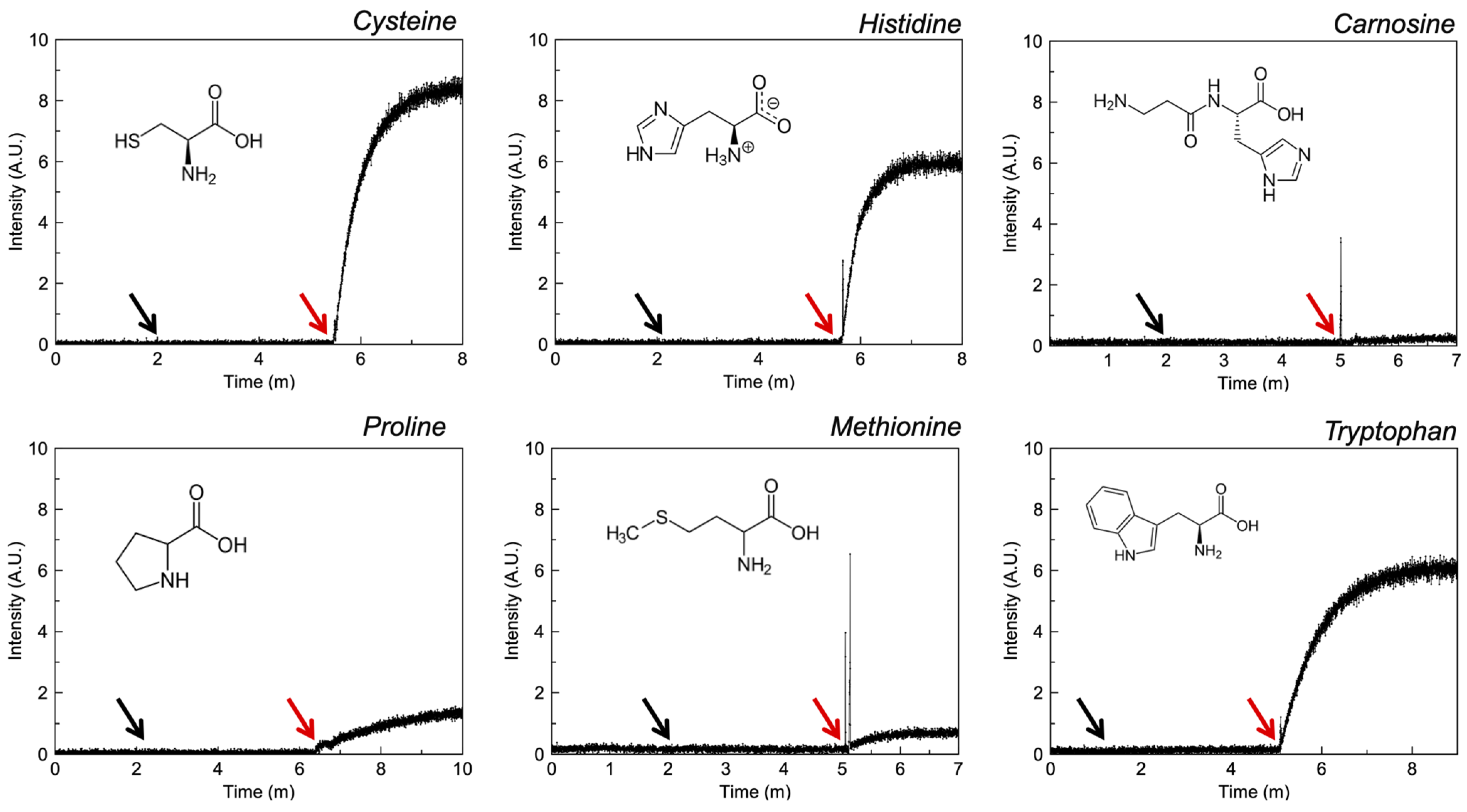
3.2.4. Amino Acids
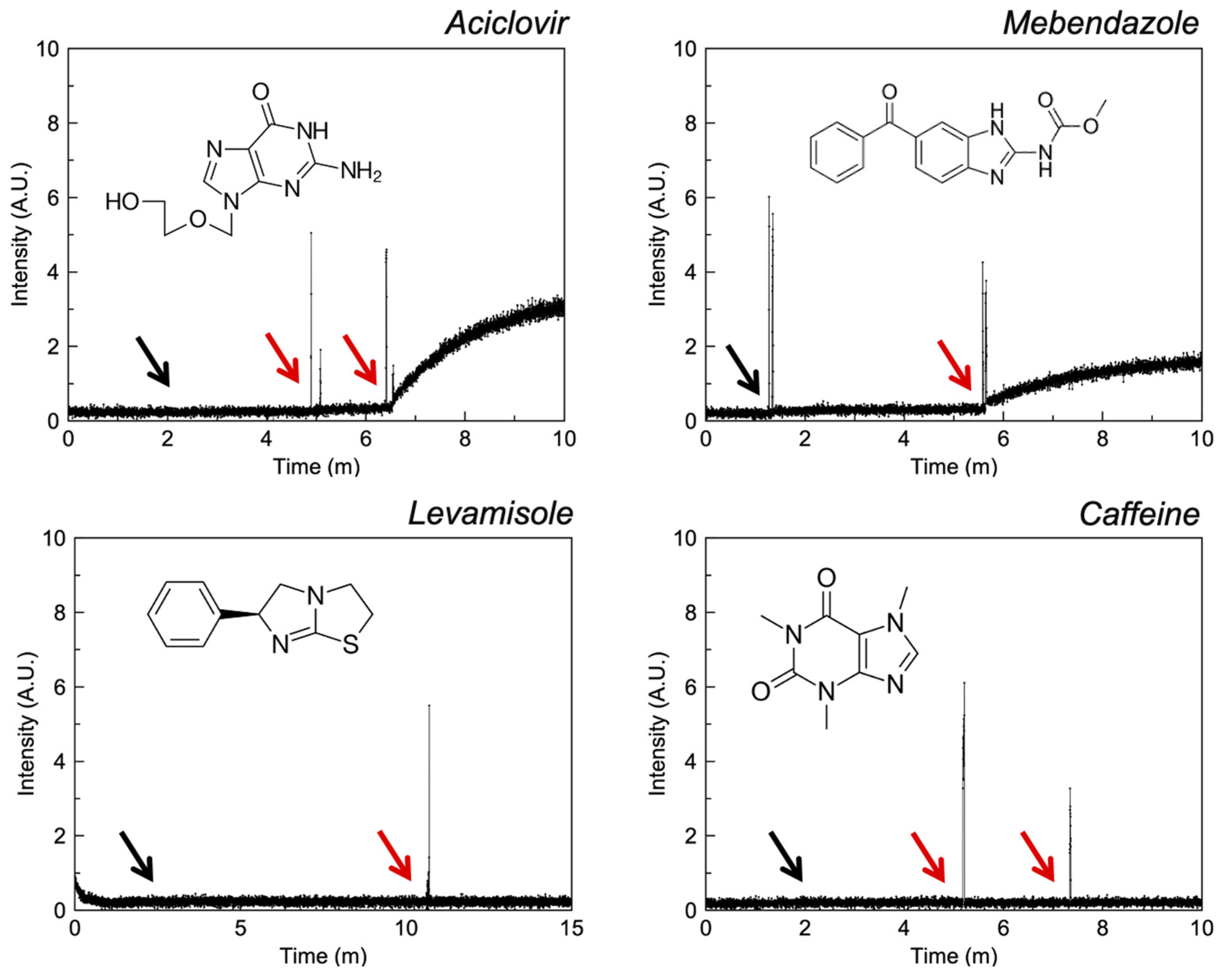
3.2.5. Imidazoles
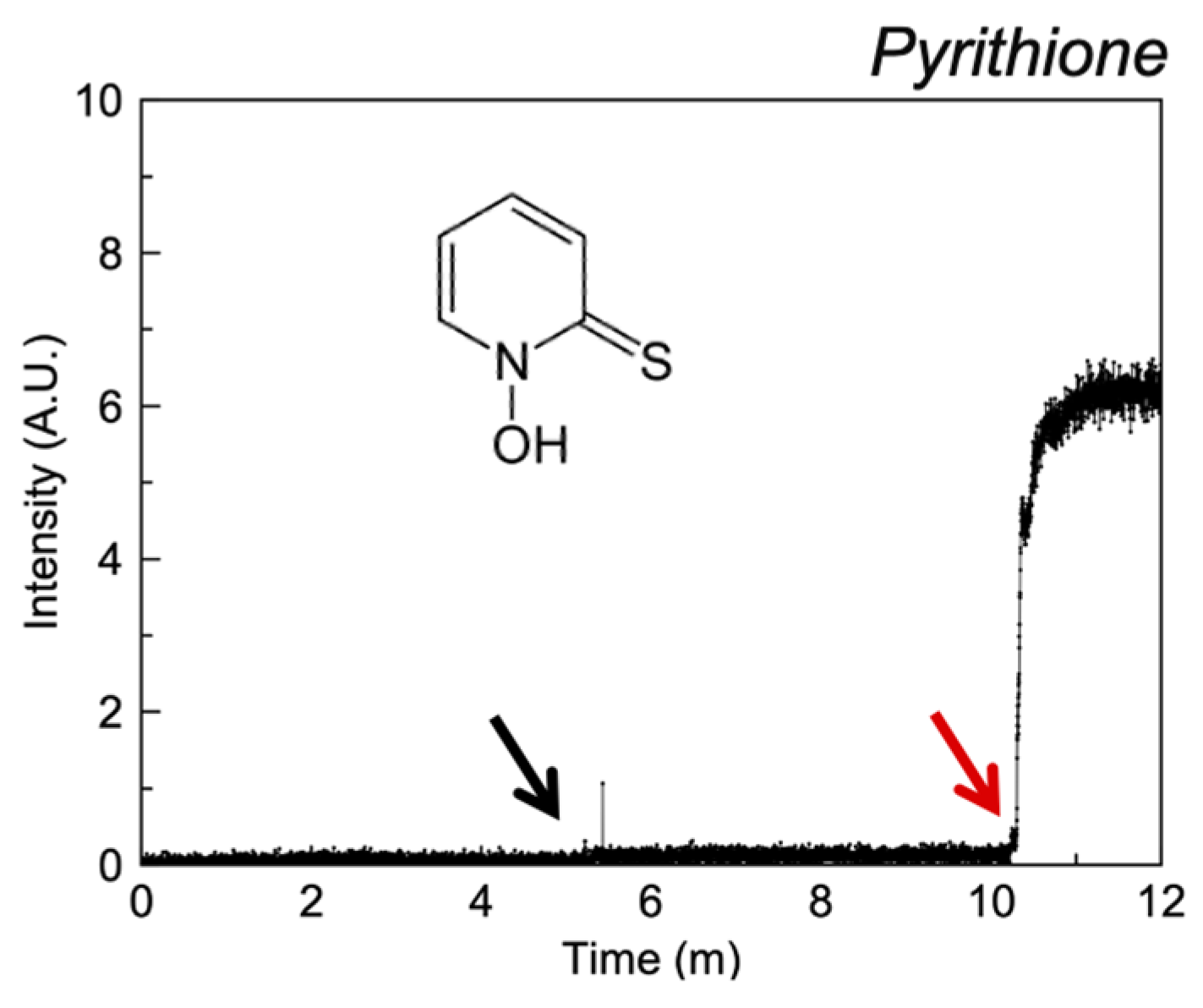
3.2.6. Miscellaneous
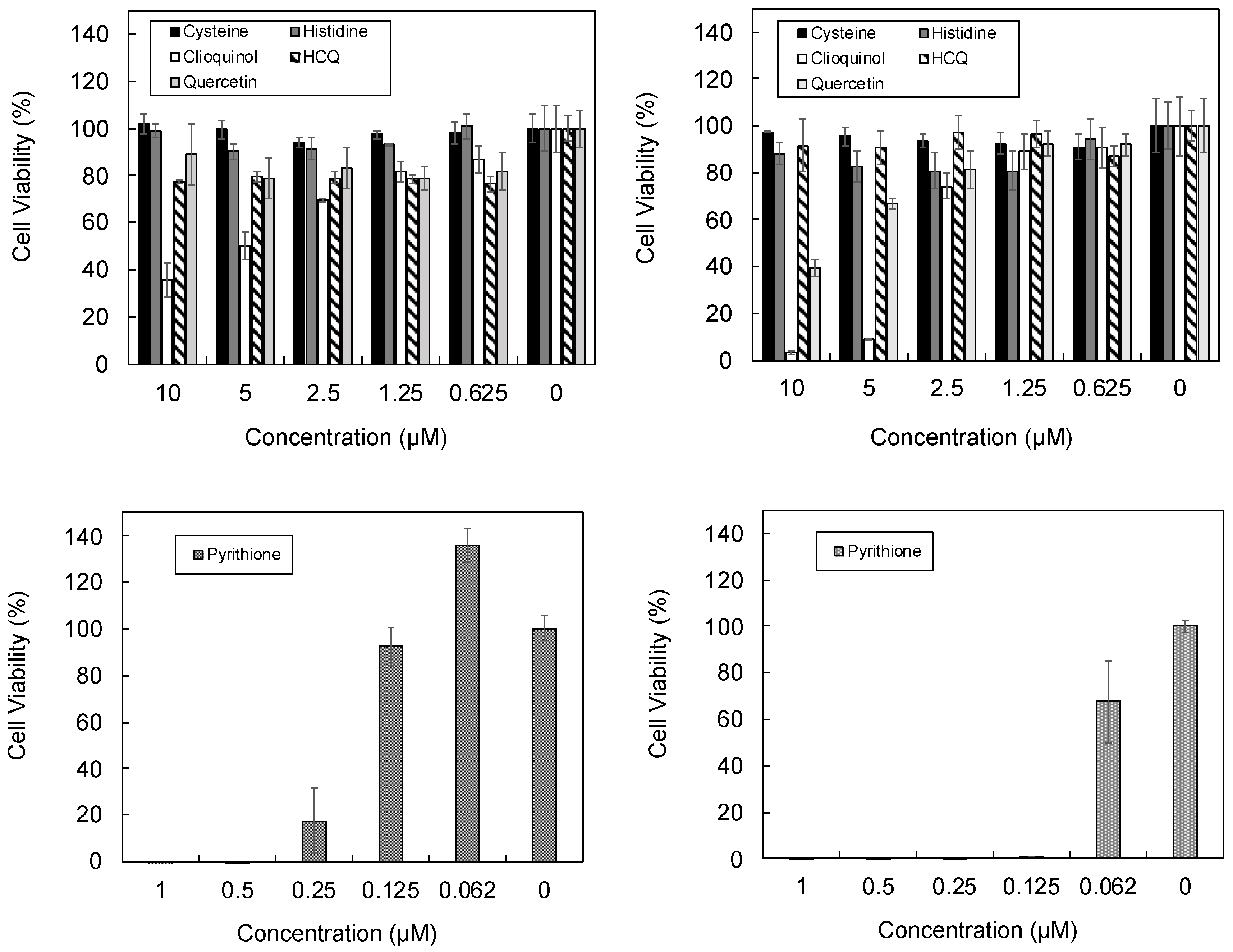
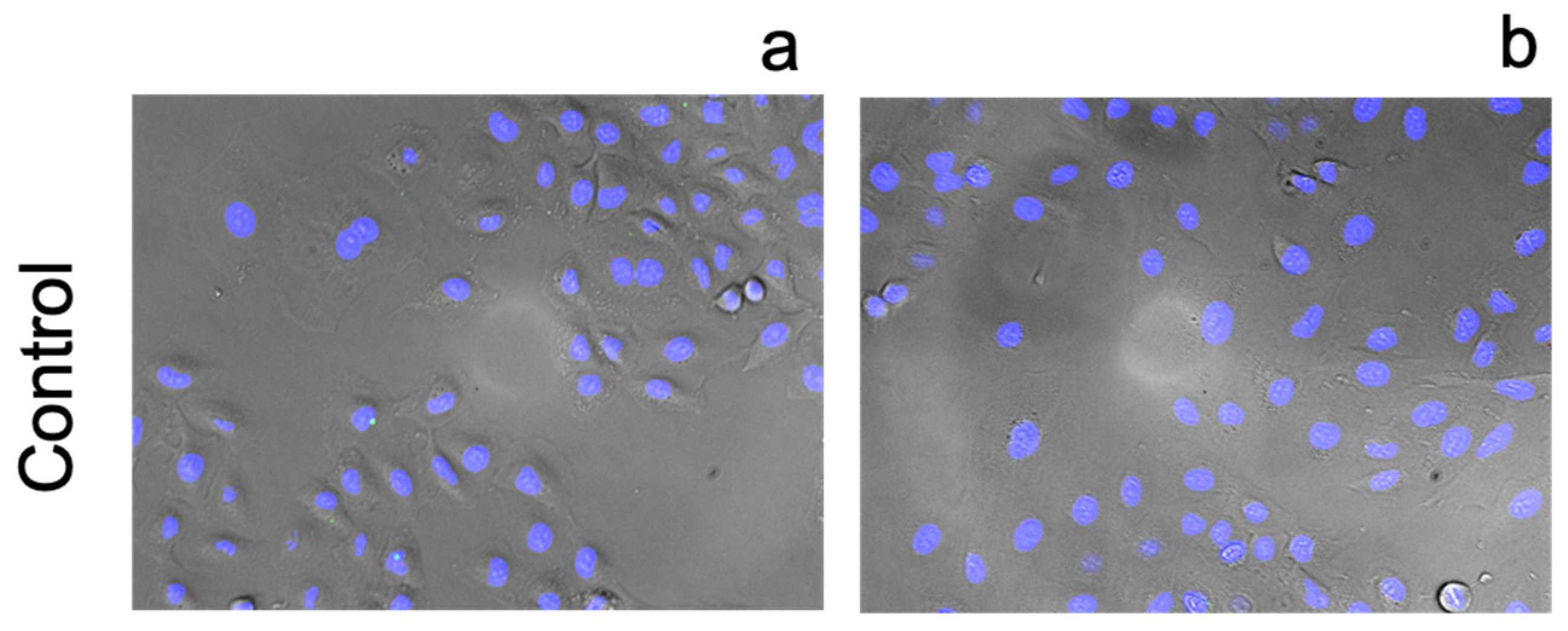
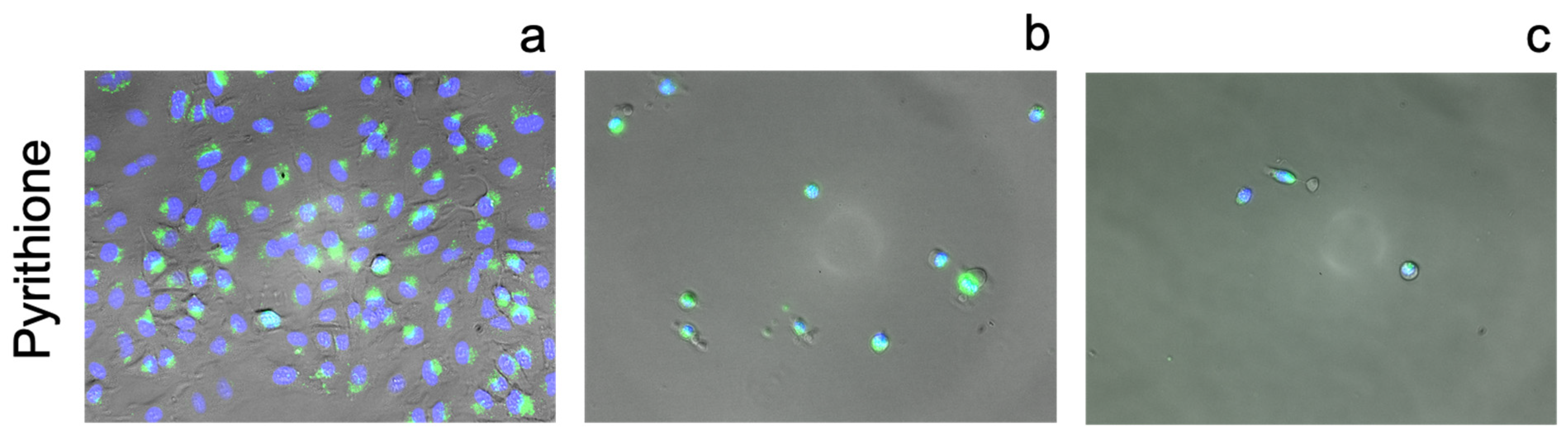
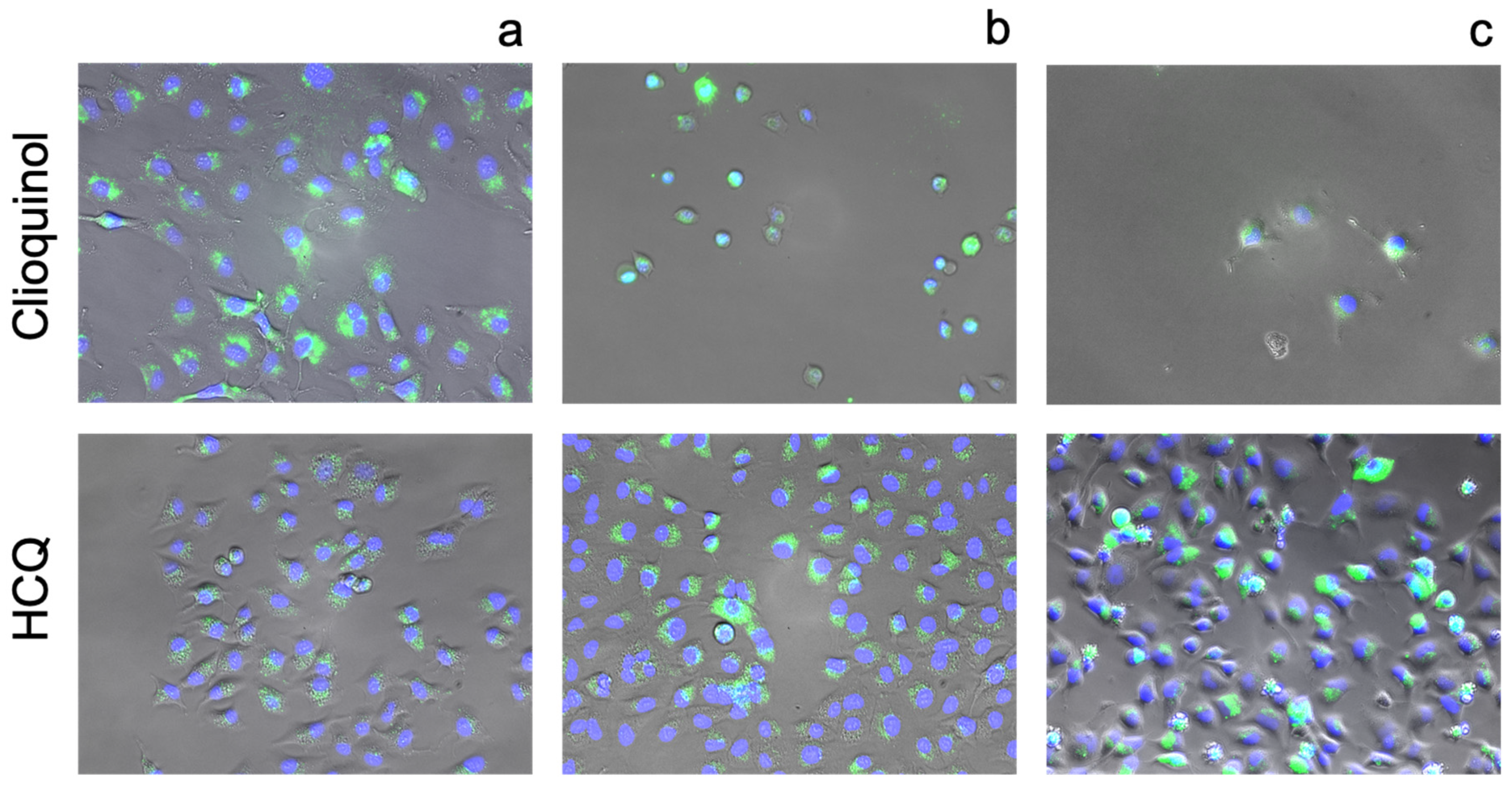
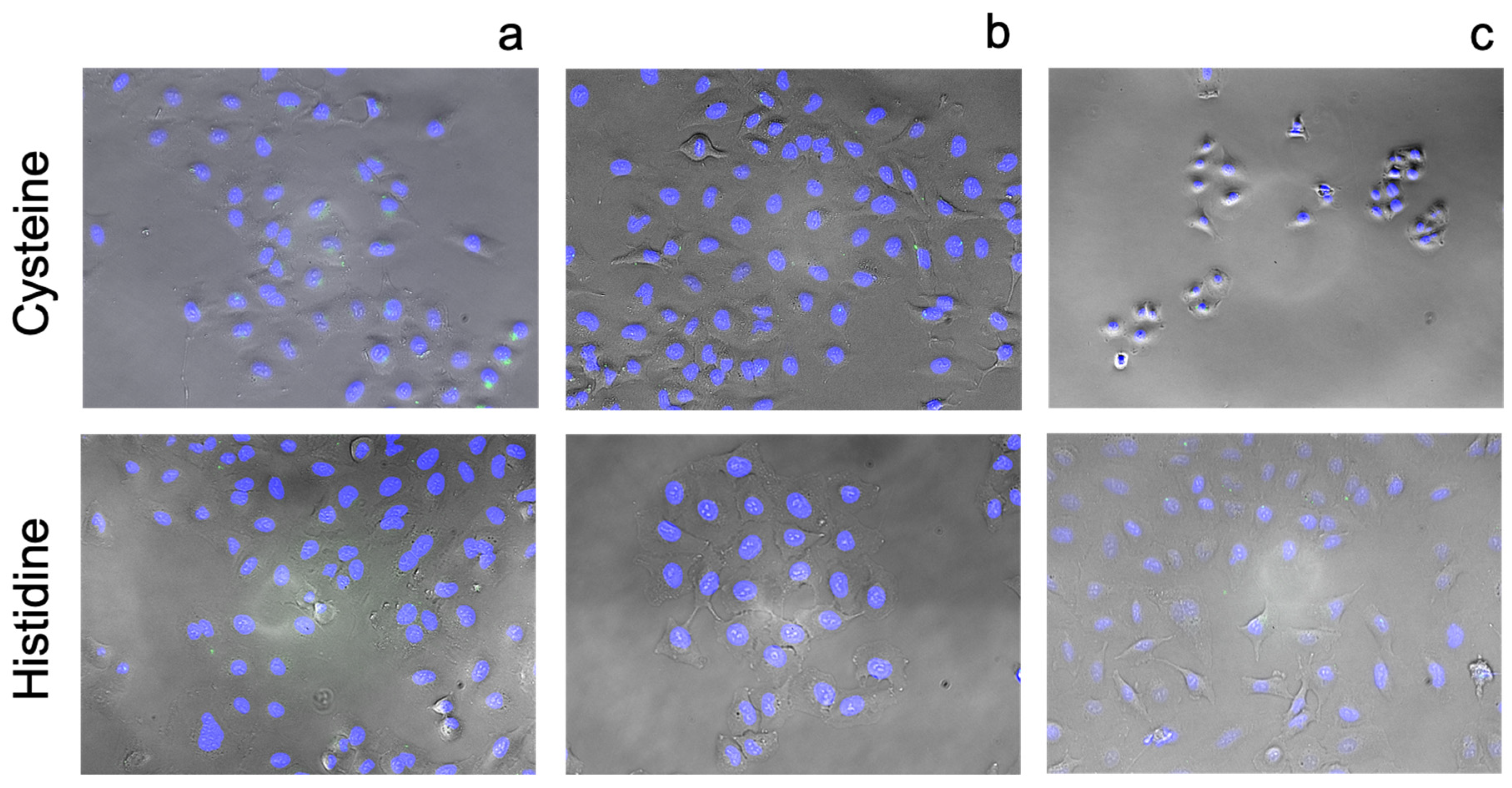
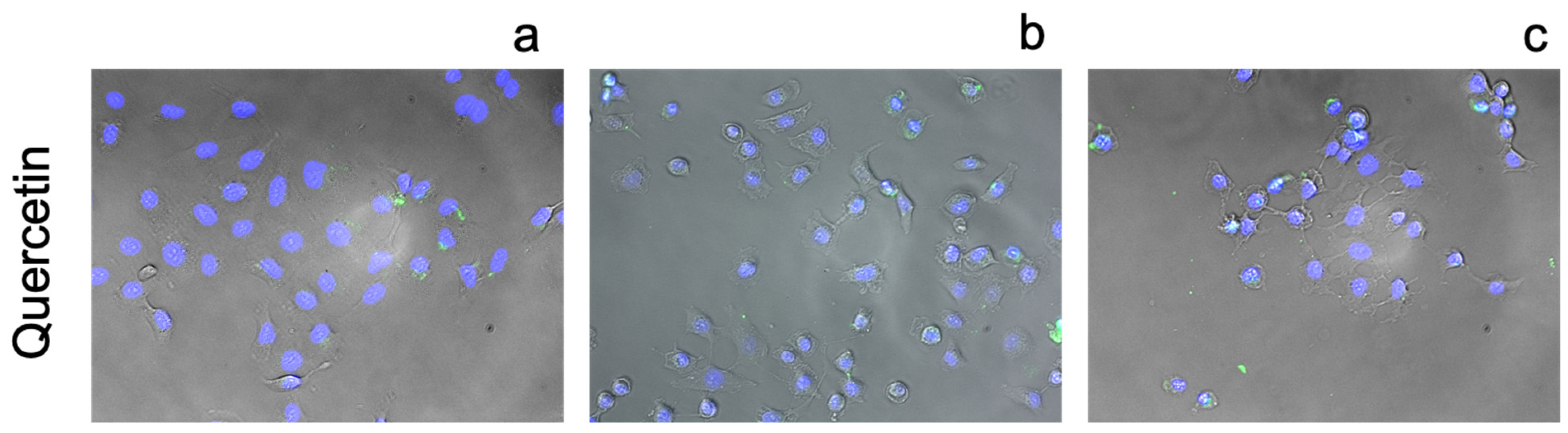
3.3. Cellular Ionophorism with Lead Compounds
4. Discussion
4.1. Identification of Effective Zinc Chelators with CSD Analysis
4.2. Quinolines
4.3. Flavonoids
4.4. Polyols
4.5. Amino Acids
4.6. Imidazoles
4.7. Pyrithione
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kavanagh, O.N.; Albadarin, A.; Croker, D.M.; Healy, A.M.; Walker, G. Maximising success in multidrug formulation development: A review. J. Control. Release 2018, 283, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Vallee, B.L.; Falchuk, K.H. The biochemical basis of zinc physiology. Physiol. Rev. 1993, 73, 79–118. [Google Scholar] [CrossRef] [PubMed]
- Frederickson, C.J.; Koh, J.Y.; Bush, A.I. The neurobiology of zinc in health and disease. Nat. Rev. Neurosci. 2005, 6, 449–462. Available online: www.nature.com/reviews/neuro (accessed on 9 September 2021). [CrossRef] [PubMed]
- Read, S.A.; Obeid, S.; Ahlenstiel, C.; Ahlenstiel, G. The Role of Zinc in Antiviral Immunity. Adv. Nutr. 2019, 10, 696–710. Available online: https://pmc/articles/PMC6628855/?report=abstract (accessed on 10 September 2021). [CrossRef] [Green Version]
- Frederickson, C.J.; Maret, W.; Cuajungco, M.P. Zinc and Excitotoxic Brain Injury: A New Model [Internet]. Neuroscientist 2004, 10, 18–25. Available online: https://pubmed.ncbi.nlm.nih.gov/14987444/ (accessed on 9 September 2021). [CrossRef] [PubMed]
- Wilson, B.R.; Bogdan, A.R.; Miyazawa, M.; Hashimoto, K.; Tsuji, Y. Siderophores in Iron Metabolism: From Mechanism to Therapy Potential. Trends Mol. Med. 2016, 22, 1077–1090. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, T.C.; Nolan, E.M. Beyond iron: Non-classical biological functions of bacterial siderophores. Dalt. Trans. 2015, 44, 6320–6339. Available online: https://pmc.com/articles/PMC4375017/?report=abstract (accessed on 11 September 2021). [CrossRef] [PubMed] [Green Version]
- Pressman, B.C. Biological Applications of Ionophores. Annu. Rev. Biochem. 1976, 45, 501–530. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, C.J. Cyclic polyethers and their complexes with metal salts. J. Am. Chem. Soc. 1967, 89, 7017–7036. [Google Scholar] [CrossRef]
- Kaushik, V.; Yakisich, J.S.; Kumar, A.; Azad, N.; Iyer, A.K. Ionophores: Potential use as anticancer drugs and chemosensitizers. Cancers 2018, 10, 360. Available online: https://pmc/articles/PMC6211070/?report=abstract (accessed on 10 September 2021). [CrossRef] [PubMed] [Green Version]
- Singh, N.; Kaur, N.; McCaughan, B.; Callan, J.F. Ratiometric fluorescent detection of Cu(II) in semi-aqueous solution using a two-fluorophore approach. Tetrahedron Lett. 2010, 51, 3385–3387. [Google Scholar] [CrossRef]
- Prasad, A.S. Zinc deficiency. BMJ 2003, 326, 409–410. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Zhou, Y.; Lind, S.E.; Ding, W.Q. Clioquinol targets zinc to lysosomes in human cancer cells. Biochem. J. 2009, 417, 133–139. Available online: https://portlandpress.com/biochemj/article/417/1/133/45106/Clioquinol-targets-zinc-to-lysosomes-in-human (accessed on 11 September 2021). [CrossRef] [PubMed] [Green Version]
- Costello, L.C.; Franklin, R.B. A Proposed Efficacious Treatment with Clioquinol (Zinc Ionophore) and Cabergoline (Prolactin Dopamine Agonist) for the Treatment of Terminal Androgen-independent Prostate Cancer. Why and How? J. Clin. Res. Oncol. 2019, 2. [Google Scholar]
- Harbison-Price, N.; Ferguson, S.A.; Heikal, A.; Taiaroa, G.; Hards, K.; Nakatani, Y.; Rennison, D.; Brimble, M.A.; El-Deeb, I.M.; Bohlmann, L.; et al. Multiple Bactericidal Mechanisms of the Zinc Ionophore PBT2. mSphere 2020, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohlmann, L.; De Oliveira, D.M.P.; El-Deeb, I.M.; Brazel, E.B.; Harbison-Price, N.; Ong, C.-L.; Rivera-Hernandez, T.; Ferguson, S.A.; Cork, A.J.; Phan, M.-D.; et al. Chemical Synergy between Ionophore PBT2 and Zinc Reverses Antibiotic Resistance. mBio 2018, 9, e02391-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecel, A.; Ostrowska, M.; Stokowa-Sołtys, K.; Wątły, J.; Dudek, D.; Miller, A.; Potocki, S.; Witkiewicz, A.M.; Martin, A.D.; Kozlowski, H.; et al. Zinc(II)—The Overlooked Éminence Grise of Chloroquine’s Fight against COVID-19? Pharmaceuticals 2020, 13, 228. Available online: https://www.mdpi.com/1424-8247/13/9/228 (accessed on 11 September 2021). [CrossRef] [PubMed]
- Puerta, A.E.; Michael O Griffin, A.E.; Jana, A.; Lewis Diego Romero-Perez, A.E.; Ricardo Garcia Francisco, J.; Villarreal, A.E.; Seth, M.; Cohen, D.T. Heterocyclic zinc-binding groups for use in next-generation matrix metalloproteinase inhibitors: Potency, toxicity, and reactivity. J. Biol. Inorg. Chem. 2006, 11, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Vaden, R.M.; Guillen, K.P.; Salvant, J.M.; Santiago, C.B.; Gibbons, J.B.; Pathi, S.S.; Arunachalam, S.; Sigman, M.S.; Looper, R.E.; Welm, B.E. A Cancer-Selective Zinc Ionophore Inspired by the Natural Product Naamidine A. ACS Chem. Biol. 2018, 14, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Bangham, A.D.; Standish, M.M.; Watkins, J.C. Diffusion of univalent ions across the lamellae of swollen phospholipids. J. Mol. Biol. 1965, 13, 238–252. [Google Scholar] [CrossRef]
- Clergeaud, G.; Dabbagh-Bazarbachi, H.; Ortiz, M.; Fernández-Larrea, J.B.; O’Sullivan, C.K. A simple liposome assay for the screening of zinc ionophore activity of polyphenols. Food Chem. 2016, 197, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Aleshin, G.Y.; Egorova, B.V.; Priselkova, A.B.; Zamurueva, L.S.; Khabirova, S.Y.; Zubenko, A.D.; Karnoukhova, V.A.; Fedorova, O.A.; Kalmykov, S.N. Zinc and copper complexes with azacrown ethers and their comparative stability in vitro and in vivo. Dalt. Trans. 2020, 49, 6249–6258. Available online: https://pubs.rsc.org/en/content/articlehtml/2020/dt/d0dt00645a (accessed on 11 September 2021). [CrossRef] [PubMed]
- Tetteh, S. Coordination Behavior of Ni2+, Cu2+, and Zn2+ in Tetrahedral 1-Methylimidazole Complexes: A DFT/CSD Study. Bioinorg. Chem. Appl. 2018, 2018, 3157969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruccolo, S.; Rauch, M.; Parkin, G. Synthesis and Structural Characterization of Tris(isopropylbenzimidazol-2-ylthio)methyl Zinc Complexes, [TitmPriBenz]ZnX: Modulation of Transannular Zn–C Interactions. Organometallics 2018, 37, 1708–1718. [Google Scholar] [CrossRef]
- Mirzaei, M.; Hassanpoor, A.; Alizadeh, H.; Gohari, M.; Blake, A.J. An eight-coordinate zinc complex containing the highly pre-organized ligand 1,10-phenanthroline-2,9-dicarboxylic acid: Solvothermal synthesis, supramolecular structure and CSD studies. J. Mol. Struct. 2018, 1171, 626–630. [Google Scholar] [CrossRef]
- Jowett, L.A.; Gale, P.A. Supramolecular methods: The chloride/nitrate transmembrane exchange assay. Supramol. Chem. 2019, 31, 297–312. [Google Scholar] [CrossRef]
- Carrel, A.; Burrows, M.T. Cultivation of tissues in vitro and its technique. J. Exp. Med. 1911, 13, 387–396. Available online: http://rupress.org/jem/article-pdf/13/3/387/1086204/387.pdf (accessed on 30 October 2021). [CrossRef] [PubMed]
- Darling, J.L. Tissue Culture Techniques. An Introduction. J. Clin. Pathol. 1996, 49, 438. Available online: https://books.google.com/books/about/Tissue_Culture_Techniques.html?id=tl3dBwAAQBAJ (accessed on 30 October 2021). [CrossRef] [Green Version]
- Wang, B.; Zhang, J.; Chen, C.; Xu, G.; Qin, X.; Hong, Y.; Bose, D.D.; Qiu, F.; Zou, Z. The size of zinc oxide nanoparticles controls its toxicity through impairing autophagic flux in A549 lung epithelial cells. Toxicol. Lett. 2018, 285, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Namazi, M.R. Zinc-levamisole combination: Powerful synergistic immunopotentiation? Med. Hypotheses 2006, 66, 1253. Available online: https://pubmed.ncbi.nlm.nih.gov/16434148/ (accessed on 24 June 2021). [CrossRef] [PubMed]
- Dabbagh-Bazarbachi, H.; Clergeaud, G.; Quesada, I.M.; Ortiz, M.; Sullivan, C.O.; Fernández-Larrea, J.B. Zinc Ionophore Activity of Quercetin and Epigallocatechin-gallate: From Hepa 1-6 Cells to a Liposome Model. J. Agric. Food Chem. 2014, 62, 8085–8093. [Google Scholar] [CrossRef] [PubMed]
- Di Vaira, M.; Bazzicalupi, C.; Orioli, P.; Messori, L.; Bruni, B.; Zatta, P. Clioquinol, a drug for Alzheimer’s disease specifically interfering with brain metal metabolism: Structural characterization of its zinc(II) and copper(II) complexes. Inorg. Chem. 2004, 43, 3795–3797. Available online: http://pubs.acs.org (accessed on 31 May 2021). [CrossRef] [PubMed]
- Figueroa, E.E.; Denton, J.S. Zinc pyrithione activates the volume-regulated anion channel through an antioxidant-sensitive mechanism. Am. J. Physiol. Physiol. 2021, 320, C1088–C1098. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Cho, Y.-J.; Lee, Y.W.; Jung, W.H. Understanding the Mechanism of Action of the Anti-Dandruff Agent Zinc Pyrithione against Malassezia restricta. Sci. Rep. 2018, 8, 12086. [Google Scholar] [CrossRef] [PubMed]
- Reeder, N.; Xu, J.; Youngquist, R.; Schwartz, J.; Rust, R.; Saunders, C. The antifungal mechanism of action of zinc pyrithione. Br. J. Dermatol. 2011, 165, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Chandler, C.J.; Segel, I.H. Mechanism of the Antimicrobial Action of Pyrithione: Effects on Membrane Transport, ATP Levels, and Protein Synthesis. Antimicrob. Agents Chemother. 1978, 14, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.S.; Hao, Y.Y.; Wang, H.; Zhou, H.F.; Liu, X.G.; Chen, M.W. The effects of crystal structure on optical absorption/photoluminescence of bis(8-hydroxyquinoline)zinc. Solid State Commun. 2005, 136, 318–322. [Google Scholar]
- Basu, S.; Hajra, A.; Chattopadhyay, A. An ambient complexation reaction of zinc acetate and ascorbic acid leads to a new form of nanoscale particles with emergent optical properties. Nanoscale Adv. 2021, 3, 3298–3305. [Google Scholar] [CrossRef]
- Tajmir-Riahi, H.A. Coordination chemistry of vitamin C. Part II. Interaction of L-ascorbic acid with Zn(II), Cd(II), Hg(II), and Mn(II) ions in the solid state and in aqueous solution. J. Inorg. Biochem. 1991, 42, 47–55. [Google Scholar] [CrossRef]
- Luedders, W.L.; Willins, R.E. An Erythromycin Compound, a Process for Preparing the Compound and Compositions Containing the Compound Useful in the Treatment of Acne. EP0001871A1, 16 May 1979. [Google Scholar]
- Sawyer, D.T. Metal-Gluconate Complexes. Available online: https://pubs.acs.org/sharingguidelines (accessed on 9 June 2021).
- Sarria, A.L.F.; Vilela, A.F.L.; Frugeri, B.M.; Fernandes, J.B.; Carlos, R.M.; Cass, Q.B.; Cardoso, C.L. Copper (II) and zinc (II) complexes with flavanone derivatives: Identification of potential cholinesterase inhibitors by on-flow assays. J. Inorg. Biochem. 2016, 164, 141–149. [Google Scholar] [CrossRef]
- Wang, H.-L.; Yang, Z.-Y.; Wang, B.-D. Synthesis, Characterization and the Antioxidative Activity of Copper(II), Zinc(II) and Nickel(II) Complexes with Naringenin. Transit. Met. Chem. 2006, 31, 470–474. [Google Scholar] [CrossRef]
- Kalinowska, M.; Świderski, G.; Matejczyk, M.; Lewandowski, W. Spectroscopic, thermogravimetric and biological studies of Na(I), Ni(II) and Zn(II) complexes of quercetin. J. Therm. Anal. Calorim. 2016, 126, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Herrero, L.A.; Cerro-Garrido, J.C.; Terrón-Homar, À. A calorimetric study of 3d metal ions–acyclovir interactions. The 2-hydroxyethoxymethyl group of acyclovir mimics the role of ribose in deoxy-guanosine and guanosine promoting the coordination through N(7). J. Inorg. Biochem. 2001, 86, 677–680. [Google Scholar] [CrossRef]
- AlAjmi, M.F.; Hussain, A.; Rehman, T.; Khan, A.A.; Alam Shaikh, P.; Khan, R.A. Design, Synthesis, and Biological Evaluation of Benzimidazole-Derived Biocompatible Copper(II) and Zinc(II) Complexes as Anticancer Chemotherapeutics. Int. J. Mol. Sci. 2018, 19, 1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamdani, H.E.L.; Amane, M.E.L. Preparation, spectral, antimicrobial properties and anticancer molecular docking studies of new metal complexes [M(caffeine) 4] (PF 6) 2; M = Fe(II), Co(II), Mn(II), Cd(II), Zn(II), Cu(II), Ni(II). J. Mol. Struct. 2019, 1184, 262–270. [Google Scholar] [CrossRef]
- Kovachev, T.B.; Stamberov, P.N.; Ivanov, D.S.; Mitcheva, M.K.; Marinova, S.P.; Astroug, H.A.; Stoychkov, J.N. Synthesis, toxicity and immunomodulating activity of Co(II), Ni(II), Cu(II) and Zn(II) complexes of L(-)-2,3,5,6-tetrahydro-6-phenylimidazo[2,1-b]thiazole (levamisole). Pharmazie 1994, 49, 25–27. [Google Scholar] [PubMed]
- Matsukura, T.; Takahashi, T.; Nishimura, Y.; Ohtani, T.; Sawada, M.; Shibata, K. Characterization of crystalline L-carnosine Zn(II) complex (Z-103), a novel anti-gastric ulcer agent: Tautomeric change of imidazole moiety upon complexation. Chem. Pharm. Bull. 1990, 38, 3140–3146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, C.E.; Antholine, W.E. Chelation Chemistry of Carnosine. Evidence that Mixed Complexes May Occur in Vivo. J. Phys. Chem. 1979, 83, 3314–3319. [Google Scholar] [CrossRef]
- Gockel, P.; Vahrenkamp, H.; Zuberbühler, A.D. Zine Complexes of Cysteine, Histidine, and Derivatives Thereof: Potentiometric determination of their compositions and stabilities. Helvetica Chim. Acta 1993, 76, 511–520. [Google Scholar] [CrossRef]
- Gao, S.; Liu, J.; Ji, M.; Yang, X.; Zhang, F.; Li, Z. Coordination behavior between zinc salts and L-α-methionine. Chin. Sci. Bull. 1998, 43, 1527–1531. [Google Scholar] [CrossRef]
- Darbre, T.; Machuqueiro, M. Zn-Proline catalyzed direct aldol reaction in aqueous media. Chem. Commun. 2003, 3, 1090–1091. Available online: http://www.rsc.org/suppdata/cc/b3/b301117h/ (accessed on 9 June 2021). [CrossRef]
- Abendrot, M.; Chęcińska, L.; Kusz, J.; Lisowska, K.; Zawadzka, K.; Felczak, A.; Kalinowska-Lis, U. Zinc(II) Complexes with Amino Acids for Potential Use in Dermatology: Synthesis, Crystal Structures, and Antibacterial Activity. Molecules 2020, 25, 951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabindra Reddy, P.; Mohan Reddy, A. Synthesis and characterization of mixed ligand complexes of zinc (II) with uridine and amino acids. In Proceedings of the Indian Academy of Sciences-Chemical Sciences; Springer: New Delhi, India, 1998; Volume 110, pp. 385–396. [Google Scholar] [CrossRef]
- Bond, A.D.; Jones, W. Synthesis and Characterisation of a Novel Zinc Pyrithione Hydrate. Mol. Cryst. Liq. Cryst. Sci. Technol Sect. A Mol. Cryst. Liq. Cryst. 2001, 356, 305–313. Available online: https://www.tandfonline.com/action/journalInformation?journalCode=gmcl20 (accessed on 9 June 2021). [CrossRef]
- Barnett, B.L.; Kretschmar, H.C.; Hartman, F.A. Structural Characterization of Bis(N-oxopyridine-2-thionato)zinc(II). Inorg. Chem. 1977, 16, 1834–1838. [Google Scholar] [CrossRef]
- Macintyre, A.C.; Cutler, D.J. Role of Lysosomes in Hepatic Accumulation of Chloroquine. J. Pharm. Sci. 1988, 77, 196–199. [Google Scholar] [CrossRef]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Hosios, A.M.; Hecht, V.C.; Danai, L.V.; Johnson, M.O.; Rathmell, J.C.; Steinhauser, M.L.; Manalis, S.R.; Heiden, M.G.V. Amino Acids Rather than Glucose Account for the Majority of Cell Mass in Proliferating Mammalian Cells. Dev. Cell 2016, 36, 540–549. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.E.; Coody, T.; Jeong, M.-Y.; Berg, J.; Winge, D.R.; Hughes, A.L. Cysteine Toxicity Drives Age-Related Mitochondrial Decline by Altering Iron Homeostasis. Cell 2020, 180, 296–310.e18. [Google Scholar] [CrossRef]
- Nakanishi, T.; Akabane, E.R.; Nanami, M.; Kiyobayashi, Y.; Moriguchi, R.; Hasuike, Y.; Otaki, Y.; Miyagawa, K.; Itahana, R.; Izumi, M. Comparison of Cytotoxicity of Cysteine and Homocysteine for Renal Epithelial Cells. Nephron Exp. Nephrol. 2005, 100, e11–e20. Available online: https://www.karger.com/Article/FullText/84108 (accessed on 24 November 2021). [CrossRef] [PubMed]
- Nishiuch, Y.; Sasaki, M.; Nakayasu, M.; Oikawa, A. Cytotoxicity of cysteine in culture media. In Vitro 1976, 12, 635–638. Available online: https://www.jstor.org/stable/20170352?seq=1#metadata_info_tab_contents (accessed on 6 August 2021). [CrossRef] [PubMed]
- Rauen, U.; Klempt, S.; De Groot, H. Histidine-induced injury to cultured liver cells, effects of histidine derivatives and of iron chelators. Cell Mol. Life Sci. 2007, 64, 192–205. Available online: https://pubmed.ncbi.nlm.nih.gov/17180300/ (accessed on 6 August 2021). [CrossRef]
- Hashemzaei, M.; Far, A.D.; Yari, A.; Heravi, R.E.; Tabrizian, K.; Taghdisi, S.M.; Sadegh, S.E.; Tsarouhas, K.; Kouretas, D.; Tzanakakis, G.; et al. Anticancer and apoptosis-inducing effects of quercetin in vitro and in vivo. Oncol. Rep. 2017, 38, 819–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, S.; Somasagara, R.R.; Hegde, M.; Nishana, M.; Tadi, S.K.; Srivastava, M.; Choudhary, B.; Raghavan, S.C. Quercetin, a Natural Flavonoid Interacts with DNA, Arrests Cell Cycle and Causes Tumor Regression by Activating Mitochondrial Pathway of Apoptosis. Sci. Rep. 2016, 6, 24049. Available online: https://www.nature.com/articles/srep24049 (accessed on 24 November 2021). [CrossRef] [PubMed] [Green Version]
- Ireland, S.M.; Martin, A.C.R. ZincBind—The database of zinc binding sites. Database 2019, 6, baz006. Available online: https://academic.oup.com/database/article/doi/10.1093/database/baz006/5304468 (accessed on 9 June 2021).
- Barnham, K.J.; Gautier, E.C.L.; Kok, G.B.; Krippner, G. 8-Hydroxy Quinoline Derivatives. WO2004007461A1, 16 July 2003. [Google Scholar]
- Egan, T.J.; Hunter, R.; Kaschula, C.H.; Marques, H.M.; Misplon, A.; Walden, J. Structure-Function Relationships in Aminoquinolines: Effect of Amino and Chloro Groups on Quinoline-Hematin Complex Formation, Inhibition of-Hematin Formation, and Antiplasmodial Activity. J. Med. Chem. 2000, 43, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Wang, W.; Zhao, B.; Zhang, S.; Miao, J. Chloroquine inhibits cell growth and induces cell death in A549 lung cancer cells. Bioorganic Med. Chem. 2006, 14, 3218–3222. [Google Scholar] [CrossRef]
- Lu, S.; Sung, T.; Lin, N.; Abraham, R.T.; Jessen, B.A. Lysosomal adaptation: How cells respond to lysosomotropic compounds. PLoS ONE 2017, 12, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundelin, S.P.; Terman, A. Different effects of chloroquine and hydroxychloroquine on lysosomal function in cultured retinal pigment epithelial cells. APMIS 2002, 110, 481–489. Available online: https://onlinelibrary.wiley.com/doi/full/10.1034/j.1600-0463.2002.100606.x (accessed on 24 November 2021). [CrossRef] [PubMed]
- Daniel, W.A.; Bickel, M.H.; Honegger, U.E. The Contribution of Lysosomal Trapping in the Uptake of Desipramine and Chloroquine by Different Tissues. Pharmacol. Toxicol. 1995, 77, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Navarro, M.; Goitia, H.; Silva, P.; Velásquez, M.; Ojeda, L.; Fraile, G. Synthesis and characterization of new copper– and zinc–chloroquine complexes and their activities on respiratory burst of polymorphonuclear leukocytes. J. Inorg. Biochem. 2005, 99, 1630–1636. [Google Scholar] [CrossRef]
- Navarro, M.; Hernández, C.; Vásquez, F.; Goitia, H.; Ojeda, L.E.; Velásquez, M.; Fraile, G. Syntheses, characterization, and biological evaluation of new zinc-and gold-chloroquine diphosphate complexes. Transit. Met. Chem. 2008, 33, 893–898. [Google Scholar] [CrossRef]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chemie Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Mori, M.; Kondo, T. Blue flower color development by anthocyanins: From chemical structure to cell physiology. Nat. Prod. Rep. 2009, 26, 884–915. Available online: https://pubs.rsc.org/en/content/articlehtml/2009/np/b800165k (accessed on 24 November 2021). [CrossRef] [PubMed]
- De Freitas, V.; Mateus, N. Chemical transformations of anthocyanins yielding a variety of colours (Review). Environ. Chem. Lett. 2006, 4, 175–183. Available online: http://link.springer.com/article/10.1007/s10311-006-0060-3 (accessed on 23 July 2021). [CrossRef]
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.; Boelens, P.G.; van Norren, K.; van Leeuwen, P.A. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. Available online: https://academic.oup.com/ajcn/article/74/4/418/4737352 (accessed on 4 August 2021). [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, 1–15. Available online: https://pmc/articles/PMC5465813/ (accessed on 4 August 2021). [CrossRef] [Green Version]
- Wei, Y.; Guo, M. Zinc-Binding Sites on Selected Flavonoids. Biol. Trace Element Res. 2014, 161, 223–230. Available online: https://link.springer.com/article/10.1007/s12011-014-0099-0 (accessed on 24 November 2021). [CrossRef] [PubMed]
- Primikyri, A.; Mazzone, G.; Lekka, C.; Tzakos, A.G.; Russo, N.; Gerothanassis, I.P. Understanding Zinc(II) Chelation with Quercetin and Luteolin: A Combined NMR and Theoretical Study. J. Phys. Chem. B 2014, 119, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Charley, P.J.; Sarkar, B.; Stitt, C.F.; Saltman, P. Chelation of iron by sugars. BBA–Biochim. Biophys. Acta 1963, 69, 313–321. [Google Scholar] [CrossRef]
- Christides, T.; Sharp, P.A. Sugars Increase Non-Heme Iron Bioavailability in Human Epithelial Intestinal and Liver Cells. PLoS ONE 2013, 8, e83031. Available online: http://pmc/articles/PMC3858368/ (accessed on 24 November 2021). [CrossRef] [PubMed] [Green Version]
- Gyurcsik, B.; Nagy, L. Carbohydrates as ligands: Coordination equilibria and structure of the metal complexes. Coord. Chem. Rev. 2000, 203, 81–149. [Google Scholar] [CrossRef]
- Allscher, T.; Klüfers, P.; Mayer, P. 4.7 Carbohydrate-Metal Complexes: Structural Chemistry of Stable Solution Species. In Glycoscience; Springer: Berlin/Heidelberg, Germany, 2008; p. 1077. ISBN 978-3-540-36154-1. [Google Scholar]
- Lim, J.H.; Jeong, Y.; Song, S.-H.; Ahn, J.-H.; Lee, J.R.; Lee, S.-M. Penetration of an antimicrobial zinc-sugar alcohol complex into Streptococcus mutans biofilms. Sci. Rep. 2018, 8, 16154. Available online: https://www.nature.com/articles/s41598-018-34366-y (accessed on 24 November 2021). [CrossRef] [PubMed]
- Morgan, A.J.; Lewis, G.; Hoven, W.V.D.; Akkerboom, P. The effect of zinc in the form of erythromycin-zinc complex (ZinerytR lotion) and zinc acetate on metallothionein expression and distribution in hamster skin. Br. J. Dermatol. 1993, 129, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.R.; James, D. Interaction of vitamin C and iron. Ann. N. Y. Acad. Sci. 1980, 355, 32–44. Available online: https://pubmed.ncbi.nlm.nih.gov/6940487/ (accessed on 5 August 2021). [CrossRef] [Green Version]
- Teucher, B.; Olivares, M.; Cori, H. Enhancers of iron absorption: Ascorbic acid and other organic acids. Int. J. Vitam. Nutr. Res. 2004, 74, 403–419. Available online: https://pubmed.ncbi.nlm.nih.gov/15743017/ (accessed on 5 August 2021). [CrossRef] [PubMed]
- Ünaleroǧlu, C.; Zümreoǧlu-Karan, B.; Mert, Y. Zinc ascorbate: A combined experimental and computational study for structure elucidation. J. Mol. Struct. 2002, 605, 227–233. [Google Scholar] [CrossRef]
- Ragsdale, N.N.; Sisler, H.D. Inhibition of ergosterol synthesis in Ustilago maydis by the fungicide triarimol. Biochem. Biophys. Res. Commun. 1972, 46, 2048–2053. [Google Scholar] [CrossRef]
- Borgers, M. Mechanism of Action of Antifungal Drugs, with Special Reference to the Imidazole Derivatives. Clin. Infect. Dis. 1980, 2, 520–534. Available online: http://cid.oxfordjournals.org/ (accessed on 24 November 2021). [CrossRef] [PubMed]
- Verras, A.; de Montellano, P.O. Protein dynamics and imidazole binding in cytochrome P450 enzymes. Biochem. Soc. Trans. 2006, 34, 1170–1172. [Google Scholar] [CrossRef] [Green Version]
- Sundberg, R.J.; Martin, R.B. Interactions of histidine and other imidazole derivatives with transition metal ions in chemical and biological systems. Chem. Rev. 1974, 74, 471–517. [Google Scholar] [CrossRef]
- López-Sandoval, H.; Lemos, M.E.L.; Garza-Velasco, R.; Poblano-Meléndez, I.; Granada-Macías, P.; Gracia-Mora, I.; Barba-Behrens, N. Synthesis, structure and biological activities of cobalt(II) and zinc(II) coordination compounds with 2-benzimidazole derivatives. J. Inorg. Biochem. 2008, 102, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, D. Synthesis, Characterisation and Biological Activity of Novel Carboxylate Complexes Incorporating Phenanthroline and Benzimidazole Ligands 2004. Available online: https://arrow.tudublin.ie/tourdoc (accessed on 4 August 2021).
- Mothilal, K.; Karunakaran, C.; Rajendran, A.; Murugesan, R. Synthesis, X-ray crystal structure, antimicrobial activity and photodynamic effects of some thiabendazole complexes. J. Inorg. Biochem. 2004, 98, 322–332. [Google Scholar] [CrossRef]
- Seidel, R.W.; Schulze, A.C.; Oppel, I.M. 1D Coordination Polymers from Zinc(II) and Cadmium(II) Halides and 2,2′-Dithiobis(pyridine N-oxide): Isostructurality and Structural Diversity. J. Inorg. Gen. Chem. 2017, 643, 317–324. Available online: https://onlinelibrary.wiley.com/doi/full/10.1002/zaac.201600417 (accessed on 24 November 2021). [CrossRef]
- Rush, A.K. Improving the Mechanistic Understanding of Zinc Pyrithione Bioavailability in Skin through Lateral and Transverse Diffusion Measurements. 2015. Available online: http://rave.ohiolink.edu/etdc/view?acc_num=ucin1439281238 (accessed on 24 June 2021).
- Oyama, T.M.; Saito, M.; Yonezawa, T.; Okano, Y.; Oyama, Y. Nanomolar concentrations of zinc pyrithione increase cell susceptibility to oxidative stress induced by hydrogen peroxide in rat thymocytes. Chemosphere 2012, 87, 1316–1322. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
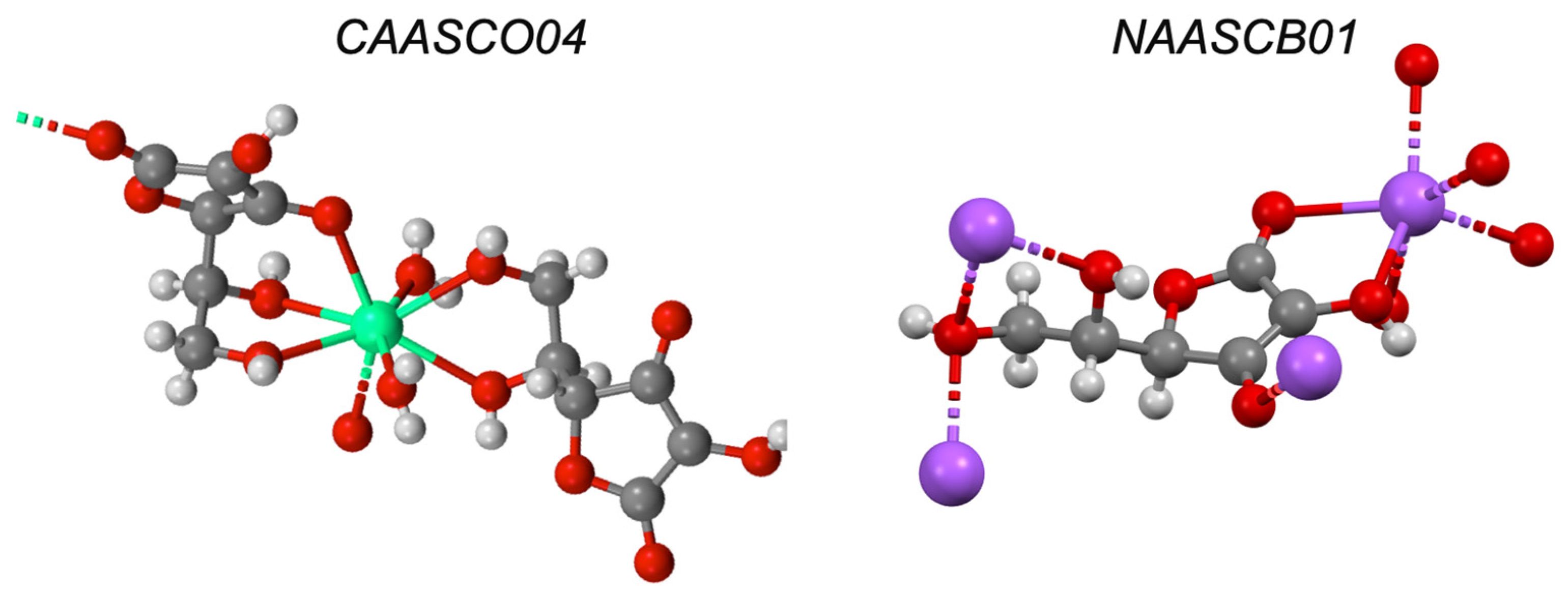
| Drug | Log P7.4 a | BCS | Solid-State Complex b | Solution Complex | Liposomal Ionophorism | |
|---|---|---|---|---|---|---|
| 0.1 mM | 1 mM | |||||
| Quinolines | ||||||
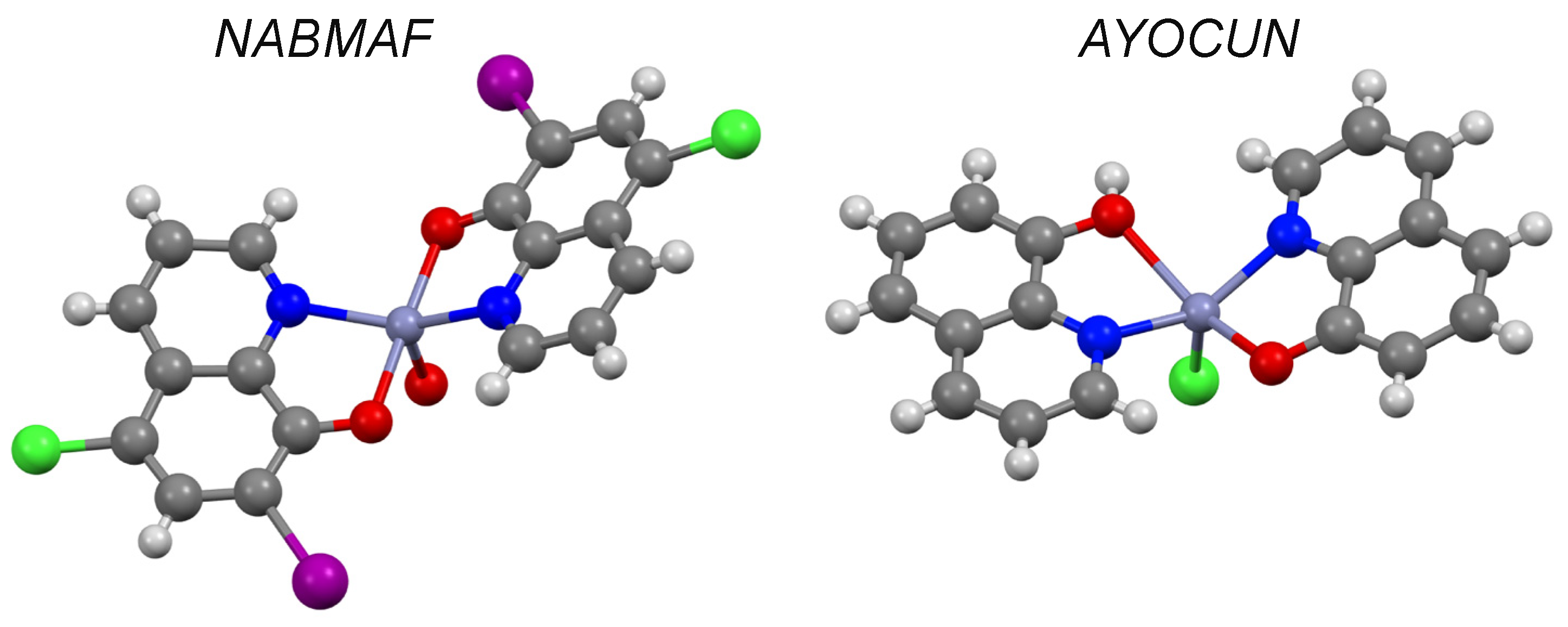
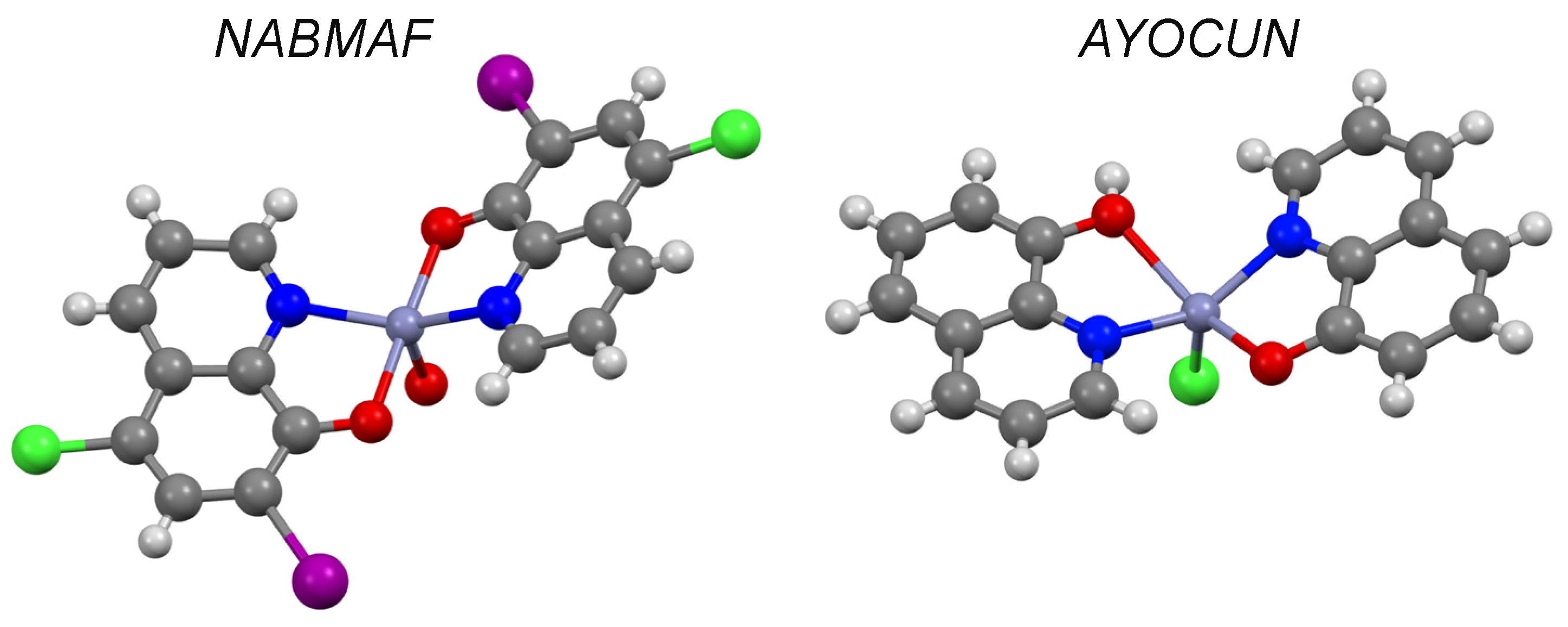
| 8-hydroxyquinoline d | 1.82 | I | AYOCUN | ✓ [37] | - | ✓ |
| Clioquinol d | 3.03 | II | NABMAF | ✓ | ✓ | |
| Hydroxychloroquine d | 0.33 | I | - | - | - | - |
| Polyols | ||||||
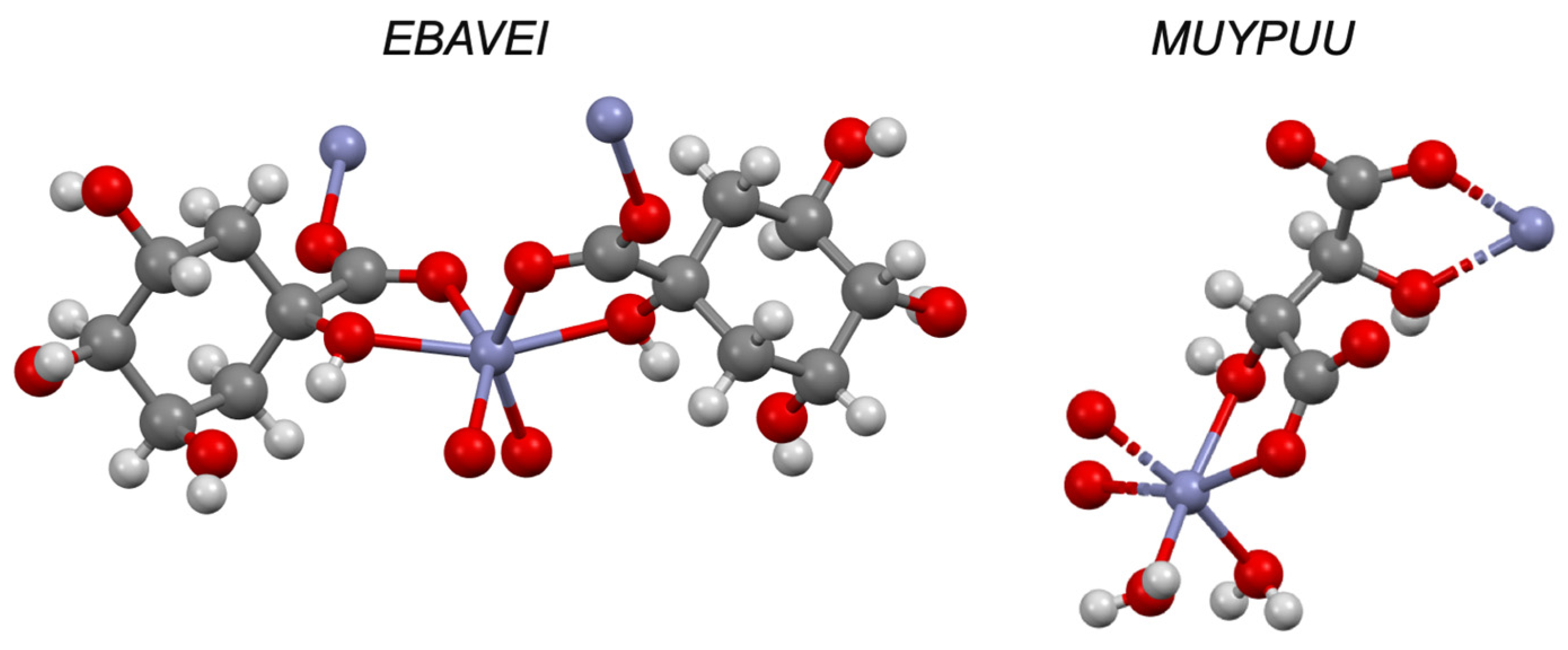
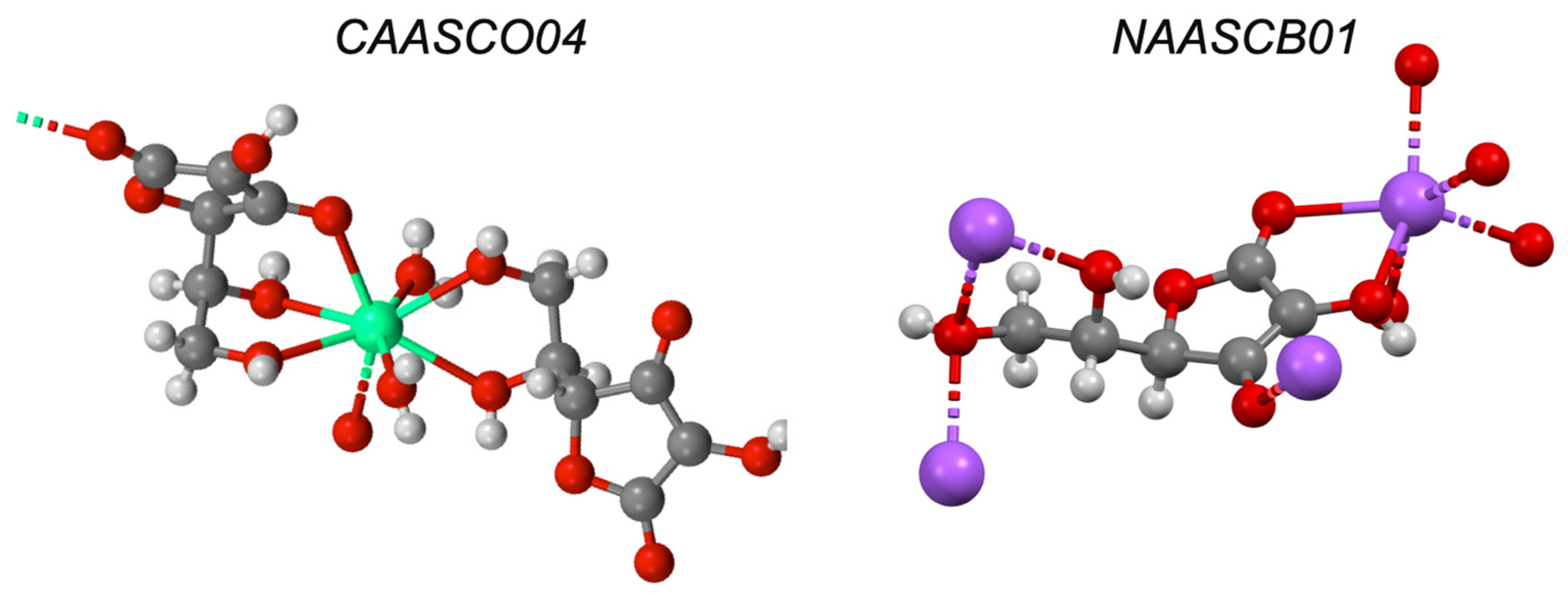
| Ascorbic acid e | −5.00 | I | ✓ [38] | ✓ [39] | ✓ | |
| Erythromycin d | 0.99 | III | BOPRON10 | ✓ [40] | - | - |
| Zinc gluconate e | - | I | ✓ [41] | ✓ [41] | - | ✓ |
| Flavonoids | ||||||
| Naringenin e | 2.70 | II | ✓ [42] | ✓ [43] | - | ✓ |
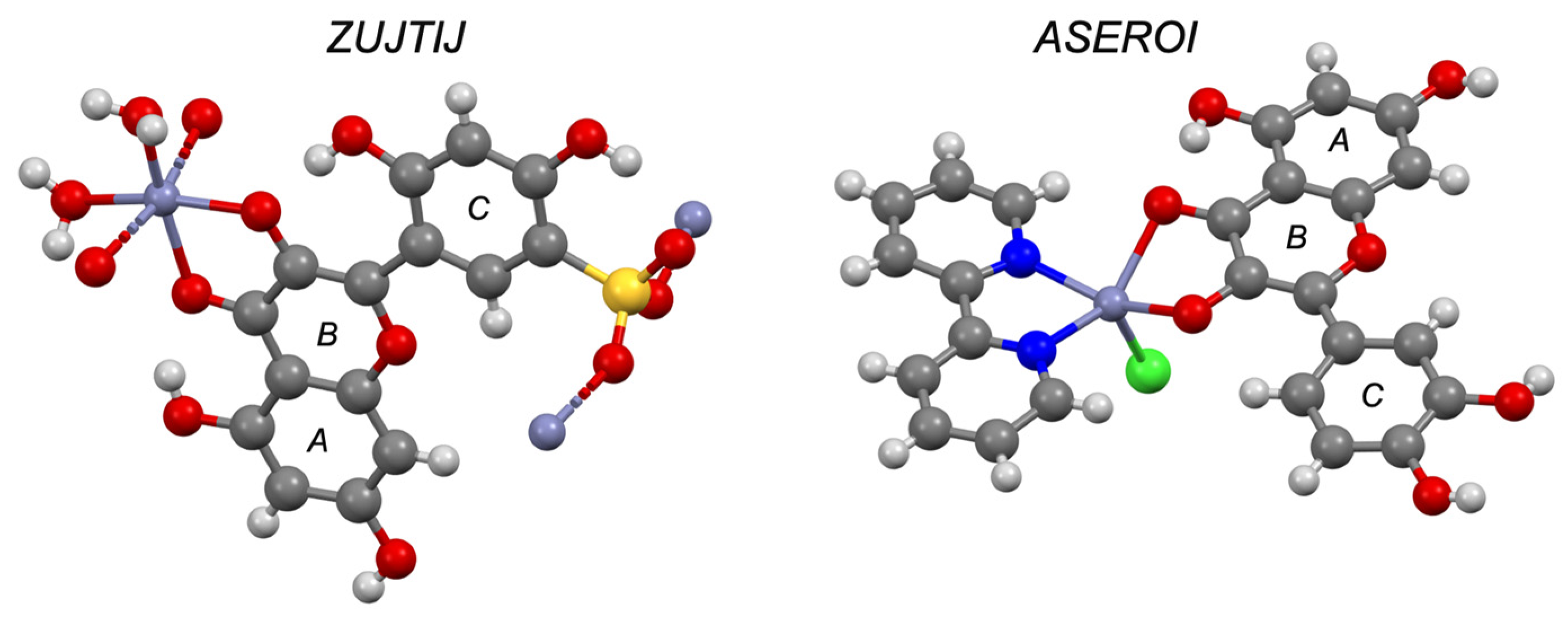
| Quercetin e | 1 | II/IV | ASEROI | ✓ [44] | ✓ | |
| Imidazoles | ||||||
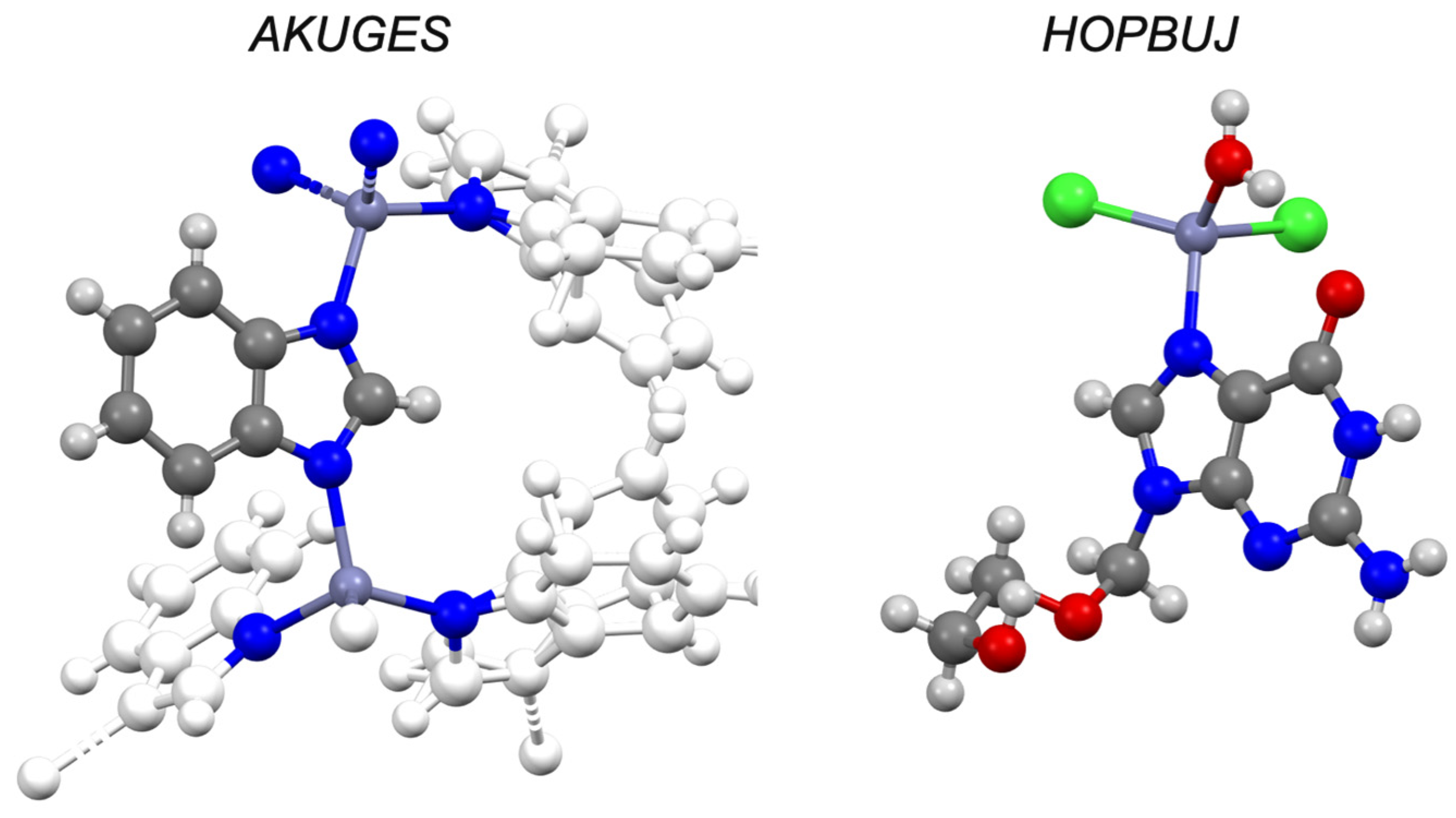
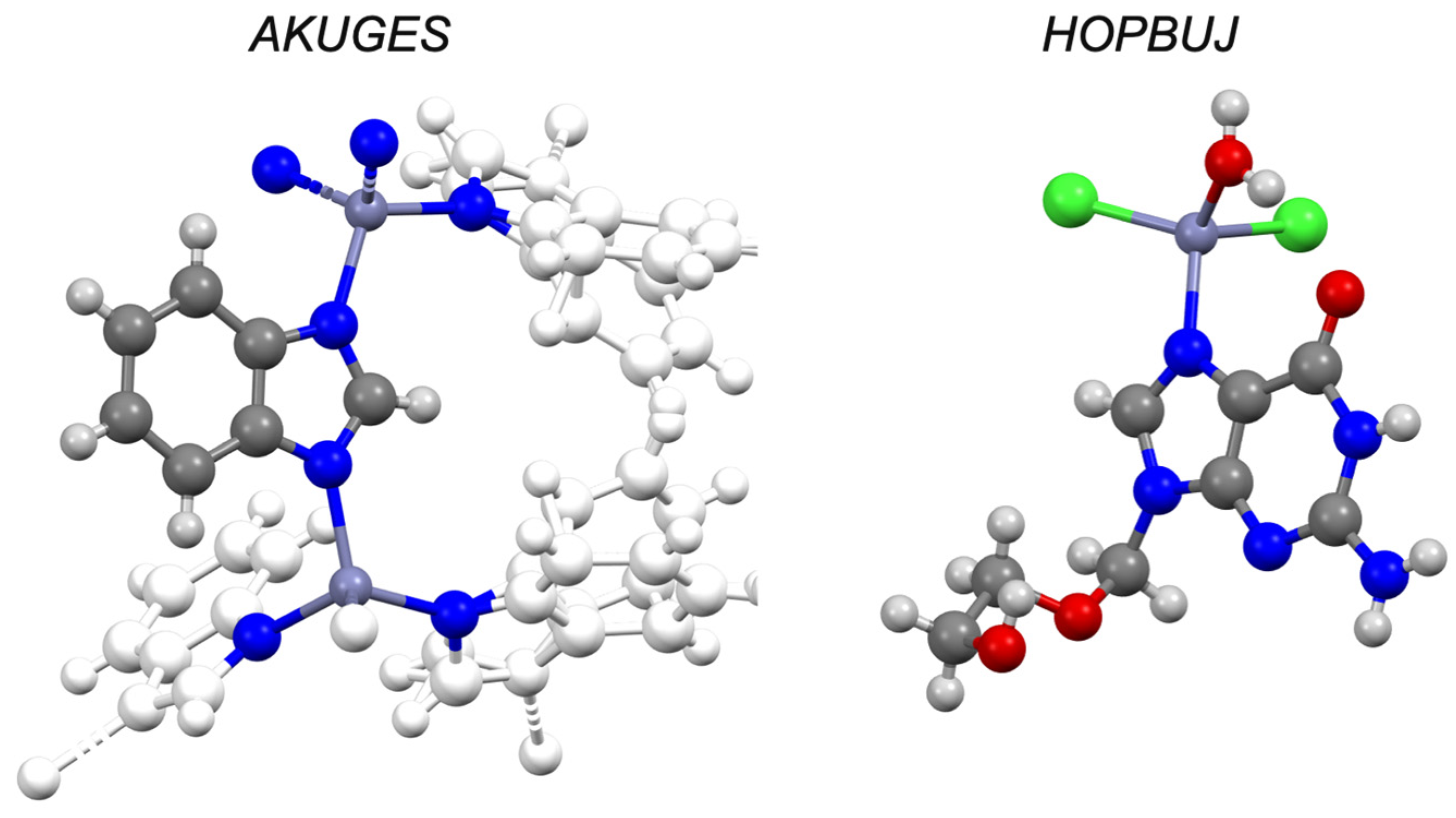
| Aciclovir d | −1.03 | IV | HOPBUJ | ✓ [45] | - | ✓ |
| Mebendazole d | 3.25 | II | - | ✓ c [46] | - | ✓ |
| Caffeine d | −0.55 | I | RITLEO | ✓ [47] | - | ✓ |
| Levamisole d | 2.22 | I | ✓ [48] | ✓ [48] | - | - |
| Amino acids | ||||||
| Carnosine d | −4.51 | I | ✓ [49] | ✓ [50] | - | - |
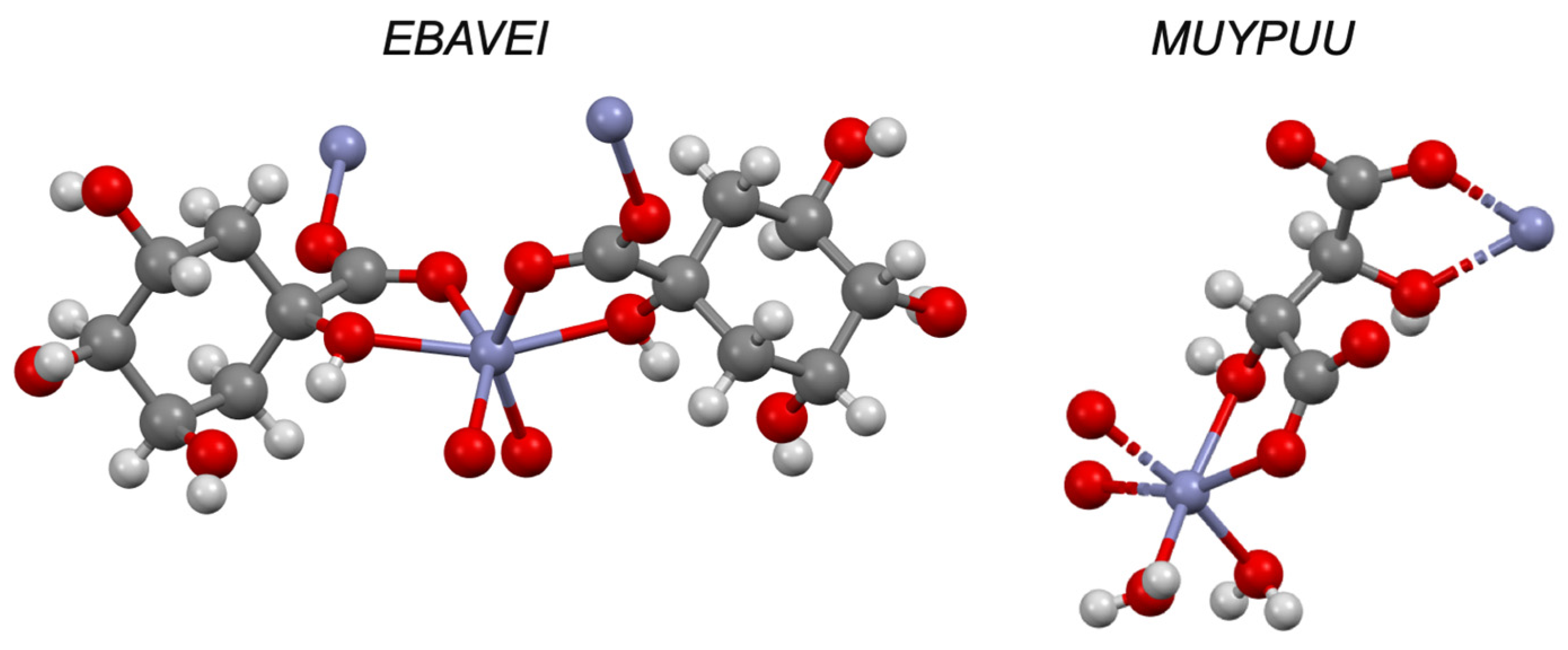
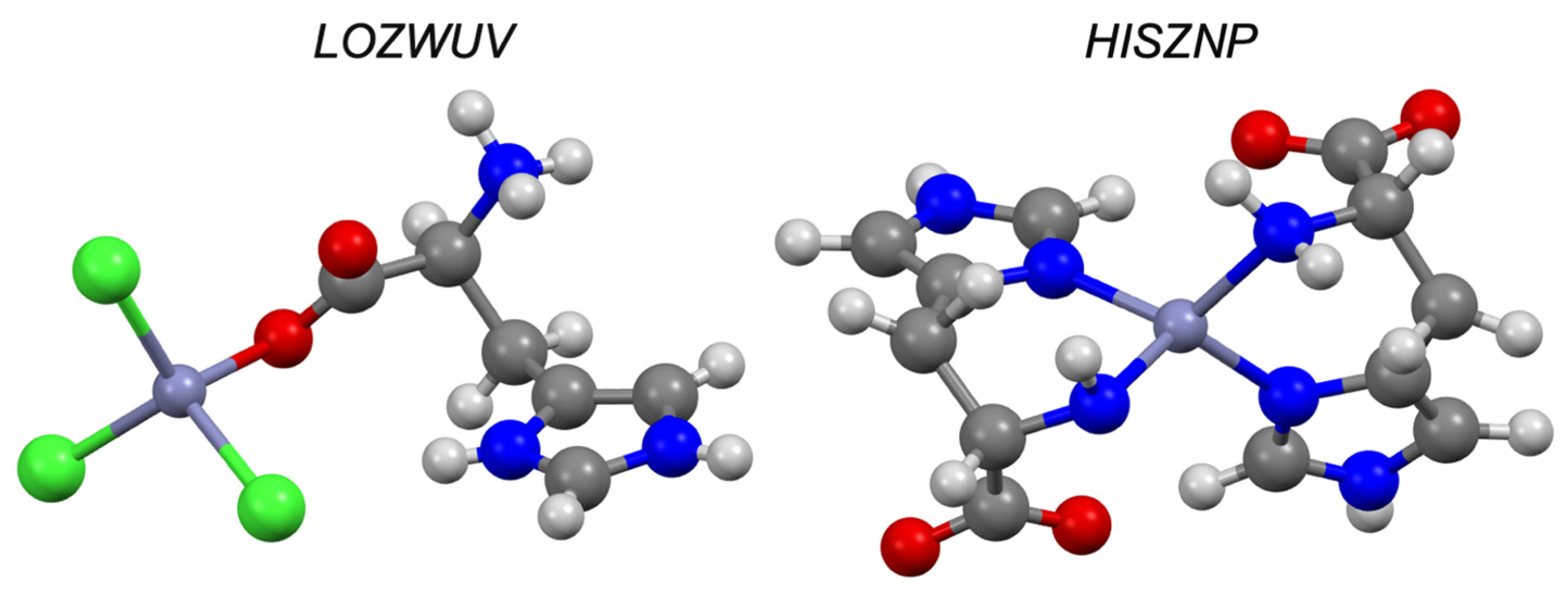
| Cysteine e | −2.80 | I | CURLUW | ✓ [51] | ✓ | |
| Histidine e | −3.64 | I | MUYFEU | ✓ [51] | ✓ | |
| Methionine e | −2.19 | I | LMETZN01 | ✓ [52] | - | - |
| Proline e | −2.57 | I | HIBTOB01 | ✓ [53] | - | - |
| Tryptophan e | −1.09 | I | ✓ [54] | ✓ [55] | - | ✓ |
| Miscellaneous | ||||||
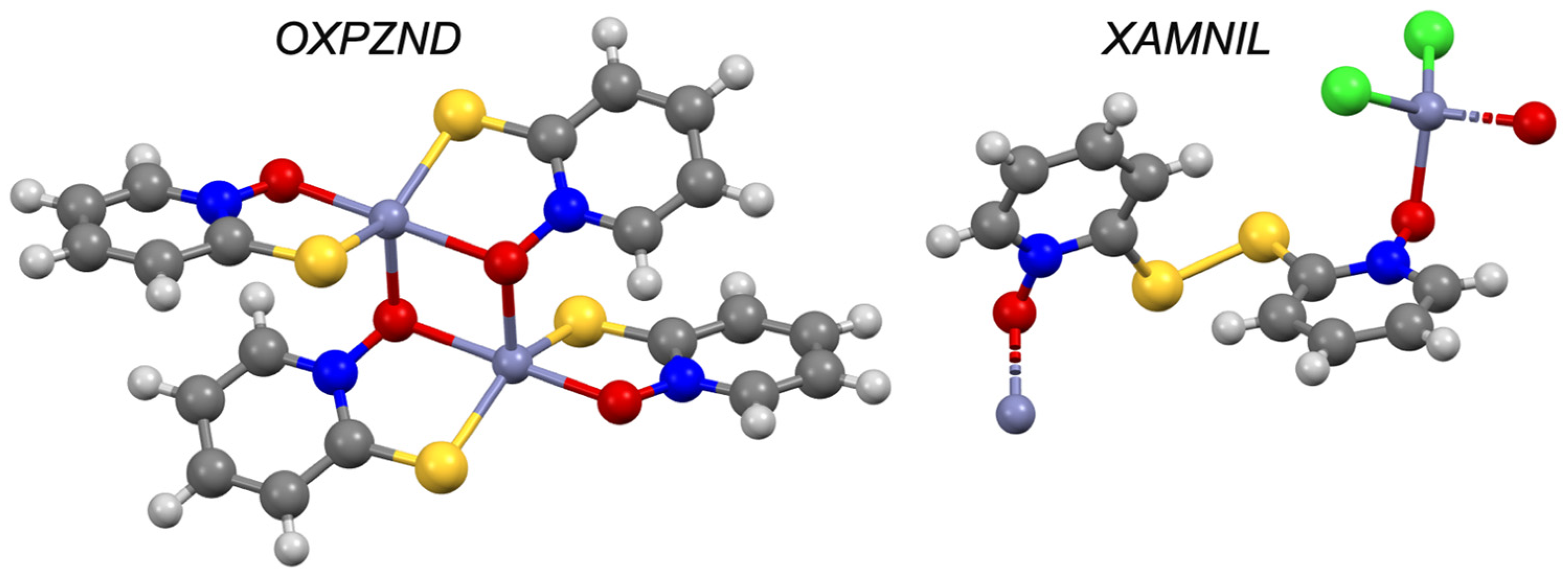
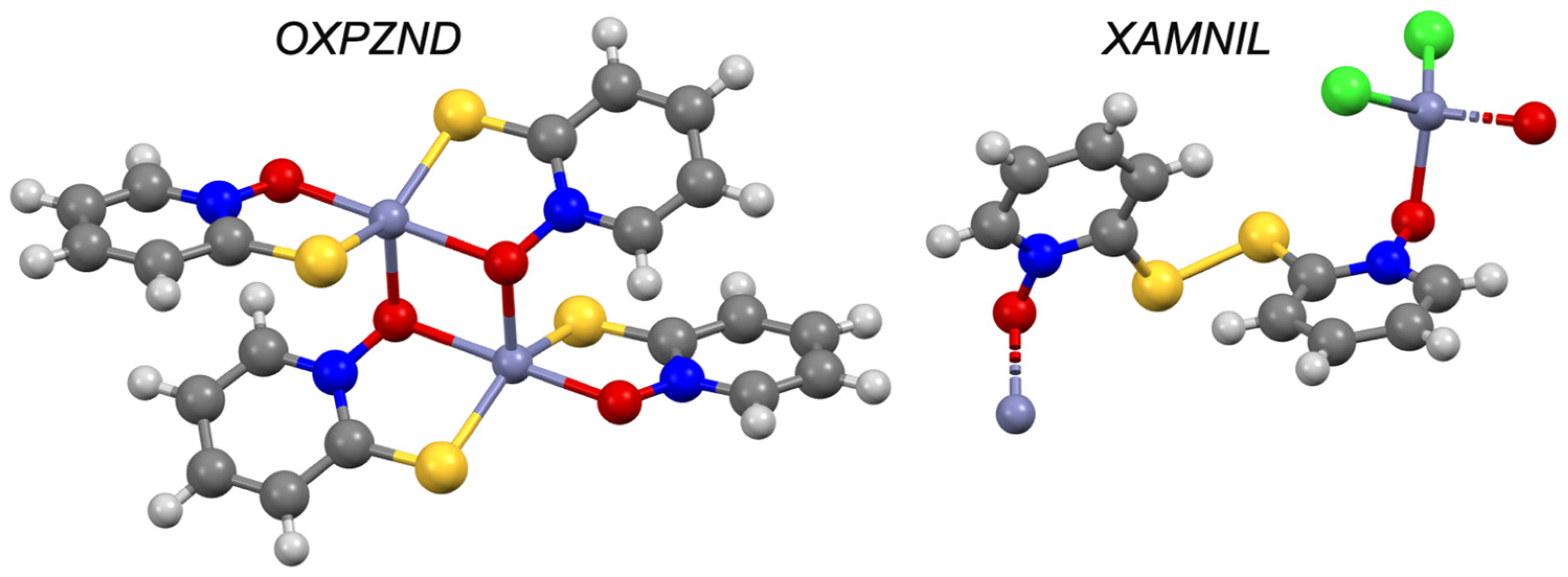
| Pyrithione d | −0.41 | III | OXPZND [56,57] | ✓ [8] | ✓ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kavanagh, O.; Elmes, R.; O’Sullivan, F.; Farragher, J.; Robinson, S.; Walker, G. Investigating Structural Property Relationships to Enable Repurposing of Pharmaceuticals as Zinc Ionophores. Pharmaceutics 2021, 13, 2032. https://doi.org/10.3390/pharmaceutics13122032
Kavanagh O, Elmes R, O’Sullivan F, Farragher J, Robinson S, Walker G. Investigating Structural Property Relationships to Enable Repurposing of Pharmaceuticals as Zinc Ionophores. Pharmaceutics. 2021; 13(12):2032. https://doi.org/10.3390/pharmaceutics13122032
Chicago/Turabian StyleKavanagh, Oisín, Robert Elmes, Finbarr O’Sullivan, John Farragher, Shane Robinson, and Gavin Walker. 2021. "Investigating Structural Property Relationships to Enable Repurposing of Pharmaceuticals as Zinc Ionophores" Pharmaceutics 13, no. 12: 2032. https://doi.org/10.3390/pharmaceutics13122032
APA StyleKavanagh, O., Elmes, R., O’Sullivan, F., Farragher, J., Robinson, S., & Walker, G. (2021). Investigating Structural Property Relationships to Enable Repurposing of Pharmaceuticals as Zinc Ionophores. Pharmaceutics, 13(12), 2032. https://doi.org/10.3390/pharmaceutics13122032