
Egr1 Gene Expression as a Potential Biomarker for In Vitro Prediction of Ocular Toxicity



 and
and 
Abstract
:
1. Introduction
2. Materials and Methods
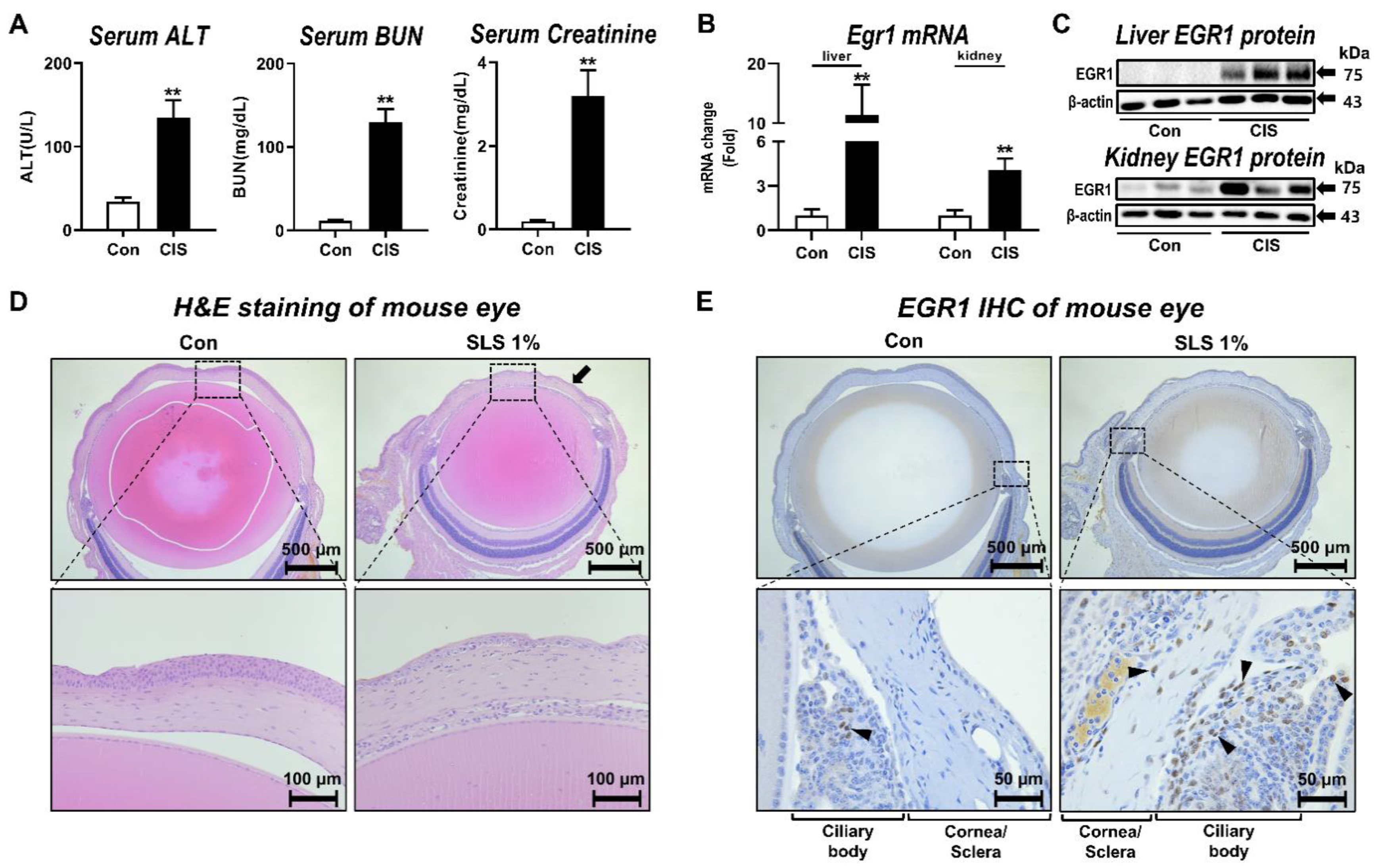
2.1. Animal Experiment
2.2. Serum Collection and Biochemical Analysis
2.3. Histology and Immunohistochemistry
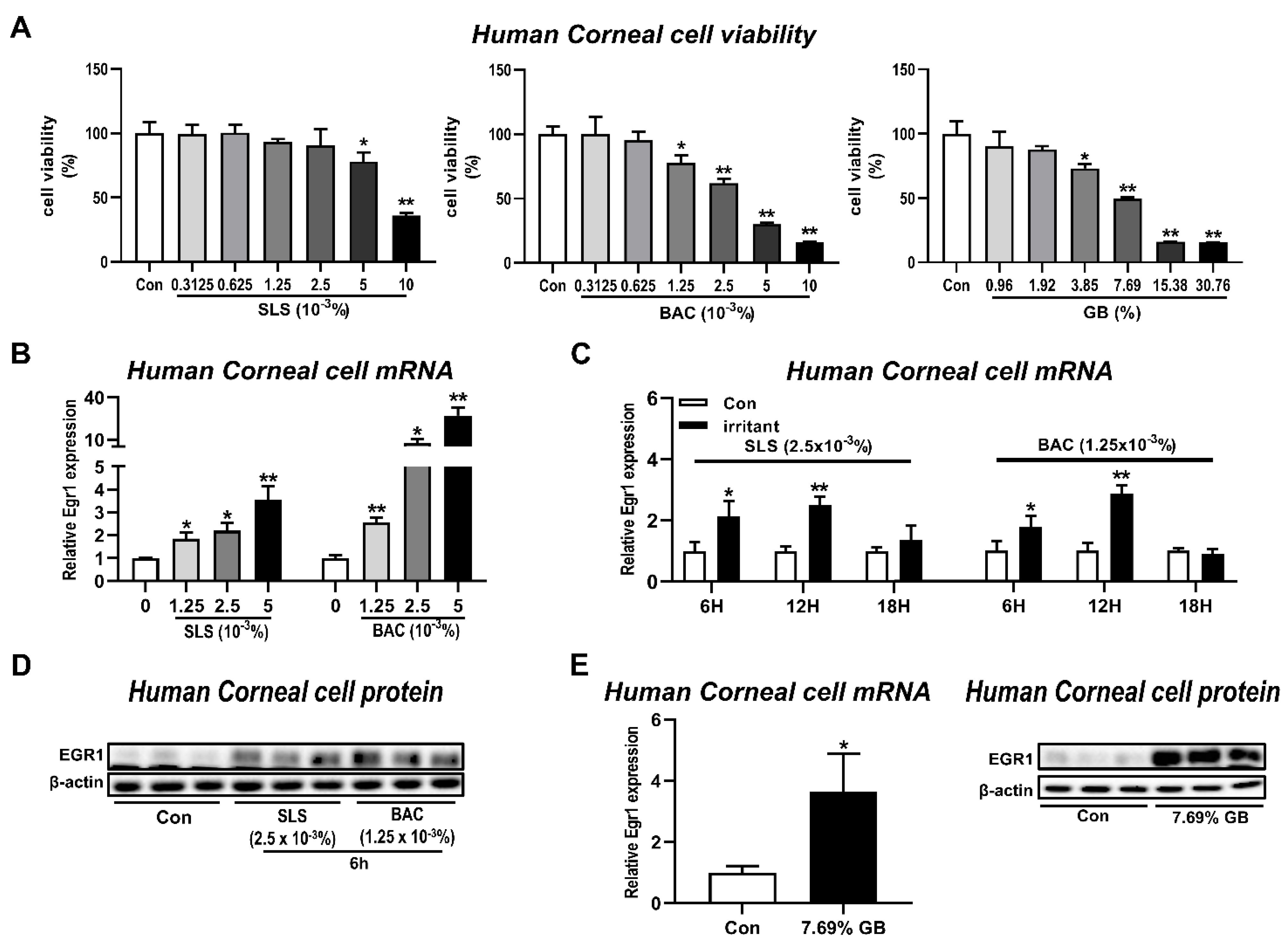
2.4. Cell Culture and Treatments
2.5. Cell Viability Assay
2.6. RNA Extraction, cDNA Synthesis, and Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.7. Droplet Digital PCR (ddPCR)
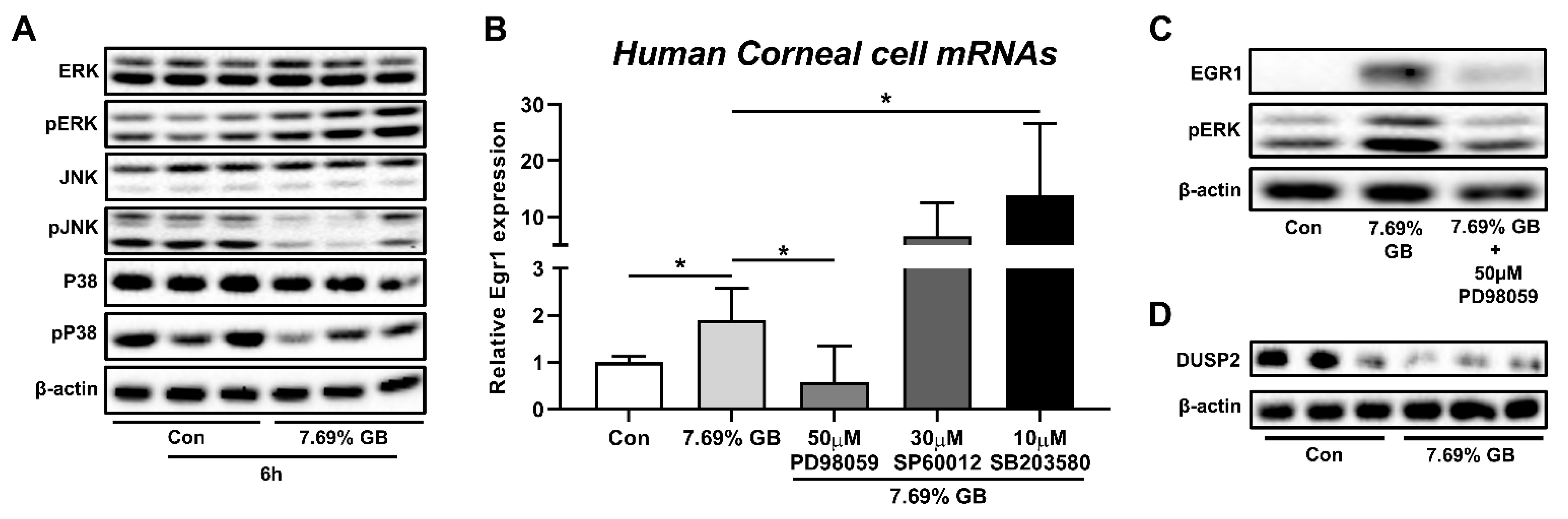
2.8. Western Blot
2.9. Putative Transcription Factor Binding Site in the Egr1 Promoter
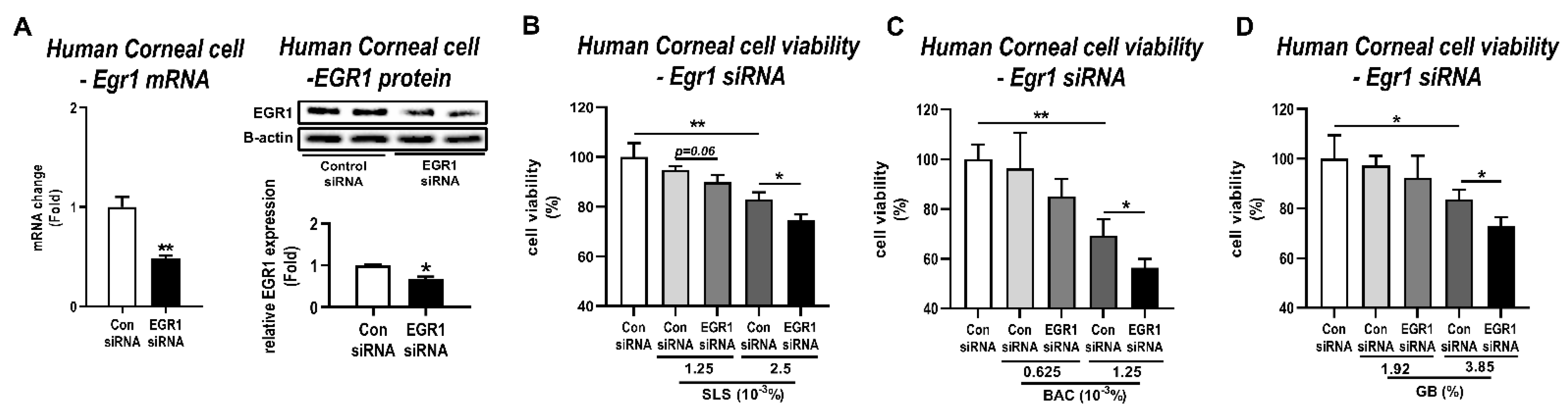
2.10. Egr1-Small Interfering RNA (siRNA) Silencing
2.11. Egr1 Promoter–Luciferase Gene Reporter Assay
2.12. Statistics
3. Results
3.1. Egr1 Expression Alterations Associated with Multiorgan Injury In Vivo
3.2. In Vitro Egr1 Expression Alterations in Human Corneal Epithelial Cells by Treatment with Various Irritants
3.3. Involvement of ERK Pathway in the Elevation of EGR1 Expression after Irritation Exposure
3.4. Effects of Egr1 Knockdown in Human Corneal Epithelial Cells Treated with Irritants
3.5. Induction of Egr1 Promoter Activity by Various Irritants
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bailey, J.; Thew, M.; Balls, M. An analysis of the use of animal models in predicting human toxicology and drug safety. Altern. Lab. Anim. 2014, 42, 181–199. [Google Scholar] [CrossRef] [Green Version]
- Choksi, N.Y.; Truax, J.; Layton, A.; Matheson, J.; Mattie, D.; Varney, T.; Tao, J.; Yozzo, K.; McDougal, A.J.; Merrill, J.; et al. United States regulatory requirements for skin and eye irritation testing. Cutan. Ocul. Toxicol. 2019, 38, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Draize, J.H. Methods for the study of irritation and toxicity of substances applied topically to the skin and mucous membranes. J. Pharmacol. Exp. Ther. 1944, 82, 377–390. [Google Scholar]
- Guideline, O.T. 405, Acute Eye Irritation/Corrosion; Organization for Economic Co-Operation and Development: Paris, France, 2002. [Google Scholar]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; Methuen: London, UK, 1959. [Google Scholar]
- Vinardell, M.P.; Mitjans, M. Alternative methods for eye and skin irritation tests: An overview. J. Pharm. Sci. 2008, 97, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Luepke, N.; Kemper, F. The HET-CAM test: An alternative to the Draize eye test. Food Chem. Toxicol. 1986, 24, 495–496. [Google Scholar] [CrossRef]
- Luepke, N.P. Hen’s egg chorioallantoic membrane test for irritation potential. Food Chem. Toxicol. 1985, 23, 287–291. [Google Scholar] [CrossRef]
- OECD. Test No. 437: Bovine corneal opacity and permeability test method for identifying i) chemicals inducing serious eye damage and ii) chemicals not requiring classification for eye irritation or serious eye damage. In OECD Guidelines for the Testing of Chemicals; OECD Publishing: Paris, France, 2017. [Google Scholar]
- OECD. Test No. 438: Isolated Chicken Eye Test Method for Identifying i) Chemicals Inducing Serious Eye Damage and ii) Chemicals Not Requiring Classification for Eye Irritation or Serious Eye Damage. In OECD Guidelines for the Testing of Chemicals; OECD Publishing: Paris, France, 2013. [Google Scholar]
- Matsuda, S.; Hisama, M.; Shibayama, H.; Itou, N.; Iwaki, M. Application of the reconstructed rabbit corneal epithelium model to assess the in vitro eye irritancy test of chemicals. Yakugaku Zasshi 2009, 129, 1113–1120. [Google Scholar] [CrossRef] [Green Version]
- Kaluzhny, Y.; Kandarova, H.; Handa, Y.; DeLuca, J.; Truong, T.; Hunter, A.; Kearney, P.; D’Argembeau-Thornton, L.; Klausner, M. The EpiOcular Eye Irritation Test (EIT) for hazard identification and labelling of eye irritating chemicals: Protocol optimisation for solid materials and the results after extended shipment. Altern. Lab. Anim. 2015, 43, 101–127. [Google Scholar] [CrossRef]
- Pfannenbecker, U.; Bessou-Touya, S.; Faller, C.; Harbell, J.; Jacob, T.; Raabe, H.; Tailhardat, M.; Alepee, N.; De Smedt, A.; De Wever, B.; et al. Cosmetics Europe multi-laboratory pre-validation of the EpiOcular reconstituted human tissue test method for the prediction of eye irritation. Toxicol. Vitr. 2013, 27, 619–626. [Google Scholar] [CrossRef]
- Jang, W.-h.; Jung, K.-m.; Yang, H.-r.; Lee, M.; Jung, H.-S.; Lee, S.-H.; Park, M.; Lim, K.-M. Evaluation of eye irritation potential of solid substance with new 3D reconstructed human cornea model, MCTT HCETM. Biomol. Ther. 2015, 23, 379. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.-M.; Lee, S.-H.; Ryu, Y.-H.; Jang, W.-H.; Jung, H.-S.; Han, J.-H.; Seok, S.-H.; Park, J.-H.; Son, Y.; Park, Y.-H. A new 3D reconstituted human corneal epithelium model as an alternative method for the eye irritation test. Toxicol. Vitr. 2011, 25, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Hamajima, F.; Ogasawara, T.; Hata, K.-i. Establishment of a new in vitro test method for evaluation of eye irritancy using a reconstructed human corneal epithelial model, LabCyte CORNEA-MODEL. Toxicol. Vitr. 2013, 27, 2184–2192. [Google Scholar] [CrossRef] [PubMed]
- Kaluzhny, Y.; Kandárová, H.; Hayden, P.; Kubilus, J.; D’Argembeau-Thornton, L.; Klausner, M. Development of the EpiOcular™ eye irritation test for hazard identification and labelling of eye irritating chemicals in response to the requirements of the EU cosmetics directive and REACH legislation. Altern. Lab. Anim. 2011, 39, 339–364. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Kim, D.-e.; Jang, W.-H.; An, S.; Cho, S.-A.; Jung, M.-S.; Lee, J.E.; Yeo, K.-W.; Koh, S.B.; Jeong, T.-C. Prevalidation trial for a novel in vitro eye irritation test using the reconstructed human cornea-like epithelial model, MCTT HCE™. Toxicol. Vitr. 2017, 39, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Cottrez, F.; Boitel, E.; Ourlin, J.C.; Peiffer, J.L.; Fabre, I.; Henaoui, I.S.; Mari, B.; Vallauri, A.; Paquet, A.; Barbry, P.; et al. SENS-IS, a 3D reconstituted epidermis based model for quantifying chemical sensitization potency: Reproducibility and predictivity results from an inter-laboratory study. Toxicol. Vitr. 2016, 32, 248–260. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, S.; Meng, T.; Zhang, L.; Dai, T.; Xiang, Q.; Su, Z.; Zhang, Q.; Huang, Y. HSP27 as a biomarker for predicting skin irritation in human skin and reconstructed organotypic skin model. Toxicol. Lett. 2014, 226, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Coquette, A.; Berna, N.; Vandenbosch, A.; Rosdy, M.; De Wever, B.; Poumay, Y. Analysis of interleukin-1alpha (IL-1alpha) and interleukin-8 (IL-8) expression and release in in vitro reconstructed human epidermis for the prediction of in vivo skin irritation and/or sensitization. Toxicol. Vitr. 2003, 17, 311–321. [Google Scholar] [CrossRef]
- Braunstein, S.G.; Deramaudt, T.G.; Rosenblum, D.G.; Dunn, M.G.; Abraham, N.G. Heme oxygenase-1 gene expression as a stress index to ocular irritation. Curr. Eye Res. 1999, 19, 115–122. [Google Scholar] [CrossRef]
- Choi, S.; Lee, M.; Lee, S.-H.; Jung, H.-S.; Kim, S.-Y.; Chung, T.-Y.; Choe, T.-b.; Chun, Y.-J.; Lim, K.-M. Identification of cornifelin and early growth response-1 gene as novel biomarkers for in vitro eye irritation using a 3D reconstructed human cornea model MCTT HCE™. Arch. Toxicol. 2015, 89, 1589–1598. [Google Scholar] [CrossRef]
- Lin, Z.; Liu, X.; Zhou, T.; Wang, Y.; Bai, L.; He, H.; Liu, Z. A mouse dry eye model induced by topical administration of benzalkonium chloride. Mol. Vis. 2011, 17, 257–264. [Google Scholar]
- Pauloin, T.; Dutot, M.; Liang, H.; Chavinier, E.; Warnet, J.M.; Rat, P. Corneal protection with high-molecular-weight hyaluronan against in vitro and in vivo sodium lauryl sulfate-induced toxic effects. Cornea 2009, 28, 1032–1041. [Google Scholar] [CrossRef]
- Hwang, D.B.; Won, D.H.; Shin, Y.S.; Kim, S.Y.; Kang, B.C.; Lim, K.M.; Che, J.H.; Nam, K.T.; Yun, J.W. Ccrn4l as a pre-dose marker for prediction of cisplatin-induced hepatotoxicity susceptibility. Free Radic. Biol. Med. 2020, 148, 128–139. [Google Scholar] [CrossRef]
- Hwang, D.-B.; Cha, M.H.; Won, D.-H.; Shin, Y.-S.; Kim, S.-Y.; Kim, C.; Lee, E.-J.; Kim, Y.Y.; Yun, J.-W. Transcriptomic analysis of rat kidney reveals a potential mechanism of sex differences in susceptibility to cisplatin-induced nephrotoxicity. Free Radic. Biol. Med. 2021, 174, 100–109. [Google Scholar] [CrossRef]
- Ye, D.-J.; Kwon, Y.-J.; Baek, H.-S.; Shin, S.; Lee, C.; Yun, J.-W.; Nam, K.T.; Lim, K.-M.; Chun, Y.-J. Discovery of ezrin expression as a potential biomarker for chemically induced ocular irritation using human corneal epithelium cell line and a reconstructed human cornea-like epithelium model. Toxicol. Sci. 2018, 165, 335–346. [Google Scholar] [CrossRef]
- UN. Globally Harmonized System of Classification and Labelling of Chemicals (GHS); United Nations: New York, NY, USA; Geneva, Switzerland, 2011. [Google Scholar]
- Sastry, J.; Kellie, S.J. Severe neurotoxicity, ototoxicity and nephrotoxicity following high-dose cisplatin and amifostine. Pediatr. Hematol. Oncol. 2005, 22, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Urbschat, A.; Obermuller, N.; Haferkamp, A. Biomarkers of kidney injury. Biomarkers 2011, 16 (Suppl. 1), S22–S30. [Google Scholar] [CrossRef]
- Church, R.J.; Watkins, P.B. The transformation in biomarker detection and management of drug-induced liver injury. Liver Int. 2017, 37, 1582–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.M. Regulation of radiation-induced apoptosis by early growth response-1 gene in solid tumors. Curr. Cancer Drug Targets 2004, 4, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiel, G.; Mayer, S.I.; Muller, I.; Stefano, L.; Rossler, O.G. Egr-1-A Ca2+-regulated transcription factor. Cell Calcium 2010, 47, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Shao, Y.; Lai, S.S.; Qiao, L.; Yang, R.L.; Xue, B.; Pan, F.Y.; Chen, H.Q.; Li, C.J. GGPPS, a new EGR-1 target gene, reactivates ERK 1/2 signaling through increasing Ras prenylation. Am. J. Pathol. 2011, 179, 2740–2750. [Google Scholar] [CrossRef] [PubMed]
- Fey, D.; Croucher, D.R.; Kolch, W.; Kholodenko, B.N. Crosstalk and signaling switches in mitogen-activated protein kinase cascades. Front. Physiol. 2012, 3, 355. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Wu, X.; Chen, Q.; Zhu, S.; Liu, Y.; Pan, D.; Chen, X.; Li, D. The anti-apoptotic and cardioprotective effects of salvianolic acid a on rat cardiomyocytes following ischemia/reperfusion by DUSP-mediated regulation of the ERK1/2/JNK pathway. PLoS ONE 2014, 9, e102292. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, K.; Nishida, E. Regulation of MAP kinases by MAP kinase phosphatases. Biochim. Biophys. Acta 2007, 1773, 1227–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, S.; Yuan, J.; Zhao, D.; Shen, N.; Chen, W.; Ding, Y.; Yu, D.; Li, J.; Pan, F.; Zhu, M.; et al. Regulation of mice liver regeneration by early growth response-1 through the GGPPS/RAS/MAPK pathway. Int. J. Biochem. Cell Biol. 2015, 64, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Guerquin, M.J.; Charvet, B.; Nourissat, G.; Havis, E.; Ronsin, O.; Bonnin, M.A.; Ruggiu, M.; Olivera-Martinez, I.; Robert, N.; Lu, Y.; et al. Transcription factor EGR1 directs tendon differentiation and promotes tendon repair. J. Clin. Investig. 2013, 123, 3564–3576. [Google Scholar] [CrossRef] [Green Version]
- Bai, Q.; Yan, H.; Sheng, Y.; Jin, Y.; Shi, L.; Ji, L.; Wang, Z. Long-term acetaminophen treatment induced liver fibrosis in mice and the involvement of Egr-1. Toxicology 2017, 382, 47–58. [Google Scholar] [CrossRef]
- Liu, C.; Calogero, A.; Ragona, G.; Adamson, E.; Mercola, D. EGR-1, the reluctant suppression factor: EGR-1 is known to function in the regulation of growth, differentiation, and also has significant tumor suppressor activity and a mechanism involving the induction of TGF-beta1 is postulated to account for this suppressor activity. Crit. Rev. Oncog. 1996, 7, 101–125. [Google Scholar]
- Thiel, G.; Cibelli, G. Regulation of life and death by the zinc finger transcription factor Egr-1. J. Cell. Physiol. 2002, 193, 287–292. [Google Scholar] [CrossRef]
- Cabodi, S.; Morello, V.; Masi, A.; Cicchi, R.; Broggio, C.; Distefano, P.; Brunelli, E.; Silengo, L.; Pavone, F.; Arcangeli, A. Convergence of integrins and EGF receptor signaling via PI3K/Akt/FoxO pathway in early gene Egr-1 expression. J. Cell. Physiol. 2009, 218, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Junttila, M.R.; Li, S.P.; Westermarck, J. Phosphatase-mediated crosstalk between MAPK signaling pathways in the regulation of cell survival. FASEB J. 2008, 22, 954–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowan, K.J.; Storey, K.B. Mitogen-activated protein kinases: New signaling pathways functioning in cellular responses to environmental stress. J. Exp. Biol. 2003, 206, 1107–1115. [Google Scholar] [CrossRef] [Green Version]
- Kohno, M.; Pouyssegur, J. Targeting the ERK signaling pathway in cancer therapy. Ann. Med. 2006, 38, 200–211. [Google Scholar] [CrossRef]
- Liu, Q.; Hofmann, P.A. Protein phosphatase 2A-mediated cross-talk between p38 MAPK and ERK in apoptosis of cardiac myocytes. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H2204–H2212. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Hofmann, P.A. Modulation of protein phosphatase 2a by adenosine A1 receptors in cardiomyocytes: Role for p38 MAPK. Am. J. Physiol Heart Circ. Physiol. 2003, 285, H97–H103. [Google Scholar] [CrossRef] [Green Version]
- Letourneux, C.; Rocher, G.; Porteu, F. B56-containing PP2A dephosphorylate ERK and their activity is controlled by the early gene IEX-1 and ERK. EMBO J. 2006, 25, 727–738. [Google Scholar] [CrossRef] [Green Version]
- Harada, T.; Morooka, T.; Ogawa, S.; Nishida, E. ERK induces p35, a neuron-specific activator of Cdk5, through induction of Egr1. Nature 2001, 3, 453–459. [Google Scholar] [CrossRef]
- Duclot, F.; Kabbaj, M. The Role of Early Growth Response 1 (EGR1) in Brain Plasticity and Neuropsychiatric Disorders. Front. Behav. Neurosci. 2017, 11, 35. [Google Scholar] [CrossRef] [Green Version]
- Shan, J.; Dudenhausen, E.; Kilberg, M.S. Induction of early growth response gene 1 (EGR1) by endoplasmic reticulum stress is mediated by the extracellular regulated kinase (ERK) arm of the MAPK pathways. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Lo, L.W.; Cheng, J.J.; Chiu, J.J.; Wung, B.S.; Liu, Y.C.; Wang, D.L. Endothelial exposure to hypoxia induces Egr-1 expression involving PKCalpha-mediated Ras/Raf-1/ERK1/2 pathway. J. Cell. Physiol. 2001, 188, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.P.; Jain, N.; Cao, X. Stress-induced immediate-early gene, egr-1, involves activation of p38/JNK1. Oncogene 1998, 16, 2915–2926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.H.; Jang, W.H.; Seo, J.A.; Park, M.; Lee, T.R.; Park, Y.H.; Kim, D.K.; Lim, K.M. Decrease of ceramides with very long-chain fatty acids and downregulation of elongases in a murine atopic dermatitis model. J. Investig. Dermatol. 2012, 132, 476–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Test Chemicals | CAS No. | Physical State | Supplier | Category a | Conc. b |
|---|---|---|---|---|---|---|
| 1 | 2-Ethoxyethyl methacrylate | 2370-63-0 | liquid | Sigma-Aldrich | NC | 10% |
| 2 | Piperonyl butoxide | 1951-03-06 | liquid | Toronto research chemicals, Toronto, Canada | NC | 10% |
| 3 | 1-Ethyl-3-methylimidazolium ethylsulfate | 342573-75-5 | liquid | Sigma-Aldrich | NC | 10% |
| 4 | Potassium tetrafluoroborate | 14075-53-7 | solid | Sigma-Aldrich | NC | 10% |
| 5 | Polyoxyl 40 hydrogenated castor oil | 61788-85-0 | viscous | Sigma-Aldrich | NC | 10% |
| 6 | Dipropyl disulfide | 629-19-6 | liquid | Sigma-Aldrich | NC | 10% |
| 7 | 2-Methyl-1-pentanol | 105-30-6 | liquid | Sigma-Aldrich | Cat2B | 1.25% |
| 8 | Diethyl tolumide | 134-62-3 | liquid | Tokyo Chemical Industry, Tokyo, Japan | Cat2B | 1.92% |
| 9 | 1,4-Dibutoxy benzene | 104-36-9 | solid | Tokyo Chemical Industry | Cat2B | 5% |
| 10 | 2,4,11,13-Tetraazatetradecane diimidamide, N,N″-bis(4chlorophenyl)-3,12-diimino-, di-d-gluconate | 18472-51-0 | liquid | Sigma-Aldrich | Cat2A | 0.05% |
| 11 | Gamma-butyrolactone | 96-48-0 | liquid | Sigma-Aldrich | Cat2A | 7.69% |
| 12 | (Ethylenediamine-propyl)-trimethoxysilane | 1760-24-3 | liquid | Sigma-Aldrich | Cat1 | 0.25% |
| 13 | Tetraethylene glycol diacrylate | 17831-71-9 | liquid | Sigma-Aldrich | Cat1 | 0.01% |
| 14 | 1,2-Benzisothiazol-3(2H)-one | 2634-33-5 | solid | Sigma-Aldrich | Cat1 | 0.01% |
| 15 | 3,5-Dimethyl-2,5-hexanediol | 110-03-2 | solid | Sigma-Aldrich | Cat1 | 0.1% |
| 16 | Disodium 2,2′-([1,1′-biphenyl]-4,4′diyldivinylene)bis-(benzenesulfonate) | 27355-41-8 | solid | Tokyo Chemical Industry | Cat1 | 1% |
| 17 | Sodium oxalate | 62-76-0 | solid | Sigma-Aldrich | Cat1 | 3% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, D.-B.; Kim, S.-Y.; Won, D.-H.; Kim, C.; Shin, Y.-S.; Park, J.-H.; Chun, Y.-J.; Lim, K.-M.; Yun, J.-W. Egr1 Gene Expression as a Potential Biomarker for In Vitro Prediction of Ocular Toxicity. Pharmaceutics 2021, 13, 1584. https://doi.org/10.3390/pharmaceutics13101584
Hwang D-B, Kim S-Y, Won D-H, Kim C, Shin Y-S, Park J-H, Chun Y-J, Lim K-M, Yun J-W. Egr1 Gene Expression as a Potential Biomarker for In Vitro Prediction of Ocular Toxicity. Pharmaceutics. 2021; 13(10):1584. https://doi.org/10.3390/pharmaceutics13101584
Chicago/Turabian StyleHwang, Da-Bin, Shin-Young Kim, Dong-Hoon Won, Changuk Kim, Yoo-Sub Shin, Jong-Hwan Park, Young-Jin Chun, Kyung-Min Lim, and Jun-Won Yun. 2021. "Egr1 Gene Expression as a Potential Biomarker for In Vitro Prediction of Ocular Toxicity" Pharmaceutics 13, no. 10: 1584. https://doi.org/10.3390/pharmaceutics13101584
APA StyleHwang, D.-B., Kim, S.-Y., Won, D.-H., Kim, C., Shin, Y.-S., Park, J.-H., Chun, Y.-J., Lim, K.-M., & Yun, J.-W. (2021). Egr1 Gene Expression as a Potential Biomarker for In Vitro Prediction of Ocular Toxicity. Pharmaceutics, 13(10), 1584. https://doi.org/10.3390/pharmaceutics13101584