Potential Herb–Drug Interactions in the Management of Age-Related Cognitive Dysfunction

 ,
,  , and
, and 
Abstract
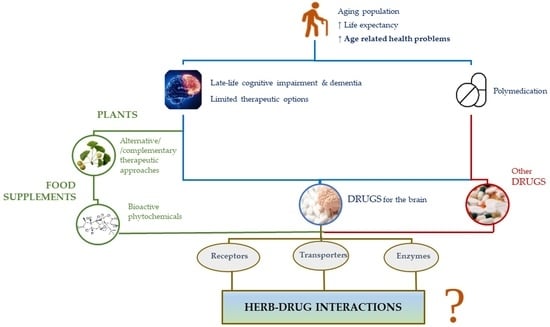
1. Introduction
2. Methodology
3. Botanicals for Cognitive Enhancement
3.1. Identification and Selection of Botanicals (and Respective Bioactives) Implicated in Cognition Enhancement
3.2. Mechanisms of Action in Cognition Enhancement
4. Chemical Characterization of Nootropic Bioactive Compounds
5. Interactions between Botanicals and Drugs
5.1. Protein Targets as Key Points for Herb–Drug Interactions
5.1.1. Cytochrome P450
5.1.2. Uridine Diphosphate-Glucuronosyltransferases
5.1.3. Drug Carriers
5.1.4. Other Targets
5.2. Drugs Used in Elderly Patients
5.3. Target Modulation by Bioactives
5.4. Assessment of HDI Potential
5.4.1. Diclofenac
5.4.2. Atorvastatin
5.5. Food Supplements
6. Final Remarks
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABC | ATP-binding cassette |
| ABCG2 | ATP Binding Cassette Subfamily G Member 2 |
| AChE | Acetylcholinesterase |
| ADME | Absorption, distribution, metabolism and elimination |
| Al | Alprazolam |
| ALOX5 | Arachidonate 5-lipoxygenase |
| AMPK | AMP-activated protein kinase |
| At | Atorvastatin |
| ATC | Anatomical Therapeutic Chemical |
| BChE | Butyrylcholinesterase |
| BCRP | Breast cancer resistance protein |
| BDDCS | Biopharmaceutics Drug Disposition Classification System |
| BSEP | Bile salt export pump |
| Cl | Clopidogrel |
| COMT | Catechol-O-methyltransferase |
| COX | Cyclooxygenase |
| CYP | Cytochrome P450 |
| Di | Diclofenac |
| ECG | Epicatechin gallate |
| EGCG | Epigallocatechin-3-gallate |
| GABA | γ-Aminobutyric acid |
| GLP-1 | Glucagon-like peptide-1 |
| HAM | High-Alert Medications |
| HDI | Herb–drug interactions |
| 5-HI | 5-hydroxyndoleacetic acid |
| 5-HT | 5-hydroxytryptamine |
| HMC | Herbal Medicines Compendium |
| HMGCoAR | 3-hydroxy-3-methyl-glutaril-CoA reductase; |
| IP | Indian Pharmacopoeia |
| ITC | International Transporter Consortium |
| JP | The Japanese Pharmacopoeia |
| L-DOPA | Levodopa |
| MAO | Monoamine oxidase |
| MATE | Multi-antimicrobial extrusion protein |
| MDR1 | Multidrug resistance protein 1 |
| Me | Memantine |
| MRP | Multidrug resistance-associated protein |
| Mt | Metformin |
| NMDA | N-methyl-d-aspartate |
| OAT | Organic anion transporter |
| OATP | Organic-anion-transporting polypeptide |
| OCT | Organic cation transport |
| PEPT | Peptide transporter |
| P-gP | Glycoprotein P |
| Ph.Eur | European Pharmacopoeia |
| Pi | Piracetam |
| PLA2G2A | Phospholipase A2 Group IIA |
| PPRC | Pharmacopoeia of People’s Republic of China |
| Pr | Propranolol |
| Se | Sertraline |
| SLC | Solute Carriers |
| SLCO | Solute carrier organic anion transporter |
| Ta | Tadalafil |
| UGT | Uridine diphosphate-glucuronosyltransferase |
| USP | United States Pharmacopoeia |
| WHO | World Health Organization |
References
- Winblad, B.; Amouyel, P.; Andrieu, S.; Ballard, C.; Brayne, C.; Brodaty, H.; Cedazo-Minguez, A.; Dubois, B.; Edvardsson, D.; Feldman, H.; et al. Defeating Alzheimer’s disease and other dementias: A priority for European science and society. Lancet Neurol. 2016, 15, 455–532. [Google Scholar] [CrossRef]
- Wu, Y.-T.; Fratiglioni, L.; Matthews, F.E.; Lobo, A.; Breteler, M.M.B.; Skoog, I.; Brayne, C. Dementia in western Europe: Epidemiological evidence and implications for policy making. Lancet Neurol. 2016, 15, 116–124. [Google Scholar] [CrossRef]
- Prince, M.; Ali, G.-C.; Guerchet, M.; Prina, A.M.; Albanese, E.; Wu, Y.-T. Recent global trends in the prevalence and incidence of dementia, and survival with dementia. Alzheimers. Res. Ther. 2016, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Dembitsky, V.M.; Dzhemileva, L.; Gloriozova, T.; D’yakonov, V. Natural and synthetic drugs used for the treatment of the dementia. Biochem. Biophys. Res. Commun. 2020, 524, 772–783. [Google Scholar] [CrossRef]
- Howes, M.-J.R.; Perry, E.K.; Perry, N.S.L.; Vásquez-londoño, C. Role of phytochemicals as nutraceuticals for cognitive functions affected in ageing. Br. J. Pharmacol. 2020, 177, 1294–1315. [Google Scholar] [CrossRef]
- Dominguez, L.J.; Barbagallo, M. Nutritional prevention of cognitive decline and dementia. Acta BioMed 2018, 89, 276–290. [Google Scholar] [CrossRef]
- Solfrizzi, V.; Agosti, P.; Lozupone, M.; Custodero, C.; Schilardi, A.; Valiani, V.; Santamato, A.; Sardone, R.; Dibello, V.; Di Lena, L.; et al. Nutritional interventions and cognitive-related outcomes in patients with late-life cognitive disorders: A systematic review. Neurosci. Biobehav. Rev. 2018, 95, 480–498. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, S.; Kan, J.; Zhang, J.; Zhou, L.; Huang, Y.; Zhang, Y. Chinese Herbal Medicine Interventions in Neurological Disorder Therapeutics by Regulating Glutamate Signaling. Curr. Neuropharmacol. 2020, 18, 260–276. [Google Scholar] [CrossRef]
- Shen, J.; Xu, L.; Qu, C.; Sun, H.; Zhang, J. Resveratrol prevents cognitive deficits induced by chronic unpredictable mild stress: Sirt1/miR-134 signalling pathway regulates CREB/BDNF expression in hippocampus in vivo and in vitro. Behav. Brain Res. 2018, 349, 1–7. [Google Scholar] [CrossRef]
- Sowndhararajan, K.; Kim, S. Neuroprotective and cognitive enhancement potentials of Angelica gigas nakai root: A review. Sci. Pharm. 2017, 85, 21. [Google Scholar] [CrossRef]
- Tan, C.-C.; Yu, J.-T.; Wang, H.-F.; Tan, M.-S.; Meng, X.-F.; Wang, C.; Jiang, T.; Zhu, X.-C.; Tan, L. Efficacy and safety of donepezil, galantamine, rivastigmine, and memantine for the treatment of Alzheimer’s disease: A systematic review and meta-analysis. J. Alzheimers. Dis. 2014, 41, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Kasture, S.; Mohan, M.; Kasture, V. Mucuna pruriens seeds in treatment of Parkinson’s disease: Pharmacological review. Orient. Pharm. Exp. Med. 2013, 13, 165–174. [Google Scholar] [CrossRef]
- Cassani, E.; Cilia, R.; Laguna, J.; Barichella, M.; Contin, M.; Cereda, E.; Isaias, I.U.; Sparvoli, F.; Akpalu, A.; Budu, K.O.; et al. Mucuna pruriens for Parkinson’s disease: Low-cost preparation method, laboratory measures and pharmacokinetics profile. J. Neurol. Sci. 2016, 365, 175–180. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Global Action Plan for the Prevention and Control of Noncommunicable Diseases 2013–2020; World Health Organization: Geneva, Switzerland, 2013; ISBN 9789241506236.
- Sut, S.; Baldan, V.; Faggian, M.; Peron, G.; Dall Acqua, S. Nutraceuticals, A New Challenge for Medicinal Chemistry. Curr. Med. Chem. 2016, 23, 3198–3223. [Google Scholar] [CrossRef]
- Tundis, R.; Loizzo, M.R.; Nabavi, S.M.; Orhan, I.E.; Skalicka-Woźniak, K.; D’Onofrio, G.; Aiello, F. Chapter 3—Natural Compounds and Their Derivatives as Multifunctional Agents for the Treatment of Alzheimer Disease. In Discovery and Development of Neuroprotective Agents from Natural Products; Brahmachari, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 63–102. ISBN 9780128095935. [Google Scholar]
- López, S.; Bastida, J.; Viladomat, F.; Codina, C. Acetylcholinesterase inhibitory activity of some Amaryllidaceae alkaloids and Narcissus extracts. Life Sci. 2002, 71, 2521–2529. [Google Scholar] [CrossRef]
- Lin, Z.; Gu, J.; Xiu, J.; Mi, T.; Dong, J.; Tiwari, J.K. Traditional chinese medicine for senile dementia. Evid. Based Complement. Altern. Med. 2012, 2012, 692621. [Google Scholar] [CrossRef]
- Kennedy, D.O.; Wightman, E.L. Herbal extracts and phytochemicals: Plant secondary metabolites and the enhancement of human brain function. Adv. Nutr. 2011, 2, 32–50. [Google Scholar] [CrossRef]
- Perry, E.; Howes, M.-J.R. Medicinal plants and dementia therapy: Herbal hopes for brain aging? CNS Neurosci. Ther. 2011, 17, 683–698. [Google Scholar] [CrossRef]
- Wightman, E.L. Potential benefits of phytochemicals against Alzheimer’s disease. Proc. Nutr. Soc. 2017, 76, 106–112. [Google Scholar] [CrossRef]
- Ahmed, F.; Ghalib, R.M.; Sasikala, P.; Ahmed, K.K.M. Cholinesterase inhibitors from botanicals. Pharmacogn. Rev. 2013, 7, 121–130. [Google Scholar] [CrossRef]
- Hostettmann, K.; Borloz, A.; Urbain, A.; Marston, A. Natural product inhibitors of acetylcholinesterase. Curr. Org. Chem. 2006, 10, 825–847. [Google Scholar] [CrossRef]
- Zhou, X.; Chan, K.; Yeung, J.H.K. Herb-drug interactions with Danshen (Salvia miltiorrhiza): A review on the role of cytochrome P450 enzymes. Drug Metabol. Drug Interact. 2012, 27, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Clairet, A.-L.; Boiteux-Jurain, M.; Curtit, E.; Jeannin, M.; Gérard, B.; Nerich, V.; Limat, S. Interaction between phytotherapy and oral anticancer agents: Prospective study and literature review. Med. Oncol. 2019, 36, 45. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, A.S.; Thomas, A.B.; Chitlange, S.S. Herb-drug interaction studies of herbs used in treatment of cardiovascular disorders-A narrative review of preclinical and clinical studies. Phytother. Res. 2020, 34, 1008–1026. [Google Scholar] [CrossRef] [PubMed]
- Kantor, E.D.; Rehm, C.D.; Haas, J.S.; Chan, A.T.; Giovannucci, E.L. Trends in Prescription Drug Use Among Adults in the United States From 1999–2012. JAMA 2015, 314, 1818–1831. [Google Scholar] [CrossRef]
- Payne, R.A. The epidemiology of polypharmacy. Clin. Med. 2016, 16, 465–469. [Google Scholar] [CrossRef]
- Council of Europe (Ed.) European Pharmacopoeia (up to 10.2), 10th ed.; EDQM: Strasbourg, France, 2019; ISBN 9789287189233. [Google Scholar]
- United States Pharmacopoeia Convention Inc. United States Pharmacopeia National Formulary USP43 NF38; United States Pharmacopoeia Convention Inc.: Rockville, MD, USA, 2019. [Google Scholar]
- Society of Japanese Pharmacopoeia (Ed.) The Japanese Pharmacopoeia (English Version), 17th ed.; Yakuji Nippo Ltd.: Tokyo, Japan, 2016; ISBN 9784840813716. [Google Scholar]
- Indian Pharmacopoeia Commission. Indian Pharmacopoeia 2018, 8th ed.; Indian Pharmacopoeia Commission: Ghaziabad, India, 2018; Volume 1–4, ISBN 9789381238196.
- Chinese Pharmacopoeia Commission (Ed.) Pharmacopoeia of the People’s Republic of China; China Medical Science Press: Beijing, China, 2015; Volume 1, ISBN 9787506789295. [Google Scholar]
- WHO Monographs on Selected Medicinal Plants. Available online: https://apps.who.int/iris/handle/10665/42052 (accessed on 9 January 2021).
- Herbal Medicines Compendium. Available online: https://hmc.usp.org/ (accessed on 9 January 2021).
- World Health Organization. WHOCC—ATC/DDD Index. Available online: https://www.whocc.no/atc_ddd_index/ (accessed on 28 November 2020).
- Golfar, Y.; Shayanfar, A. Prediction of Biopharmaceutical Drug Disposition Classification System (BDDCS) by Structural Parameters. J. Pharm. Pharm. Sci. 2019, 22, 247–269. [Google Scholar] [CrossRef]
- Benet, L.Z.; Broccatelli, F.; Oprea, T.I. BDDCS applied to over 900 drugs. AAPS J. 2011, 13, 519–547. [Google Scholar] [CrossRef]
- Mora, M.J.; Onnainty, R.; Granero, G.E. Comparative Oral Drug Classification Systems: Acetazolamide, Azithromycin, Clopidogrel, and Efavirenz Case Studies. Mol. Pharm. 2018, 15, 3187–3196. [Google Scholar] [CrossRef]
- Benet, L.Z. The role of BCS (biopharmaceutics classification system) and BDDCS (biopharmaceutics drug disposition classification system) in drug development. J. Pharm. Sci. 2013, 102, 34–42. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, X.; Benet, L.Z. Reliability of In Vitro and In Vivo Methods for Predicting the Effect of P-Glycoprotein on the Delivery of Antidepressants to the Brain. Clin. Pharmacokinet. 2016, 55, 143–167. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Benet, L.Z.; Okochi, H.; Chen, X. pH Dependent but not P-gp Dependent Bidirectional Transport Study of S-propranolol: The Importance of Passive Diffusion. Pharm. Res. 2015, 32, 2516–2526. [Google Scholar] [CrossRef] [PubMed]
- Hosey, C.M.; Chan, R.; Benet, L.Z. BDDCS Predictions, Self-Correcting Aspects of BDDCS Assignments, BDDCS Assignment Corrections, and Classification for more than 175 Additional Drugs. AAPS J. 2016, 18, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Institute for Safe Medication Practices. ISMP List of High-Alert Medications in Acute Care Settings 2018; Institute for Safe Medication Practices: Horsham, PA, USA, 2020. [Google Scholar]
- Maroyi, A. Albizia adianthifolia: Botany, Medicinal Uses, Phytochemistry, and Pharmacological Properties. Sci. World J. 2018, 2018, 7463584. [Google Scholar] [CrossRef] [PubMed]
- Sonibare, M.A.; Ayoola, I.O.; Elufioye, T.O. Antioxidant and acetylcholinesterase inhibitory activities of leaf extract and fractions of Albizia adianthifolia (Schumach) W.F. Wright. J. Basic Clin. Physiol. Pharmacol. 2017, 28, 143–148. [Google Scholar] [CrossRef]
- Toukea, D.D.; Kamto, E.L.D.; Simo, L.M.; Mbing, J.N.; Antheaume, C.; Haddad, M.; Noté, O.P.; Pegnyemb, D.E. New triterpenoid saponin from the stems of Albizia adianthifolia (Schumach.) W.Wight. Nat. Prod. Res. 2020, 1–9. [Google Scholar] [CrossRef]
- Ferraz, A.C.; Angelucci, M.E.; da Costa, M.L.; Batista, I.R.; de Oliveira, B.H.; da Cunha, C. Pharmacological evaluation of ricinine, a central nervous system stimulant isolated from Ricinus communis. Pharmacol. Biochem. Behav. 1999, 63, 367–375. [Google Scholar] [CrossRef]
- Patel, K.; Patel, D.K. Medicinal significance, pharmacological activities, and analytical aspects of ricinine: A concise report. J. Coast. Life Med. 2016, 4, 663–667. [Google Scholar] [CrossRef]
- Popiołek, A.K.; Chyrek-Tomaszewska, A.; Stachowicz-Karpińska, A.; Bieliński, M.K.; Borkowska, A. Biochemical parameters in cognitive functions. Neuropsychiatr. Dis. Treat. 2020, 16, 2479–2489. [Google Scholar] [CrossRef]
- Sekhar, V.C.; Viswanathan, G.; Baby, S. Insights Into the Molecular Aspects of Neuroprotective Bacoside A and Bacopaside I. Curr. Neuropharmacol. 2019, 17, 438–446. [Google Scholar] [CrossRef]
- Rezende, D.A.D.C.S.; das Graças Cardoso, M.; Souza, R.V.; Teixeira, M.L.; Brandão, R.M.; Ferreira, V.R.F.; e Nogueira, J.O.; Magalhães, M.L.; Marcussi, S.; Nelson, D.L.; et al. Essential oils from Mentha piperita, Cymbopogon citratus, Rosmarinus officinalis, Peumus boldus and Foeniculum vulgare: Inhibition of phospholipase A2 and cytotoxicity to human erythrocytes. Am. J. Plant Sci. 2017, 8, 2196. [Google Scholar] [CrossRef][Green Version]
- Gray, N.E.; Alcazar Magana, A.; Lak, P.; Wright, K.M.; Quinn, J.; Stevens, J.F.; Maier, C.S.; Soumyanath, A. Centella asiatica—Phytochemistry and mechanisms of neuroprotection and cognitive enhancement. Phytochem. Rev. 2018, 17, 161–194. [Google Scholar] [CrossRef] [PubMed]
- Rajabian, A.; Hosseini, A.; Hosseini, M.; Sadeghnia, H.R. A review of potential efficacy of Saffron (Crocus sativus L.) in cognitive dysfunction and seizures. Prev. Nutr. Food Sci. 2019, 24, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; More, S.V.; Han, S.-D.; Choi, J.-Y.; Choi, D.-K. Promising therapeutics with natural bioactive compounds for improving learning and memory—A review of randomized trials. Molecules 2012, 17, 10503–10539. [Google Scholar] [CrossRef]
- Sharma, V.; Singh, I.; Chaudhary, P. Acorus calamus (The Healing Plant): A review on its medicinal potential, micropropagation and conservation. Nat. Prod. Res. 2014, 28, 1454–1466. [Google Scholar] [CrossRef]
- Shiksharthi, A.R.; Mittal, S.; Ramana, J. Systematic review of herbals as potential memory enhancers. Int. J. Herb. Med. 2011, 3, 20–24. [Google Scholar]
- Pak, M.E.; Kim, Y.R.; Kim, H.N.; Ahn, S.M.; Shin, H.K.; Baek, J.U.; Choi, B.T. Studies on medicinal herbs for cognitive enhancement based on the text mining of Dongeuibogam and preliminary evaluation of its effects. J. Ethnopharmacol. 2016, 179, 383–390. [Google Scholar] [CrossRef]
- Ishola, I.O.; Awoyemi, A.A.; Afolayan, G.O. Involvement of antioxidant system in the amelioration of scopolamine-induced memory impairment by Grains of Paradise (Aframomum melegueta K. Schum.) extract. Drug Res. 2016, 66, 455–463. [Google Scholar] [CrossRef]
- Adefegha, S.A.; Oboh, G. Acetylcholinesterase (AChE) inhibitory activity, antioxidant properties and phenolic composition of two Aframomum species. J. Basic Clin. Physiol. Pharmacol. 2012, 23, 153–161. [Google Scholar] [CrossRef]
- Shi, S.-H.; Zhao, X.; Liu, B.; Li, H.; Liu, A.-J.; Wu, B.; Bi, K.-S.; Jia, Y. The effects of sesquiterpenes-rich extract of Alpinia oxyphylla Miq. on amyloid-β-induced cognitive impairment and neuronal abnormalities in the cortex and hippocampus of mice. Oxid. Med. Cell. Longev. 2014, 2014, 451802. [Google Scholar] [CrossRef]
- Wei, W.-L.; Zeng, R.; Gu, C.-M.; Qu, Y.; Huang, L.-F. Angelica sinensis in China-A review of botanical profile, ethnopharmacology, phytochemistry and chemical analysis. J. Ethnopharmacol. 2016, 190, 116–141. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.C.; Hsieh, C.L. Pharmacological effects of Radix Angelica sinensis (Danggui) on cerebral infarction. Chin. Med. 2011, 6, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Nisar, M.; Khan, N.; Saeed, M.; Nadeem, S.; Ali, F.; Karim, N.; Kaleem, W.A.; Qayum, M.; Ahmad, H.; et al. Structural insights to investigate Conypododiol as a dual cholinesterase inhibitor from Asparagus adscendens. Fitoterapia 2010, 81, 1020–1025. [Google Scholar] [CrossRef] [PubMed]
- Pahwa, P.; Goel, R.K. Asparagus adscendens root extract enhances cognition and protects against scopolamine induced amnesia: An in-silico and in-vivo studies. Chem. Biol. Interact. 2016, 260, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.S.; Al Mamun, A.; Kabir, M.T.; Jakaria, M.; Mathew, B.; Barreto, G.E.; Ashraf, G.M. Nootropic and Anti-Alzheimer’s Actions of Medicinal Plants: Molecular Insight into Therapeutic Potential to Alleviate Alzheimer’s Neuropathology. Mol. Neurobiol. 2019, 56, 4925–4944. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-G.; Jan, Y.-S.; Tsai, P.-W.; Norimoto, H.; Michihara, S.; Murayama, C.; Wang, C.-C. Anti-inflammatory and Antinociceptive Constituents of Atractylodes japonica Koidzumi. J. Agric. Food Chem. 2016, 64, 2254–2262. [Google Scholar] [CrossRef] [PubMed]
- Dubey, T.; Chinnathambi, S. Brahmi (Bacopa monnieri): An ayurvedic herb against the Alzheimer’s disease. Arch. Biochem. Biophys. 2019, 676, 108153. [Google Scholar] [CrossRef]
- Peth-Nui, T.; Wattanathorn, J.; Muchimapura, S.; Tong-Un, T.; Piyavhatkul, N.; Rangseekajee, P.; Ingkaninan, K.; Vittaya-Areekul, S. Effects of 12-week Bacopa monnieri consumption on attention, cognitive processing, working memory, and functions of both cholinergic and monoaminergic systems in healthy elderly volunteers. Evid. Based Complement. Altern. Med. 2012, 2012, 606424. [Google Scholar] [CrossRef]
- Devendra, P.; Patel, S.S.; Birwal, P.; Basu, S.; Deshmukh, G.; Datir, R. Brahmi (Bacopa monnieri) as functional food ingredient in food processing industry. J. Pharmacogn. Phytochem. 2018, 7, 189–194. [Google Scholar]
- Muhammad, A.; Dauda, U.; Jibril, S.; Sirat, H.M. Acetylcholinesterase inhibitory activity of a cyclitol isolated from the leaves of Bauhinia rufescens. BAJOPAS 2019, 11, 431. [Google Scholar] [CrossRef]
- Ata, A.; Iverson, C.D.; Kalhari, K.S.; Akhter, S.; Betteridge, J.; Meshkatalsadat, M.H.; Orhan, I.; Sener, B. Triterpenoidal alkaloids from Buxus hyrcana and their enzyme inhibitory, anti-fungal and anti-leishmanial activities. Phytochemistry 2010, 71, 1780–1786. [Google Scholar] [CrossRef] [PubMed]
- Mancini, E.; Beglinger, C.; Drewe, J.; Zanchi, D.; Lang, U.E.; Borgwardt, S. Green tea effects on cognition, mood and human brain function: A systematic review. Phytomedicine 2017, 34, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Van Dusseldorp, T.A.; Stratton, M.T.; Bailly, A.R.; Holmes, A.J.; Alesi, M.G.; Feito, Y.; Mangine, G.T.; Hester, G.M.; Esmat, T.A.; Barcala, M.; et al. Safety of Short-Term Supplementation with Methylliberine (Dynamine®) Alone and in Combination with TeaCrine® in Young Adults. Nutrients 2020, 12, 654. [Google Scholar] [CrossRef] [PubMed]
- Keshavarz, M.; Farrokhi, M.R.; Amiri, A.; Hosseini, M. The contribution of S100B to the glioprotective effects of valproic and arundic acids. Iran. J. Basic Med. Sci. 2019, 22, 557–562. [Google Scholar] [CrossRef]
- Pham, H.M.; Xu, A.; Schriner, S.E.; Sevrioukov, E.A.; Jafari, M. Cinnamaldehyde Improves Lifespan and Healthspan in Drosophila melanogaster Models for Alzheimer’s Disease. Biomed Res. Int. 2018, 2018, 3570830. [Google Scholar] [CrossRef]
- Irie, Y. Effects of eugenol on the central nervous system: Its possible application to treatment of Alzheimer’s disease, depression, and Parkinson’s disease. CBC 2006, 2, 57–66. [Google Scholar] [CrossRef]
- Kumar, S.; Kumari, R.; Mishra, S. Pharmacological properties and their medicinal uses of Cinnamomum: A review. J. Pharm. Pharmacol. 2019, 71, 1735–1761. [Google Scholar] [CrossRef]
- Stohs, S.J. Safety, Efficacy, and Mechanistic Studies Regarding Citrus aurantium (Bitter Orange) Extract and p-Synephrine. Phytother. Res. 2017, 31, 1463–1474. [Google Scholar] [CrossRef]
- Bello, M.L.; Walker, A.J.; McFadden, B.A.; Sanders, D.J.; Arent, S.M. The effects of TeaCrine® and caffeine on endurance and cognitive performance during a simulated match in high-level soccer players. J. Int. Soc. Sports Nutr. 2019, 16, 20. [Google Scholar] [CrossRef]
- Pomeroy, D.E.; Tooley, K.L.; Probert, B.; Wilson, A.; Kemps, E. A systematic review of the effect of dietary supplements on cognitive performance in healthy young adults and military personnel. Nutrients 2020, 12, 545. [Google Scholar] [CrossRef]
- Owona, B.A.; Zug, C.; Schluesener, H.J.; Zhang, Z.-Y. Protective effects of forskolin on behavioral deficits and neuropathological changes in a mouse model of cerebral amyloidosis. J. Neuropathol. Exp. Neurol. 2016, 75, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Amin, H.; Sharma, R.; Vyas, M.; Prajapati, P.K.; Dhiman, K. Shankhapushpi (Convolvulus pluricaulis Choisy): Validation of the Ayurvedic therapeutic claims through contemporary studies. Int. J. Green Pharm. 2014, 8, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Kuo, P.-C.; Yang, M.-L.; Hwang, T.-L.; Lai, Y.-Y.; Li, Y.-C.; Thang, T.D.; Wu, T.-S. Anti-inflammatory diterpenoids from Croton tonkinensis. J. Nat. Prod. 2013, 76, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Montaño, J.M.; Burgos-Morón, E.; Pérez-Guerrero, C.; López-Lázaro, M. A review on the dietary flavonoid kaempferol. Mini Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef]
- Tohda, C.; Yang, X.; Matsui, M.; Inada, Y.; Kadomoto, E.; Nakada, S.; Watari, H.; Shibahara, N. Diosgenin-rich yam extract enhances cognitive function: A placebo-controlled, randomized, double-blind, crossover study of healthy adults. Nutrients 2017, 9, 1160. [Google Scholar] [CrossRef]
- Chiu, C.-S.; Chiu, Y.-J.; Wu, L.-Y.; Lu, T.-C.; Huang, T.-H.; Hsieh, M.-T.; Lu, C.-Y.; Peng, W.-H. Diosgenin ameliorates cognition deficit and attenuates oxidative damage in senescent mice induced by D-galactose. Am. J. Chin. Med. 2011, 39, 551–563. [Google Scholar] [CrossRef]
- Patocka, J. Bioactivity of Echium amoenum: A mini review. BJSTR 2019, 20, 14915–14917. [Google Scholar] [CrossRef]
- Rabiei, Z.; Setorki, M. Effect of hydroalcoholic Echium amoenum extract on scopolamine-induced learning and memory impairment in rats. Pharm. Biol. 2018, 56, 672–677. [Google Scholar] [CrossRef]
- Kim, Y.H.; Cho, M.L.; Kim, D.B.; Shin, G.H.; Lee, J.H.; Lee, J.S.; Park, S.O.; Lee, S.J.; Shin, H.M.; Lee, O.H. The antioxidant activity and their major antioxidant compounds from Acanthopanax senticosus and A. koreanum. Molecules 2015, 20, 13281–13295. [Google Scholar] [CrossRef]
- Kwan, C.-Y.; Zhang, W.-B.; Sim, S.-M.; Deyama, T.; Nishibe, S. Vascular effects of Siberian ginseng (Eleutherococcus senticosus): Endothelium-dependent No- and EDHF-mediated relaxation depending on vessel size. Naunyn. Schmiedebergs. Arch. Pharmacol. 2004, 369, 473–480. [Google Scholar] [CrossRef]
- Lee, D.; Park, J.; Yoon, J.; Kim, M.-Y.; Choi, H.-Y.; Kim, H. Neuroprotective effects of Eleutherococcus senticosus bark on transient global cerebral ischemia in rats. J. Ethnopharmacol. 2012, 139, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Moni, D.A.; Sonawane, K.D.; Paek, K.Y.; Shohael, A.M. A comprehensive in silico exploration of pharmacological properties, bioactivities and COX-2 inhibitory potential of eleutheroside B from Eleutherococcus senticosus (Rupr. & Maxim.) Maxim. J. Biomol. Struct. Dyn. 2020, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Joshi, H.; Parle, M. Cholinergic basis of memory-strengthening effect of Foeniculum vulgare Linn. J. Med. Food 2006, 9, 413–417. [Google Scholar] [CrossRef]
- Sayah, K.; El Omari, N.; Kharbach, M.; Bouyahya, A.; Kamal, R.; Marmouzi, I.; Cherrah, Y.; Faouzi, M.E.A. Comparative Study of Leaf and Rootstock Aqueous Extracts of Foeniculum vulgare on Chemical Profile and In Vitro Antioxidant and Antihyperglycemic Activities. Adv. Pharmacol. Pharm. Sci. 2020, 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nemati, M.; Hemmati, A.A.; Najafzadeh, H.; Mansouri, M.T.; Khodayar, M.J. Evaluation of the effects of Foeniculum vulgare essence on behavioral-motor disorders of Parkinson’s Disease induced by reserpine in ovariectomized and non ovariectomized rats. Jundishapur J. Nat. Pharm. Prod. 2018, 13, e67391. [Google Scholar] [CrossRef]
- Nguyen, T.; Alzahrani, T. Ginkgo Biloba. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Ben-Eliezer, D.; Yechiam, E. Hypericum perforatum as a cognitive enhancer in rodents: A meta-analysis. Sci. Rep. 2016, 6, 35700. [Google Scholar] [CrossRef]
- Oliveira, A.I.; Pinho, C.; Sarmento, B.; Dias, A.C.P. Neuroprotective Activity of Hypericum perforatum and Its Major Components. Front. Plant Sci. 2016, 7, 1004. [Google Scholar] [CrossRef]
- Widy-Tyszkiewicz, E.; Piechal, A.; Joniec, I.; Blecharz-Klin, K. Long term administration of Hypericum perforatum improves spatial learning and memory in the water maze. Biol. Pharm. Bull. 2002, 25, 1289–1294. [Google Scholar] [CrossRef]
- Dinamarca, M.C.; Cerpa, W.; Garrido, J.; Hancke, J.L.; Inestrosa, N.C. Hyperforin prevents beta-amyloid neurotoxicity and spatial memory impairments by disaggregation of Alzheimer’s amyloid-beta-deposits. Mol. Psychiatry 2006, 11, 1032–1048. [Google Scholar] [CrossRef]
- Karioti, A.; Bilia, A.R. Hypericins as potential leads for new therapeutics. Int. J. Mol. Sci. 2010, 11, 562–594. [Google Scholar] [CrossRef]
- da Silva, N.L.P.; Cabrera, L.P.B.; Medeiros, L.L.M.; Formigoni, M.; Fuchs, R.H.B.; Droval, A.A.; Reitz, F.A.C. Medicinal effects of Peruvian maca (Lepidium meyenii): A review. Food Funct. 2020, 11, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhu, F. Chemical composition and health effects of maca (Lepidium meyenii). Food Chem. 2019, 288, 422–443. [Google Scholar] [CrossRef] [PubMed]
- Alasmari, M.; Böhlke, M.; Kelley, C.; Maher, T.; Pino-Figueroa, A. Inhibition of Fatty Acid Amide Hydrolase (FAAH) by Macamides. Mol. Neurobiol. 2019, 56, 1770–1781. [Google Scholar] [CrossRef]
- Almukadi, H.; Wu, H.; Böhlke, M.; Kelley, C.J.; Maher, T.J.; Pino-Figueroa, A. The Macamide N-3-Methoxybenzyl-Linoleamide Is a Time-Dependent Fatty Acid Amide Hydrolase (FAAH) Inhibitor. Mol. Neurobiol. 2013, 48, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, G.F.; Gonzales-Castañeda, C. The Methyltetrahydro-beta-Carbolines in Maca (Lepidium meyenii). Evid. Based Complement. Altern. Med. 2009, 6, 315–316. [Google Scholar] [CrossRef]
- López-Ríos, L.; Wiebe, J.C.; Vega-Morales, T.; Gericke, N. Central nervous system activities of extract Mangifera indica L. J. Ethnopharmacol. 2020, 260, 112996. [Google Scholar] [CrossRef]
- Nonato, C.D.F.A.; Leite, D.O.D.; Pereira, R.C.; Boligon, A.A.; Ribeiro-Filho, J.; Rodrigues, F.F.G.; da Costa, J.G.M. Chemical analysis and evaluation of antioxidant and antimicrobial activities of fruit fractions of Mauritia flexuosa L. f. (Arecaceae). PeerJ 2018, 2018, e5991. [Google Scholar] [CrossRef]
- Ali-Shtayeh, M.S.; Jamous, R.M.; Zaitoun, S.Y.A.; Qasem, I.B. In-vitro screening of acetylcholinesterase inhibitory activity of extracts from Palestinian indigenous flora in relation to the treatment of Alzheimer’s disease. Funct. Foods Health Dis. 2014, 4, 381–400. [Google Scholar] [CrossRef]
- Herrlinger, K.A.; Nieman, K.M.; Sanoshy, K.D.; Fonseca, B.A.; Lasrado, J.A.; Schild, A.L.; Maki, K.C.; Wesnes, K.A.; Ceddia, M.A. Spearmint Extract Improves Working Memory in Men and Women with Age-Associated Memory Impairment. J. Altern. Complement. Med. 2018, 24, 37–47. [Google Scholar] [CrossRef]
- Qu, J.; Zhou, Q.; Du, Y.; Zhang, W.; Bai, M.; Zhang, Z.; Xi, Y.; Li, Z.; Miao, J. Rutin protects against cognitive deficits and brain damage in rats with chronic cerebral hypoperfusion. Br. J. Pharmacol. 2014, 171, 3702–3715. [Google Scholar] [CrossRef]
- Wang, K.; Sun, W.; Zhang, L.; Guo, W.; Xu, J.; Liu, S.; Zhou, Z.; Zhang, Y. Oleanolic acid ameliorates Aβ25-35 injection-induced memory deficit in Alzheimer’s disease model rats by maintaining synaptic plasticity. CNS Neurol. Disord. Drug Targets 2018, 17, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Heitman, E.; Ingram, D.K. Cognitive and neuroprotective effects of chlorogenic acid. Nutr. Neurosci. 2017, 20, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Akomolafe, T.L.; Adefegha, S.A.; Adetuyi, A.O. Attenuation of cyclophosphamide-induced neurotoxicity in rat by yellow dye extract from root of Brimstone tree (Morinda lucida). Exp. Toxicol. Pathol. 2012, 64, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Elufioye, T.O.; Hameed, H.A. Cognitive-enhancing properties of Morinda lucida (Rubiaceae) and Peltophorum pterocarpum (Fabaceae) in scopolamine-induced amnesic mice. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 136–141. [Google Scholar] [CrossRef]
- Chokki, M.; Cudălbeanu, M.; Zongo, C.; Dah-Nouvlessounon, D.; Ghinea, I.O.; Furdui, B.; Raclea, R.; Savadogo, A.; Baba-Moussa, L.; Avamescu, S.M.; et al. Exploring Antioxidant and Enzymes (A-Amylase and B-Glucosidase) Inhibitory Activity of Morinda lucida and Momordica charantia Leaves from Benin. Foods 2020, 9, 434. [Google Scholar] [CrossRef]
- Elufioye, T.O.; Obuotor, E.; Agbedahunsi, J.M.; Adesanya, S.A. Acetyl and Butyrylcholinesterase Inhibiting Constituent from Morinda lucida Benth (Rubiaceae). Br. J. Pharm. Res. 2015, 6, 358–365. [Google Scholar] [CrossRef]
- Singh, B.; Sharma, R.A. Indian Morinda species: A review. Phytother. Res. 2020, 34, 924–1007. [Google Scholar] [CrossRef]
- Sachan, A.; Singh, S.; Singh, H.; Shankar, P.; Kumar, D.; Sachan, A.K.; Nath, R.; Dixi, R.K. An experimental study to evaluate the effect of Mucuna pruriens on learning and memory in mice. IJISR 2015, 4, 144–148. [Google Scholar]
- Iannello, C.; Pigni, N.B.; Antognoni, F.; Poli, F.; Maxia, A.; de Andrade, J.P.; Bastida, J. A potent acetylcholinesterase inhibitor from Pancratium illyricum L. Fitoterapia 2014, 92, 163–167. [Google Scholar] [CrossRef]
- Konstantinos, F.; Heun, R. The effects of Guarana (Paullinia cupana) supplementation on the cognitive performance of young healthy adults—A Systematic Review. Glob. Psychiatry 2019, 2, 171–182. [Google Scholar] [CrossRef]
- Heckman, M.A.; Weil, J.; Gonzalez de Mejia, E. Caffeine (1, 3, 7-trimethylxanthine) in foods: A comprehensive review on consumption, functionality, safety, and regulatory matters. J. Food Sci. 2010, 75, R77–R87. [Google Scholar] [CrossRef] [PubMed]
- Herraiz, T.; Guillén, H. Monoamine Oxidase-A Inhibition and Associated Antioxidant Activity in Plant Extracts with Potential Antidepressant Actions. Biomed Res. Int. 2018, 2018, 4810394. [Google Scholar] [CrossRef] [PubMed]
- Gruss, M.; Appenroth, D.; Flubacher, A.; Enzensperger, C.; Bock, J.; Fleck, C.; Gille, G.; Braun, K. 9-Methyl-β-carboline-induced cognitive enhancement is associated with elevated hippocampal dopamine levels and dendritic and synaptic proliferation. J. Neurochem. 2012, 121, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, Y.; Deng, G.; Wang, Y.; Qi, S.; Cheng, X.; Ma, Y.; Xie, Y.; Wang, C. Exposure characteristics of the analogous β-carboline alkaloids harmaline and harmine based on the efflux transporter of multidrug resistance protein 2. Front. Pharmacol. 2017, 8, 541. [Google Scholar] [CrossRef] [PubMed]
- Keller, S.; Polanski, W.H.; Enzensperger, C.; Reichmann, H.; Hermann, A.; Gille, G. 9-Methyl-β-carboline inhibits monoamine oxidase activity and stimulates the expression of neurotrophic factors by astrocytes. J. Neural Transm. 2020, 127, 999–1012. [Google Scholar] [CrossRef]
- Charoenteeraboon, J.; Ngamkitidechakul, C.; Soonthornchareonnon, N.; Jaijoy, K.; Sireeratawong, S. Others Antioxidant activities of the standardized water extract from fruit of Phyllanthus emblica Linn. Songklanakarin J. Sci. Technol. 2010, 32, 599–604. [Google Scholar]
- Kumar, R.; Sharma, S.; Parihar, L. Evaluation of memory enhancing potential of ethanolic extract of Terminalia belerica (EETB) aganinst scopolamine induced amnesia in Wistar rats. World J. Pharm. Pharm. Sci. 2020, 9, 1505–1515. [Google Scholar] [CrossRef]
- Joshi, H.; Parle, M. Evaluation of antiamnestic potentials of [6]-gingerol and phyllanthin in mice. Nat. Prod. Bioprospect. 2006, 2, 109–117. [Google Scholar]
- Chonpathompikunlert, P.; Wattanathorn, J.; Muchimapura, S. Piperine, the main alkaloid of Thai black pepper, protects against neurodegeneration and cognitive impairment in animal model of cognitive deficit like condition of Alzheimer’s disease. Food Chem. Toxicol. 2010, 48, 798–802. [Google Scholar] [CrossRef] [PubMed]
- Wightman, E.L.; Reay, J.L.; Haskell, C.F.; Williamson, G.; Dew, T.P.; Kennedy, D.O. Effects of resveratrol alone or in combination with piperine on cerebral blood flow parameters and cognitive performance in human subjects: A randomised, double-blind, placebo-controlled, cross-over investigation. Br. J. Nutr. 2014, 112, 203–213. [Google Scholar] [CrossRef]
- Silva, A.R.; Grosso, C.; Delerue-Matos, C.; Rocha, J.M. Comprehensive review on the interaction between natural compounds and brain receptors: Benefits and toxicity. Eur. J. Med. Chem. 2019, 174, 87–115. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-S.; Lee, D.-S.; Bae, G.-S.; Park, S.-J.; Kang, D.-G.; Lee, H.-S.; Oh, H.; Kim, Y.-C. The inhibition of JNK MAPK and NF-κB signaling by tenuifoliside A isolated from Polygala tenuifolia in lipopolysaccharide-induced macrophages is associated with its anti-inflammatory effect. Eur. J. Pharmacol. 2013, 721, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.-Z.; Huang, C.-L.; Yu, B.-Y.; Hu, Y.; Mu, L.-H.; Liu, P. Effect of Tenuifoliside A isolated from Polygala tenuifolia on the ERK and PI3K pathways in C6 glioma cells. Phytomedicine 2014, 21, 1178–1188. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-M.; Li, Z.-Y.; Hu, H.; Xu, S.-P.; Chang, Q.; Liao, Y.-H.; Pan, R.-L.; Liu, X.-M. Tenuifolin, a secondary saponin from hydrolysates of polygalasaponins, counteracts the neurotoxicity induced by Aβ25-35 peptides in vitro and in vivo. Pharmacol. Biochem. Behav. 2015, 128, 14–22. [Google Scholar] [CrossRef]
- Li, Z.; Liu, Y.; Wang, L.; Liu, X.; Chang, Q.; Guo, Z.; Liao, Y.; Pan, R.; Fan, T.-P. Memory-Enhancing Effects of the Crude Extract of Polygala tenuifolia on Aged Mice. Evid. Based Complement. Altern. Med. 2014, 2014, 392324. [Google Scholar] [CrossRef]
- Park, C.H.; Choi, S.H.; Koo, J.-W.; Seo, J.-H.; Kim, H.-S.; Jeong, S.-J.; Suh, Y.-H. Novel cognitive improving and neuroprotective activities of Polygala tenuifolia Willdenow extract, BT-11. J. Neurosci. Res. 2002, 70, 484–492. [Google Scholar] [CrossRef]
- Wu, D.; He, J.; Jiang, Y.; Yang, B. Quality analysis of Polygala tenuifolia root by ultrahigh performance liquid chromatography-tandem mass spectrometry and gas chromatography-mass spectrometry. J. Food Drug Anal. 2015, 23, 144–151. [Google Scholar] [CrossRef]
- Wu, Y.; Shi, Q.; Lei, H.; Liu, X.; Luan, L. Studies on the total synthesis of tenuifoliside B. Tetrahedron 2014, 70, 3757–3761. [Google Scholar] [CrossRef]
- Huang, H.-J.; Huang, C.-Y.; Lee, M.; Lin, J.-Y.; Hsieh-Li, H.M. Puerariae Radix prevents anxiety and cognitive deficits in mice under oligomeric Aβ-induced stress. Am. J. Chin. Med. 2019, 47, 1459–1481. [Google Scholar] [CrossRef]
- Wang, Z.; Huang, X.; Zhao, P.; Zhao, L.; Wang, Z.-Y. Catalpol inhibits amyloid-β generation through promoting α-cleavage of APP in Swedish mutant APP overexpressed N2a Cells. Front. Aging Neurosci. 2018, 10, 66. [Google Scholar] [CrossRef]
- Leong, P.K.; Chen, J.; Ko, K.M. Chapter 4—Development of Chinese herbal health products for the prevention of aging-associated diseases. In Natural Products and Drug Discovery; Mandal, S.C., Mandal, V., Konishi, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 73–104. ISBN 9780081020814. [Google Scholar]
- Shiksharthi, A.R.; Mittal, S.; Ramana, J.; Road, R.; Pradesh, H. Systematic Review of Herbals as Potential Memory Enhancers. Int. J. Res. Pharm. Biomed. Sci. 2011, 2, 918–925. [Google Scholar]
- Mao, J.; Huang, S.; Liu, S.; Feng, X.-L.; Yu, M.; Liu, J.; Sun, Y.E.; Chen, G.; Yu, Y.; Zhao, J.; et al. A herbal medicine for Alzheimer’s disease and its active constituents promote neural progenitor proliferation. Aging Cell 2015, 14, 784–796. [Google Scholar] [CrossRef]
- Panossian, A.; Wikman, G.; Sarris, J. Rosenroot (Rhodiola rosea): Traditional use, chemical composition, pharmacology and clinical efficacy. Phytomedicine 2010, 17, 481–493. [Google Scholar] [CrossRef] [PubMed]
- Cropley, M.; Banks, A.P.; Boyle, J. The effects of Rhodiola rosea L. extract on anxiety, stress, cognition and other mood symptoms. Phytother. Res. 2015, 29, 1934–1939. [Google Scholar] [CrossRef]
- Amsterdam, J.D.; Panossian, A.G. Rhodiola rosea L. as a putative botanical antidepressant. Phytomedicine 2016, 23, 770–783. [Google Scholar] [CrossRef]
- Vepsäläinen, S.; Koivisto, H.; Pekkarinen, E.; Mäkinen, P.; Dobson, G.; McDougall, G.J.; Stewart, D.; Haapasalo, A.; Karjalainen, R.O.; Tanila, H.; et al. Anthocyanin-enriched bilberry and blackcurrant extracts modulate amyloid precursor protein processing and alleviate behavioral abnormalities in the APP/PS1 mouse model of Alzheimer’s disease. J. Nutr. Biochem. 2013, 24, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.O.; Dodd, F.L.; Robertson, B.C.; Okello, E.J.; Reay, J.L.; Scholey, A.B.; Haskell, C.F. Monoterpenoid extract of sage (Salvia lavandulaefolia) with cholinesterase inhibiting properties improves cognitive performance and mood in healthy adults. J. Psychopharmacol. 2011, 25, 1088–1100. [Google Scholar] [CrossRef] [PubMed]
- Chong, C.-M.; Su, H.; Lu, J.-J.; Wang, Y. The effects of bioactive components from the rhizome of Salvia miltiorrhiza (Danshen) on the characteristics of Alzheimer’s disease. Chin. Med. 2019, 14, 19. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Houghton, P.J.; Hider, R.C.; Howes, M.-J.R. Novel diterpenoid acetylcholinesterase inhibitors from Salvia miltiorhiza. Planta Med. 2004, 70, 201–204. [Google Scholar] [CrossRef]
- Kim, S.R.; Lee, K.Y.; Koo, K.A.; Sung, S.H.; Lee, N.-G.; Kim, J.; Kim, Y.C. Four new neuroprotective iridoid glycosides from Scrophularia buergeriana roots. J. Nat. Prod. 2002, 65, 1696–1699. [Google Scholar] [CrossRef]
- Kim, S.R.; Koo, K.A.; Sung, S.H.; Ma, C.J.; Yoon, J.S.; Kim, Y.C. Iridoids from Scrophularia buergeriana attenuate glutamate-induced neurotoxicity in rat cortical cultures. J. Neurosci. Res. 2003, 74, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Sumiyoshi, E.; Matsuzaki, K.; Sugimoto, N.; Tanabe, Y.; Hara, T.; Katakura, M.; Miyamoto, M.; Mishima, S.; Shido, O. Sub-chronic consumption of dark chocolate enhances cognitive function and releases nerve growth factors: A parallel-group randomized trial. Nutrients 2019, 11, 2800. [Google Scholar] [CrossRef] [PubMed]
- Nehlig, A. The neuroprotective effects of cocoa flavanol and its influence on cognitive performance. Br. J. Clin. Pharmacol. 2013, 75, 716–727. [Google Scholar] [CrossRef] [PubMed]
- Marsh, C.E.; Carter, H.H.; Guelfi, K.J.; Smith, K.J.; Pike, K.E.; Naylor, L.H.; Green, D.J. Brachial and cerebrovascular functions are enhanced in postmenopausal women after ingestion of chocolate with a high concentration of cocoa. J. Nutr. 2017, 147, 1686–1692. [Google Scholar] [CrossRef] [PubMed]
- Occhiuto, F.; Palumbo, D.R.; Samperi, S.; Zangla, G.; Pino, A.; De Pasquale, R.; Circosta, C. The isoflavones mixture from Trifolium pratense L. protects HCN 1-A neurons from oxidative stress. Phytother. Res. 2009, 23, 192–196. [Google Scholar] [CrossRef]
- Zameer, S.; Najmi, A.K.; Vohora, D.; Akhtar, M. A review on therapeutic potentials of Trigonella foenum graecum (fenugreek) and its chemical constituents in neurological disorders: Complementary roles to its hypolipidemic, hypoglycemic, and antioxidant potential. Nutr. Neurosci. 2018, 21, 539–545. [Google Scholar] [CrossRef]
- Karcheva-Bahchevanska, D.; Lukova, P. Therapeutic effects of anthocyannins from Vaccinium genus L. Int. J. Med. Res. Pharm. 2017, 4, 4–19. [Google Scholar] [CrossRef]
- Subash, S.; Essa, M.M.; Al-Adawi, S.; Memon, M.A.; Manivasagam, T.; Akbar, M. Neuroprotective effects of berry fruits on neurodegenerative diseases. Neural Regen. Res. 2014, 9, 1557–1566. [Google Scholar] [CrossRef]
- Nyakas, C.; Felszeghy, K.; Szabó, R.; Keijser, J.N.; Luiten, P.G.M.; Szombathelyi, Z.; Tihanyi, K. Neuroprotective effects of vinpocetine and its major metabolite cis-apovincaminic acid on NMDA-induced neurotoxicity in a rat entorhinal cortex lesion model. CNS Neurosci. Ther. 2009, 15, 89–99. [Google Scholar] [CrossRef]
- Kim, J.; Seo, Y.H.; Kim, J.; Goo, N.; Jeong, Y.; Bae, H.J.; Jung, S.Y.; Lee, J.; Ryu, J.H. Casticin ameliorates scopolamine-induced cognitive dysfunction in mice. J. Ethnopharmacol. 2020, 259, 112843. [Google Scholar] [CrossRef]
- Kakkar, S.; Bais, S. A review on protocatechuic Acid and its pharmacological potential. ISRN Pharmacol. 2014, 2014, 952943. [Google Scholar] [CrossRef] [PubMed]
- Zahiruddin, S.; Basist, P.; Parveen, A.; Parveen, R.; Khan, W.; Ahmad, S. Ashwagandha in brain disorders: A review of recent developments. J. Ethnopharmacol. 2020, 257, 112876. [Google Scholar] [CrossRef] [PubMed]
- Dar, N.J. MuzamilAhmad Neurodegenerative diseases and Withania somnifera (L.): An update. J. Ethnopharmacol. 2020, 256, 112769. [Google Scholar] [CrossRef] [PubMed]
- Tandon, N.; Yadav, S.S. Safety and clinical effectiveness of Withania Somnifera (Linn.) Dunal root in human ailments. J. Ethnopharmacol. 2020, 255, 112768. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, D.; Bhattacharyya, S.; Bose, S. Efficacy and Safety of Ashwagandha (Withania somnifera (L.) Dunal) Root Extract in Improving Memory and Cognitive Functions. J. Diet. Suppl. 2017, 14, 599–612. [Google Scholar] [CrossRef]
- Gupta, M.; Kaur, G. Withania somnifera (L.) Dunal ameliorates neurodegeneration and cognitive impairments associated with systemic inflammation. BMC Complement. Altern. Med. 2019, 19, 217. [Google Scholar] [CrossRef]
- Saenghong, N.; Wattanathorn, J.; Muchimapura, S.; Tongun, T.; Piyavhatkul, N.; Banchonglikitkul, C.; Kajsongkram, T. Zingiber officinale improves cognitive function of the middle-aged healthy women. Evid. Based Complement. Altern. Med. 2012, 2012, 383062. [Google Scholar] [CrossRef]
- Wattanathorn, J.; Jittiwat, J.; Tongun, T.; Muchimapura, S.; Ingkaninan, K. Zingiber officinale Mitigates Brain Damage and Improves Memory Impairment in Focal Cerebral Ischemic Rat. Evid. Based Complement. Altern. Med. 2011, 2011, 429505. [Google Scholar] [CrossRef]
- Mao, Q.-Q.; Xu, X.-Y.; Cao, S.-Y.; Gan, R.-Y.; Corke, H.; Beta, T.; Li, H.-B. Bioactive compounds and bioactivities of Ginger (Zingiber officinale Roscoe). Foods 2019, 8, 185. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, J.-G.; Yang, W.; Xu, P.; Xiao, Y.-L.; Zhang, H.-T. 6-Gingerol attenuates LPS-induced neuroinflammation and cognitive impairment partially via suppressing astrocyte overactivation. Biomed. Pharmacother. 2018, 107, 1523–1529. [Google Scholar] [CrossRef]
- Park, E.; Ryu, M.J.; Kim, N.K.; Bae, M.H.; Seo, Y.; Kim, J.; Yeo, S.; Kanwal, M.; Choi, C.W.; Heo, J.Y.; et al. Synergistic neuroprotective effect of Schisandra chinensis and Ribes fasciculatum on neuronal cell death and scopolamine-induced cognitive impairment in rats. Int. J. Mol. Sci. 2019, 20, 4517. [Google Scholar] [CrossRef] [PubMed]
- McLennan, S.N.; Lam, A.K.; Mathias, J.L.; Koblar, S.A.; Hamilton-Bruce, M.A.; Jannes, J. Role of vasodilation in cognitive impairment. Int. J. Stroke 2011, 6, 280. [Google Scholar] [CrossRef] [PubMed]
- Hitzenberger, G.; Sommer, W.; Grandt, R. Influence of vinpocetine on warfarin-induced inhibition of coagulation. Int. J. Clin. Pharmacol. Ther. Toxicol. 1990, 28, 323–328. [Google Scholar] [PubMed]
- Wurglics, M.; Schubert-Zsilavecz, M. Hypericum perforatum: A “modern” herbal antidepressant: Pharmacokinetics of active ingredients. Clin. Pharmacokinet. 2006, 45, 449–468. [Google Scholar] [CrossRef]
- Sasaki, K.; El Omri, A.; Kondo, S.; Han, J.; Isoda, H. Rosmarinus officinalis polyphenols produce anti-depressant like effect through monoaminergic and cholinergic functions modulation. Behav. Brain Res. 2013, 238, 86–94. [Google Scholar] [CrossRef]
- Bak, L.K.; Schousboe, A.; Waagepetersen, H.S. The glutamate/GABA-glutamine cycle: Aspects of transport, neurotransmitter homeostasis and ammonia transfer. J. Neurochem. 2006, 98, 641–653. [Google Scholar] [CrossRef]
- Hajirahimkhan, A.; Dietz, B.M.; Bolton, J.L. Botanical modulation of menopausal symptoms: Mechanisms of action? Planta Med. 2013, 79, 538–553. [Google Scholar] [CrossRef]
- Camfield, D.A.; Stough, C.; Farrimond, J.; Scholey, A.B. Acute effects of tea constituents L-theanine, caffeine, and epigallocatechin gallate on cognitive function and mood: A systematic review and meta-analysis. Nutr. Rev. 2014, 72, 507–522. [Google Scholar] [CrossRef]
- Liu, R.H. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr. 2004, 134, 3479S–3485S. [Google Scholar] [CrossRef]
- Campos-Vega, R.; Oomah, B.D. Chemistry and classification of phytochemicals. In Handbook of Plant Food Phytochemicals; Tiwari, B.K., Brunton, N.P., Brennan, C.S., Eds.; John Wiley & Sons Ltd.: Oxford, UK, 2013; pp. 5–48. ISBN 9781118464717. [Google Scholar]
- Giada, M.D.L.R. Food Phenolic Compounds: Main Classes, Sources and Their Antioxidant Power. In Oxidative Stress and Chronic Degenerative Diseases; Morales-González, J.A., Ed.; IntechOpen: Rijeka, Croatia, 2013; pp. 87–112. [Google Scholar]
- Liu, R.H. Health-promoting components of fruits and vegetables in the diet. Adv. Nutr. 2013, 4, 384S–392S. [Google Scholar] [CrossRef]
- Verpoorte, R. Alkaloids. In Encyclopedia of Analytical Science, 2nd ed.; Worsfold, P., Townshend, A., Poole, C., Eds.; Elsevier: Oxford, UK, 2005; pp. 56–61. ISBN 9780123693976. [Google Scholar]
- Debnath, B.; Singh, W.S.; Das, M.; Goswami, S.; Singh, M.K.; Maiti, D.; Manna, K. Role of plant alkaloids on human health: A review of biological activities. Mater. Today Chem. 2018, 9, 56–72. [Google Scholar] [CrossRef]
- Konrath, E.L.; Passos, C.D.S.; Klein, L.C., Jr.; Henriques, A.T. Alkaloids as a source of potential anticholinesterase inhibitors for the treatment of Alzheimer’s disease. J. Pharm. Pharmacol. 2013, 65, 1701–1725. [Google Scholar] [CrossRef] [PubMed]
- Brahmkshatriya, P.P.; Brahmkshatriya, P.S. Terpenes: Chemistry, Biological Role, and Therapeutic Applications. In Natural Products: Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 2665–2691. ISBN 9783642221446. [Google Scholar]
- Pearlson, G. Chapter 9—Chemistry, chemical analysis, and extraction. Terpenes to tinctures. In Weed Science; Academic Press: Cambridge, MA, USA, 2020; pp. 217–242. ISBN 9780128181744. [Google Scholar]
- Agus, H.H. Chapter 4—Terpene toxicity and oxidative stress. In Toxicology: Oxidative Stress and Dietary Antioxidants; Patel, V.B., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 33–42. ISBN 9780128190937. [Google Scholar]
- Bell, L.; Lamport, D.J.; Butler, L.T.; Williams, C.M. A review of the cognitive effects observed in humans following acute supplementation with flavonoids, and their associated mechanisms of action. Nutrients 2015, 7, 10290–10306. [Google Scholar] [CrossRef] [PubMed]
- Nouri, Z.; Fakhri, S.; El-Senduny, F.F.; Sanadgol, N.; Abd-Elghani, G.E.; Farzaei, M.H.; Chen, J.T. On the neuroprotective effects of naringenin: Pharmacological targets, signaling pathways, molecular mechanisms, and clinical perspective. Biomolecules 2019, 9, 690. [Google Scholar] [CrossRef]
- Tutunchi, H.; Naeini, F.; Ostadrahimi, A.; Hosseinzadeh-Attar, M.J. Naringenin, a flavanone with antiviral and anti-inflammatory effects: A promising treatment strategy against COVID-19. Phytother. Res. 2020, 34, 3137–3147. [Google Scholar] [CrossRef]
- Umukoro, S.; Kalejaye, H.A.; Ben-Azu, B.; Ajayi, A.M. Naringenin attenuates behavioral derangements induced by social defeat stress in mice via inhibition of acetylcholinesterase activity, oxidative stress and release of pro-inflammatory cytokines. Biomed. Pharmacother. 2018, 105, 714–723. [Google Scholar] [CrossRef]
- Piironen, V.; Lampi, A.-M. Chapter 9—Rye as a Source of Phytosterols, Tocopherols, and Tocotrienols. In Rye and Health; Poutanen, K., Åman, P., Eds.; AACC International Press: Washington, DC, USA, 2014; pp. 131–158. ISBN 9781891127816. [Google Scholar]
- Izzo, A.A. Interactions between herbs and conventional drugs: Overview of the clinical data. Med. Princ. Pract. 2012, 21, 404–428. [Google Scholar] [CrossRef]
- Ondieki, G.; Nyagblordzro, M.; Kikete, S.; Liang, R.; Wang, L.; He, X. Cytochrome P450 and P-Glycoprotein-Mediated Interactions Involving African Herbs Indicated for Common Noncommunicable Diseases. Evid. Based Complement. Altern. Med. 2017, 2017, 2582463. [Google Scholar] [CrossRef]
- Zhou, S.; Gao, Y.; Jiang, W.; Huang, M.; Xu, A.; Paxton, J.W. Interactions of herbs with cytochrome P450. Drug Metab. Rev. 2003, 35, 35–98. [Google Scholar] [CrossRef]
- Cermak, R. Effect of dietary flavonoids on pathways involved in drug metabolism. Expert Opin. Drug Metab. Toxicol. 2008, 4, 17–35. [Google Scholar] [CrossRef]
- Liang, Y.; Li, S.; Chen, L. The physiological role of drug transporters. Protein Cell 2015, 6, 334–350. [Google Scholar] [CrossRef] [PubMed]
- Grimstein, M.; Huang, S.-M. A regulatory science viewpoint on botanical-drug interactions. J. Food Drug Anal. 2018, 26, S12–S25. [Google Scholar] [CrossRef] [PubMed]
- Roth, M.; Obaidat, A.; Hagenbuch, B. OATPs, OATs and OCTs: The organic anion and cation transporters of the SLCO and SLC22A gene superfamilies. Br. J. Pharmacol. 2012, 165, 1260–1287. [Google Scholar] [CrossRef] [PubMed]
- Zamek-Gliszczynski, M.J.; Taub, M.E.; Chothe, P.P.; Chu, X.; Giacomini, K.M.; Kim, R.B.; Ray, A.S.; Stocker, S.L.; Unadkat, J.D.; Wittwer, M.B.; et al. Transporters in Drug Development: 2018 ITC Recommendations for Transporters of Emerging Clinical Importance. Clin. Pharmacol. Ther. 2018, 104, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, F.A. Cyclooxygenase enzymes: Regulation and function. Curr. Pharm. Des. 2004, 10, 577–588. [Google Scholar] [CrossRef]
- Churihar, R.; Solanki, P.; Vyas, S.; Hemant Tanwani, H.; Shubham Atal, S. Analgesic activity of cinnamaldehyde per se and it’s interaction with diclofenac sodium and pentazocine in Swiss albino mice. Int. J. Phamacog. 2016, 3, 97–102. [Google Scholar] [CrossRef]
- Finberg, J.P.M.; Rabey, J.M. Inhibitors of MAo-A and MAo-B in Psychiatry and Neurology. Front. Pharmacol. 2016, 7, 340. [Google Scholar] [CrossRef]
- Hritcu, L.; Ionita, R.; Postu, P.A.; Gupta, G.K.; Turkez, H.; Lima, T.C.; Carvalho, C.U.S.; de Sousa, D.P. Antidepressant Flavonoids and Their Relationship with Oxidative Stress. Oxid. Med. Cell. Longev. 2017, 2017, 5762172. [Google Scholar] [CrossRef]
- Jewett, B.E.; Thapa, B. Physiology, NMDA Receptor. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Page, A.T.; Falster, M.O.; Litchfield, M.; Pearson, S.-A.; Etherton-Beer, C. Polypharmacy among older Australians, 2006–2017: A population-based study. Med. J. Aust. 2019, 211, 71–75. [Google Scholar] [CrossRef]
- National Center for Health Statistics (US). Health, United States, 2018; National Center for Health Statistics (US): Hyattsville, MD, USA, 2019.
- Nies, A.T.; Koepsell, H.; Damme, K.; Schwab, M. Organic cation transporters (OCTs, MATEs), in vitro and in vivo evidence for the importance in drug therapy. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2011; Volume 201, pp. 105–167. [Google Scholar] [CrossRef]
- Koepsell, H. Organic Cation Transporters in Health and Disease. Pharmacol. Rev. 2020, 72, 253–319. [Google Scholar] [CrossRef]
- Koepsell, H. Role of organic cation transporters in drug-drug interaction. Expert Opin. Drug Metab. Toxicol. 2015, 11, 1619–1633. [Google Scholar] [CrossRef] [PubMed]
- Benet, L.Z.; Cummins, C.L.; Wu, C.Y. Transporter-enzyme interactions: Implications for predicting drug-drug interactions from in vitro data. Curr. Drug Metab. 2003, 4, 393–398. [Google Scholar] [CrossRef]
- Liu, X. Transporter-Mediated Drug-Drug Interactions and Their Significance. Adv. Exp. Med. Biol. 2019, 1141, 241–291. [Google Scholar] [CrossRef] [PubMed]
- Smolders, E.J.; de Kanter, C.T.M.M.; de Knegt, R.J.; van der Valk, M.; Drenth, J.P.H.; Burger, D.M. Drug–Drug Interactions Between Direct-Acting Antivirals and Psychoactive Medications. Clin. Pharmacokinet. 2016, 55, 1471–1494. [Google Scholar] [CrossRef] [PubMed]
- DrugBank Online. Available online: https://go.drugbank.com/ (accessed on 23 November 2020).
- Zhou, S.-F.; Zhou, Z.-W.; Yang, L.-P.; Cai, J.-P. Substrates, inducers, inhibitors and structure-activity relationships of human Cytochrome P450 2C9 and implications in drug development. Curr. Med. Chem. 2009, 16, 3480–3675. [Google Scholar] [CrossRef]
- Vildhede, A.; Karlgren, M.; Svedberg, E.K.; Wisniewski, J.R.; Lai, Y.; Norén, A.; Artursson, P. Hepatic uptake of atorvastatin: Influence of variability in transporter expression on uptake clearance and drug-drug interactions. Drug Metab. Dispos. 2014, 42, 1210–1218. [Google Scholar] [CrossRef]
- Elsby, R.; Martin, P.; Surry, D.; Sharma, P.; Fenner, K. Solitary Inhibition of the Breast Cancer Resistance Protein Efflux Transporter Results in a Clinically Significant Drug-Drug Interaction with Rosuvastatin by Causing up to a 2-Fold Increase in Statin Exposure. Drug Metab. Dispos. 2016, 44, 398–408. [Google Scholar] [CrossRef]
- Shugarts, S.; Benet, L.Z. The role of transporters in the pharmacokinetics of orally administered drugs. Pharm. Res. 2009, 26, 2039–2054. [Google Scholar] [CrossRef]
- Clarke, T.A.; Waskell, L.A. The metabolism of clopidogrel is catalyzed by human cytochrome P450 3A and is inhibited by atorvastatin. Drug Metab. Dispos. 2003, 31, 53–59. [Google Scholar] [CrossRef]
- Kim, S.-J.; Yoshikado, T.; Ieiri, I.; Maeda, K.; Kimura, M.; Irie, S.; Kusuhara, H.; Sugiyama, Y. Clarification of the Mechanism of Clopidogrel-Mediated Drug-Drug Interaction in a Clinical Cassette Small-dose Study and Its Prediction Based on In Vitro Information. Drug Metab. Dispos. 2016, 44, 1622–1632. [Google Scholar] [CrossRef]
- Varma, M.V.S.; Bi, Y.-A.; Lazzaro, S.; West, M. Clopidogrel as a Perpetrator of Drug-Drug Interactions: A Challenge for Quantitative Predictions? Clin. Pharmacol. Ther. 2019, 105, 1295–1299. [Google Scholar] [CrossRef] [PubMed]
- Lagas, J.S.; van der Kruijssen, C.M.M.; van de Wetering, K.; Beijnen, J.H.; Schinkel, A.H. Transport of diclofenac by breast cancer resistance protein (ABCG2) and stimulation of multidrug resistance protein 2 (ABCC2)-mediated drug transport by diclofenac and benzbromarone. Drug Metab. Dispos. 2009, 37, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Kindla, J.; Müller, F.; Mieth, M.; Fromm, M.F.; König, J. Influence of non-steroidal anti-inflammatory drugs on organic anion transporting polypeptide (OATP) 1B1- and OATP1B3-mediated drug transport. Drug Metab. Dispos. 2011, 39, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Müller, F.; Weitz, D.; Derdau, V.; Sandvoss, M.; Mertsch, K.; König, J.; Fromm, M.F. Contribution of MATE1 to Renal Secretion of the NMDA Receptor Antagonist Memantine. Mol. Pharm. 2017, 14, 2991–2998. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.I.; Betterton, R.D.; Davis, T.P.; Ronaldson, P.T. Transporter-Mediated Delivery of Small Molecule Drugs to the Brain: A Critical Mechanism That Can Advance Therapeutic Development for Ischemic Stroke. Pharmaceutics 2020, 12, 154. [Google Scholar] [CrossRef]
- Beconi, M.G.; Howland, D.; Park, L.; Lyons, K.; Giuliano, J.; Dominguez, C.; Munoz-Sanjuan, I.; Pacifici, R. Pharmacokinetics of memantine in rats and mice. PLoS Curr. 2011, 3, RRN1291. [Google Scholar] [CrossRef]
- Li, Y.; Meng, Q.; Yang, M.; Liu, D.; Hou, X.; Tang, L.; Wang, X.; Lyu, Y.; Chen, X.; Liu, K.; et al. Current trends in drug metabolism and pharmacokinetics. Acta Pharm. Sin. B 2019, 9, 1113–1144. [Google Scholar] [CrossRef]
- Lee, J.O.; Lee, S.K.; Kim, J.H.; Kim, N.; You, G.Y.; Moon, J.W.; Kim, S.J.; Park, S.H.; Kim, H.S. Metformin regulates glucose transporter 4 (GLUT4) translocation through AMP-activated protein kinase (AMPK)-mediated Cbl/CAP signaling in 3T3-L1 preadipocyte cells. J. Biol. Chem. 2012, 287, 44121–44129. [Google Scholar] [CrossRef]
- Kim, J.; Yang, G.; Kim, Y.; Kim, J.; Ha, J. AMPK activators: Mechanisms of action and physiological activities. Exp. Mol. Med. 2016, 48, e224. [Google Scholar] [CrossRef]
- Hemauer, S.J.; Patrikeeva, S.L.; Nanovskaya, T.N.; Hankins, G.D.V.; Ahmed, M.S. Role of human placental apical membrane transporters in the efflux of glyburide, rosiglitazone, and metformin. Am. J. Obstet. Gynecol. 2010, 202, 383-e1. [Google Scholar] [CrossRef]
- Gong, L.; Goswami, S.; Giacomini, K.M.; Altman, R.B.; Klein, T.E. Metformin pathways: Pharmacokinetics and pharmacodynamics. Pharmacogenet. Genom. 2012, 22, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Alsarrani, A.; Kaplita, P.V. In Silico and in vitro evaluation of brain penetration properties of selected nootropic agents. Future Drug Discov. 2019, 1, FDD9. [Google Scholar] [CrossRef]
- Masubuchi, Y.; Hosokawa, S.; Horie, T.; Suzuki, T.; Ohmori, S.; Kitada, M.; Narimatsu, S. Cytochrome P450 isozymes involved in propranolol metabolism in human liver microsomes. The role of CYP2D6 as ring-hydroxylase and CYP1A2 as N-desisopropylase. Drug Metab. Dispos. 1994, 22, 909–915. [Google Scholar] [PubMed]
- Dudley, A.J.; Bleasby, K.; Brown, C.D. The organic cation transporter OCT2 mediates the uptake of beta-adrenoceptor antagonists across the apical membrane of renal LLC-PK(1) cell monolayers. Br. J. Pharmacol. 2000, 131, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Steiner, A.; Walle, T. Potent inhibition of MAO mediated propranolol metabolism by dimethyl sulfoxide in Hep G2 cells. Res. Commun. Chem. Pathol. Pharmacol. 1992, 75, 373–376. [Google Scholar] [PubMed]
- Goldberg, M.R.; Sciberras, D.; De Smet, M.; Lowry, R.; Tomasko, L.; Lee, Y.; Olah, T.V.; Zhao, J.; Vyas, K.P.; Halpin, R.; et al. Influence of beta-adrenoceptor antagonists on the pharmacokinetics of rizatriptan, a 5-HT1B/1D agonist: Differential effects of propranolol, nadolol and metoprolol. Br. J. Clin. Pharmacol. 2001, 52, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Obach, R.S.; Cox, L.M.; Tremaine, L.M. Sertraline is metabolized by multiple cytochrome P450 enzymes, monoamine oxidases, and glucuronyl transferases in human: An in vitro study. Drug Metab. Dispos. 2005, 33, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, C.U.; Frølund, S.; Abdulhadi, S.; Sari, H.; Langthaler, L.; Nøhr, M.K.; Kall, M.A.; Brodin, B.; Holm, R. Sertraline inhibits the transport of PAT1 substrates in vivo and in vitro. Br. J. Pharmacol. 2013, 170, 1041–1052. [Google Scholar] [CrossRef]
- Feng, S.; Zheng, L.; Tang, S.; Gu, J.; Jiang, X.; Wang, L. In-vitro and in situ assessment of the efflux of five antidepressants by breast cancer resistance protein. J. Pharm. Pharmacol. 2019, 71, 1133–1141. [Google Scholar] [CrossRef]
- Ring, B.J.; Patterson, B.E.; Mitchell, M.I.; Vandenbranden, M.; Gillespie, J.; Bedding, A.W.; Jewell, H.; Payne, C.D.; Forgue, S.T.; Eckstein, J.; et al. Effect of tadalafil on cytochrome P450 3A4-mediated clearance: Studies in vitro and in vivo. Clin. Pharmacol. Ther. 2005, 77, 63–75. [Google Scholar] [CrossRef]
- Kopečná-Zapletalová, M.; Krasulová, K.; Anzenbacher, P.; Hodek, P.; Anzenbacherová, E. Interaction of isoflavonoids with human liver microsomal cytochromes P450: Inhibition of CYP enzyme activities. Xenobiotica 2017, 47, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Chao, W.-W.; Lin, B.-F. Bioactivities of major constituents isolated from Angelica sinensis (Danggui). Chin. Med. 2011, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dewanjee, S.; Dua, T.K.; Bhattacharjee, N.; Das, A.; Gangopadhyay, M.; Khanra, R.; Joardar, S.; Riaz, M.; De Feo, V.; Zia-Ul-Haq, M. Natural products as alternative choices for P-glycoprotein (P-gp) inhibition. Molecules 2017, 22, 871. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wolkoff, A.W.; Morris, M.E. Flavonoids as a novel class of human organic anion-transporting polypeptide OATP1B1 (OATP-C) modulators. Drug Metab. Dispos. 2005, 33, 1666–1672. [Google Scholar] [CrossRef] [PubMed]
- Šarić Mustapić, D.; Debeljak, Ž.; Maleš, Ž.; Bojić, M. The Inhibitory Effect of Flavonoid Aglycones on the Metabolic Activity of CYP3A4 Enzyme. Molecules 2018, 23, 2553. [Google Scholar] [CrossRef]
- Mandery, K.; Bujok, K.; Schmidt, I.; Keiser, M.; Siegmund, W.; Balk, B.; König, J.; Fromm, M.F.; Glaeser, H. Influence of the flavonoids apigenin, kaempferol, and quercetin on the function of organic anion transporting polypeptides 1A2 and 2B1. Biochem. Pharmacol. 2010, 80, 1746–1753. [Google Scholar] [CrossRef]
- Chan, T.; Li, Z.; Zheng, J.; Cheung, F.S.G.; Zhu, L.; Zhou, F. Inhibitory effects of apigenin and kaempferol on the essential solute carrier transporters. World J. Gastrointest. Pharmacol. Ther. 2013, 2, 115–121. [Google Scholar] [CrossRef]
- Saeed, M.; Kadioglu, O.; Khalid, H.; Sugimoto, Y.; Efferth, T. Activity of the dietary flavonoid, apigenin, against multidrug-resistant tumor cells as determined by pharmacogenomics and molecular docking. J. Nutr. Biochem. 2015, 26, 44–56. [Google Scholar] [CrossRef]
- Mandery, K.; Balk, B.; Bujok, K.; Schmidt, I.; Fromm, M.F.; Glaeser, H. Inhibition of hepatic uptake transporters by flavonoids. Eur. J. Pharm. Sci. 2012, 46, 79–85. [Google Scholar] [CrossRef]
- Meng, X.; Liao, S.; Wang, X.; Wang, S.; Zhao, X.; Jia, P.; Pei, W.; Zheng, X.; Zheng, X. Reversing P-glycoprotein-mediated multidrug resistance in vitro by α-asarone and β-asarone, bioactive cis-trans isomers from Acorus tatarinowii. Biotechnol. Lett. 2014, 36, 685–691. [Google Scholar] [CrossRef]
- Liu, H.-J.; Lai, X.; Xu, Y.; Miao, J.-K.; Li, C.; Liu, J.-Y.; Hua, Y.-Y.; Ma, Q.; Chen, Q. α-Asarone Attenuates Cognitive Deficit in a Pilocarpine-Induced Status Epilepticus Rat Model via a Decrease in the Nuclear Factor-κB Activation and Reduction in Microglia Neuroinflammation. Front. Neurol. 2017, 8, 661. [Google Scholar] [CrossRef] [PubMed]
- Cartus, A.T.; Schrenk, D. Metabolism of carcinogenic alpha-asarone by human cytochrome P450 enzymes. Naunyn. Schmiedebergs. Arch. Pharmacol. 2020, 393, 213–223. [Google Scholar] [CrossRef]
- Pandit, S.; Mukherjee, P.K.; Ponnusankar, S.; Venkatesh, M.; Srikanth, N. Metabolism mediated interaction of α-asarone and Acorus calamus with CYP3A4 and CYP2D6. Fitoterapia 2011, 82, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Páez, L.; Juárez-Sanchez, M.; Antúnez-Solís, J.; Baeza, I.; Wong, C. Alpha-asarone inhibits HMG-CoA reductase, lowers serum LDL-cholesterol levels and reduces biliary CSI in hypercholesterolemic rats. Phytomedicine 2003, 10, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Chellian, R.; Pandy, V.; Mohamed, Z. Pharmacology and toxicology of α- and β-Asarone: A review of preclinical evidence. Phytomedicine 2017, 32, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.-W.; Cheong, Y.-J.; Koo, Y.-M.; Kim, S.; Noh, C.-K.; Son, Y.-H.; Kang, C.; Sohn, N.-W. α-Asarone Ameliorates Memory Deficit in Lipopolysaccharide-Treated Mice via Suppression of Pro-Inflammatory Cytokines and Microglial Activation. Biomol. Ther. 2014, 22, 17–26. [Google Scholar] [CrossRef]
- Das, B.K.; Swamy, A.V.; Koti, B.C.; Gadad, P.C. Experimental evidence for use of Acorus calamus (asarone) for cancer chemoprevention. Heliyon 2019, 5, e01585. [Google Scholar] [CrossRef]
- Guo, L.; Cui, Y.; Hao, K. Effects of glycyrrhizin on the pharmacokinetics of asiatic acid in rats and its potential mechanism. Pharm. Biol. 2018, 56, 119–123. [Google Scholar] [CrossRef]
- Pan, Y.; Abd-Rashid, B.A.; Ismail, Z.; Ismail, R.; Mak, J.W.; Pook, P.C.K.; Er, H.M.; Ong, C.E. In vitro modulatory effects on three major human cytochrome P450 enzymes by multiple active constituents and extracts of Centella asiatica. J. Ethnopharmacol. 2010, 130, 275–283. [Google Scholar] [CrossRef]
- Wright, K.M.; Magana, A.A.; Laethem, R.M.; Moseley, C.L.; Banks, T.T.; Maier, C.S.; Stevens, J.F.; Quinn, J.F.; Soumyanath, A. Centella asiatica Water Extract Shows Low Potential for Cytochrome P450-Mediated Drug Interactions. Drug Metab. Dispos. 2020, 48, 1053–1063. [Google Scholar] [CrossRef]
- Cheng, Q.; Liao, M.; Hu, H.; Li, H.; Wu, L. Asiatic Acid (AA) Sensitizes Multidrug-Resistant Human Lung Adenocarcinoma A549/DDP Cells to Cisplatin (DDP) via Downregulation of P-Glycoprotein (MDR1) and Its Targets. Cell. Physiol. Biochem. 2018, 47, 279–292. [Google Scholar] [CrossRef]
- Zhang, Q.; Cao, Y.F.; Ran, R.X.; Li, R.S.; Wu, X.; Dong, P.P.; Zhang, Y.Y.; Hu, C.M.; Wang, W.M. Strong Specific Inhibition of UDP-glucuronosyltransferase 2B7 by Atractylenolide I and III. Phytother. Res. 2016, 30, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Gul, S.; Gul, H.; Bangash, M.H. Anti-inflammatory and anti-platelet activities of Avena sativa are mediated through the inhibition of cyclooxygenase and lipoxygenase enzymes. IJEHSR 2013, 1, 62–65. [Google Scholar] [CrossRef]
- Scarpa, E.S.; Mari, M.; Antonini, E.; Palma, F.; Ninfali, P. Natural and synthetic avenanthramides activate caspases 2, 8, 3 and downregulate hTERT, MDR1 and COX-2 genes in CaCo-2 and Hep3B cancer cells. Food Funct. 2018, 9, 2913–2921. [Google Scholar] [CrossRef]
- Ramasamy, V.S.; Samidurai, M.; Park, H.J.; Wang, M.; Park, R.Y.; Yu, S.Y.; Kang, H.K.; Hong, S.; Choi, W.-S.; Lee, Y.Y.; et al. Avenanthramide-C Restores Impaired Plasticity and Cognition in Alzheimer’s Disease Model Mice. Mol. Neurobiol. 2020, 57, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, S.; Kiew, L.V.; Chung, L.Y. Inhibition of human cytochrome P450 enzymes by Bacopa monnieri standardized extract and constituents. Molecules 2014, 19, 2588–2601. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, L.; Xie, L.; Gan, D.; Jiang, X. Effects of berberine on the pharmacokinetics of losartan and its metabolite EXP3174 in rats and its mechanism. Pharm. Biol. 2016, 54, 2886–2894. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.-L.; Yu, H.-L.; Li, D.; Meng, X.-L.; Zhou, Z.-Y.; Yu, Q.; Zhang, X.-Y.; Wang, F.-J.; Guo, C. In vitro inhibition of Huanglian [Rhizoma coptidis (L.)] and its six active alkaloids on six cytochrome P450 isoforms in human liver microsomes. Phytother. Res. 2011, 25, 1660–1665. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Qiu, F.; Jiang, J.; Gao, C.; Tan, Y. Intestinal absorption mechanisms of berberine, palmatine, jateorhizine, and coptisine: Involvement of P-glycoprotein. Xenobiotica 2011, 41, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-P.; Huang, C.-Y.; Lin, S.-P.; Hou, Y.-C. Activation of P-glycoprotein and CYP 3A by Coptidis Rhizoma in vivo: Using cyclosporine as a probe substrate in rats. J. Food Drug Anal. 2018, 26, S125–S132. [Google Scholar] [CrossRef]
- Min, Y.D.; Yang, M.C.; Lee, K.H.; Kim, K.R.; Choi, S.U.; Lee, K.R. Protoberberine alkaloids and their reversal activity of P-gp expressed multidrug resistance (MDR) from the rhizome of Coptis japonica Makino. Arch. Pharm. Res. 2006, 29, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.W.; Li, Y.; Paxton, J.W.; Birch, N.P.; Scheepens, A. Identification of novel dietary phytochemicals inhibiting the efflux transporter breast cancer resistance protein (BCRP/ABCG2). Food Chem. 2013, 138, 2267–2274. [Google Scholar] [CrossRef]
- Chen, C.; Wu, Z.-T.; Ma, L.-L.; Ni, X.; Lin, Y.-F.; Wang, L.; Chen, K.-P.; Huang, C.-G.; Pan, G. Organic anion-transporting polypeptides contribute to the hepatic uptake of berberine. Xenobiotica 2015, 45, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Obach, R.S. Inhibition of human cytochrome P450 enzymes by constituents of St. John’s Wort, an herbal preparation used in the treatment of depression. J. Pharmacol. Exp. Ther. 2000, 294, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Gutmann, H.; Bruggisser, R.; Schaffner, W.; Bogman, K.; Botomino, A.; Drewe, J. Transport of amentoflavone across the blood-brain barrier in vitro. Planta Med. 2002, 68, 804–807. [Google Scholar] [CrossRef] [PubMed]
- Umegaki, K.; Taki, Y.; Endoh, K.; Taku, K.; Tanabe, H.; Shinozuka, K.; Sugiyama, T. Bilobalide in Ginkgo biloba extract is a major substance inducing hepatic CYPs. J. Pharm. Pharmacol. 2007, 59, 871–877. [Google Scholar] [CrossRef]
- Yaro, P.; Nie, J.; Xu, M.; Zeng, K.; He, H.; Yao, J.; Wang, R.; Zeng, S. Influence of organic anion transporter 1/3 on the pharmacokinetics and renal excretion of ginkgolides and bilobalide. J. Ethnopharmacol. 2019, 243, 112098. [Google Scholar] [CrossRef]
- Weichel, O.; Hilgert, M.; Chatterjee, S.S.; Lehr, M.; Klein, J. Bilobalide, a constituent of Ginkgo biloba, inhibits NMDA-induced phospholipase A2 activation and phospholipid breakdown in rat hippocampus. Naunyn. Schmiedebergs. Arch. Pharmacol. 1999, 360, 609–615. [Google Scholar] [CrossRef]
- Srinivas, N.R. Biochanin A: Understanding the complexities in the paradoxical drug-drug interaction potential. Eur. J. Drug Metab. Pharmacokinet. 2015, 40, 119–125. [Google Scholar] [CrossRef]
- Zhang, S.; Morris, M.E. Effects of the flavonoids biochanin A, morin, phloretin, and silymarin on P-glycoprotein-mediated transport. J. Pharmacol. Exp. Ther. 2003, 304, 1258–1267. [Google Scholar] [CrossRef]
- Bircsak, K.M.; Aleksunes, L.M. Interaction of Isoflavones with the BCRP/ABCG2 Drug Transporter. Curr. Drug Metab. 2015, 16, 124–140. [Google Scholar] [CrossRef] [PubMed]
- Sissung, T.M.; Baum, C.E.; Kirkland, C.T.; Gao, R.; Gardner, E.R.; Figg, W.D. Pharmacogenetics of membrane transporters: An update on current approaches. Mol. Biotechnol. 2010, 44, 152–167. [Google Scholar] [CrossRef] [PubMed]
- An, G.; Morris, M.E. The sulfated conjugate of biochanin A is a substrate of breast cancer resistant protein (ABCG2). Biopharm. Drug Dispos. 2011, 32, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Armutcu, F.; Akyol, S.; Ustunsoy, S.; Turan, F.F. Therapeutic potential of caffeic acid phenethyl ester and its anti-inflammatory and immunomodulatory effects (Review). Exp. Ther. Med. 2015, 9, 1582–1588. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, H.; Jana, S. Evaluation of Inhibitory Effects of Caffeic acid and Quercetin on Human Liver Cytochrome P450 Activities. Phytother. Res. 2014, 28, 1873–1878. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.-N.; Wang, C.C.N.; Liao, W.-C.; Lan, Y.-H.; Hung, C.-C. Caffeic Acid Attenuates Multi-Drug Resistance in Cancer Cells by Inhibiting Efflux Function of Human P-glycoprotein. Molecules 2020, 25, 247. [Google Scholar] [CrossRef]
- Hong, Y.-J.; Yang, S.-Y.; Nam, M.-H.; Koo, Y.-C.; Lee, K.-W. Caffeic acid inhibits the uptake of 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP) by inducing the efflux transporters expression in Caco-2 cells. Biol. Pharm. Bull. 2015, 38, 201–207. [Google Scholar] [CrossRef]
- Mora, F.; Molina, J.D.; Zubillaga, E.; López-Muñoz, F.; Álamo, C. CYP450 and Its Implications in the Clinical Use of Antipsychotic Drugs. Clin. Exp. Pharmacol. 2015, 5, 1000176. [Google Scholar] [CrossRef]
- Ding, R.; Shi, J.; Pabon, K.; Scotto, K.W. Xanthines Down-Regulate the Drug Transporter ABCG2 and Reverse Multidrug Resistance. Mol. Pharmacol. 2012, 81, 328–337. [Google Scholar] [CrossRef]
- Cao, R.; Peng, W.; Wang, Z.; Xu, A. beta-Carboline alkaloids: Biochemical and pharmacological functions. Curr. Med. Chem. 2007, 14, 479–500. [Google Scholar] [CrossRef]
- Passos, C.D.S.; Simoes-Pires, C.; Henriques, A.; Cuendet, M.; Carrupt, P.-A.; Christen, P. Chapter 4—Alkaloids as Inhibitors of Monoamine Oxidases and Their Role in the Central Nervous System. In Studies in Natural Products Chemistry; Rahman, A.U., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 43, pp. 123–144. [Google Scholar]
- Ma, Y.; Wink, M. The beta-carboline alkaloid harmine inhibits BCRP and can reverse resistance to the anticancer drugs mitoxantrone and camptothecin in breast cancer cells. Phytother. Res. 2010, 24, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; He, Y.-Q.; Wang, J.; Ding, K.-M.; Wang, C.-H.; Wang, Z.-T. Inhibition of human cytochrome P450 enzymes 3A4 and 2D6 by β-carboline alkaloids, harmine derivatives. Phytother. Res. 2011, 25, 1671–1677. [Google Scholar] [CrossRef] [PubMed]
- Mohos, V.; Bencsik, T.; Boda, G.; Fliszár-Nyúl, E.; Lemli, B.; Kunsági-Máté, S.; Poór, M. Interactions of casticin, ipriflavone, and resveratrol with serum albumin and their inhibitory effects on CYP2C9 and CYP3A4 enzymes. Biomed. Pharmacother. 2018, 107, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chen, K.; Lu, Y.; Fang, Z.; Yu, G. Catalpol provides a protective effect on fibrillary Aβ1-42 -induced barrier disruption in an in vitro model of the blood-brain barrier. Phytother. Res. 2018, 32, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Bao, Q.; Shen, X.; Qian, L.; Gong, C.; Nie, M.; Dong, Y. Anti-diabetic activities of catalpol in db/db mice. Korean J. Physiol. Pharmacol. 2016, 20, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Cao, X.; Li, T.; Li, X. Effects of catalpol on the activity of human liver cytochrome P450 enzymes. Xenobiotica 2019, 49, 1289–1295. [Google Scholar] [CrossRef]
- Knop, J.; Misaka, S.; Singer, K.; Hoier, E.; Müller, F.; Glaeser, H.; König, J.; Fromm, M.F. Inhibitory Effects of Green Tea and (-)-Epigallocatechin Gallate on Transport by OATP1B1, OATP1B3, OCT1, OCT2, MATE1, MATE2-K and P-Glycoprotein. PLoS ONE 2015, 10, e0139370. [Google Scholar] [CrossRef]
- Satoh, T.; Fujisawa, H.; Nakamura, A.; Takahashi, N.; Watanabe, K. Inhibitory effects of eight green tea catechins on cytochrome P450 1A2, 2C9, 2D6, and 3A4 activities. J. Pharm. Pharm. Sci. 2016, 19, 188–197. [Google Scholar] [CrossRef]
- Reddy, D.B.; Reddy, T.C.M.; Jyotsna, G.; Sharan, S.; Priya, N.; Lakshmipathi, V.; Reddanna, P. Chebulagic acid, a COX-LOX dual inhibitor isolated from the fruits of Terminalia chebula Retz., induces apoptosis in COLo-205 cell line. J. Ethnopharmacol. 2009, 124, 506–512. [Google Scholar] [CrossRef]
- Achari, C.; Reddy, G.V.; Reddy, T.C.M.; Reddanna, P. Chebulagic Acid Synergizes the Cytotoxicity of Doxorubicin in Human Hepatocellular Carcinoma Through COX-2 Dependant Modulation of MDR-1. Med. Chem. 2011, 7, 432–442. [Google Scholar] [CrossRef]
- Clifford, M.N.; Kerimi, A.; Williamson, G. Bioavailability and metabolism of chlorogenic acids (acyl-quinic acids) in humans. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1299–1352. [Google Scholar] [CrossRef] [PubMed]
- Najar, I.A.; Sachin, B.S.; Sharma, S.C.; Satti, N.K.; Suri, K.A.; Johri, R.K. Modulation of P-glycoprotein ATPase activity by some phytoconstituents. Phytother. Res. 2010, 24, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Fan, Y.; Su, H.; Wu, L.; Huang, Y.; Zhao, L.; Han, B.; Shu, G.; Xiang, M.; Yang, J.-M. Chlorogenic acid methyl ester exerts strong anti-inflammatory effects via inhibiting the COX-2/NLRP3/NF-κB pathway. Food Funct. 2018, 9, 6155–6164. [Google Scholar] [CrossRef] [PubMed]
- Pang, C.; Sheng, Y.-C.; Jiang, P.; Wei, H.; Ji, L.-L. Chlorogenic acid prevents acetaminophen-induced liver injury: The involvement of CYP450 metabolic enzymes and some antioxidant signals. J. Zhejiang Univ. Sci. B 2015, 16, 602–610. [Google Scholar] [CrossRef]
- Meng, S.; Cao, J.; Feng, Q.; Peng, J.; Hu, Y. Roles of chlorogenic acid on regulating glucose and lipids metabolism: A review. Evid. Based Complement. Altern. Med. 2013, 2013, 81457. [Google Scholar] [CrossRef]
- Mei, Y.; Pan, D.; Jiang, Y.; Zhang, W.; Yao, X.; Dai, Y.; Yu, Y.; Yao, X. Target discovery of chlorogenic acid derivatives from the flower buds of Lonicera macranthoides and their MAO B inhibitory mechanism. Fitoterapia 2019, 134, 297–304. [Google Scholar] [CrossRef]
- Miyazawa, M.; Shindo, M.; Shimada, T. Oxidation of 1,8-cineole, the monoterpene cyclic ether originated from eucalyptus polybractea, by cytochrome P450 3A enzymes in rat and human liver microsomes. Drug Metab. Dispos. 2001, 29, 200–205. [Google Scholar]
- Xi, J.; Yun, M.; Lee, D.; Park, M.-N.; Kim, E.-O.; Sohn, E.J.; Kwon, B.-M. Cinnamaldehyde Derivative (CB-PIC) Sensitizes Chemo-Resistant Cancer Cells to Drug-Induced Apoptosis via Suppression of MDR1 and its Upstream STAT3 and AKT Signalling. Cell. Physiol. Biochem. 2015, 35, 1821–1830. [Google Scholar] [CrossRef]
- Huang, B.; Yuan, H.D.; Kim, D.Y.; Quan, H.Y.; Chung, S.H. Cinnamaldehyde prevents adipocyte differentiation and adipogenesis via regulation of peroxisome proliferator-activated receptor-γ (PPARγ) and AMP-activated protein kinase (AMPK) pathways. J. Agric. Food Chem. 2011, 59, 3666–3673. [Google Scholar] [CrossRef]
- Guo, J.-Y.; Huo, H.-R.; Zhao, B.-S.; Liu, H.-B.; Li, L.-F.; Ma, Y.-Y.; Guo, S.-Y.; Jiang, T.-L. Cinnamaldehyde reduces IL-1beta-induced cyclooxygenase-2 activity in rat cerebral microvascular endothelial cells. Eur. J. Pharmacol. 2006, 537, 174–180. [Google Scholar] [CrossRef]
- Hasegawa, A.; Yoshino, M.; Nakamura, H.; Ishii, I.; Watanabe, T.; Kiuchi, M.; Ishikawa, T.; Ohmori, S.; Kitada, M. Identification of inhibitory component in cinnamon--o-methoxycinnamaldehyde inhibits CYP1A2 and CYP2E1-. Drug Metab. Pharmacokinet. 2002, 17, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.H.; Lawson, J.; Backwell, F.R. Inhibition and inactivation of monoamine oxidase by 3-amino-1-phenyl-prop-1-enes. Biochim. Biophys. Acta 1992, 1119, 111–117. [Google Scholar] [CrossRef]
- Chen, C.; Wu, C.; Lu, X.; Yan, Z.; Gao, J.; Zhao, H.; Li, S. Coniferyl ferulate, a strong inhibitor of glutathione s-transferase isolated from radix Angelicae sinensis, reverses multidrug resistance and downregulates P-glycoprotein. Evid. Based Complement. Altern. Med. 2013, 2013, 639093. [Google Scholar] [CrossRef]
- Ro, J.S.; Lee, S.S.; Lee, K.S.; Lee, M.K. Inhibition of type A monoamine oxidase by coptisine in mouse brain. Life Sci. 2001, 70, 639–645. [Google Scholar] [CrossRef]
- Li, L.; Sun, S.; Weng, Y.; Song, F.; Zhou, S.; Bai, M.; Zhou, H.; Zeng, S.; Jiang, H. Interaction of six protoberberine alkaloids with human organic cation transporters 1, 2 and 3. Xenobiotica 2016, 46, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-A.; Kang, S.-I.; Shin, H.-S.; Kang, S.-W.; Kim, J.-H.; Ko, H.-C.; Kim, S.-J. p-Coumaric acid modulates glucose and lipid metabolism via AMP-activated protein kinase in L6 skeletal muscle cells. Biochem. Biophys. Res. Commun. 2013, 432, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Luceri, C.; Guglielmi, F.; Lodovici, M.; Giannini, L.; Messerini, L.; Dolara, P. Plant phenolic 4-coumaric acid protects against intestinal inflammation in rats. Scand. J. Gastroenterol. 2004, 39, 1128–1133. [Google Scholar] [CrossRef]
- Yang, F.; Wang, Y.; Li, G.; Xue, J.; Chen, Z.-L.; Jin, F.; Luo, L.; Zhou, X.; Ma, Q.; Cai, X.; et al. Effects of corilagin on alleviating cholestasis via farnesoid X receptor-associated pathways in vitro and in vivo. Br. J. Pharmacol. 2018, 175, 810–829. [Google Scholar] [CrossRef]
- Lautenschläger, M.; Sendker, J.; Hüwel, S.; Galla, H.J.; Brandt, S.; Düfer, M.; Riehemann, K.; Hensel, A. Intestinal formation of trans-crocetin from saffron extract (Crocus sativus L.) and in vitro permeation through intestinal and blood brain barrier. Phytomedicine 2015, 22, 36–44. [Google Scholar] [CrossRef]
- Neyshaburinezhad, N.; Hashemi, M.; Ramezani, M.; Arabzadeh, S.; Behravan, J.; Kalalinia, F. The effects of crocetin, extracted from saffron, in chemotherapy against the incidence of multiple drug resistance phenotype. Iran. J. Basic Med. Sci. 2018, 21, 1192–1197. [Google Scholar] [CrossRef]
- Berger, F.; Hensel, A.; Nieber, K. Trans-crocetin is involved in the inhibition of the glutamatergic synaptic transmission on rat cortical neurones by saffron extract. Planta Med. 2010, 76, P589. [Google Scholar] [CrossRef]
- Kamyar, M.; Razavi, B.M.; Hasani, F.V.; Mehri, S.; Foroutanfar, A.; Hosseinzadeh, H. Crocin prevents haloperidol-induced orofacial dyskinesia: Possible an antioxidant mechanism. Iran. J. Basic Med. Sci. 2016, 19, 1070–1079. [Google Scholar] [PubMed]
- Dovrtělová, G.; Nosková, K.; Juřica, J.; Turjap, M.; Zendulka, O. Can bioactive compounds of Crocus sativus L. Influence the metabolic activity of selected CYP enzymes in the rat? Physiol. Res. 2015, 64, S453–S458. [Google Scholar] [CrossRef] [PubMed]
- Razavi, S.M.S.; Vaziri, R.M.; Karimi, G.; Arabzadeh, S.; Keyvani, V.; Behravan, J.; Kalalinia, F. Crocin Increases Gastric Cancer Cells’ Sensitivity to Doxorubicin. Asian Pac. J. Cancer Prev. 2020, 21, 1959–1967. [Google Scholar] [CrossRef]
- Lopes-Rodrigues, V.; Sousa, E.; Vasconcelos, M.H. Curcumin as a Modulator of P-Glycoprotein in Cancer: Challenges and Perspectives. Pharmaceuticals 2016, 9, 71. [Google Scholar] [CrossRef]
- Wortelboer, H.M.; Usta, M.; van der Velde, A.E.; Boersma, M.G.; Spenkelink, B.; van Zanden, J.J.; Rietjens, I.M.C.M.; van Bladeren, P.J.; Cnubben, N.H.P. Interplay between MRP inhibition and metabolism of MRP inhibitors: The case of curcumin. Chem. Res. Toxicol. 2003, 16, 1642–1651. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, F.; Chen, C.; Guo, Z.; Liu, J.; Yu, J.; Xu, Y.; Zhong, D.; Jiang, H. Impact of curcumin on the pharmacokinetics of rosuvastatin in rats and dogs based on the conjugated metabolites. Xenobiotica 2017, 47, 267–275. [Google Scholar] [CrossRef]
- Li, H.; Krstin, S.; Wink, M. Modulation of multidrug resistant in cancer cells by EGCG, tannic acid and curcumin. Phytomedicine 2018, 50, 213–222. [Google Scholar] [CrossRef]
- Naganuma, M.; Saruwatari, A.; Okamura, S.; Tamura, H. Turmeric and curcumin modulate the conjugation of 1-naphthol in Caco-2 cells. Biol. Pharm. Bull. 2006, 29, 1476–1479. [Google Scholar] [CrossRef]
- Basu, N.K.; Kole, L.; Kubota, S.; Owens, I.S. Human UDP-glucuronosyltransferases show atypical metabolism of mycophenolic acid and inhibition by curcumin. Drug Metab. Dispos. 2004, 32, 768–773. [Google Scholar] [CrossRef]
- Volak, L.P.; Court, M.H. Role for protein kinase C delta in the functional activity of human UGT1A6: Implications for drug-drug interactions between PKC inhibitors and UGT1A6. Xenobiotica 2010, 40, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Ricci, J.W.; Lovato, D.; Larson, R.S. ABCG2 Inhibitors: Will They Find Clinical Relevance? J. Develop. Drugs 2015, 4, 138. [Google Scholar] [CrossRef]
- Gameiro, M.; Silva, R.; Rocha-Pereira, C.; Carmo, H.; Carvalho, F.; Bastos, M.D.L.; Remião, F. Cellular Models and In Vitro Assays for the Screening of modulators of P-gp, MRP1 and BCRP. Molecules 2017, 22, 600. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, J.; Guo, C.; Xing, H.; Xu, J.; Wen, Y.; Qiu, Z.; Zhang, Q.; Zheng, Y.; Chen, X.; et al. Pharmacokinetic effects of curcumin on docetaxel mediated by OATP1B1, OATP1B3 and CYP450s. Drug Metab. Pharmacokinet. 2016, 31, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Zapletalova, M.K.; Lecianova, A.; Spicakova, A.; Anzenbacherova, E.; Anzenbacher, P. Interaction of anthocyanins with human liver microsomal cytochromes P450. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech. Repub. 2013, 157, S56–S59. [Google Scholar]
- Dreiseitel, A.; Oosterhuis, B.; Vukman, K.V.; Schreier, P.; Oehme, A.; Locher, S.; Hajak, G.; Sand, P.G. Berry anthocyanins and anthocyanidins exhibit distinct affinities for the efflux transporters BCRP and MDR1. Br. J. Pharmacol. 2009, 158, 1942–1950. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, A.H.; Rønsted, N.; Güler, S.; Jäger, A.K.; Sendra, J.R.; Brodin, B. In-vitro evaluation of the P-glycoprotein interactions of a series of potentially CNS-active Amaryllidaceae alkaloids. J. Pharm. Pharmacol. 2012, 64, 1667–1677. [Google Scholar] [CrossRef]
- Wang, Q.; Xia, M.; Liu, C.; Guo, H.; Ye, Q.; Hu, Y.; Zhang, Y.; Hou, M.; Zhu, H.; Ma, J.; et al. Cyanidin-3-O-beta-glucoside inhibits iNOS and COX-2 expression by inducing liver X receptor alpha activation in THP-1 macrophages. Life Sci. 2008, 83, 176–184. [Google Scholar] [CrossRef]
- Srovnalova, A.; Svecarova, M.; Zapletalova, M.K.; Anzenbacher, P.; Bachleda, P.; Anzenbacherova, E.; Dvorak, Z. Effects of anthocyanidins and anthocyanins on the expression and catalytic activities of CYP2A6, CYP2B6, CYP2C9, and CYP3A4 in primary human hepatocytes and human liver microsomes. J. Agric. Food Chem. 2014, 62, 789–797. [Google Scholar] [CrossRef]
- Fuchikami, H.; Satoh, H.; Tsujimoto, M.; Ohdo, S.; Ohtani, H.; Sawada, Y. Effects of herbal extracts on the function of human organic anion-transporting polypeptide OATP-B. Drug Metab. Dispos. 2006, 34, 577–582. [Google Scholar] [CrossRef]
- Atherton, K.M.; Mutch, E.; Ford, D. Metabolism of the soyabean isoflavone daidzein by CYP1A2 and the extra-hepatic CYPs 1A1 and 1B1 affects biological activity. Biochem. Pharmacol. 2006, 72, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.X.; Wang, L.S.; Li, H.D.; Abd El-Aty, A.M.; Chen, G.L.; Zhou, H.H. Evidence for the involvement of human liver microsomes CYP1A2 in the mono-hydroxylation of daidzein. Clin. Chim. Acta 2003, 334, 77–85. [Google Scholar] [CrossRef]
- Okura, T.; Ibe, M.; Umegaki, K.; Shinozuka, K.; Yamada, S. Effects of dietary ingredients on function and expression of P-glycoprotein in human intestinal epithelial cells. Biol. Pharm. Bull. 2010, 33, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Pritchett, L.E.; Atherton, K.M.; Mutch, E.; Ford, D. Glucuronidation of the soyabean isoflavones genistein and daidzein by human liver is related to levels of UGT1A1 and UGT1A9 activity and alters isoflavone response in the MCF-7 human breast cancer cell line. J. Nutr. Biochem. 2008, 19, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Aty, A.M.; Shah, S.S.; Kim, B.M.; Choi, J.H.; Cho, H.J.; Chang, B.J.; Shin, H.C.; Lee, K.B.; Shimoda, M.; Shime, J.-H. In Vitro inhibitory potential of decursin and decursinol angelate on the catalytic activity of cytochrome P-450 1A1/2, 2D15, and 3A12 isoforms in canine hepatic microsomes. Arch. Pharm. Res. 2008, 31, 1425–1435. [Google Scholar] [CrossRef]
- Abdallah, H.M.; Al-Abd, A.M.; El-Dine, R.S.; El-Halawany, A.M. P-glycoprotein inhibitors of natural origin as potential tumor chemo-sensitizers: A review. J. Adv. Res. 2015, 6, 45–62. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, L.; Tang, S.; Hale, T.W.; Xing, C.; Jiang, C.; Lü, J. Cytochrome P450 Isoforms in the Metabolism of Decursin and Decursinol Angelate from Korean Angelica. Am. J. Chin. Med. 2015, 43, 1211–1230. [Google Scholar] [CrossRef]
- Riha, J.; Brenner, S.; Srovnalova, A.; Klameth, L.; Dvorak, Z.; Jäger, W.; Thalhammer, T. Effects of anthocyanins on the expression of organic anion transporting polypeptides (SLCOs/OATPs) in primary human hepatocytes. Food Funct. 2015, 6, 772–779. [Google Scholar] [CrossRef]
- Liagre, B.; Vergne-Salle, P.; Corbiere, C.; Charissoux, J.L.; Beneytout, J.L. Diosgenin, a plant steroid, induces apoptosis in human rheumatoid arthritis synoviocytes with cyclooxygenase-2 overexpression. Arthritis Res. Ther. 2004, 6, R373–R383. [Google Scholar] [CrossRef]
- Manda, V.K.; Avula, B.; Wong, Y.H.; Smillie, T.J.; Kahn, I.A.; Khan, S.I. Characterization of in Vitro ADME Properties of Diosgenin and Dioscin from Dioscorea villosa. Planta Med. 2013, 79, 1421–1428. [Google Scholar] [CrossRef]
- Vijayakumar, T.M.; Ilango, K.; Kumar, R.M.; Agrawal, A.; Dubey, G.P. Effect of Dioscorea bulbifera and its Major Bioactive Compound, Diosgenin on CYP450 Mediated Drug Metabolism. JBAPN 2015, 5, 313–321. [Google Scholar] [CrossRef]
- González-Sarrías, A.; Azorín-Ortuño, M.; Yáñez-Gascón, M.J.; Tomás-Barberán, F.A.; García-Conesa, M.T.; Espín, J.C. Dissimilar in vitro and in vivo effects of ellagic acid and its microbiota-derived metabolites, urolithins, on the cytochrome P450 1A1. J. Agric. Food Chem. 2009, 57, 5623–5632. [Google Scholar] [CrossRef] [PubMed]
- Barch, D.H.; Rundhaugen, L.M.; Thomas, P.E.; Kardos, P.; Pillay, N.S. Dietary ellagic acid inhibits the enzymatic activity of CYP1A1 without altering hepatic concentrations of CYP1A1 or CYP1A1 mRNA. Biochem. Biophys. Res. Commun. 1994, 201, 1477–1482. [Google Scholar] [CrossRef] [PubMed]
- Celik, G.; Semiz, A.; Karakurt, S.; Arslan, S.; Adali, O.; Sen, A. A comparative study for the evaluation of two doses of ellagic acid on hepatic drug metabolizing and antioxidant enzymes in the rat. Biomed Res. Int. 2013, 2013, 358945. [Google Scholar] [CrossRef]
- Whitley, A.C.; Sweet, D.H.; Walle, T. The dietary polyphenol ellagic acid is a potent inhibitor of hOAT1. Drug Metab. Dispos. 2005, 33, 1097–1100. [Google Scholar] [CrossRef]
- Van Wert, A.L.; Gionfriddo, M.R.; Sweet, D.H. Organic anion transporters: Discovery, pharmacology, regulation and roles in pathophysiology. Biopharm. Drug Dispos. 2010, 31, 1–71. [Google Scholar] [CrossRef]
- Ahn, D.; Putt, D.; Kresty, L.; Stoner, G.D.; Fromm, D.; Hollenberg, P.F. The effects of dietary ellagic acid on rat hepatic and esophageal mucosal cytochromes P450 and phase II enzymes. Carcinogenesis 1996, 17, 821–828. [Google Scholar] [CrossRef]
- Tamaki, N.; Matsunami, K.; Otsuka, H.; Shinzato, T.; Aramoto, M.; Takeda, Y. Rearranged ent-kauranes from the stems of Tricalysia dubia and their biological activities. J. Nat. Med. 2008, 62, 314–320. [Google Scholar] [CrossRef]
- Sul, Y.H.; Lee, M.S.; Cha, E.Y.; Thuong, P.T.; Khoi, N.M.; Song, I.S. An ent-kaurane diterpenoid from Croton tonkinensis induces apoptosis by regulating AMP-activated protein kinase in SK-HEP1 human hepatocellular carcinoma cells. Biol. Pharm. Bull. 2013, 36, 158–164. [Google Scholar] [CrossRef]
- Lee, M.; Kim, S.H.; Lee, H.K.; Cho, Y.; Kang, J.; Sung, S.H. Ent-kaurane and ent-pimarane diterpenes from Siegesbeckia pubescens inhibit lipopolysaccharide-induced nitric oxide production in BV2 microglia. Biol. Pharm. Bull. 2014, 37, 152–157. [Google Scholar] [CrossRef]
- Chiba, T.; Sato, Y.; Suzuki, S.; Umegaki, K. Concomitant use of dietary supplements and medicines in patients due to miscommunication with physicians in Japan. Nutrients 2015, 7, 2947–2960. [Google Scholar] [CrossRef] [PubMed]
- Roth, M.; Timmermann, B.N.; Hagenbuch, B. Interactions of green tea catechins with organic anion-transporting polypeptides. Drug Metab. Dispos. 2011, 39, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Jodoin, J.; Demeule, M.; Beliveau, R. Inhibition of the multidrug resistance P-glycoprotein activity by green tea polyphenols. Biochim. Biophys. Acta 2002, 1542, 149–159. [Google Scholar] [CrossRef]
- Greenmol BD | Nature Derived Molecule Database. Available online: https://www.greenmolbd.gov.bd/search?search_in=target&term=Solute%20carrier%20organic%20anion%20transporter%20family%20member%201B1%20(OATP1B1 (accessed on 28 November 2020).
- Wen, Y.; Zhao, R.; Gupta, P.; Fan, Y.; Zhang, Y.; Huang, Z.; Li, X.; Su, Y.; Liao, L.; Xie, Y.-A.; et al. The epigallocatechin gallate derivative Y6 reverses drug resistance mediated by the ABCB1 transporter both in vitro and in vivo. Acta Pharm. Sin. B 2019, 9, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Muto, S.; Fujita, K.; Yamazaki, Y.; Kamataki, T. Inhibition by green tea catechins of metabolic activation of procarcinogens by human cytochrome P450. Mutat. Res. 2001, 479, 197–206. [Google Scholar] [CrossRef]
- Isaka, S.M.; Awabe, K.K.; Noue, S.O.; Erba, J.P.W.; Iroli, M.G.; Amaki, S.T.; An, T.K.; Imura, J.K.; Atanabe, H.W.; Amada, S.Y. Effects of Green Tea Catechins on Cytochrome P450 2B6, 2C8, 2C19, 2D6 and 3A Activities in Human Liver and Intestinal Microsomes. Drug Metab. Pharmacokinet. 2013, 28, 244–249. [Google Scholar] [CrossRef]
- Kitagawa, S.; Nabekura, T.; Kamiyama, S. Inhibition of P-glycoprotein function by tea catechins in KB-C2 cells. J. Pharm. Pharmacol. 2004, 56, 1001–1005. [Google Scholar] [CrossRef]
- Alvarez, A.I.; Real, R.; Pérez, M.; Mendoza, G.; Prieto, J.G.; Merino, G. Modulation of the activity of ABC transporters (P-glycoprotein, MRP2, BCRP) by flavonoids and drug response. J. Pharm. Sci. 2010, 99, 598–617. [Google Scholar] [CrossRef]
- Wang, E.J.; Barecki-Roach, M.; Johnson, W.W. Elevation of P-glycoprotein function by a catechin in green tea. Biochem. Biophys. Res. Commun. 2002, 297, 412–418. [Google Scholar] [CrossRef]
- Farabegoli, F.; Papi, A.; Bartolini, G.; Ostan, R.; Orlandi, M. (-)-Epigallocatechin-3-gallate downregulates Pg-P and BCRP in a tamoxifen resistant MCF-7 cell line. Phytomedicine 2010, 17, 356–362. [Google Scholar] [CrossRef]
- Iwano, H.; Ujita, W.; Nishikawa, M.; Ishii, S.; Inoue, H.; Yokota, H. Effect of dietary eugenol on xenobiotic metabolism and mediation of UDP-glucuronosyltransferase and cytochrome P450 1A1 expression in rat liver. Int. J. Food Sci. Nutr. 2014, 65, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Jeong, K.J.; Kim, D.Y.; Quan, H.-Y.; Jo, H.K.; Kim, G.W.; Chung, S.H. Effects of eugenol on hepatic glucose production and AMPK signaling pathway in hepatocytes and C57BL/6J mice. Fitoterapia 2014, 93, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Oh, O.-J.; Min, H.-Y.; Park, E.-J.; Kim, Y.; Park, H.J.; Nam Han, Y.; Lee, S.K. Eugenol suppresses cyclooxygenase-2 expression in lipopolysaccharide-stimulated mouse macrophage RAW264.7 cells. Life Sci. 2003, 73, 337–348. [Google Scholar] [CrossRef]
- Gardner, I.; Wakazono, H.; Bergin, P.; de Waziers, I.; Beaune, P.; Kenna, J.G.; Caldwell, J. Cytochrome P450 mediated bioactivation of methyleugenol to 1’-hydroxymethyleugenol in Fischer 344 rat and human liver microsomes. Carcinogenesis 1997, 18, 1775–1783. [Google Scholar] [CrossRef]
- Harb, A.A.; Bustanji, Y.K.; Almasri, I.M.; Abdalla, S.S. Eugenol Reduces LDL Cholesterol and Hepatic Steatosis in Hypercholesterolemic Rats by Modulating TRPV1 Receptor. Sci. Rep. 2019, 9, 14003. [Google Scholar] [CrossRef]
- Wie, M.B.; Won, M.H.; Lee, K.H.; Shin, J.H.; Lee, J.C.; Suh, H.W.; Song, D.K.; Kim, Y.H. Eugenol protects neuronal cells from excitotoxic and oxidative injury in primary cortical cultures. Neurosci. Lett. 1997, 225, 93–96. [Google Scholar] [CrossRef]
- Nabekura, T.; Kawasaki, T.; Furuta, M.; Kaneko, T.; Uwai, Y. Effects of Natural Polyphenols on the Expression of Drug Efflux Transporter P-Glycoprotein in Human Intestinal Cells. ACS Omega 2018, 3, 1621–1626. [Google Scholar] [CrossRef]
- Yu, L.; Zhang, Y.; Ma, R.; Bao, L.; Fang, J.; Yu, T. Potent protection of ferulic acid against excitotoxic effects of maternal intragastric administration of monosodium glutamate at a late stage of pregnancy on developing mouse fetal brain. Eur. Neuropsychopharmacol. 2006, 16, 170–177. [Google Scholar] [CrossRef]
- Muthusamy, G.; Balupillai, A.; Ramasamy, K.; Shanmugam, M.; Gunaseelan, S.; Mary, B.; Prasad, N.R. Ferulic acid reverses ABCB1-mediated paclitaxel resistance in MDR cell lines. Eur. J. Pharmacol. 2016, 786, 194–203. [Google Scholar] [CrossRef]
- Zhuang, X.-M.; Chen, L.; Tan, Y.; Yang, H.-Y.; Lu, C.; Gao, Y.; Li, H. Identification of human cytochrome P450 and UGT enzymes involved in the metabolism of ferulic acid, a major bioactive component in traditional Chinese medicines. Chin. J. Nat. Med. 2017, 15, 695–702. [Google Scholar] [CrossRef]
- Mathew, B.; Baek, S.C.; Parambi, D.G.T.; Lee, J.P.; Mathew, G.E.; Jayanthi, S.; Vinod, D.; Rapheal, C.; Devikrishna, V.; Kondarath, S.S.; et al. Potent and highly selective dual-targeting monoamine oxidase-B inhibitors: Fluorinated chalcones of morpholine versus imidazole. Archiv der Pharmazie 2019, 352, 1800309. [Google Scholar] [CrossRef]
- Wang, E.J.; Casciano, C.N.; Clement, R.P.; Johnson, W.W. Active transport of fluorescent P-glycoprotein substrates: Evaluation as markers and interaction with inhibitors. Biochem. Biophys. Res. Commun. 2001, 289, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Virgona, N.; Yokotani, K.; Yamazaki, Y.; Shimura, F.; Chiba, T.; Taki, Y.; Yamada, S.; Shinozuka, K.; Murata, M.; Umegaki, K. Coleus forskohlii extract induces hepatic cytochrome P450 enzymes in mice. Food Chem. Toxicol. 2012, 50, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Yokotani, K.; Chiba, T.; Sato, Y.; Taki, Y.; Yamada, S.; Shinozuka, K.; Murata, M.; Umegaki, K. Hepatic cytochrome P450 mediates interaction between warfarin and Coleus forskohlii extract in vivo and in vitro. J. Pharm. Pharmacol. 2012, 64, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Hurley, R.L.; Barré, L.K.; Wood, S.D.; Anderson, K.A.; Kemp, B.E.; Means, A.R.; Witters, L.A. Regulation of AMP-activated protein kinase by multisite phosphorylation in response to agents that elevate cellular cAMP. J. Biol. Chem. 2006, 281, 36662–36672. [Google Scholar] [CrossRef] [PubMed]
- Egawa, M.; Kamata, H.; Kushiyama, A.; Sakoda, H.; Fujishiro, M.; Horike, N.; Yoneda, M.; Nakatsu, Y.; Ying, G.; Jun, Z.; et al. Long-term forskolin stimulation induces AMPK activation and thereby enhances tight junction formation in human placental trophoblast BeWo cells. Placenta 2008, 29, 1003–1008. [Google Scholar] [CrossRef]
- El-Agroudy, N.N.; El-Naga, R.N.; El-Razeq, R.A.; El-Demerdash, E. Forskolin, a hedgehog signalling inhibitor, attenuates carbon tetrachloride-induced liver fibrosis in rats. Br. J. Pharmacol. 2016, 173, 3248–3260. [Google Scholar] [CrossRef]
- Ríos-Silva, M.; Trujillo, X.; Trujillo-Hernández, B.; Sánchez-Pastor, E.; Urzúa, Z.; Mancilla, E.; Huerta, M. Effect of chronic administration of forskolin on glycemia and oxidative stress in rats with and without experimental diabetes. Int. J. Med. Sci. 2014, 11, 448–452. [Google Scholar] [CrossRef]
- Kihira, Y.; Burentogtokh, A.; Itoh, M.; Izawa-Ishizawa, Y.; Ishizawa, K.; Ikeda, Y.; Tsuchiya, K.; Tamaki, T. Hypoxia decreases glucagon-like peptide-1 secretion from the GLUTag cell line. Biol. Pharm. Bull. 2015, 38, 514–521. [Google Scholar] [CrossRef]
- Grey, K.B.; Burrell, B.D. Forskolin induces NMDA receptor-dependent potentiation at a central synapse in the leech. J. Neurophysiol. 2008, 99, 2719–2724. [Google Scholar] [CrossRef]
- Morris, D.I.; Speicher, L.A.; Ruoho, A.E.; Tew, K.D.; Seamon, K.B. Interaction of forskolin with the P-glycoprotein multidrug transporter. Biochemistry 1991, 30, 8371–8379. [Google Scholar] [CrossRef] [PubMed]
- Namanja, H.A.; Emmert, D.; Pires, M.M.; Hrycyna, C.A.; Chmielewski, J. Inhibition of human P-glycoprotein transport and substrate binding using a galantamine dimer. Biochem. Biophys. Res. Commun. 2009, 388, 672–676. [Google Scholar] [CrossRef] [PubMed]
- McNulty, J.; Nair, J.J.; Singh, M.; Crankshaw, D.J.; Holloway, A.C.; Bastida, J. Selective cytochrome P450 3A4 inhibitory activity of Amaryllidaceae alkaloids. Bioorg. Med. Chem. Lett. 2009, 19, 3233–3237. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Fu, Y. A review of clinical pharmacokinetics and pharmacodynamics of galantamine, a reversible acetylcholinesterase inhibitor for the treatment of Alzheimer’s disease, in healthy subjects and patients. Curr. Clin. Pharmacol. 2010, 5, 115–124. [Google Scholar] [CrossRef]
- Jann, M.W.; Shirley, K.L.; Small, G.W. Clinical pharmacokinetics and pharmacodynamics of cholinesterase inhibitors. Clin. Pharmacokinet. 2002, 41, 719–739. [Google Scholar] [CrossRef]
- Cacabelos, R. Pharmacogenetic considerations when prescribing cholinesterase inhibitors for the treatment of Alzheimer’s disease. Expert Opin. Drug Metab. Toxicol. 2020, 16, 673–701. [Google Scholar] [CrossRef]
- Amaravani, M.; Prasad, N.K.; Ramakrishna, V. COX-2 structural analysis and docking studies with gallic acid structural analogues. Springerplus 2012, 1, 58. [Google Scholar] [CrossRef]
- Ponnusankar, S.; Pandit, S.; Babu, R.; Bandyopadhyay, A.; Mukherjee, P.K. Cytochrome P450 inhibitory potential of Triphala—A Rasayana from Ayurveda. J. Ethnopharmacol. 2011, 133, 120–125. [Google Scholar] [CrossRef]
- Athukuri, B.L.; Neerati, P. Enhanced oral bioavailability of metoprolol with gallic acid and ellagic acid in male Wistar rats: Involvement of CYP2D6 inhibition. Drug Metab. Pers. Ther. 2016, 31, 229–234. [Google Scholar] [CrossRef]
- Chhillar, R.; Dhingra, D. Antidepressant-like activity of gallic acid in mice subjected to unpredictable chronic mild stress. Fundam. Clin. Pharmacol. 2013, 27, 409–418. [Google Scholar] [CrossRef]
- Kitagawa, S.; Nabekura, T.; Kamiyama, S.; Takahashi, T.; Nakamura, Y.; Kashiwada, Y.; Ikeshiro, Y. Effects of alkyl gallates on P-glycoprotein function. Biochem. Pharmacol. 2005, 70, 1262–1266. [Google Scholar] [CrossRef] [PubMed]
- Kahkeshani, N.; Farzaei, F.; Fotouhi, M.; Alavi, S.S.; Bahramsoltani, R.; Naseri, R.; Momtaz, S.; Abbasabadi, Z.; Rahimi, R.; Farzaei, M.H.; et al. Pharmacological effects of gallic acid in health and diseases: A mechanistic review. Iran. J. Basic Med. Sci. 2019, 22, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, K.; Zheng, J.; Cheung, F.S.G.; Chan, T.; Zhu, L.; Zhou, F. Interactions of the active components of Punica granatum (pomegranate) with the essential renal and hepatic human Solute Carrier transporters. Pharm. Biol. 2014, 52, 1510–1517. [Google Scholar] [CrossRef] [PubMed]
- Palacios-González, B.; Zarain-Herzberg, A.; Flores-Galicia, I.; Noriega, L.G.; Alemán-Escondrillas, G.; Zariñan, T.; Ulloa-Aguirre, A.; Torres, N.; Tovar, A.R. Genistein stimulates fatty acid oxidation in a leptin receptor-independent manner through the JAK2-mediated phosphorylation and activation of AMPK in skeletal muscle. Biochim. Biophys. Acta 2014, 1841, 132–140. [Google Scholar] [CrossRef]
- Tuli, H.S.; Tuorkey, M.J.; Thakral, F.; Sak, K.; Kumar, M.; Sharma, A.K.; Sharma, U.; Jain, A.; Aggarwal, V.; Bishayee, A. Molecular Mechanisms of Action of Genistein in Cancer: Recent Advances. Front. Pharmacol. 2019, 10, 1336. [Google Scholar] [CrossRef]
- Lepri, S.R.; Sartori, D.; Semprebon, S.C.; Baranoski, A.; Coatti, G.C.; Mantovani, M.S. Genistein Affects Expression of Cytochrome P450 (CYP450) Genes in Hepatocellular Carcinoma (HEPG2/C3A) Cell Line. Drug Metab. Lett. 2018, 12, 138–144. [Google Scholar] [CrossRef]
- Notarnicola, M.; Messa, C.; Orlando, A.; D’Attoma, B.; Tutino, V.; Rivizzigno, R.; Caruso, M.G. Effect of genistein on cholesterol metabolism-related genes in a colon cancer cell line. Genes Nutr. 2008, 3, 35–40. [Google Scholar] [CrossRef]
- Zarmouh, N.O.; Messeha, S.S.; Elshami, F.M.; Soliman, K.F.A. Evaluation of the Isoflavone Genistein as Reversible Human Monoamine Oxidase-A and -B Inhibitor. Evid. Based Complement. Altern. Med. 2016, 2016, 1423052. [Google Scholar] [CrossRef]
- Huang, R.; Singh, M.; Dillon, G.H. Genistein directly inhibits native and recombinant NMDA receptors. Neuropharmacology 2010, 58, 1246–1251. [Google Scholar] [CrossRef]
- Cho, H.-J.; Yoon, I.-S. Pharmacokinetic interactions of herbs with cytochrome p450 and p-glycoprotein. Evid. Based Complement. Altern. Med. 2015, 2015, 736431. [Google Scholar] [CrossRef]
- Qiu, J.X.; Zhou, Z.W.; He, Z.X.; Zhang, X.; Zhou, S.F.; Zhu, S. Estimation of the binding modes with important human cytochrome P450 enzymes, drug interaction potential, pharmacokinetics, and hepatotoxicity of ginger components using molecular docking, computational, and pharmacokinetic modeling studies. Drug Des. Dev. Ther. 2015, 9, 841–866. [Google Scholar] [CrossRef] [PubMed]
- Bogacz, A.; Deka-Pawlik, D.; Bartkowiak-Wieczorek, J.; Karasiewicz, M.; Kujawski, R.; Kowalska, A.; Chałas, A.; Czerny, B.; Grześkowiak, E.; Mrozikiewicz, P.M. The effect of herbal materials on the p-glycoprotein activity and function. Herb. Pol. 2013, 59, 129–141. [Google Scholar] [CrossRef][Green Version]
- Li, Y.; Wu, Y.; Yao, X.; Hao, F.; Yu, C.; Bao, Y.; Wu, Y.; Song, Z.; Sun, Y.; Zheng, L.; et al. Ginkgolide a ameliorates LPS-induced inflammatory responses in vitro and in vivo. Int. J. Mol. Sci. 2017, 18, 794. [Google Scholar] [CrossRef] [PubMed]
- Kuo, L.-C.; Song, Y.-Q.; Yao, C.-A.; Cheng, I.H.; Chien, C.-T.; Lee, G.-C.; Yang, W.-C.; Lin, Y. Ginkgolide A Prevents the Amyloid-β-Induced Depolarization of Cortical Neurons. J. Agric. Food Chem. 2019, 67, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, M.R.; Penzak, S.R. Pharmacokinetic Drug Interactions with Panax ginseng. Eur. J. Drug Metab. Pharmacokinet. 2017, 42, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Liu, J.; Hou, J.; Zhan, T.; Yuan, L.; Liu, F.; Xiong, Y.; Hu, J.; Xia, C. Interactions of the major effective components in Shengmai formula with breast cancer resistance protein at the cellular and vesicular levels. Biomed. Pharmacother. 2020, 133, 110939. [Google Scholar] [CrossRef]
- Jiang, R.; Dong, J.; Li, X.; Du, F.; Jia, W.; Xu, F.; Wang, F.; Yang, J.; Niu, W.; Li, C. Molecular mechanisms governing different pharmacokinetics of ginsenosides and potential for ginsenoside-perpetrated herb-drug interactions on OATP1B3. Br. J. Pharmacol. 2015, 172, 1059–1073. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L.; Kim, S.-H.; Hagerman, A.E.; Lü, J. Anti-cancer, anti-diabetic and other pharmacologic and biological activities of penta-galloyl-glucose. Pharm. Res. 2009, 26, 2066–2080. [Google Scholar] [CrossRef]
- Lee, S.-J.; Lee, I.-S.; Mar, W. Inhibition of inducible nitric oxide synthase and cyclooxygenase-2 activity by 1,2,3,4,6-penta-O-galloyl-beta-d-glucose in murine macrophage cells. Arch. Pharm. Res. 2003, 26, 832–839. [Google Scholar] [CrossRef]
- Wen, F.; Shi, M.; Bian, J.; Zhang, H.; Gui, C. Identification of natural products as modulators of OATP2B1 using LC-MS/MS to quantify OATP-mediated uptake. Pharm. Biol. 2016, 54, 293–302. [Google Scholar] [CrossRef]
- Tai, T.; Huang, X.; Su, Y.; Ji, J.; Su, Y.; Jiang, Z.; Zhang, L. Glycyrrhizin accelerates the metabolism of triptolide through induction of CYP3A in rats. J. Ethnopharmacol. 2014, 152, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.-C.; Lin, S.-P.; Chao, P.-D.L. Liquorice reduced cyclosporine bioavailability by activating P-glycoprotein and CYP 3A. Food Chem. 2012, 135, 2307–2312. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, L.; Zhang, H.; Huang, S.; Xiong, Y.; Xia, C. Interaction of Sulfonylureas with Liver Uptake Transporters OATP1B1 and OATP1B3. Basic Clin. Pharmacol. Toxicol. 2018, 123, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Ishola, A.A.; Adewole, K.E. In Silico screening of anticholinesterase alkaloids for cyclooxygenase-2 (COX-2) and matrix metalloproteinase 8 (MMP-8) inhibitory potentials as multi-target inhibitors of Alzheimer’s disease. Med. Chem. Res. 2019, 28, 1704–1717. [Google Scholar] [CrossRef]
- Modarai, M.; Suter, A.; Kortenkamp, A.; Heinrich, M. The interaction potential of herbal medicinal products: A luminescence-based screening platform assessing effects on cytochrome P450 and its use with devil’s claw (Harpagophyti radix) preparations: Screening for CYP P450-HMP interactions. J. Pharm. Pharmacol. 2011, 63, 429–438. [Google Scholar] [CrossRef]
- Anauate, M.C.; Torres, L.M.; de Mello, S.B.V. Effect of isolated fractions of Harpagophytum procumbens D.C. (devil’s claw) on COX-1, COX-2 activity and nitric oxide production on whole-blood assay. Phytother. Res. 2010, 24, 1365–1369. [Google Scholar] [CrossRef]
- Zhang, L.; Feng, L.; Jia, Q.; Xu, J.; Wang, R.; Wang, Z.; Wu, Y.; Li, Y. Effects of β-glucosidase hydrolyzed products of harpagide and harpagoside on cyclooxygenase-2 (COX-2) in vitro. Bioorg. Med. Chem. 2011, 19, 4882–4886. [Google Scholar] [CrossRef]
- Unger, M.; Frank, A. Simultaneous determination of the inhibitory potency of herbal extracts on the activity of six major cytochrome P450 enzymes using liquid chromatography/mass spectrometry and automated online extraction. Rapid Commun. Mass Spectrom. 2004, 18, 2273–2281. [Google Scholar] [CrossRef]
- Fei, Z.; Hu, M.; Baum, L.; Kwan, P.; Hong, T.; Zhang, C. The potential role of human multidrug resistance protein 1 (MDR1) and multidrug resistance-associated protein 2 (MRP2) in the transport of Huperzine A in vitro. Xenobiotica 2020, 50, 354–362. [Google Scholar] [CrossRef]
- Ferreira, A.; Rodrigues, M.; Fortuna, A.; Falcão, A.; Alves, G. Huperzine A from Huperzia serrata: A review of its sources, chemistry, pharmacology and toxicology. Phytochem. Rev. 2016, 15, 51–85. [Google Scholar] [CrossRef]
- Ma, X.; Wang, H.; Xin, J.; Zhang, T.; Tu, Z. Identification of cytochrome P450 1A2 as enzyme involved in the microsomal metabolism of Huperzine A. Eur. J. Pharmacol. 2003, 461, 89–92. [Google Scholar] [CrossRef]
- Madabushi, R.; Frank, B.; Drewelow, B.; Derendorf, H.; Butterweck, V. Hyperforin in St. John’s wort drug interactions. Eur. J. Clin. Pharmacol. 2006, 62, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ji, Z.L.; Chen, Y.Z. TTD: Therapeutic Target Database. Nucleic Acids Res. 2002, 30, 412–415. [Google Scholar] [CrossRef] [PubMed]
- Koeberle, A.; Rossi, A.; Bauer, J.; Dehm, F.; Verotta, L.; Northoff, H.; Sautebin, L.; Werz, O. Hyperforin, an Anti-Inflammatory Constituent from St. John’s Wort, Inhibits Microsomal Prostaglandin E(2) Synthase-1 and Suppresses Prostaglandin E(2) Formation in vivo. Front. Pharmacol. 2011, 2, 7. [Google Scholar] [CrossRef] [PubMed]
- Quiney, C.; Billard, C.; Salanoubat, C.; Fourneron, J.D.; Kolb, J.P. Hyperforin, a new lead compound against the progression of cancer and leukemia? Leukemia 2006, 20, 1519–1525. [Google Scholar] [CrossRef]
- Albert, D.; Zündorf, I.; Dingermann, T.; Müller, W.E.; Steinhilber, D.; Werz, O. Hyperforin is a dual inhibitor of cyclooxygenase-1 and 5-lipoxygenase. Biochem. Pharmacol. 2002, 64, 1767–1775. [Google Scholar] [CrossRef]
- Schäfer, A.M.; Potterat, O.; Seibert, I.; Fertig, O.; Meyer Zu Schwabedissen, H.E. Hyperforin-Induced Activation of the Pregnane X Receptor Is Influenced by the Organic Anion-Transporting Polypeptide 2B1. Mol. Pharmacol. 2019, 95, 313–323. [Google Scholar] [CrossRef]
- Nicolussi, S.; Drewe, J.; Butterweck, V.; Meyer Zu Schwabedissen, H.E. Clinical relevance of St. John’s wort drug interactions revisited. Br. J. Pharmacol. 2020, 177, 1212–1226. [Google Scholar] [CrossRef]
- Kumar, V.; Mdzinarishvili, A.; Kiewert, C.; Abbruscato, T.; Bickel, U.; van der Schyf, C.J.; Klein, J. NMDA receptor-antagonistic properties of hyperforin, a constituent of St. John’s Wort. J. Pharmacol. Sci. 2006, 102, 47–54. [Google Scholar] [CrossRef]
- Tirona, R.G.; Leake, B.F.; Wolkoff, A.W.; Kim, R.B. Human organic anion transporting polypeptide-C (SLC21A6) is a major determinant of rifampin-mediated pregnane X receptor activation. J. Pharmacol. Exp. Ther. 2003, 304, 223–228. [Google Scholar] [CrossRef]
- Gutmann, H.; Poller, B.; Büter, K.B.; Pfrunder, A.; Schaffner, W.; Drewe, J. Hypericum perforatum: Which constituents may induce intestinal MDR1 and CYP3A4 mRNA expression? Planta Med. 2006, 72, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.F.; Acharya, M.R.; Desai, N.; Figg, W.D.; Sparreboom, A. Identification of OATP1B3 as a high-affinity hepatocellular transporter of paclitaxel. Cancer Biol. Ther. 2005, 4, 815–818. [Google Scholar] [CrossRef] [PubMed]
- Quiney, C.; Billard, C.; Faussat, A.-M.; Salanoubat, C.; Kolb, J.-P. Hyperforin inhibits P-gp and BCRP activities in chronic lymphocytic leukaemia cells and myeloid cells. Leuk. Lymphoma 2007, 48, 1587–1599. [Google Scholar] [CrossRef] [PubMed]
- You, M.-K.; Kim, H.-J.; Kook, J.H.; Kim, H.-A. St. John’s Wort Regulates Proliferation and Apoptosis in MCF-7 Human Breast Cancer Cells by Inhibiting AMPK/mTOR and Activating the Mitochondrial Pathway. Int. J. Mol. Sci. 2018, 19, 966. [Google Scholar] [CrossRef]
- Novelli, M.; Masiello, P.; Beffy, P.; Menegazzi, M. Protective Role of St. John’s Wort and Its Components Hyperforin and Hypericin against Diabetes through Inhibition of Inflammatory Signaling: Evidence from In Vitro and In Vivo Studies. Int. J. Mol. Sci. 2020, 21, 8108. [Google Scholar] [CrossRef]
- Suzuki, O.; Katsumata, Y.; Oya, M.; Bladt, S.; Wagner, H. Inhibition of monoamine oxidase by hypericin. Planta Med. 1984, 50, 272–274. [Google Scholar] [CrossRef]
- Cott, J.M. In vitro receptor binding and enzyme inhibition by Hypericum perforatum extract. Pharmacopsychiatry 1997, 30 (Suppl. 2), 108–112. [Google Scholar] [CrossRef]
- Šemeláková, M.; Jendželovský, R.; Fedoročko, P. Drug membrane transporters and CYP3A4 are affected by hypericin, hyperforin or aristoforin in colon adenocarcinoma cells. Biomed. Pharmacother. 2016, 81, 38–47. [Google Scholar] [CrossRef]
- Khot, M.I.; Perry, S.L.; Maisey, T.; Armstrong, G.; Andrew, H.; Hughes, T.A.; Kapur, N.; Jayne, D.G. Inhibiting ABCG2 could potentially enhance the efficacy of hypericin-mediated photodynamic therapy in spheroidal cell models of colorectal cancer. Photodiagnosis Photodyn. Ther. 2018, 23, 221–229. [Google Scholar] [CrossRef]
- Song, M.; Hong, M.; Lee, M.Y.; Jee, J.-G.; Lee, Y.M.; Bae, J.-S.; Jeong, T.C.; Lee, S. Selective inhibition of the cytochrome P450 isoform by hyperoside and its potent inhibition of CYP2D6. Food Chem. Toxicol. 2013, 59, 549–553. [Google Scholar] [CrossRef]
- Li, G.; Simmler, C.; Chen, L.; Nikolic, D.; Chen, S.-N.; Pauli, G.F.; van Breemen, R.B. Cytochrome P450 inhibition by three licorice species and fourteen licorice constituents. Eur. J. Pharm. Sci. 2017, 109, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, J.; Zeng, S. Involvement of P-glycoprotein in regulating cellular levels of Ginkgo flavonols: Quercetin, kaempferol, and isorhamnetin. J. Pharm. Pharmacol. 2005, 57, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Mohos, V.; Fliszár-Nyúl, E.; Ungvári, O.; Kuffa, K.; Needs, P.W.; Kroon, P.A.; Telbisz, Á.; Özvegy-Laczka, C.; Poór, M. Inhibitory Effects of Quercetin and Its Main Methyl, Sulfate, and Glucuronic Acid Conjugates on Cytochrome P450 Enzymes, and on OATP, BCRP and MRP2 Transporters. Nutrients 2020, 12, 2306. [Google Scholar] [CrossRef]
- Park, J.-W.; Choi, J.-S. Role of kaempferol to increase bioavailability and pharmacokinetics of nifedipine in rats. Chin. J. Nat. Med. 2019, 17, 690–697. [Google Scholar] [CrossRef]
- Piao, Y.; Shin, S.-C.; Choi, J.-S. Effects of oral kaempferol on the pharmacokinetics of tamoxifen and one of its metabolites, 4-hydroxytamoxifen, after oral administration of tamoxifen to rats. Biopharm. Drug Dispos. 2008, 29, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.; Oliveira, P.J.; Dias, A.; Malva, J.O. Quercetin, kaempferol and biapigenin from Hypericum perforatum are neuroprotective against excitotoxic insults. Neurotox. Res. 2008, 13, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Navrátilová, L.; Applová, L.; Horký, P.; Mladěnka, P.; Pávek, P.; Trejtnar, F. Interaction of soy isoflavones and their main metabolites with hOATP2B1 transporter. Naunyn. Schmiedebergs. Arch. Pharmacol. 2018, 391, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Vautier, S.; Milane, A.; Fernandez, C.; Buyse, M.; Chacun, H.; Farinotti, R. Interactions between antiparkinsonian drugs and ABCB1/P-glycoprotein at the blood-brain barrier in a rat brain endothelial cell model. Neurosci. Lett. 2008, 442, 19–23. [Google Scholar] [CrossRef]
- Blanchet, P.J.; Papa, S.M.; Metman, L.V.; Mouradian, M.M.; Chase, T.N. Modulation of levodopa-induced motor response complications by NMDA antagonists in Parkinson’s disease. Neurosci. Biobehav. Rev. 1997, 21, 447–453. [Google Scholar] [CrossRef]
- Hwang, J.-T.; Park, O.J.; Lee, Y.K.; Sung, M.J.; Hur, H.J.; Kim, M.S.; Ha, J.H.; Kwon, D.Y. Anti-tumor effect of luteolin is accompanied by AMP-activated protein kinase and nuclear factor-κB modulation in HepG2 hepatocarcinoma cells. Int. J. Mol. Med. 2011, 28, 25–31. [Google Scholar] [CrossRef]
- Harris, G.K.; Qian, Y.; Leonard, S.S.; Sbarra, D.C.; Shi, X. Luteolin and chrysin differentially inhibit cyclooxygenase-2 expression and scavenge reactive oxygen species but similarly inhibit prostaglandin-E2 formation in RAW 264.7 cells. J. Nutr. 2006, 136, 1517–1521. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Kwara, A.; Greenblatt, D.J. Metabolic interactions between acetaminophen (paracetamol) and two flavonoids, luteolin and quercetin, through in-vitro inhibition studies. J. Pharm. Pharmacol. 2017, 69, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- Quintieri, L.; Palatini, P.; Nassi, A.; Ruzza, P.; Floreani, M. Flavonoids diosmetin and luteolin inhibit midazolam metabolism by human liver microsomes and recombinant CYP 3A4 and CYP3A5 enzymes. Biochem. Pharmacol. 2008, 75, 1426–1437. [Google Scholar] [CrossRef] [PubMed]
- Wong, T.Y.; Lin, S.-M.; Leung, L.K. The Flavone Luteolin Suppresses SREBP-2 Expression and Post-Translational Activation in Hepatic Cells. PLoS ONE 2015, 10, e0135637. [Google Scholar] [CrossRef] [PubMed]
- Han, X.H.; Hong, S.S.; Hwang, J.S.; Lee, M.K.; Hwang, B.Y.; Ro, J.S. Monoamine oxidase inhibitory components from Cayratia japonica. Arch. Pharm. Res. 2007, 30, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Boersma, M.G.; van der Woude, H.; Bogaards, J.; Boeren, S.; Vervoort, J.; Cnubben, N.H.P.; van Iersel, M.L.P.S.; van Bladeren, P.J.; Rietjens, I.M.C.M. Regioselectivity of phase II metabolism of luteolin and quercetin by UDP-glucuronosyl transferases. Chem. Res. Toxicol. 2002, 15, 662–670. [Google Scholar] [CrossRef] [PubMed]
- McNulty, J.; Nair, J.J.; Singh, M.; Crankshaw, D.J.; Holloway, A.C.; Bastida, J. Cytochrome P450 3A4 inhibitory activity studies within the lycorine series of alkaloids. Nat. Prod. Commun. 2010, 5, 1195–1200. [Google Scholar] [CrossRef]
- Hanley, M.J.; Masse, G.; Harmatz, J.S.; Cancalon, P.F.; Dolnikowski, G.G.; Court, M.H.; Greenblatt, D.J. Effect of blueberry juice on clearance of buspirone and flurbiprofen in human volunteers. Br. J. Clin. Pharmacol. 2013, 75, 1041–1052. [Google Scholar] [CrossRef]
- Li, Y.; Xu, Y.; Xie, J.; Chen, W. Malvidin-3-O-arabinoside ameliorates ethyl carbamate-induced oxidative damage by stimulating AMPK-mediated autophagy. Food Funct. 2020, 11, 10317–10328. [Google Scholar] [CrossRef]
- Showande, S.J.; Fakeye, T.O.; Kajula, M.; Hokkanen, J.; Tolonen, A. Potential inhibition of major human cytochrome P450 isoenzymes by selected tropical medicinal herbs—Implication for herb–drug interactions. Food Sci. Nutr. 2019, 7, 44–55. [Google Scholar] [CrossRef]
- Rodeiro, I.; José Gómez-Lechón, M.; Perez, G.; Hernandez, I.; Herrera, J.A.; Delgado, R.; Castell, J.V.; Teresa Donato, M. Mangifera indica L. extract and mangiferin modulate cytochrome P450 and UDP-glucuronosyltransferase enzymes in primary cultures of human hepatocytes. Phytother. Res. 2013, 27, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Leslie, E.M.; Mao, Q.; Oleschuk, C.J.; Deeley, R.G.; Cole, S.P. Modulation of multidrug resistance protein 1 (MRP1/ABCC1) transport and atpase activities by interaction with dietary flavonoids. Mol. Pharmacol. 2001, 59, 1171–1180. [Google Scholar] [CrossRef] [PubMed]
- Surya Sandeep, M.; Sridhar, V.; Puneeth, Y.; Ravindra Babu, P.; Naveen Babu, K. Enhanced oral bioavailability of felodipine by naringenin in Wistar rats and inhibition of P-glycoprotein in everted rat gut sacs in vitro. Drug Dev. Ind. Pharm. 2014, 40, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Zygmunt, K.; Faubert, B.; MacNeil, J.; Tsiani, E. Naringenin, a citrus flavonoid, increases muscle cell glucose uptake via AMPK. Biochem. Biophys. Res. Commun. 2010, 398, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.-L.; Weng, C.-S.; Chang, N.-C.; Lin, J.-S.; Kao, S.-T.; Ho, F.-M. Naringenin more effectively inhibits inducible nitric oxide synthase and cyclooxygenase-2 expression in macrophages than in microglia. Nutr. Res. 2010, 30, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Zeng, X.; Wu, H.; Su, W.; Fan, W.; Bai, Y.; Yao, H.; Li, P. Effects of Naringin on the Activity and mRNA Expression of CYP Isozymes in Rats. Nat. Prod. Commun. 2019, 14, 1–8. [Google Scholar] [CrossRef]
- Ho, P.C.; Saville, D.J.; Coville, P.F.; Wanwimolruk, S. Content of CYP3A4 inhibitors, naringin, naringenin and bergapten in grapefruit and grapefruit juice products. Pharm. Acta Helv. 2000, 74, 379–385. [Google Scholar] [CrossRef]
- Lu, W.J.; Ferlito, V.; Xu, C.; Flockhart, D.A.; Caccamese, S. Enantiomers of naringenin as pleiotropic, stereoselective inhibitors of cytochrome P450 isoforms. Chirality 2011, 23, 891–896. [Google Scholar] [CrossRef]
- Lee, S.H.; Park, Y.B.; Bae, K.H.; Bok, S.H.; Kwon, Y.K.; Lee, E.S.; Choi, M.S. Cholesterol-lowering activity of naringenin via inhibition of 3-hydroxy-3-methylglutaryl coenzyme A reductase and acyl coenzyme A:cholesterol acyltransferase in rats. Ann. Nutr. Metab. 1999, 43, 173–180. [Google Scholar] [CrossRef]
- Rani, N.; Bharti, S.; Krishnamurthy, B.; Bhatia, J.; Sharma, C.; Kamal, M.A.; Ojha, S.; Arya, D.S. Pharmacological Properties and Therapeutic Potential of Naringenin: A Citrus Flavonoid of Pharmaceutical Promise. Curr. Pharm. Des. 2016, 22, 4341–4359. [Google Scholar] [CrossRef]
- Satoh, H.; Yamashita, F.; Tsujimoto, M.; Murakami, H.; Koyabu, N.; Ohtani, H.; Sawada, Y. Citrus juices inhibit the function of human organic anion-transporting polypeptide OATP-B. Drug Metab. Dispos. 2005, 33, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.S.; Seo, K.; Lim, S.-C.; Han, H.-K. Interaction characteristics of flavonoids with human organic anion transporter 1 (hOAT1) and 3 (hOAT3). Pharmacol. Res. 2007, 56, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Eagling, V.A.; Profit, L.; Back, D.J. Inhibition of the CYP3A4-mediated metabolism and P-glycoprotein-mediated transport of the HIV-1 protease inhibitor saquinavir by grapefruit juice components. Br. J. Clin. Pharmacol. 1999, 48, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Sui, G.-G.; Xiao, H.-B.; Lu, X.-Y.; Sun, Z.-L. Naringin Activates AMPK Resulting in Altered Expression of SREBPs, PCSK9, and LDLR To Reduce Body Weight in Obese C57BL/6J Mice. J. Agric. Food Chem. 2018, 66, 8983–8990. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Zhen, Y.; Chen, Y.; Zou, L.; Zhang, Y.; Hu, F.; Feng, J.; Shen, J.; Wei, B. Naringin inhibits growth and induces apoptosis by a mechanism dependent on reduced activation of NF-κB/COX-2-caspase-1 pathway in HeLa cervical cancer cells. Int. J. Oncol. 2014, 45, 1929–1936. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.S.; Do, K.M.; Park, Y.S.; Jeon, S.M.; Jeong, T.S.; Lee, Y.K.; Lee, M.K.; Bok, S.H. Effect of naringin supplementation on cholesterol metabolism and antioxidant status in rats fed high cholesterol with different levels of vitamin E. Ann. Nutr. Metab. 2001, 45, 193–201. [Google Scholar] [CrossRef]
- Shirasaka, Y.; Shichiri, M.; Mori, T.; Nakanishi, T.; Tamai, I. Major active components in grapefruit, orange, and apple juices responsible for OATP2B1-mediated drug interactions. J. Pharm. Sci. 2013, 102, 280–288. [Google Scholar] [CrossRef]
- Nabekura, T.; Yamaki, T.; Kitagawa, S. Effects of chemopreventive citrus phytochemicals on human P-glycoprotein and multidrug resistance protein 1. Eur. J. Pharmacol. 2008, 600, 45–49. [Google Scholar] [CrossRef]
- Huang, H.; Li, L.; Shi, W.; Liu, H.; Yang, J.; Yuan, X.; Wu, L. The Multifunctional Effects of Nobiletin and Its Metabolites In Vivo and In Vitro. Evid. Based Complement. Altern. Med. 2016, 2016, 2918796. [Google Scholar] [CrossRef]
- Weiss, J.; Gattuso, G.; Barreca, D.; Haefeli, W.E. Nobiletin, sinensetin, and tangeretin are the main perpetrators in clementines provoking food-drug interactions in vitro. Food Chem. 2020, 319, 126578. [Google Scholar] [CrossRef]
- Bajraktari-Sylejmani, G.; Weiss, J. Potential Risk of Food-Drug Interactions: Citrus Polymethoxyflavones and Flavanones as Inhibitors of the Organic Anion Transporting Polypeptides (OATP) 1B1, 1B3, and 2B1. Eur. J. Drug Metab. Pharmacokinet. 2020, 45, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Braga, F.; Ayres-Saraiva, D.; Gattass, C.R.; Capella, M.A.M. Oleanolic acid inhibits the activity of the multidrug resistance protein ABCC1 (MRP1) but not of the ABCB1 (P-glycoprotein): Possible use in cancer chemotherapy. Cancer Lett. 2007, 248, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, H.; Hu, K.; Xu, Q.; Wen, X.; Cheng, K.; Chen, C.; Yuan, H.; Dai, L.; Sun, H. Synthesis and anti-inflammatory activity of saponin derivatives of δ-oleanolic acid. Eur. J. Med. Chem. 2020, 112932. [Google Scholar] [CrossRef] [PubMed]
- Martínez-González, J.; Rodríguez-Rodríguez, R.; González-Díez, M.; Rodríguez, C.; Herrera, M.D.; Ruiz-Gutierrez, V.; Badimon, L. Oleanolic acid induces prostacyclin release in human vascular smooth muscle cells through a cyclooxygenase-2-dependent mechanism. J. Nutr. 2008, 138, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-A.; Lee, J.-S.; Park, H.-J.; Kim, J.-W.; Kim, C.-J.; Shim, I.-S.; Kim, N.-J.; Han, S.-M.; Lim, S. Inhibition of cytochrome P450 activities by oleanolic acid and ursolic acid in human liver microsomes. Life Sci. 2004, 74, 2769–2779. [Google Scholar] [CrossRef]
- Shen, C.; Huang, L.; Xiang, H.; Deng, M.; Gao, H.; Zhu, Z.; Liu, M.; Luo, G. Inhibitory effects on the HMG-CoA reductase in the chemical constituents of the Cassia mimosoides Linn. Rev. Romana Med. Lab. 2016, 24, 413–422. [Google Scholar] [CrossRef]
- Fajemiroye, J.O.; Polepally, P.R.; Chaurasiya, N.D.; Tekwani, B.L.; Zjawiony, J.K.; Costa, E.A. Oleanolic acid acrylate elicits antidepressant-like effect mediated by 5-HT1A receptor. Sci. Rep. 2015, 5, 11582. [Google Scholar] [CrossRef]
- Rodrigues, M.; Alves, G.; Falcão, A. Investigating herb-drug interactions: The effect of Citrus aurantium fruit extract on the pharmacokinetics of amiodarone in rats. Food Chem. Toxicol. 2013, 60, 153–159. [Google Scholar] [CrossRef]
- Tiesjema, B.; Jeurissen, S.M.F.; De Wit, L.; Mol, H.; Fragki, S.; Razenberg, L. Risk Assessment of Synephrine; Ministry of Health, Welfare and Sport: Bilthoven, The Netherlands, 2017.
- Suzuki, O.; Matsumoto, T.; Oya, M.; Katsumata, Y. Oxidation of synephrine by type A and type B monoamine oxidase. Experientia 1979, 35, 1283–1284. [Google Scholar] [CrossRef]
- Morsy, M.A.; Abdel-Aziz, A.M.; Abdel-Hafez, S.M.N.; Venugopala, K.N.; Nair, A.B.; Abdel-Gaber, S.A. The Possible Contribution of P-Glycoprotein in the Protective Effect of Paeonol against Methotrexate-Induced Testicular Injury in Rats. Pharmaceuticals 2020, 13, 223. [Google Scholar] [CrossRef]
- Li, M.; Tan, S.-Y.; Wang, X.-F. Paeonol exerts an anticancer effect on human colorectal cancer cells through inhibition of PGE₂ synthesis and COX-2 expression. Oncol. Rep. 2014, 32, 2845–2853. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-X.; Hu, Y.; Liu, Y.; He, Y.-Q.; Li, W.; Yang, L. CYP1A2 is the major isoform responsible for paeonol o-demethylation in human liver microsomes. Xenobiotica 2009, 39, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Chen, S.; Zhang, W.; Hu, S.; Lu, J.; Xing, J.; Dong, Y. Paeonol reverses paclitaxel resistance in human breast cancer cells by regulating the expression of transgelin 2. Phytomedicine 2014, 21, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.D.; Cheng, C.H.K.; Tan, R.X. Inhibition of MAO A and B by some plant-derived alkaloids, phenols and anthraquinones. J. Ethnopharmacol. 2004, 91, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Ekeuku, S.O.; Pang, K.-L.; Chin, K.-Y. Palmatine as an Agent Against Metabolic Syndrome and Its Related Complications: A Review. Drug Des. Dev. Ther. 2020, 14, 4963–4974. [Google Scholar] [CrossRef]
- Lo, S.-N.; Chang, Y.-P.; Tsai, K.-C.; Chang, C.-Y.; Wu, T.-S.; Ueng, Y.-F. Inhibition of CYP1 by berberine, palmatine, and jatrorrhizine: Selectivity, kinetic characterization, and molecular modeling. Toxicol. Appl. Pharmacol. 2013, 272, 671–680. [Google Scholar] [CrossRef]
- Vrba, J.; Papouskova, B.; Pyszkova, M.; Zatloukalova, M.; Lemr, K.; Ulrichova, J.; Vacek, J. Metabolism of palmatine by human hepatocytes and recombinant cytochromes P450. J. Pharm. Biomed. Anal. 2015, 102, 193–198. [Google Scholar] [CrossRef]
- Tarabasz, D.; Kukula-Koch, W. Palmatine: A review of pharmacological properties and pharmacokinetics. Phytother. Res. 2020, 34, 33–50. [Google Scholar] [CrossRef]
- Sukhaphirom, N.; Vardhanabhuti, N.; Chirdchupunseree, H.; Pramyothin, P.; Jianmongkol, S. Phyllanthin and hypophyllanthin inhibit function of P-gp but not MRP2 in Caco-2 cells. J. Pharm. Pharmacol. 2013, 65, 292–299. [Google Scholar] [CrossRef]
- Taesotikul, T.; Dumrongsakulchai, W.; Wattanachai, N.; Navinpipat, V.; Somanabandhu, A.; Tassaneeyakul, W.; Tassaneeyakul, W. Inhibitory effects of Phyllanthus amarus and its major lignans on human microsomal cytochrome P450 activities: Evidence for CYP3A4 mechanism-based inhibition. Drug Metab. Pharmacokinet. 2011, 26, 154–161. [Google Scholar] [CrossRef]
- Zhang, F.; Ai, W.; Hu, X.; Meng, Y.; Yuan, C.; Su, H.; Wang, L.; Zhu, X.; Gao, P.; Shu, G.; et al. Phytol stimulates the browning of white adipocytes through the activation of AMP-activated protein kinase (AMPK) α in mice fed high-fat diet. Food Funct. 2018, 9, 2043–2050. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Ray, P.; Khalipha, A.B.R.; Hafiz Hassan, S.M.; Khan, M.R.; Rouf, R. Molecular docking study of the phytol and its derivatives against COX-2 induced inflammation: A combined density functional study. RRST 2020, 12, 1–5. [Google Scholar] [CrossRef]
- de Moraes, J.; de Oliveira, R.N.; Costa, J.P.; Junior, A.L.G.; de Sousa, D.P.; Freitas, R.M.; Allegretti, S.M.; Pinto, P.L.S. Phytol, a diterpene alcohol from chlorophyll, as a drug against neglected tropical disease Schistosomiasis mansoni. PLoS Negl. Trop. Dis. 2014, 8, e2617. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-J.; Yang, Y.-J.; Li, Y.-S.; Zhang, W.K.; Tang, H.-B. α-Pinene, linalool, and 1-octanol contribute to the topical anti-inflammatory and analgesic activities of frankincense by inhibiting COX-2. J. Ethnopharmacol. 2016, 179, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Wilderman, P.R.; Shah, M.B.; Jang, H.-H.; Stout, C.D.; Halpert, J.R. Structural and thermodynamic basis of (+)-α-pinene binding to human cytochrome P450 2B6. J. Am. Chem. Soc. 2013, 135, 10433–10440. [Google Scholar] [CrossRef]
- Ueno, H.; Shimada, A.; Suemitsu, S.; Murakami, S.; Kitamura, N.; Wani, K.; Matsumoto, Y.; Okamoto, M.; Ishihara, T. Attenuation Effects of Alpha-Pinene Inhalation on Mice with Dizocilpine-Induced Psychiatric-Like Behaviour. Evid. Based Complement. Altern. Med. 2019, 2019, 2745453. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, H.Y.; Back, S.Y.; Han, H.-K. Piperine-mediated drug interactions and formulation strategy for piperine: Recent advances and future perspectives. Expert Opin. Drug Metab. Toxicol. 2018, 14, 43–57. [Google Scholar] [CrossRef]
- Han, Y.; Tan, T.M.C.; Lim, L.-Y. In vitro and in vivo evaluation of the effects of piperine on P-gp function and expression. Toxicol. Appl. Pharmacol. 2008, 230, 283–289. [Google Scholar] [CrossRef]
- Li, S.; Lei, Y.; Jia, Y.; Li, N.; Wink, M.; Ma, Y. Piperine, a piperidine alkaloid from Piper nigrum re-sensitizes P-gp, MRP1 and BCRP dependent multidrug resistant cancer cells. Phytomedicine 2011, 19, 83–87. [Google Scholar] [CrossRef]
- Li, Z.-L.; Dong, X.-Z.; Wang, D.-X.; Dong, R.-H.; Guo, T.-T.; Sun, Y.; Liu, P. Effect of oligosaccharide esters and polygalaxanthone Ill from Polygala tenuifolia willd towards cytochrome P450. Zhongguo Zhong Yao Za Zhi 2014, 39, 4459–4463. [Google Scholar]
- Krajka-Kuźniak, V.; Szaefer, H.; Baer-Dubowska, W. Modulation of cytochrome P450 and phase II enzymes by protocatechuic acid in mouse liver and kidney. Toxicology 2005, 216, 24–31. [Google Scholar] [CrossRef]
- Zhang, M.-F.; Liu, Y.-X.; Jiang, K.-Y.; Niu, H.-M.; Jiang, J.-L.; Dong, S.-T.; Wang, X.; Wang, D.-F.; Meng, S.-N. Alteration of UDP-glucuronosyltransferase 1a1, 1a7 and P-glycoprotein expression in hepatic fibrosis rats and the impact on pharmacokinetics of puerarin. Phytomedicine 2019, 52, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shan, H.; Wang, B.; Wang, N.; Zhou, Z.; Pan, C.; Wang, F. Puerarin Attenuates Osteoarthritis via Upregulating AMP-Activated Protein Kinase/Proliferator-Activated Receptor-γ Coactivator-1 Signaling Pathway in Osteoarthritis Rats. Pharmacology 2018, 102, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Yang, X.; Zhe, C.; Zhang, Q.; Sun, L.; Cao, K. Puerarin inhibits iNOS, COX-2 and CRP expression via suppression of NF-κB activation in LPS-induced RAW264.7 macrophage cells. Pharmacol. Rep. 2011, 63, 781–789. [Google Scholar] [CrossRef]
- Ge, B.; Zhang, Z.; Lam, T.T.; Zuo, Z. Puerarin offsets the anticoagulation effect of warfarin in rats by inducing rCyps, upregulating vitamin K epoxide reductase and inhibiting thrombomodulin. Biopharm. Drug Dispos. 2017, 38, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Chen, B.; Jiang, B.; Zeng, L.; Tang, Z.-R.; Fan, L.; Zhou, H.-H. The effects of puerarin on CYP2D6 and CYP1A2 activities in vivo. Arch. Pharm. Res. 2010, 33, 243–246. [Google Scholar] [CrossRef]
- Guo, Y.-J.; Liang, D.-L.; Xu, Z.-S.; Ye, Q. In vivo inhibitory effects of puerarin on selected rat cytochrome P450 isoenzymes. Pharmazie 2014, 69, 367–370. [Google Scholar]
- Hwang, Y.P.; Choi, C.Y.; Chung, Y.C.; Jeon, S.S.; Jeong, H.G. Protective effects of puerarin on carbon tetrachloride-induced hepatotoxicity. Arch. Pharm. Res. 2007, 30, 1309–1317. [Google Scholar] [CrossRef]
- Chung, M.J.; Sung, N.-J.; Park, C.-S.; Kweon, D.-K.; Mantovani, A.; Moon, T.-W.; Lee, S.-J.; Park, K.-H. Antioxidative and hypocholesterolemic activities of water-soluble puerarin glycosides in HepG2 cells and in C57 BL/6J mice. Eur. J. Pharmacol. 2008, 578, 159–170. [Google Scholar] [CrossRef]
- Zhang, R.; Guo, H.-N.; Wu, H.-Q.; Cheng, H.-X.; Wang, H.-Q. Effect of puerarin on the expression of NMDA receptor in the hippocampus CA1 region after focal cerebral ischemia in rats. Sichuan Da Xue Xue Bao Yi Xue Ban 2011, 42, 52–55. [Google Scholar]
- Borska, S.; Sopel, M.; Chmielewska, M.; Zabel, M.; Dziegiel, P. Quercetin as a potential modulator of P-glycoprotein expression and function in cells of human pancreatic carcinoma line resistant to daunorubicin. Molecules 2010, 15, 857–870. [Google Scholar] [CrossRef]
- Yang, T.; Liu, Y.; Huang, X.; Zhang, R.; Yang, C.; Zhou, J.; Zhang, Y.; Wan, J.; Shi, S. Quercetin-3-O-β-d-glucoside decreases the bioavailability of cyclosporin A through regulation of drug metabolizing enzymes, transporters and nuclear receptors in rats. Mol. Med. Rep. 2018, 18, 2599–2612. [Google Scholar] [CrossRef]
- Elbarbry, F.; Ung, A.; Abdelkawy, K. Studying the Inhibitory Effect of Quercetin and Thymoquinone on Human Cytochrome P450 Enzyme Activities. Pharmacogn. Mag. 2018, 13, S895–S899. [Google Scholar] [CrossRef]
- Walsky, R.L.; Gaman, E.A.; Obach, R.S. Examination of 209 drugs for inhibition of cytochrome P450 2C8. J. Clin. Pharmacol. 2005, 45, 68–78. [Google Scholar] [CrossRef]
- Chimenti, F.; Cottiglia, F.; Bonsignore, L.; Casu, L.; Casu, M.; Floris, C.; Secci, D.; Bolasco, A.; Chimenti, P.; Granese, A.; et al. Quercetin as the active principle of Hypericum hircinum exerts a selective inhibitory activity against MAo-A: Extraction, biological analysis, and computational study. J. Nat. Prod. 2006, 69, 945–949. [Google Scholar] [CrossRef]
- Park, J.-Y.; Lim, M.-S.; Kim, S.-I.; Lee, H.J.; Kim, S.-S.; Kwon, Y.-S.; Chun, W. Quercetin-3-O-β-d-Glucuronide Suppresses Lipopolysaccharide-Induced JNK and ERK Phosphorylation in LPS-Challenged RAW264.7 Cells. Biomol. Ther. 2016, 24, 610–615. [Google Scholar] [CrossRef]
- Aires, V.; Colin, D.J.; Doreau, A.; Pietro, A.D.; Heydel, J.M.; Artur, Y.; Latruffe, N.; Delmas, D. P-glycoprotein 1 affects chemoactivities of resveratrol against human colorectal cancer cells. Nutrients 2019, 11, 2098. [Google Scholar] [CrossRef]
- Detampel, P.; Beck, M.; Krähenbühl, S.; Huwyler, J. Drug interaction potential of resveratrol. Drug Metab. Rev. 2012, 44, 253–265. [Google Scholar] [CrossRef]
- Dao, T.-M.A.; Waget, A.; Klopp, P.; Serino, M.; Vachoux, C.; Pechere, L.; Drucker, D.J.; Champion, S.; Barthélemy, S.; Barra, Y.; et al. Resveratrol increases glucose induced GLP-1 secretion in mice: A mechanism which contributes to the glycemic control. PLoS ONE 2011, 6, e20700. [Google Scholar] [CrossRef]
- Cho, I.J.; Ahn, J.Y.; Kim, S.; Choi, M.S.; Ha, T.Y. Resveratrol attenuates the expression of HMG-CoA reductase mRNA in hamsters. Biochem. Biophys. Res. Commun. 2008, 367, 190–194. [Google Scholar] [CrossRef]
- Zhang, Z.; Hamada, H.; Gerk, P.M. Selectivity of Dietary Phenolics for Inhibition of Human Monoamine Oxidases A and B. Biomed Res. Int. 2019, 2019, 8361858. [Google Scholar] [CrossRef]
- Maier-Salamon, A.; Böhmdorfer, M.; Riha, J.; Thalhammer, T.; Szekeres, T.; Jaeger, W. Interplay between metabolism and transport of resveratrol. Ann. N. Y. Acad. Sci. 2013, 1290, 98–106. [Google Scholar] [CrossRef]
- Takehana, S.; Kubota, Y.; Uotsu, N.; Yui, K.; Iwata, K.; Shimazu, Y.; Takeda, M. The dietary constituent resveratrol suppresses nociceptive neurotransmission via the NMDA receptor. Mol. Pain 2017, 13. [Google Scholar] [CrossRef]
- van de Wetering, K.; Burkon, A.; Feddema, W.; Bot, A.; de Jonge, H.; Somoza, V.; Borst, P. Intestinal breast cancer resistance protein (BCRP)/Bcrp1 and multidrug resistance protein 3 (MRP3)/Mrp3 are involved in the pharmacokinetics of resveratrol. Mol. Pharmacol. 2009, 75, 876–885. [Google Scholar] [CrossRef]
- Thu, O.K.; Nilsen, O.G.; Hellum, B. In vitro inhibition of cytochrome P-450 activities and quantification of constituents in a selection of commercial Rhodiola rosea products. Pharm. Biol. 2016, 54, 3249–3256. [Google Scholar] [CrossRef]
- van Diermen, D.; Marston, A.; Bravo, J.; Reist, M.; Carrupt, P.A.; Hostettmann, K. Monoamine oxidase inhibition by Rhodiola rosea L. roots. J. Ethnopharmacol. 2009, 122, 397–401. [Google Scholar] [CrossRef]
- Kim, S.-B.; Kim, K.-S.; Kim, D.-D.; Yoon, I.-S. Metabolic interactions of rosmarinic acid with human cytochrome P450 monooxygenases and uridine diphosphate glucuronosyltransferases. Biomed. Pharmacother. 2019, 110, 111–117. [Google Scholar] [CrossRef]
- Li, M.; Yin, D.; Li, J.; Shao, F.; Zhang, Q.; Jiang, Q.; Zhang, M.; Yang, Y. Rosmarinic acid, the active component of, improves gliquidone transport by regulating the expression and function of P-gp and BCRP in Caco-2 cells. Pharmazie 2020, 75, 18–22. [Google Scholar] [CrossRef]
- Mohana, S.; Ganesan, M.; Agilan, B.; Karthikeyan, R.; Srithar, G.; Beaulah Mary, R.; Ananthakrishnan, D.; Velmurugan, D.; Rajendra Prasad, N.; Ambudkar, S.V. Screening dietary flavonoids for the reversal of P-glycoprotein-mediated multidrug resistance in cancer. Mol. Biosyst. 2016, 12, 2458–2470. [Google Scholar] [CrossRef]
- Seo, S.; Lee, M.-S.; Chang, E.; Shin, Y.; Oh, S.; Kim, I.-H.; Kim, Y. Rutin Increases Muscle Mitochondrial Biogenesis with AMPK Activation in High-Fat Diet-Induced Obese Rats. Nutrients 2015, 7, 8152–8169. [Google Scholar] [CrossRef]
- Fideles, L.D.S.; de Miranda, J.A.L.; Martins, C.D.S.; Barbosa, M.L.L.; Pimenta, H.B.; Pimentel, P.V.D.S.; Teixeira, C.S.; Scafuri, M.A.S.; Façanha, S.; de Façanha, S.O.; et al. Role of Rutin in 5-Fluorouracil-Induced Intestinal Mucositis: Prevention of Histological Damage and Reduction of Inflammation and Oxidative Stress. Molecules 2020, 25, 2786. [Google Scholar] [CrossRef]
- Karakurt, S. Modulatory effects of rutin on the expression of cytochrome P450s and antioxidant enzymes in human hepatoma cells. Acta Pharm. 2016, 66, 491–502. [Google Scholar] [CrossRef]
- Suganya, S.; Nandagopal, B.; Anbarasu, A. Natural Inhibitors of HMG-CoA Reductase-An In silico Approach Through Molecular Docking and Simulation Studies. J. Cell. Biochem. 2017, 118, 52–57. [Google Scholar] [CrossRef]
- Azam, F.; Abodabos, H.S.; Taban, I.M.; Rfieda, A.R.; Mahmood, D.; Anwar, M.J.; Khan, S.; Sizochenko, N.; Poli, G.; Tuccinardi, T.; et al. Rutin as promising drug for the treatment of Parkinson’s disease: An assessment of MAo-B inhibitory potential by docking, molecular dynamics and DFT studies. Mol. Simul. 2019, 45, 1563–1571. [Google Scholar] [CrossRef]
- Ogura, J.; Koizumi, T.; Segawa, M.; Yabe, K.; Kuwayama, K.; Sasaki, S.; Kaneko, C.; Tsujimoto, T.; Kobayashi, M.; Yamaguchi, H.; et al. Quercetin-3-rhamnoglucoside (rutin) stimulates transport of organic anion compounds mediated by organic anion transporting polypeptide 2B1. Biopharm. Drug Dispos. 2014, 35, 173–182. [Google Scholar] [CrossRef]
- Iriti, M.; Kubina, R.; Cochis, A.; Sorrentino, R.; Varoni, E.M.; Kabała-Dzik, A.; Azzimonti, B.; Dziedzic, A.; Rimondini, L.; Wojtyczka, R.D. Rutin, a Quercetin Glycoside, Restores Chemosensitivity in Human Breast Cancer Cells. Phytother. Res. 2017, 31, 1529–1538. [Google Scholar] [CrossRef]
- Dogra, A.; Kotwal, P.; Gour, A.; Bhatt, S.; Singh, G.; Mukherjee, D.; Nandi, U. Description of Druglike Properties of Safranal and Its Chemistry behind Low Oral Exposure. ACS Omega 2020, 5, 9885–9891. [Google Scholar] [CrossRef]
- Hellum, B.H.; Tosse, A.; Hoybakk, K.; Thomsen, M.; Rohloff, J.; Georg Nilsen, O. Potent in vitro Inhibition of CYP3A4 and P-Glycoprotein by Rhodiola rosea. Planta Med. 2010, 76, 331–338. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, H.; Liu, X.; Chen, H.; Feng, N.; Zhang, J.; Wang, C.; Qiu, M.; Yang, J.; Zhou, X. Frontline Science: Reprogramming COX-2, 5-LOX, and CYP4A-mediated arachidonic acid metabolism in macrophages by salidroside alleviates gouty arthritis. J. Leukoc. Biol. 2019, 105, 11–24. [Google Scholar] [CrossRef]
- Chiang, H.-M.; Chen, H.-C.; Wu, C.-S.; Wu, P.-Y.; Wen, K.-C. Rhodiola plants: Chemistry and biological activity. J. Food Drug Anal. 2015, 23, 359–369. [Google Scholar] [CrossRef]
- Wei, Y.-L.; Du, H.-J.; Lin, Y.-P.; Wu, M.-L.; Xu, R.-A. Effects of salidroside on rat CYP enzymes by a cocktail of probe drugs. Iran. J. Basic Med. Sci. 2018, 21, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Coors, A.; Brosch, M.; Kahl, E.; Khalil, R.; Michels, B.; Laub, A.; Franke, K.; Gerber, B.; Fendt, M. Rhodiola rosea root extract has antipsychotic-like effects in rodent models of sensorimotor gating. J. Ethnopharmacol. 2019, 235, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Zhao, W.; Yang, Y.; Wu, S.; Lv, B. Salidroside could enhance the cytotoxic effect of L-OHP on colorectal cancer cells. Mol. Med. Rep. 2018, 17, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Wang, S.; Liang, W.; Wang, W.; Wang, H.; Zhao, M.; Liu, X. Salvianolic acid B reverses multidrug resistance in HCT-8/VCR human colorectal cancer cells by increasing ROS levels. Mol. Med. Rep. 2017, 15, 724–730. [Google Scholar] [CrossRef]
- Qin, T.; Rasul, A.; Sarfraz, A.; Sarfraz, I.; Hussain, G.; Anwar, H.; Riaz, A.; Liu, S.; Wei, W.; Li, J.; et al. Salvianolic acid A & B: Potential cytotoxic polyphenols in battle against cancer via targeting multiple signaling pathways. Int. J. Biol. Sci. 2019, 15, 2256–2264. [Google Scholar] [CrossRef]
- Wang, Q.-L.; Wu, Q.; Tao, Y.-Y.; Liu, C.-H.; El-Nezami, H. Salvianolic acid B modulates the expression of drug-metabolizing enzymes in HepG2 cells. Hepatobiliary Pancreat. Dis. Int. 2011, 10, 502–508. [Google Scholar] [CrossRef]
- Li, J.; Olaleye, O.E.; Yu, X.; Jia, W.; Yang, J.; Lu, C.; Liu, S.; Yu, J.; Duan, X.; Wang, Y.; et al. High degree of pharmacokinetic compatibility exists between the five-herb medicine XueBiJing and antibiotics comedicated in sepsis care. Acta Pharm. Sin. B 2019, 9, 1035–1049. [Google Scholar] [CrossRef]
- Küblbeck, J.; Hakkarainen, J.J.; Petsalo, A.; Vellonen, K.-S.; Tolonen, A.; Reponen, P.; Forsberg, M.M.; Honkakoski, P. Genetically Modified Caco-2 Cells With Improved Cytochrome P450 Metabolic Capacity. J. Pharm. Sci. 2016, 105, 941–949. [Google Scholar] [CrossRef]
- Ren, L.-X.; Luo, Y.-F.; Li, X.; Zuo, D.-Y.; Wu, Y.-L. Antidepressant-like effects of sarsasapogenin from Anemarrhena asphodeloides BUNGE (Liliaceae). Biol. Pharm. Bull. 2006, 29, 2304–2306. [Google Scholar] [CrossRef]
- Qiangrong, P.; Wang, T.; Lu, Q.; Hu, X. Schisandrin B—A novel inhibitor of P-glycoprotein. Biochem. Biophys. Res. Commun. 2005, 335, 406–411. [Google Scholar] [CrossRef]
- Li, W.L.; Xin, H.W.; Su, M.W.; Xiong, L. Inhibitory effects of schisandrin A and schisandrin B on CYP3A activity. Methods Find. Exp. Clin. Pharmacol. 2010, 32, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Lin, J.; Kang, J.; Zheng, X.; Fang, K.; Luo, Y.; Liu, A.; Yang, J. Schisandrin B and Schisandrol B induce mouse CYP2b10 associated with CAR not PXR. Phytochem. Lett. 2020, 35, 164–170. [Google Scholar] [CrossRef]
- Chiu, P.Y.; Leung, H.Y.; Poon, M.K.T.; Lee, S.S.T.; Ko, K.M. Schisandrin B induced antioxidant response is partly mediated by cytochrome P-4502E1 catalyzed reaction in mouse liver. Mol. Cell. Biochem. 2006, 293, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Hu, Q.; Chen, L.; Zhang, H.; Huang, S.; Xiong, Y.; Xia, C. Interaction of deoxyschizandrin and schizandrin B with liver uptake transporters OATP1B1 and OATP1B3. Xenobiotica 2019, 49, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Singla, R.; Jaitak, V. Shatavari (Asparagus racemosus wild): A review on its cultivation, morphology, phytochemistry and pharmacological importance. Int. J. Pharm. Sci. Res. 2014, 5, 742–757. [Google Scholar] [CrossRef]
- Azam, F.; Amer, A.M.; Abulifa, A.R.; Elzwawi, M.M. Ginger components as new leads for the design and development of novel multi-targeted anti-Alzheimer’s drugs: A computational investigation. Drug Des. Dev. Ther. 2014, 8, 2045–2059. [Google Scholar] [CrossRef]
- Kim, J. Effects of 6-Shogaol, A Major Component of Zingiber officinale Roscoe, on Human Cytochrome P450 Enzymes in vitro. Korean J. Med. Crop. Sci. 2016, 24, 7–13. [Google Scholar] [CrossRef]
- Fajrin, F.A.; Nugroho, A.E.; Nurrochmad, A.; Susilowati, R. Ginger extract and its compound, 6-shogaol, attenuates painful diabetic neuropathy in mice via reducing TRPV1 and NMDAR2B expressions in the spinal cord. J. Ethnopharmacol. 2020, 249, 112396. [Google Scholar] [CrossRef]
- Wang, P.; Zhao, Y.; Zhu, Y.; Sang, S. Glucuronidation and its impact on the bioactivity of [6]-shogaol. Mol. Nutr. Food Res. 2017, 61, 1700023. [Google Scholar] [CrossRef]
- Silva, N.; Salgueiro, L.; Fortuna, A.; Cavaleiro, C. P-glycoprotein Mediated Efflux Modulators of Plant Origin: A Short Review. Nat. Prod. Commun. 2016, 11, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.-L.; Kim, H.-N.; Jung, H.-H.; Kim, J.-E.; Choi, D.-K.; Hur, J.-M.; Lee, J.-Y.; Song, H.; Song, K.-S.; Huh, T.-L. Beneficial effects of beta-sitosterol on glucose and lipid metabolism in L6 myotube cells are mediated by AMP-activated protein kinase. Biochem. Biophys. Res. Commun. 2008, 377, 1253–1258. [Google Scholar] [CrossRef]
- Sun, Y.; Gao, L.; Hou, W.; Wu, J. β-Sitosterol Alleviates Inflammatory Response via Inhibiting the Activation of ERK/p38 and NF-κB Pathways in LPS-Exposed BV2 Cells. Biomed Res. Int. 2020, 2020, 7532306. [Google Scholar] [CrossRef] [PubMed]
- Nair, V.D.P.; Foster, B.C.; Thor Arnason, J.; Mills, E.J.; Kanfer, I. In vitro evaluation of human cytochrome P450 and P-glycoprotein-mediated metabolism of some phytochemicals in extracts and formulations of African potato. Phytomedicine 2007, 14, 498–507. [Google Scholar] [CrossRef]
- Rosenblat, M.; Volkova, N.; Aviram, M. Pomegranate phytosterol (β-sitosterol) and polyphenolic antioxidant (punicalagin) addition to statin, significantly protected against macrophage foam cells formation. Atherosclerosis 2013, 226, 110–117. [Google Scholar] [CrossRef]
- Wang, Z.; Zhan, Y.; Xu, J.; Wang, Y.; Sun, M.; Chen, J.; Liang, T.; Wu, L.; Xu, K. β-Sitosterol Reverses Multidrug Resistance via BCRP Suppression by Inhibiting the p53–MDM2 Interaction in Colorectal Cancer. J. Agric. Food Chem. 2020, 68, 3850–3858. [Google Scholar] [CrossRef]
- El-Readi, M.Z.; Hamdan, D.; Farrag, N.; El-Shazly, A.; Wink, M. Inhibition of P-glycoprotein activity by limonin and other secondary metabolites from Citrus species in human colon and leukaemia cell lines. Eur. J. Pharmacol. 2010, 626, 139–145. [Google Scholar] [CrossRef]
- Beppe, G.J.; Dongmo, A.B.; Foyet, H.S.; Tsabang, N.; Olteanu, Z.; Cioanca, O.; Hancianu, M.; Dimo, T.; Hritcu, L. Memory-enhancing activities of the aqueous extract of Albizia adianthifolia leaves in the 6-hydroxydopamine-lesion rodent model of Parkinson’s disease. BMC Complement. Altern. Med. 2014, 14, 1–11. [Google Scholar] [CrossRef]
- Nakamura, K.; Deyama, Y.; Yoshimura, Y.; Hashimoto, M.; Kaga, M.; Suzuki, K.; Yawaka, Y. Tannin-fluoride preparation attenuates prostaglandin E2 production by dental pulp cells. Mol. Med. Rep. 2011, 4, 641–644. [Google Scholar] [CrossRef][Green Version]
- Yao, H.-T.; Chang, Y.-W.; Lan, S.-J.; Yeh, T.-K. The inhibitory effect of tannic acid on cytochrome P450 enzymes and NADPH-CYP reductase in rat and human liver microsomes. Food Chem. Toxicol. 2008, 46, 645–653. [Google Scholar] [CrossRef]
- Karakurt, S.; Adali, O. Tannic Acid Inhibits Proliferation, Migration, Invasion of Prostate Cancer and Modulates Drug Metabolizing and Antioxidant Enzymes. Anticancer Agents Med. Chem. 2016, 16, 781–789. [Google Scholar] [CrossRef]
- Baer-Dubowska, W.; Szaefer, H.; Majchrzak-Celińska, A.; Krajka-Kuźniak, V. Tannic Acid: Specific Form of Tannins in Cancer Chemoprevention and Therapy-Old and New Applications. Curr. Pharmacol. Rep. 2020, 6, 28–37. [Google Scholar] [CrossRef]
- Liao, X.; Gao, Y.; Liu, J.; Tao, L.; Xie, J.; Gu, Y.; Liu, T.; Wang, D.; Xie, D.; Mo, S. Combination of Tanshinone IIA and Cisplatin Inhibits Esophageal Cancer by Downregulating NF-κB/COX-2/VEGF Pathway. Front. Oncol. 2020, 10, 1756. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-X.; Cao, Y.-F.; Wang, L.-X.; Yuan, X.-L.; Fang, Z.-Z. Inhibitory effects of tanshinones towards the catalytic activity of UDP-glucuronosyltransferases (UGTs). Pharm. Biol. 2017, 55, 1703–1709. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Li, L.; Tian, D.-D. Salvia miltiorrhiza roots against cardiovascular disease: Consideration of Herb-Drug Interactions. Biomed. Res. Int. 2017, 2017, 9868694. [Google Scholar] [CrossRef]
- Jing, J.; Zheng, H.; Wang, J.; Lin, P.; Zhang, J.; Xiong, Z.-J.; Wu, Y.-Y.; Ren, J.-J.; Yang, H.-L.; Wang, X.-J. Growth inhibition and multidrug resistance-reversing effect of tanshinone I A on human breast cancer cell with estrogen receptor negative. Sichuan Da Xue Xue Bao Yi Xue Ban 2007, 38, 391–395. [Google Scholar]
- Shu, Q. Is Brain-derived Neurotrophic Factor a Possible Mechanism Underlying Risperidone Sensitization in Adolescent Rats? Biochem. Pharmacol. 2013, S1, 4. [Google Scholar] [CrossRef]
- Chen, S.; Jia, J. Tenuifolin Attenuates Amyloid-β42-Induced Neuroinflammation in Microglia Through the NF-κB Signaling Pathway. J. Alzheimers. Dis. 2020, 76, 195–205. [Google Scholar] [CrossRef]
- Gates, S.; Miners, J.O. Cytochrome P450 isoform selectivity in human hepatic theobromine metabolism. Br. J. Clin. Pharmacol. 1999, 47, 299–305. [Google Scholar] [CrossRef]
- Lin, Y.; Zhao, W.R.; Shi, W.T.; Zhang, J.; Zhang, K.Y.; Ding, Q.; Chen, X.L.; Tang, J.Y.; Zhou, Z.Y. Pharmacological Activity, Pharmacokinetics, and Toxicity of Timosaponin AIII, a Natural Product Isolated From Anemarrhena asphodeloides Bunge: A Review. Front. Pharmacol. 2020, 11, 764. [Google Scholar] [CrossRef]
- Kanaga Sabapathi, S.; Swamy, V. Evaluation of cytochrome P450 inhibition properties of trigonelline hydrochloride by using Tandem Mass Spectrometry. World J. Pharm. Pharm. Sci. 2015, 4, 772–782. [Google Scholar]
- Isshiki, M.; Umezawa, K.; Tamura, H. Coffee induces breast cancer resistance protein expression in Caco-2 cells. Biol. Pharm. Bull. 2011, 34, 1624–1627. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Picking, D.; Chambers, B.; Barker, J.; Shah, I.; Porter, R.; Naughton, D.P.; Delgoda, R. Inhibition of Cytochrome P450 Activities by Extracts of Hyptis verticillata Jacq.: Assessment for Potential HERB-Drug Interactions. Molecules 2018, 23, 430. [Google Scholar] [CrossRef] [PubMed]
- Jinhua, W.; Ying, Z.; Yuhua, L. PXR-ABC drug transporters/CYP-mediated ursolic acid transport and metabolism in vitro and vivo. Arch. Pharm. (Weinheim) 2020, 353, e2000082. [Google Scholar] [CrossRef] [PubMed]
- Singla, R.K.; Scotti, L.; Dubey, A.K. In Silico Studies Revealed Multiple Neurological Targets for the Antidepressant Molecule Ursolic Acid. Curr. Neuropharmacol. 2017, 15, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Nabekura, T.; Yamaki, T.; Hiroi, T.; Ueno, K.; Kitagawa, S. Inhibition of anticancer drug efflux transporter P-glycoprotein by rosemary phytochemicals. Pharmacol. Res. 2010, 61, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Hua, W.J.; Hua, W.X.; Nan, F.Y.; Jiang, W.A.; Yan, C. The influence of herbal medicine ursolic acid on the uptake of rosuvastatin mediated by OATP1B1*1a and *5. Eur. J. Drug Metab. Pharmacokinet. 2014, 39, 221–230. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Uesawa, Y.; Takeuchi, T.; Mohri, K. Integrated analysis on the physicochemical properties of dihydropyridine calcium channel blockers in grapefruit juice interactions. Curr. Pharm. Biotechnol. 2012, 13, 1705–1717. [Google Scholar] [CrossRef]
- Savai, J.; Pandita, N.; Chintamaneni, M. Investigation of CYP1A interaction potential of Withania somnifera in rat and human liver microsomes. Indian J. Pharm. Sci. 2014, 76, 138–147. [Google Scholar]
- Wu, Z.; Uchi, H.; Morino-Koga, S.; Nakamura-Satomura, A.; Kita, K.; Shi, W.; Furue, M. Z-Ligustilide inhibits benzo(a)pyrene-induced CYP1A1 upregulation in cultured human keratinocytes via ROS-dependent Nrf2 activation. Exp. Dermatol. 2014, 23, 260–265. [Google Scholar] [CrossRef]
- Gonçalves, B.M.F.; Cardoso, D.S.P.; Ferreira, M.-J.U. Overcoming Multidrug Resistance: Flavonoid and Terpenoid Nitrogen-Containing Derivatives as ABC Transporter Modulators. Molecules 2020, 25, 3364. [Google Scholar] [CrossRef]
- Misaka, S.; Abe, O.; Sato, H.; Ono, T.; Shikama, Y.; Onoue, S.; Yabe, H.; Kimura, J. Lack of pharmacokinetic interaction between fluvastatin and green tea in healthy volunteers. Eur. J. Clin. Pharmacol. 2018, 74, 601–609. [Google Scholar] [CrossRef]
- Li, M.; Lan, J.; Li, X.; Xin, M.; Wang, H.; Zhang, F.; Lu, X.; Zhuang, Z.; Wu, X. Novel ultra-small micelles based on ginsenoside Rb1: A potential nanoplatform for ocular drug delivery. Drug Deliv. 2019, 26, 481–489. [Google Scholar] [CrossRef] [PubMed]
- González-Lugo, O.E.; Pozos-Guillén, A.; Ponce-Peña, P.; Lares-Asseff, I.; Escobar-García, D.M.; Campos-Cantón, I.; Vértiz-Hernández, A.A. Synergistic interaction between 4-allyl-1-hydroxy-2-methoxybenzene (eugenol) and diclofenac: An isobolograpic analysis in Wistar rats. Drug Dev. Res. 2020, 81, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Matejczyk, M.; Ofman, P.; Dąbrowska, K.; Świsłocka, R.; Lewandowski, W. The study of biological activity of transformation products of diclofenac and its interaction with chlorogenic acid. J. Environ. Sci. 2020, 91, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Bedada, S.K.; Yellu, N.R.; Neerati, P. Effect of Resveratrol Treatment on the Pharmacokinetics of Diclofenac in Healthy Human Volunteers. Phytother. Res. 2016, 30, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Buckley, D.; Klaassen, C.D. Regulation of hepatic bile acid transporters Ntcp and Bsep expression. Biochem. Pharmacol. 2007, 74, 1665–1676. [Google Scholar] [CrossRef] [PubMed]
- Dawson, S.; Stahl, S.; Paul, N.; Barber, J.; Kenna, J.G. In vitro inhibition of the bile salt export pump correlates with risk of cholestatic drug-induced liver injury in humans. Drug Metab. Dispos. 2012, 40, 130–138. [Google Scholar] [CrossRef]
- DeBose-Boyd, R.A. Feedback regulation of cholesterol synthesis: Sterol-accelerated ubiquitination and degradation of HMG CoA reductase. Cell Res. 2008, 18, 609–621. [Google Scholar] [CrossRef]
- Sama, V.; Pagilla, B.; Chiluka, R.; Alvala, R.; Pola, R.K.; Mullangi, R. Bioenhancing effects of naringin on atorvastatin. ADMET DMPK 2019, 7, 174–182. [Google Scholar] [CrossRef]
- Kong, W.; Wei, J.; Abidi, P.; Lin, M.; Inaba, S.; Li, C.; Wang, Y.; Wang, Z.; Si, S.; Pan, H.; et al. Berberine is a novel cholesterol-lowering drug working through a unique mechanism distinct from statins. Nat. Med. 2004, 10, 1344–1351. [Google Scholar] [CrossRef]
- Feng, P.; Zhao, L.; Guo, F.; Zhang, B.; Fang, L.; Zhan, G.; Xu, X.; Fang, Q.; Liang, Z.; Li, B. The enhancement of cardiotoxicity that results from inhibitIon of CYP 3A4 activity and hERG channel by berberine in combination with statins. Chem. Biol. Interact. 2018, 293, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xiao, H.; Lin, C.; Sun, W.; Wu, T.; Wang, J.; Chen, B.; Chen, X.; Cheng, D. Synergistic effects of liposomes encapsulating atorvastatin calcium and curcumin and targeting dysfunctional endothelial cells in reducing atherosclerosis. Int. J. Nanomed. 2019, 14, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Bo, L.; Baosheng, Z.; Yang, L.; Mingmin, T.; Beiran, L.; Zhiqiang, L.; Huaqiang, Z. Herb-drug enzyme-mediated interactions and the associated experimental methods: A review. J. Tradit. Chin. Med. 2016, 36, 392–408. [Google Scholar] [CrossRef]
- NEUROTHERATM. Available online: https://klaire.com/nro-neurothera (accessed on 22 December 2020).
- Maxgars Memory Booster. Available online: https://www.maxgars.com/product/maxgars-memory-booster-60-capsules/ (accessed on 22 December 2020).
- Palpu, P.; Rao, C.V.; Kishore, K.; Gupta, Y.K.; Kartik, R.; Govindrajan, R. Herbal Formulation as Memory Enhancer in Alzheimer Condition. U.S. Patent 7429397, 30 September 2008. [Google Scholar]
- Shaw, D.; Graeme, L.; Pierre, D.; Elizabeth, W.; Kelvin, C. Pharmacovigilance of herbal medicine. J. Ethnopharmacol. 2012, 140, 513–518. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
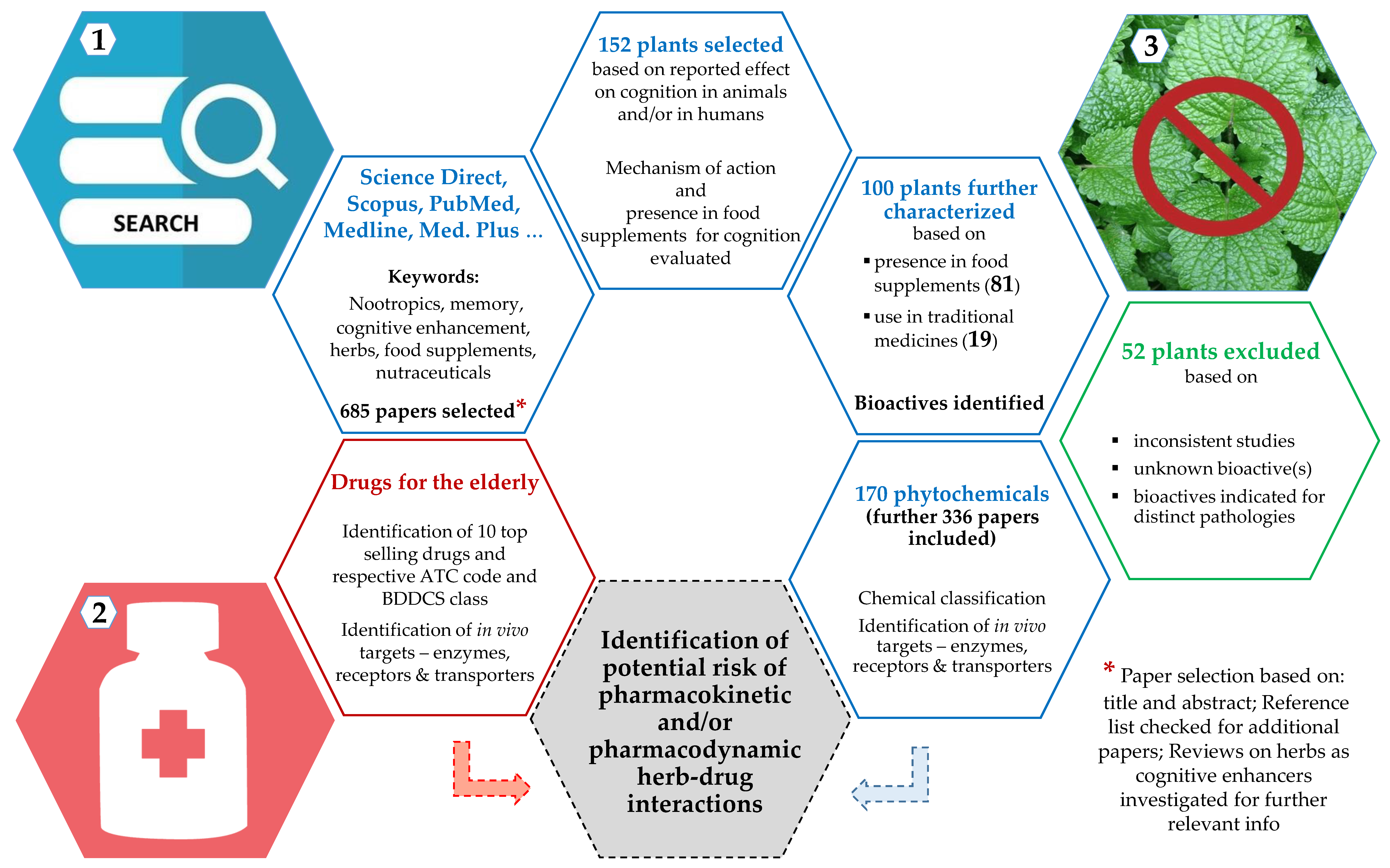{kind=link}
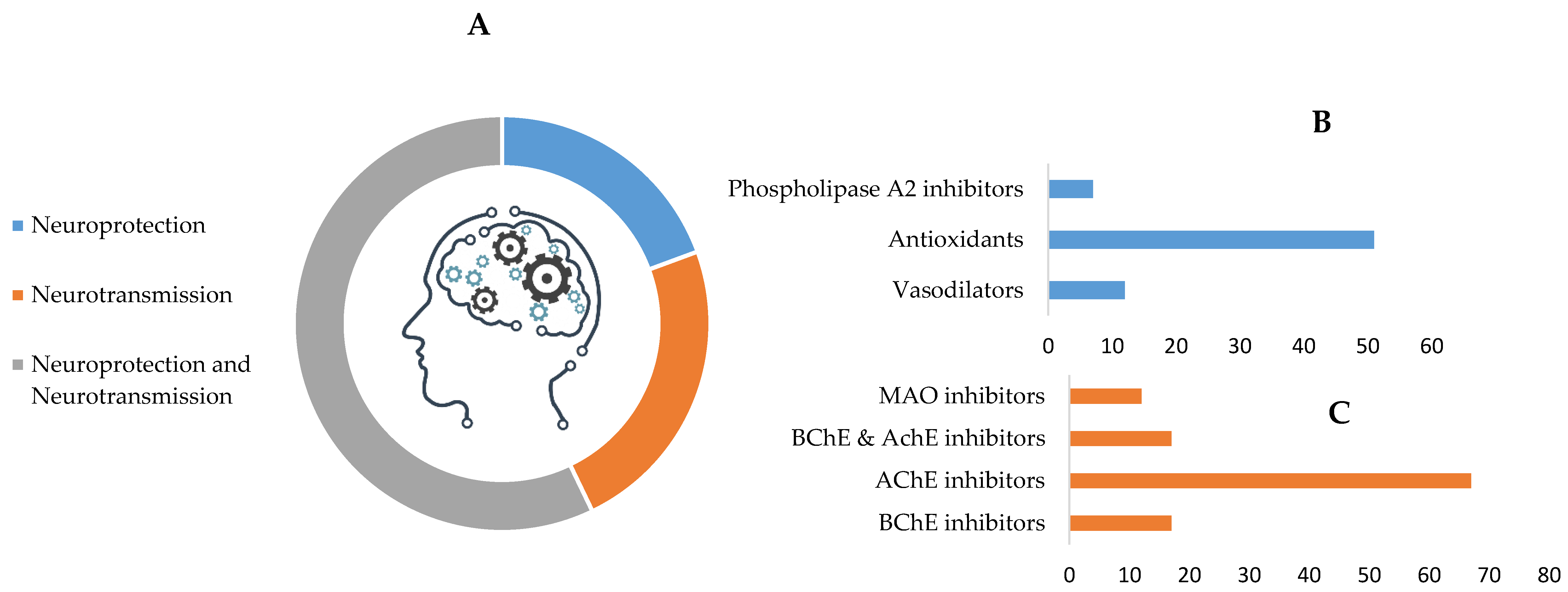{kind=link}
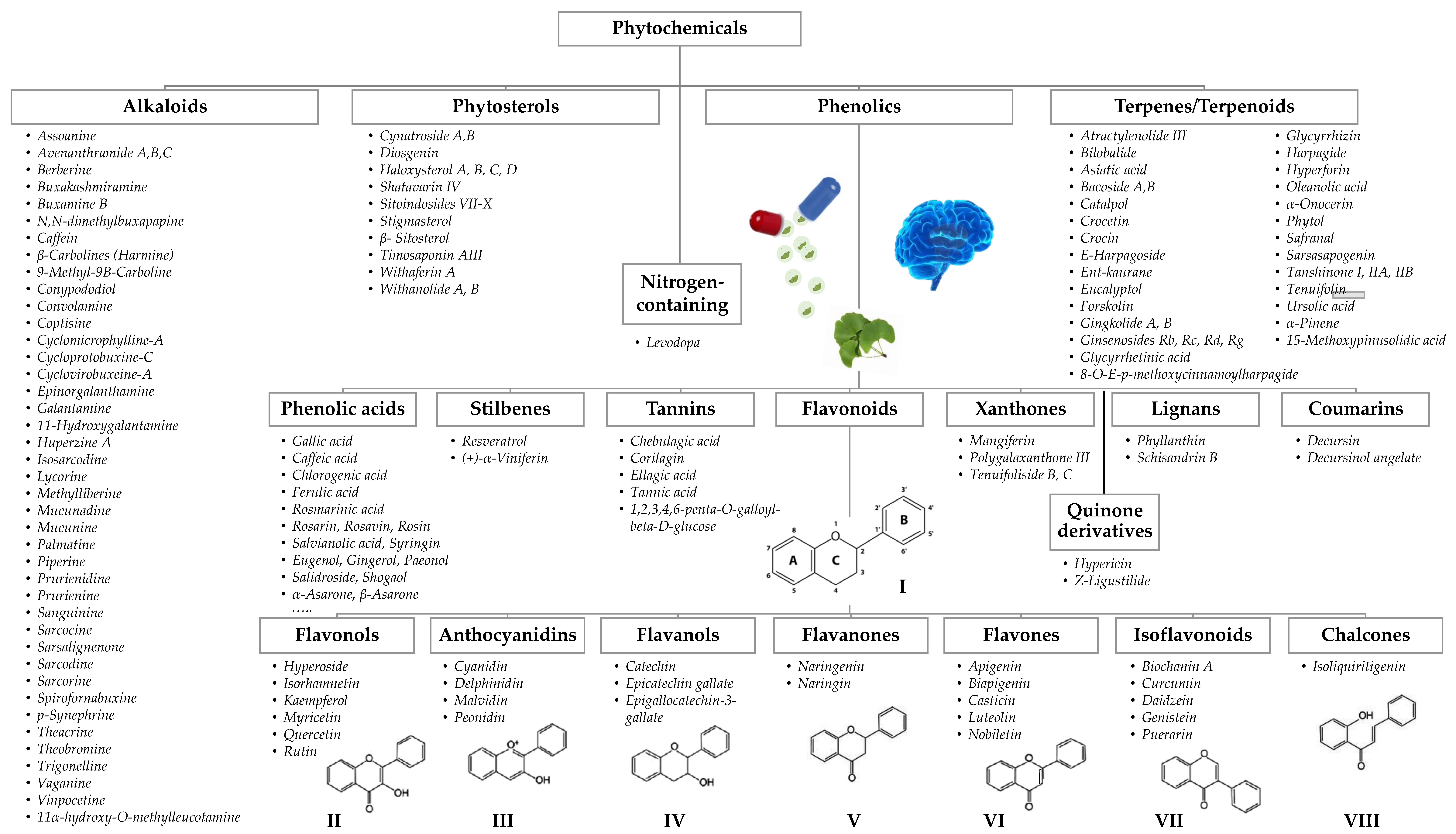{kind=link}
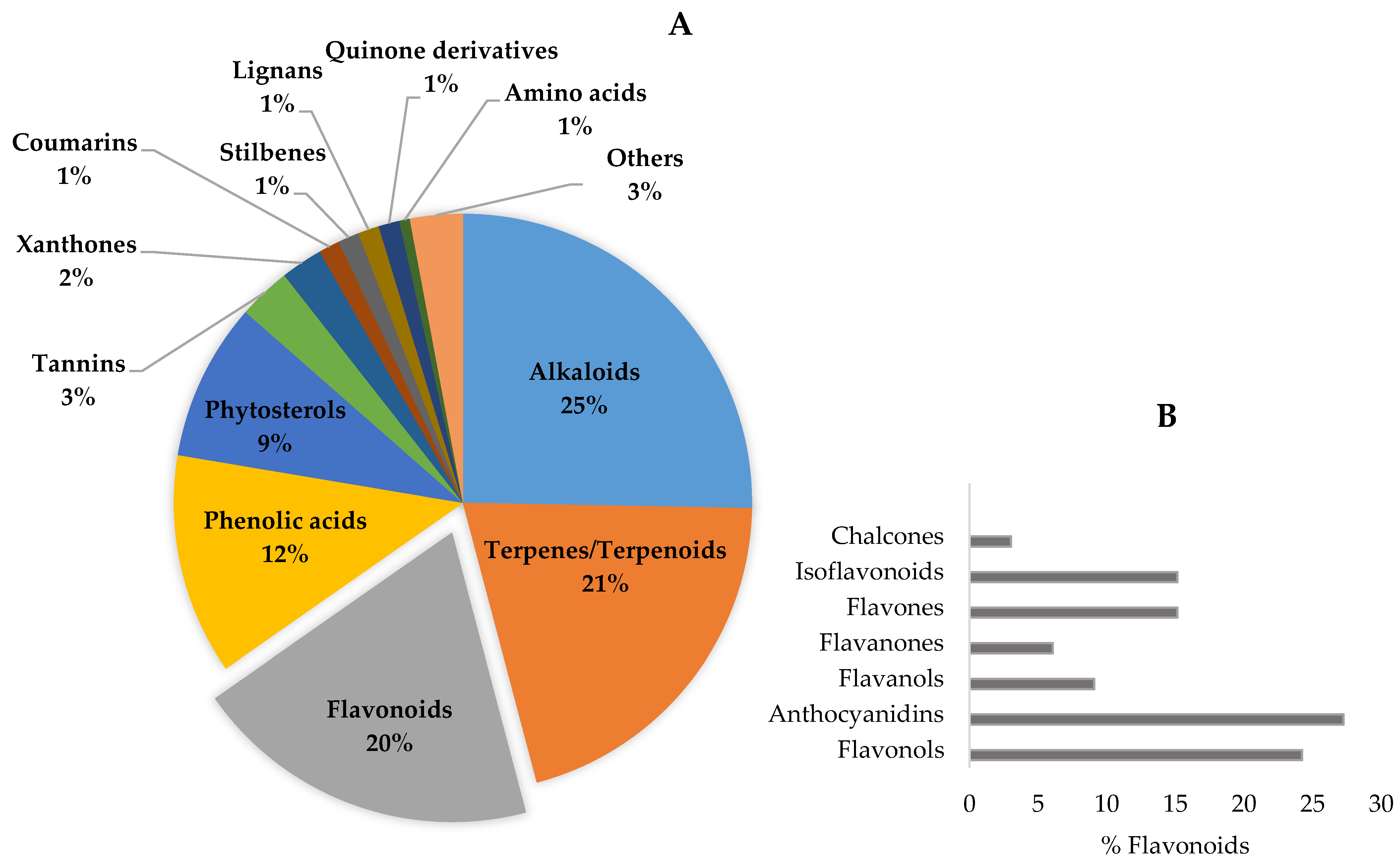{kind=link}
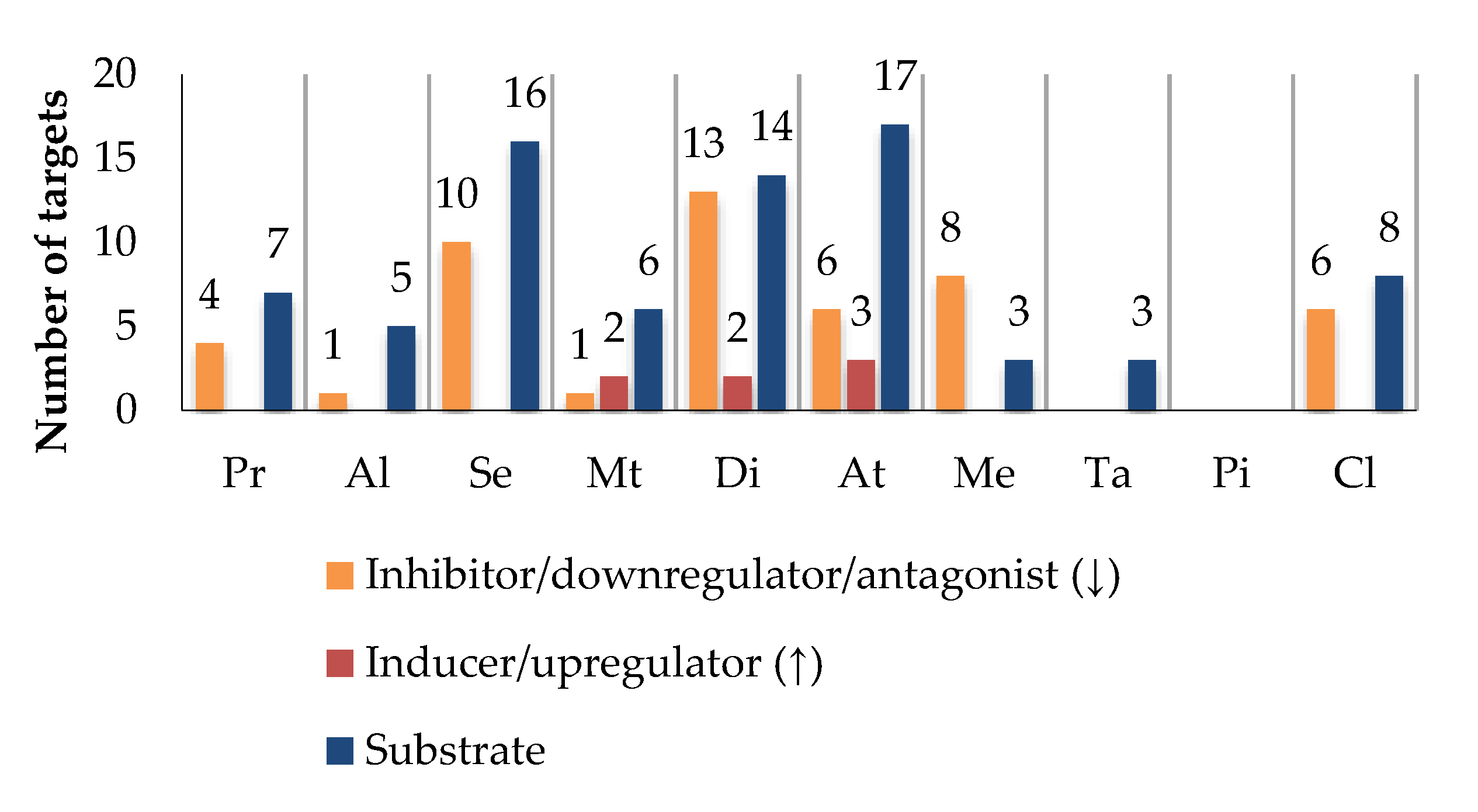{kind=link}
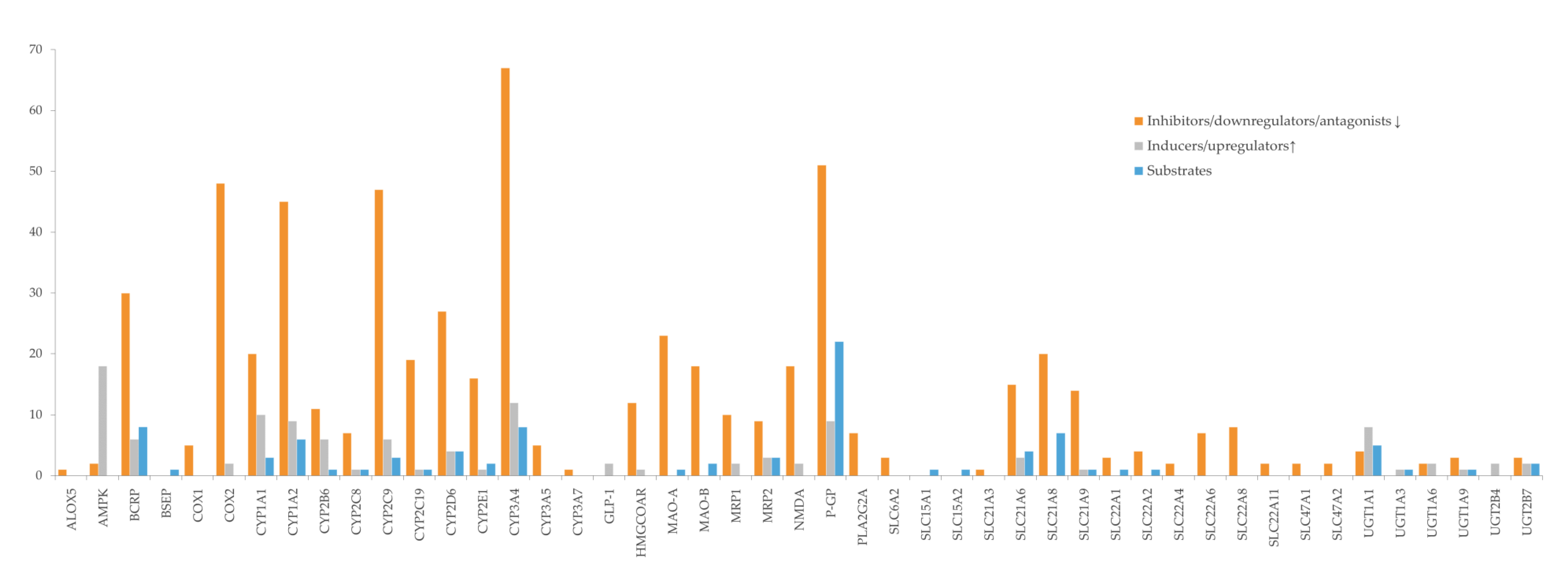{kind=link}
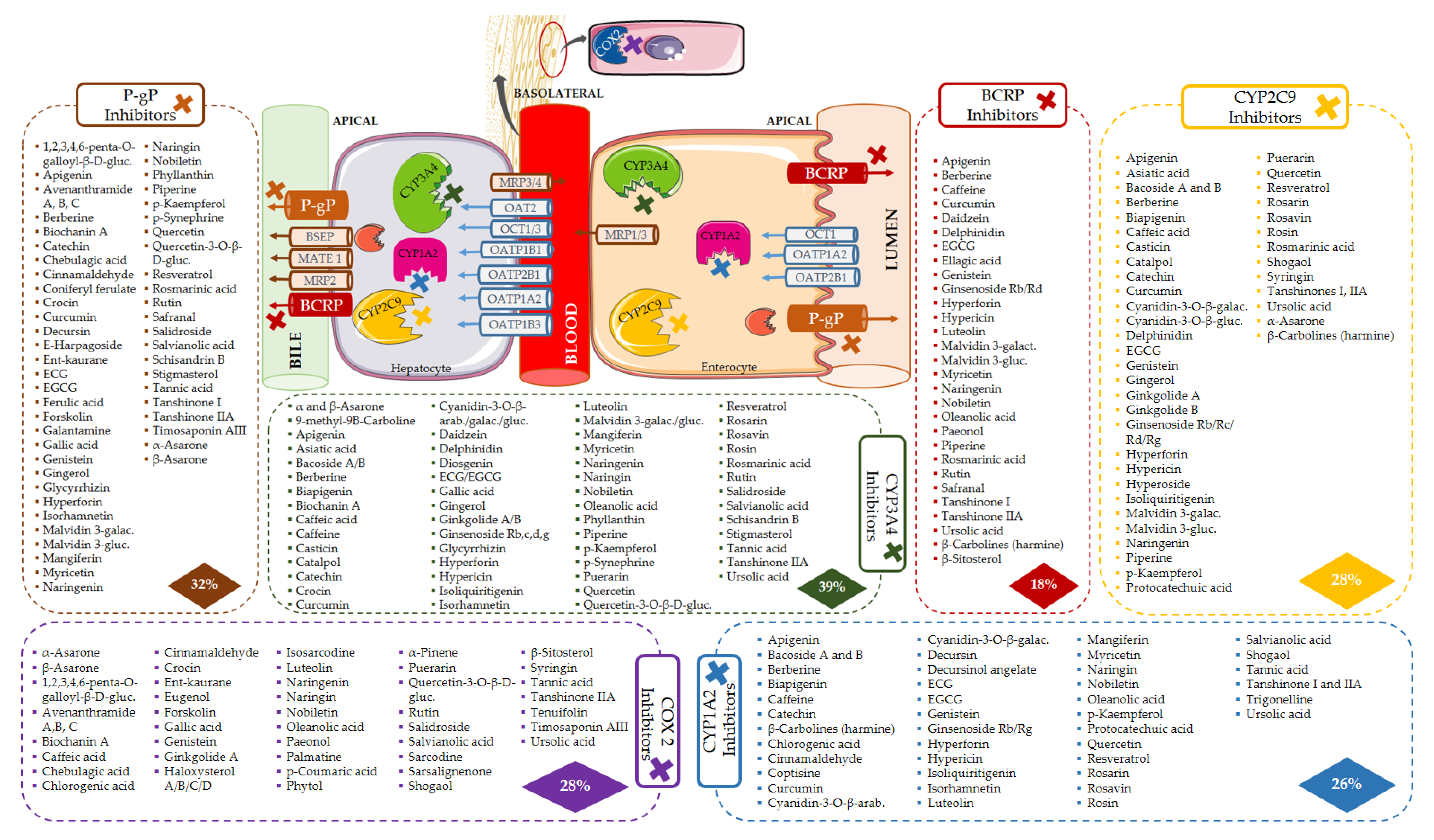{kind=link}
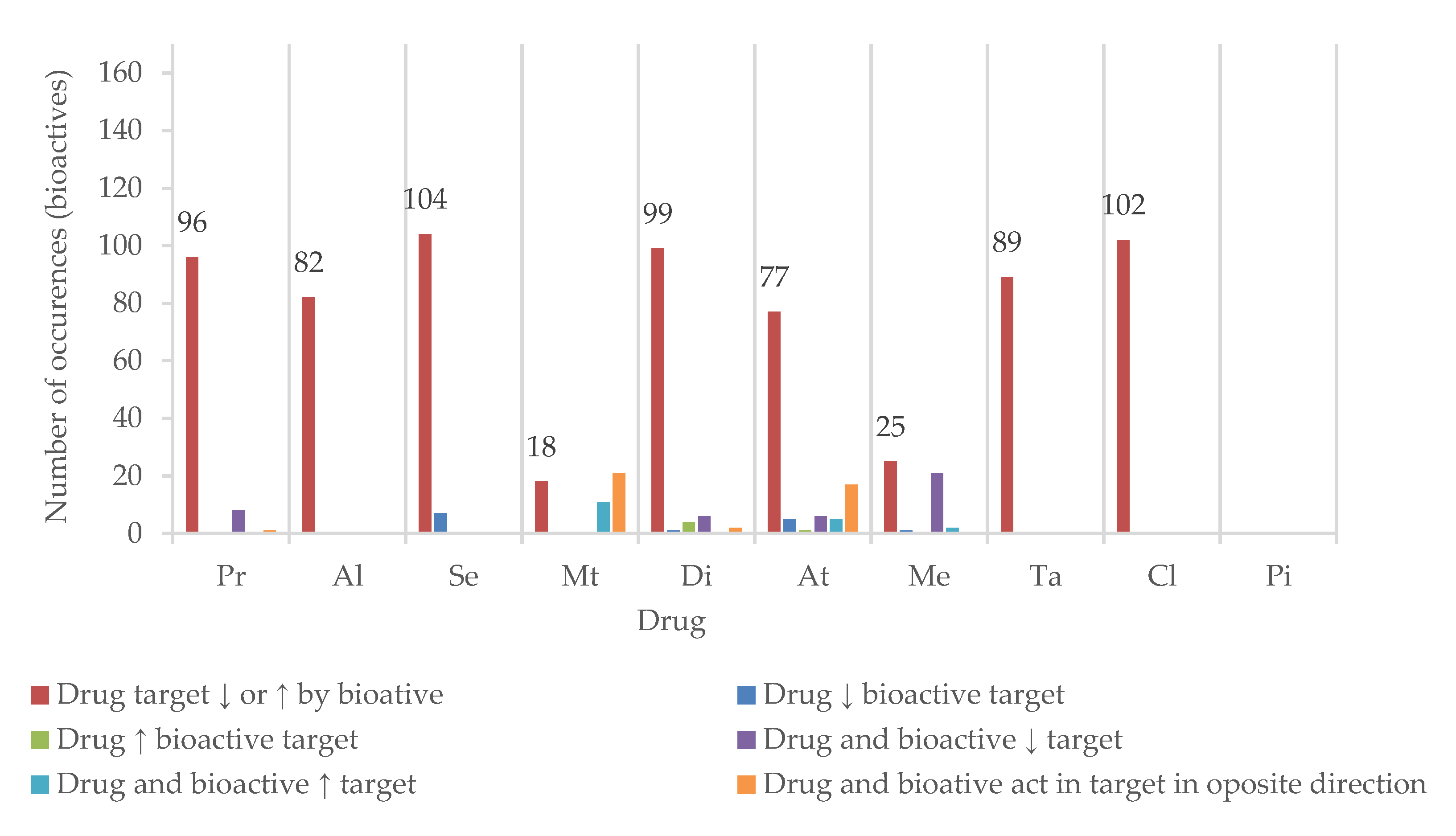{kind=link}
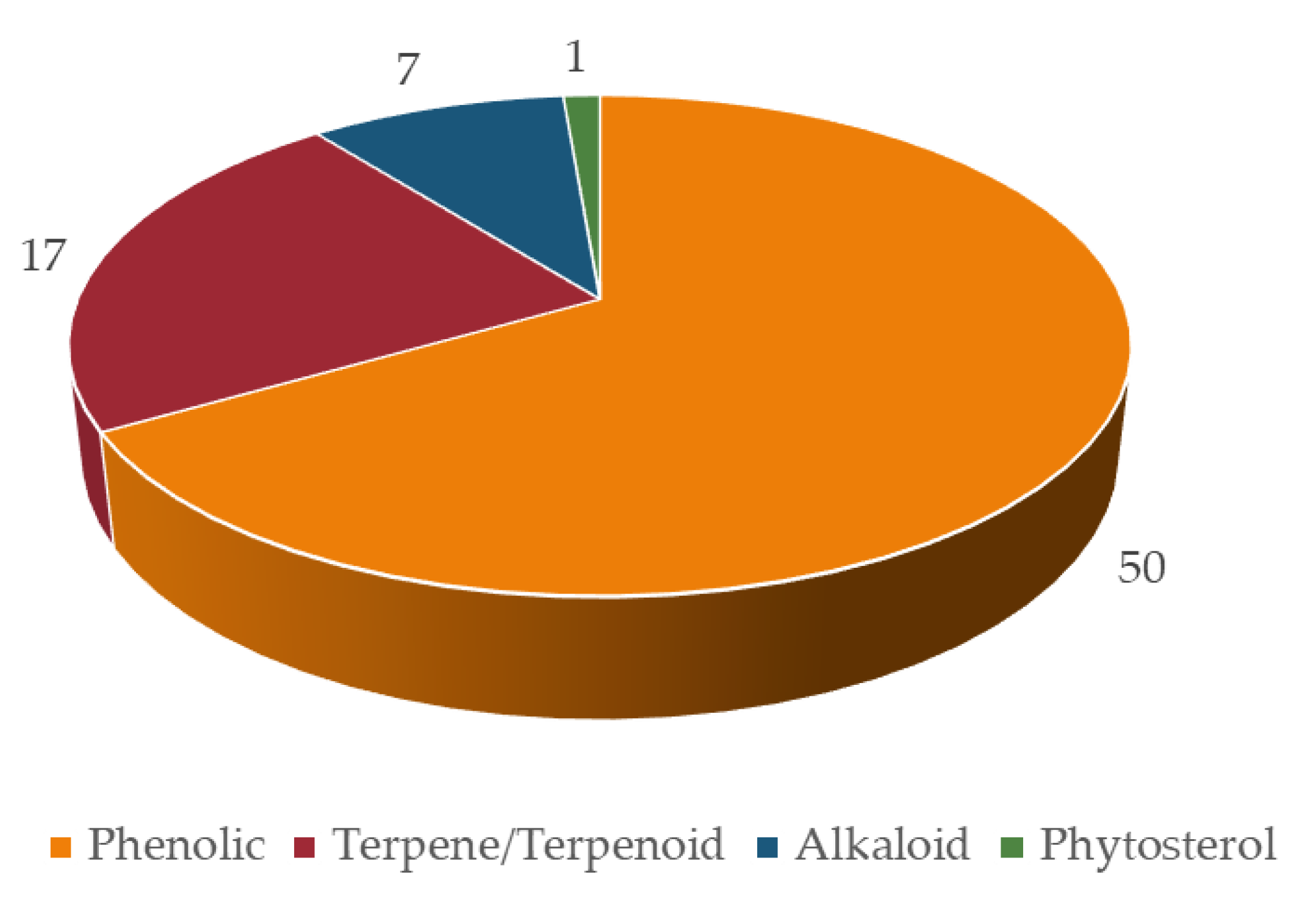{kind=link}
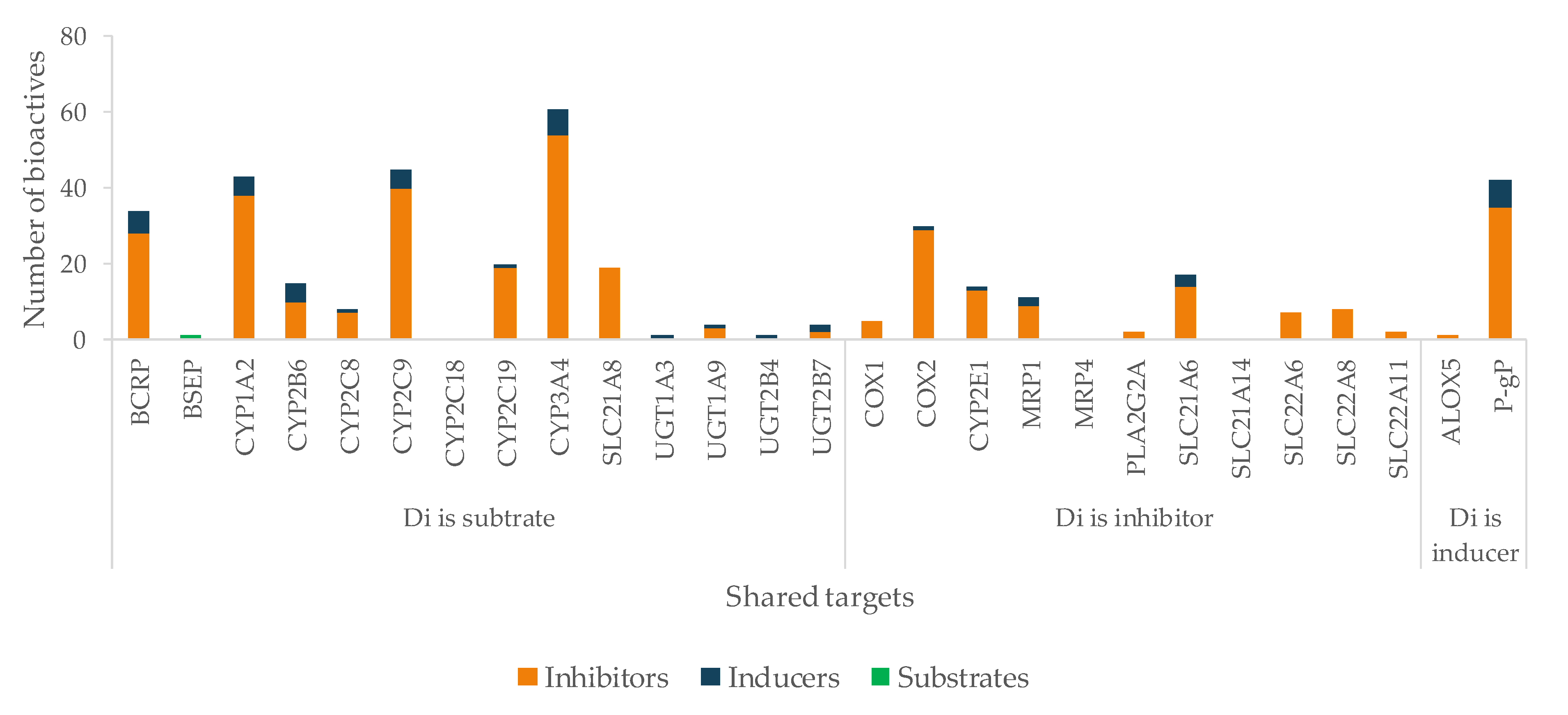{kind=link}
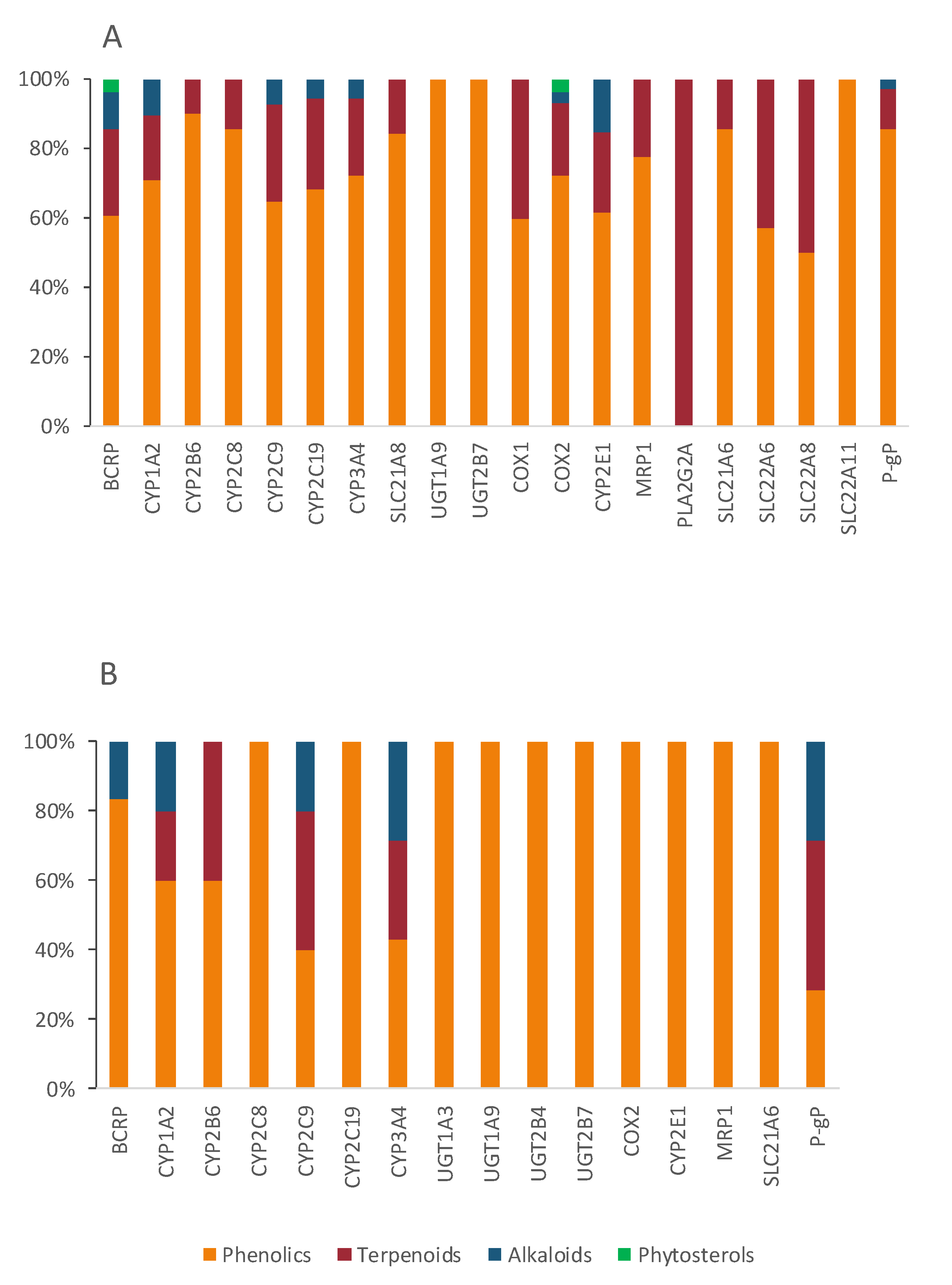{kind=link}
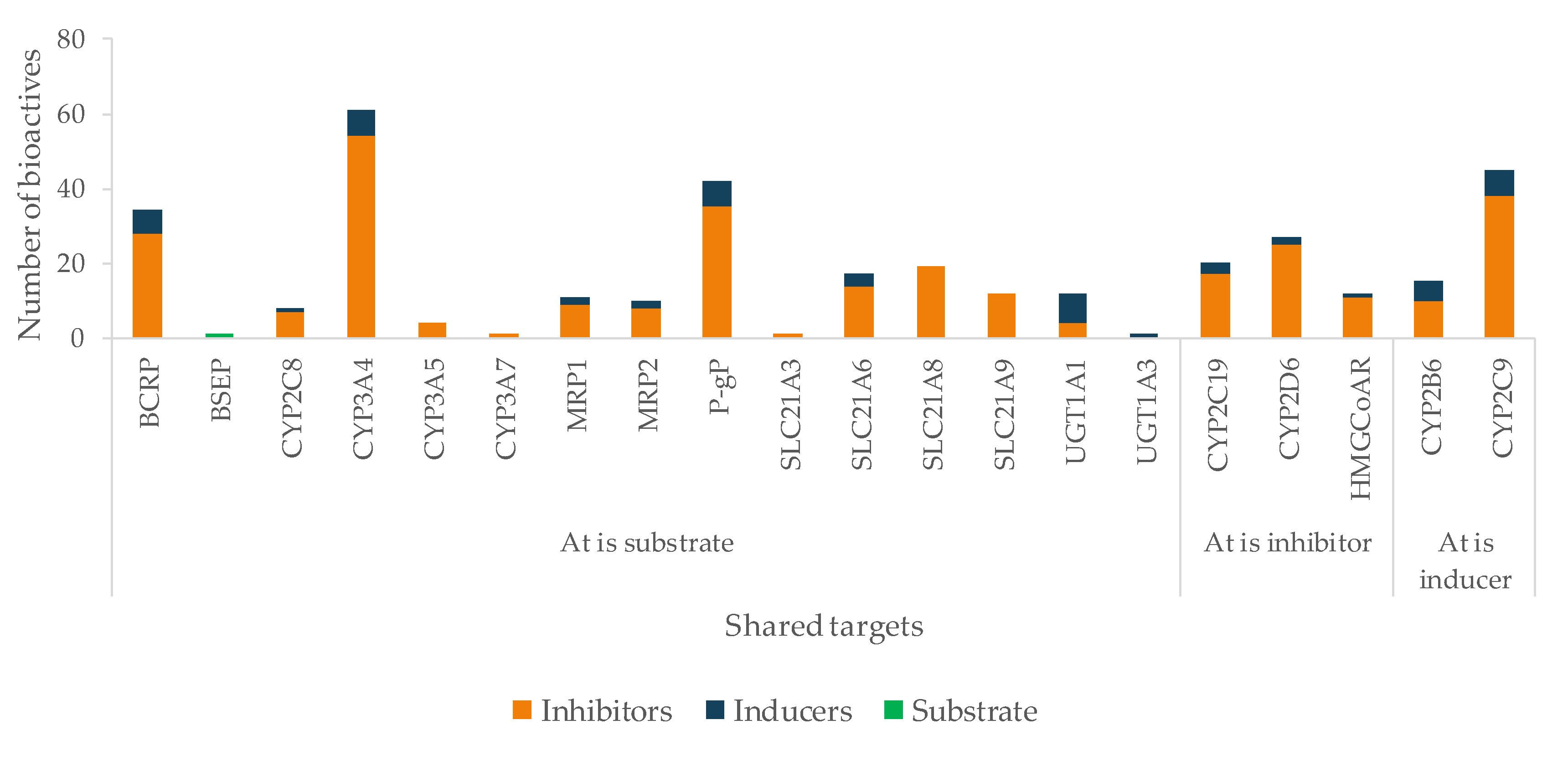{kind=link}
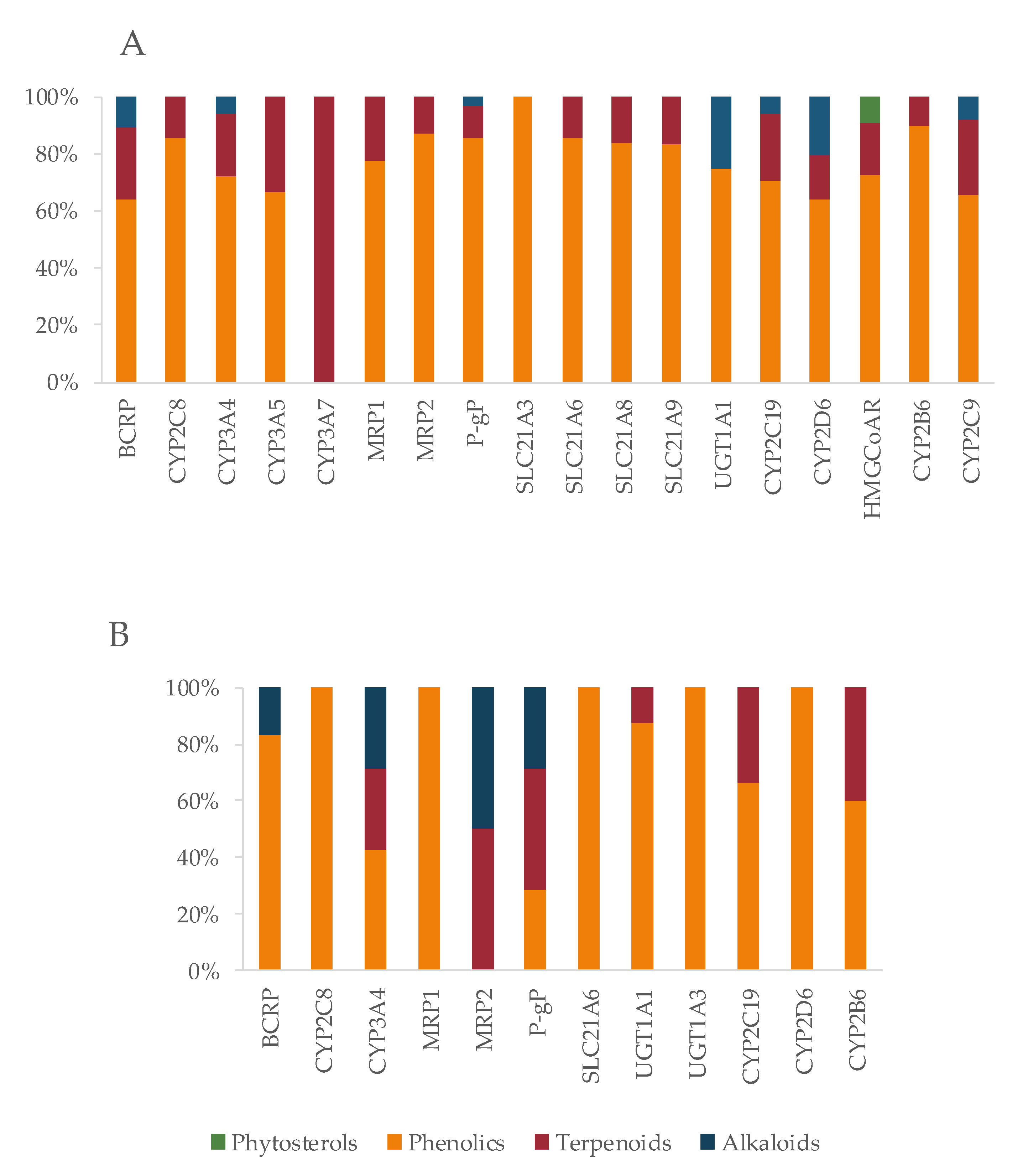{kind=link}
| Plant | Main Bioactive(S) for Cognition | Effect on Brain/Cognition | References |
|---|---|---|---|
| Acorus calamus+⬪ | α-Asarone β-Asarone | Neurotransmission | [55,56,57] |
| Acorus gramineus | α-Asarone β-Asarone | Neuroprotection and Neurotransmission | [58] |
| Aframomum melegueta | Gingerol Shogaol Quercetin p-Kaempferol | Neuroprotection and Neurotransmission | [59,60] |
| Alpinia oxyphylla■⬪ | Protocatechuic acid | Neuroprotection and Neurotransmission | [61] |
| Anemarrhena asphodeloides●■⬪ | Timosaponin AIII | Neuroprotection and Neurotransmission | [58] |
| Angelica gigas○ | Decursin Decursinol angelate | Neuroprotection and Neurotransmission | [58] |
| Angelica sinensis●▲⬪ | Z-ligustilide Coniferyl ferulate 11-Angeloylsenkyunolide F Ferulic acid | Neuroprotection and Neurotransmission | [18,62,63] |
| Asparagus adscendens | Shatavarin IV (Asparinin B) Conypododiol | Neuroprotection and Neurotransmission | [64,65] |
| Asparagus racemosus+ | Shatavarin IV (Asparinin B) Sarsasapogenin | Neuroprotection and Neurotransmission | [57,66] |
| Atractylodes japonica■ | Atractylenolide III | Neuroprotection | [67] |
| Atractylodes lancea●■⬪ | Atractylenolide III (Codonolactone) Stigmasterolβ-Sitosterol | Neurotransmission | [58] |
| Avena sativa | Avenanthramides A, B and C | Neuroprotection | [58] |
| Bacopa monnieri∗+ | Bacoside A and B | Neuroprotection and Neurotransmission | [51,68,69,70] |
| Bauhinia rufescens | d-pinitol (3-O-methyl-d-inositol) p-Coumaric acid Ferulic acid Hyperoside | Neuroprotection and Neurotransmission | [71] |
| Buxus hyrcana | Buxamine B | Neurotransmission | [22,72] |
| Buxus papillosa | Buxakashmiramine Cycloprotobuxine-C Cyclovirobuxeine-A Cyclomicrophylline-A N,N-dimethyl buxapapine | Neurotransmission | [22,72] |
| Camellia sinensis●∗⬪ | Caffein Epigallocatechin-3-gallate Epicatechin gallate Methylliberine | Neuroprotection and Neurotransmission | [55,57,73,74] |
| Caragana chamiague+ | (+)-α-Viniferin | Neurotransmission | [22] |
| Centella asiatica●∗▲+⬪ | Asiatic acid | Neuroprotection and Neurotransmission | [55,57,66] |
| Cinnamomum wilsonii (extract) | Cinnamaldehyde Eugenol | Neuroprotection and Neurotransmission | [75,76,77,78] |
| Citrus aurantium●■+⬪ | p-Synephrine | Neurotransmission | [79] |
| Citrus reticulata●■⬪○ | Nobiletin | Neuroprotection and Neurotransmission | [58] |
| Clitoria ternatea+ | Kaempferol | Neuroprotection and Neurotransmission | [57,66] |
| Coffee arabica | Caffein Methylliberine Theacrine | Neuroprotection and Neurotransmission | [55,74,80,81] |
| Coleus forskohlii+ | Forskolin (Colforsin) | Neuroprotection and Neurotransmission | [82] |
| Convolvulus pluricaulis+ | Kaempferol Kaempferol-3-glucoside Caffeic acid Convolamine Β-Sitosterol | Neuroprotection | [83] |
| Coptis chinensis●■▲⬪ | Berberine Coptisine Palmatine | Neurotransmission | [57] |
| Coptis japonica■▲ | Berberine | Neuroprotection | [57] |
| Corydalis speciosa | Palmatine | Neurotransmission | [22] |
| Crocus sativus●■▲+⬪ | Quercetin Crocins/picrocrocin Crocetin Safranal | Neuroprotection and Neurotransmission | [55,57,66] |
| Croton tonkinensis | Ent-kaurane | Neuroprotection and Neurotransmission | [16,84] |
| Curcuma longa●∗■▲+⬪ | Curcumin | Neuroprotection and Neurotransmission | [55,57] |
| Cuscuta japonica | Hyperoside Kaempferol | Neuroprotection and Neurotransmission | [85] |
| Cynanchum atratum⬪ | Cynatroside A Cynatroside B | Neurotransmission | [22] |
| Cyperus rotundus■+⬪ | Quercetin Kaempferol Catechin | Neuroprotection and Neurotransmission | [85] |
| Dioscorea polystachya■○ | Diosgenin | Neuroprotection | [58] |
| Dioscorea oppositifolia●■⬪ | Diosgenin | Neuroprotection | [86,87] |
| Echium amoenum | Cyanidin-3-glucoside | Neuroprotection | [88,89] |
| Eleutherococcus senticosus●∗■▲ (Siberian ginseng) | Syringin (Eleutheroside B) | Neuroprotection and Neurotransmission | [90,91,92,93] |
| Eucharis grandiflora | Sanguinine (O-Desmethylgalantamine) | Neuroprotection and Neurotransmission | [22] |
| Foeniculum vulgare●∗■▲+⬪ | Caffeic acid Chlorogenic acid 1,8-Cineole | Neuroprotection and Neurotransmission | [52,94,95,96] |
| Galanthus nivalis | Galantamine | Neurotransmission | [22,57] |
| Galanthus woronowii (or ikariae) | Galantamine Lycorine | Neurotransmission | [20,22,55] |
| Ginkgo biloba●∗▲⬪ | Ginkgolide A Ginkgolide B Bilobalide Isorhamnetin Protocatechuic acid | Neuroprotection and Neurotransmission | [20,55,57,81,97] |
| Glycyrrhiza glabra●∗■▲+⬪ | Glycyrrhizin Glycyrrhetinic acid | Neuroprotection | [57] |
| Glycyrrhiza uralensis●■▲⬪ | Isoliquiritigenin | Neuroprotection | [58] |
| Haloxylon recurvum | Haloxysterols A, B, C, D | Neurotransmission | [22] |
| Huperzia serrata | Huperzine A | Neuroprotection and Neurotransmission | [8,20,55,57] |
| Hypericum perforatum●∗▲⬪ | Hypericin Hyperforin Biapigenin (I3,II8-biapigenin) Quercetin Chlorogenic acid Rutin Hyperoside Kaempferol | Neuroprotection and Neurotransmission | [98,99,100,101,102] |
| Ilex paraguariensis● | Chlorogenic acid Caffein Theobromine Quercetin Kaempferol | Neurotransmission | [22,57] |
| Lepidium meyenii○ (Maca Root Extract) | Quercetin β-Carbolines N-Benzylhexadecanamide N-Acetylbenzylamine N-3-Methoxybenzyl-linoleamide | Neuroprotection and Neurotransmission | [57,103,104,105,106,107] |
| Lespedeza bicolor | Catechin Rutin Daidzein Luteolin Naringenin Genistein | Neuroprotection and Neurotransmission | [22] |
| Lycopodium clavatum | α-Onocerin | Neurotransmission | [22] |
| Lycoris radiata | Galantamine | Neurotransmission | [22,57] |
| Mangifera indica+○ | Mangiferin | Neuroprotection and Neurotransmission | [108] |
| Matricaria chamomilla●∗▲ | Chlorogenic acid Caffeic acid Catechin Rutin Luteolin Apigenin | Neuroprotection and Neurotransmission | [22] |
| Mauritia flexuosa | Rutin | Neuroprotection and Neurotransmission | [22,109] |
| Melissa officinalis●▲ | Luteolin Apigenin Rosmarinic acid Protocatechuic acid | Neuroprotection and Neurotransmission | [20,55,57] |
| Mentha spicata | Rosmarinic acid Salvianolic acid | Neuroprotection and Neurotransmission | [110,111] |
| Morinda lucida | Phytol Oleanolic acid Chlorogenic acid p-Coumaric acid Daidzein Rutin Naringin Quercetin Naringenin Genistein | Neuroprotection and Neurotransmission | [112,113,114,115,116,117,118,119] |
| Moringa peregrina | Rutin Myricetin β-Sitosterol | Neuroprotection and Neurotransmission | [22] |
| Mucuna pruriens+○ | Gallic acid Genistein Levodopa (l-Dopa) Mucunadine Mucunine Prurienidine Prurienine β-Carbolines (Harmine) β-Sitosterol | Neuroprotection and Neurotransmission | [12,17,22,120] |
| Narcissus assoanus | Assoanine | Neurotransmission | [22] |
| Narcissus confusus | Galantamine Epinorgalantamine | Neurotransmission | [17,22,57] |
| Narcissus poeticus | 11-Hydroxygalantamine | Neurotransmission | [22] |
| Paeonia lactiflora●■▲⬪○ | Paeonol | Neurotransmission | [22,58] |
| Panax ginseng●∗■▲⬪○ | Ginsenosides Rb, Rc, Rd, Rg1 | Neuroprotection | [8,55,57,58,81] |
| Pancratium illyricum | 11-α-hydroxy-O-methylleucotamine Galantamine Sanguinine Lycorine | Neuroprotection and Neurotransmission | [8,20,55,57,58,81,121] |
| Paullinia cupana●○ (Guarana) | Caffein | Neuroprotection and Neurotransmission | [81,122,123] |
| Peganum harmala | β-Carbolines (Harmine, 9-Methyl-9B-Carboline) | Neuroprotection and Neurotransmission | [124,125,126,127] |
| Peltophorum pterocarpum | Hyperoside Quercetin-3-O-β-d-glucuronide | Neuroprotection and Neurotransmission | [116] |
| Phyllanthus emblica+⬪ (Emblica officinalis) | Ellagic acid Gallic acid Chebulagic acid Apigenin Quercetin Corilagin Luteolin Phyllanthin | Neuroprotection and Neurotransmission | [57,128,129,130] |
| Piper nigrum●∗+⬪ | Piperine | Neuroprotection | [131,132] |
| Platycladus orientalis⬪ | 15-Methoxypinusolidic acid | Neurotransmission | [57,58,133] |
| Polygala tenuifolia●■⬪ | Tenuifoliside B Tenuifoliside C Tenuifolin Polygalaxanthone III | Neuroprotection and Neurotransmission | [134,135,136,137,138,139,140] |
| Puerariae lobate●∗■⬪ | Daidzein Puerarin | Neuroprotection | [141] |
| Rehmannia glutinosa■▲⬪○ | Catalpol | Neuroprotection | [58,142,143] |
| Rhizoma acori+⬪ | Eugenol β-Asarone | Neuroprotection | [144,145] |
| Rhodiola rosea∗○ | Rosavin Rosin Rosarin Salidroside | Neuroprotection and Neurotransmission | [146,147,148] |
| Ribes nigrum● (Blackcurrant) | Myricetin Quercetin Isorhamnetin | Neuroprotection and Neurotransmission | [22,149] |
| Rosmarinus officinalis●∗▲ | Caffeic acid Chlorogenic acid Oleanolic acid Rosmarinic acid Ursolic acid α-Pinene Eucalyptol (Cineole) Luteolin Protocatechuic acid | Neuroprotection and Neurotransmission | [22] |
| Salvia lavandulaefolia● | Eucalyptol α-Pinene | Neuroprotection and Neurotransmission | [20,22,55,57,150] |
| Salvia miltiorrhiza●∗■⬪○ | Tanshinone I Tanshinone IIA Tanshinone IIB Salvianolic acid | Neurotransmission | [24,151,152] |
| Sarcococca saligna | Sarsalignenone Vaganine Sarcocine Sarcodine Sarcorine Isosarcodine | Neurotransmission | [22] |
| Saussurea costus●■ | Apigenin Quercetin | Neurotransmission | [22,58] |
| Schisandra chinensis●■▲⬪○ | Schisandrin B (Gomisin N) | Neuroprotection and Neurotransmission | [57,58] |
| Scrophularia buergeriana | E-Harpagoside 8-O-E-p-methoxycinnamoylharpagide Harpagide | Neuroprotection and Neurotransmission | [153,154] |
| Terminalia chebula▲+⬪○ | Ellagic acid Tannic acid Chebulagic acid Corilagin Gallic acid 1,2,3,4,6-penta-O-galloyl-β-d-glucose | Neuroprotection and Neurotransmission | [22,57,66] |
| Theobroma cacao●∗■ | Theobromine Caffein Catechin | Neuroprotection and Neurotransmission | [155,156,157] |
| Theobroma grandiflorum | Theacrine | Neuroprotection | [74] |
| Trifolium pretense∗▲ (Red clover) | Biochanin A | Neuroprotection | [158] |
| Trigonella foenum-graecum●∗▲+⬪○ | Diosgenin | Neuroprotection and Neurotransmission | [159] |
| Vaccinium myrtillus●∗▲ (Bilberry) | Cyanidin-3-O-β-glucoside | Neuroprotection | [160] |
| Vaccinium uliginosum L. (Bog bilberry) | Quercetin Rutin (Quercetin-3-rutinoside) Malvidin-3-glucoside Cyanidin-3-O-β-glucoside Delphinidin | Neuroprotection and Neurotransmission | [22,160] |
| Vaccinium angustifolium (Lowbush Blueberry or Wild blueberry) | Malvidin 3-glucoside Malvidin 3-galactoside Caffeic acid Cyanidin-3-O-β-galactoside Cyanidin-3-O-β-glucoside Cyanidin-3-O-β-arabinose Malvidin-3-O-β-arabinose Peonidin-3-O-β-arabinose Delphinidin-3-O-β-galactoside | Neuroprotection and Neurotransmission | [161] |
| Vinca minor | Vinpocetine | Neuroprotection and Neurotransmission | [20,162] |
| Vitex agnus-castus●∗▲+○ (Chasteberry Extract) | Casticin | Neurotransmission | [163] |
| Vitis vinifera∗+ | Resveratrol Protocatechuic acid | Neuroprotection | [57,164] |
| Withania somnifera∗▲+ | Withaferin A Withanolide A, B Sitoindosides VII-X | Neuroprotection | [57,66,165,166,167,168,169] |
| Zingiber officinale Roscoe ●∗■▲+⬪ | Gingerol Shogaol | Neuroprotection and Neurotransmission | [22,57,170,171,172,173] |
| Drug Name and Chemical Structure | HAM | ATC Class (Code) | BDDCS Class | ALOX5 | AMPK | BSEP | COX | CYP | GLP-1 | HMGCoAR | MAO | NMDA | ABC | PLA2G2A | SLC | UGT | References | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BCRP | MRP | Pgp | PEPT; OCT; OAT, MATE, OATP | ||||||||||||||||
Alprazolam | Y | Benzodiazepine derivatives (N05BA12) | I | - | - | - | - | 3A4S↓ 3A5S 3A7S 2C9S 2C19S | - | - | - | - | - | - | - | - | - | - | [217,218,219] |
Atorvastatin | Y | HMG CoA reductase inhibitors (C10AA05) | II | - | - | S | - | 3A4S↑ 3A5S 3A7S 2B6↑ 2C8S↓ 2C9↑ 2C19↓ 2D6↓ | - | ↓ | - | - | S | 1S 2S 4S 5S | S↓ | - | 21A3S 21A6S↓ 21A8S 21A9S | 1A1S 1A3S | [218,219,220,221,222] |
Clopidogrel | Y | Platelet aggregation inhibitors excl. heparin (B01AC04) | II | - | - | - | - | 1A2S 3A4S 3A5S 2B6S↓ 2C8↓ 2C9S↓ 2C19S | - | - | - | - | ↓ | - | S | - | 21A8S 22A1↓22A2↓ | - | [218,219,223,224,225] |
Diclofenac | Y | Anti-inflammatory agents, non-steroids (S01BC03) | II | ↑ | - | S | 1↓ 2↓ | 1A2S 3A4S↓ 2B6S 2C8S 2C9S↓ 2C18S 2C19S 2E1↓ | - | - | - | - | S | 1↓ 4↓ | ↑ | ↓ | 21A6↓ 21A8S 21A14↓ 22A11↓ 22A6↓ 22A8↓ | 1A3s 1A9s 2B4s 2B7s | [218,219,226,227] |
Memantine | N | Other anti-dementia drugs (N06DX01) | III | - | - | - | - | 2B6↓ 2C19↓ | - | - | - | ANT | - | - | - | - | 15A1↓ 15A2↓ 22A8↓ 22A1↓ 22A2S↓ 22A4S 47A1S | - | [218,222,228,229,230,231] |
Metformin | Y | Biguanides (A10BA02) | III | ↑ | - | - | - | ↑ | - | - | - | S | - | - | - | 22A1S 22A2S↓ 22A3S 47A1S 47A2S | - | [218,222,232,233,234,235] | |
Piracetam | N | Other psychostimulants and nootropics (N06BX03) | III | - | - | - | - | - | - | - | - | - | - | - | 0 | - | - | - | [236] |
Propranolol | Y | β-blocking agents, non-selective (C07AA05) | I | - | - | - | - | 1A1↓ 1A2S 3A4S 3A5S 3A7S 2C19S 2D6S↓ | - | - | A↓ | - | - | - | S | - | 22A2↓ | - | [218,237,238,239,240] |
Sertraline | N | Selective serotonin reuptake inhibitors (N06AB06) | I | - | - | - | - | 1A2↓ 3A4S 2B6S↓ 2C9S↓ 2C19S↓ 2D6S↓ 2E1S | - | - | AS BS | - | S | - | S↓ | - | 6A2↓ 6A3↓S 6A4↓S 36A1↓ | 1A3S 1A6S 2B4S 2B7S | [217,218,219,241,242,243] |
Tadalafil | N | Drugs used in erectile dysfunction (G04BE08) | II | - | - | - | - | 3A4S 3A5S | - | - | - | - | - | - | S | - | - | - | [218,219,244] |
| Bioactive | Plant Sources | ALOX5 | AMPK | BSEP | COX | CYP | GLP-1 | HMGCoAR | MAO | NMDA | ABC | PLA2G2A | SLC | UGT | Reference | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BCRP | MRP | PgP | PEPT; OCT; OAT, MATE, OATP | ||||||||||||||
| 11-Angeloylsenkyunolide F * | A. sinensis | — | — | — | — | 3A4↑2D6↑ | — | — | — | — | — | — | — | — | — | — | [246] |
| Apigenin * | M. chamomilla M. officinalis P. emblica S. costus | — | — | — | — | 1A2↓ 2C9↓ 2C19↓ 3A4↓ | — | — | — | — | ↓ | — | ↓ | — | 21A6↓ 21A8↓ 21A9↓ 22A8↓ 22A11↓ 22A4↓ | 1A1↑ | [200,247,248,249,250,251,252,253] |
| α-Asarone | A. calamus A. gramineus | — | — | — | 2↓ | 1A1↓ 3A4↓ 2B6↓ 2C8↓ 2C9↓ 2C19↓ 2D6↓ 2E1↓ | — | ↓ | A↓ B↓ | Ant | — | — | ↓ | — | — | — | [254,255,256,257,258,259,260] |
| β-Asarone | A. calamus A. gramineus R. acori | — | ↑ | — | 2↓ | 3A4↓ | — | — | A↓ B↓ | — | — | — | ↓ | — | — | — | [255,259,261] |
| Asiatic acid * | C. asiatica | — | — | — | — | 3A4↓ 2D6↓ 2C9↓ | — | — | — | — | — | — | S | — | — | — | [262,263,264,265] |
| Assoanine | N. assoanus | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Atractylenolide III * | A. japonica A. lancea | — | — | — | — | — | — | — | — | — | — | — | — | — | — | 2B7↓ | [266] |
| Avenanthramide A * | A. sativa | — | — | — | 2↓ | — | — | — | — | — | — | — | ↓ | — | — | — | [267,268] |
| Avenanthramide B * | A. sativa | — | — | — | 2↓ | — | — | — | — | — | — | — | ↓ | — | — | — | [268] |
| Avenanthramide C * | A. sativa | — | ↑ | — | 2↓ | — | — | — | — | — | — | — | ↓ | — | — | — | [268,269] |
| Bacoside A * and B * | B. monnieri | — | — | — | — | 1A2↓ 3A4↓ 2C9↓ 2C19↓ | — | — | — | — | — | — | — | — | — | — | [270] |
| N-Acetyl Benzylamine * | L. meyenii | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| N-Benzylhexadecanamide * | L. meyenii | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Berberine * | C. chinensis C. japonica | — | — | — | — | 1A2↓ 3A4S↓ 2C9↓ 2D6↓ | — | — | — | — | ↓ | — | ↓↑S | — | 21A8S | — | [271,272,273,274,275,276,277] |
| Biapigenin * | H. perforatum | — | — | — | — | 1A2↓ 3A4↓ 2C9↓ 2D6↓ | — | — | A↓ | Ant | — | — | S | — | — | — | [124,200,278,279] |
| Bilobalide * | G. biloba | — | — | — | — | 1A1↑ 1A2↑ 3A4↑ 2B6↑ 2C9↑ 2E1↑ | — | — | — | ↓ | — | — | — | ↓ | 6A2↓ 22A6↓ 22A8↓ | — | [218,280,281,282] |
| Biochanin A * | T. pratense | — | — | — | 2↓ | 3A4↓ | — | — | B↓ | ↓ | ↓S | — | ↓ | — | 21A6↓ 21A8↓ | — | [245,248,283,284,285,286,287] |
| Buxakashmiramine | B. papillosa | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Buxamine B | B. hyrcana | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Caffeic acid * | C. pluricaulis F. vulgare M. chamomilla M. officinalis R. officinalis V. angustifolium | — | — | — | 1,2↓ | 1A1↓ 3A4↓ 2C9↓ 2C19↓ 2D6↓ | — | — | — | — | ↑ | — | — | ↓ | — | — | [288,289,290,291] |
| Caffein * | C. arabica I. paraguariensis P. cupana T. cacao C. sinensis | — | — | — | — | 1A2↓ 3A4↓ | — | — | — | — | ↓ | — | — | — | — | — | [292,293] |
| β-Carbolines (Harmine) * | L. meyenii M. pruriens P. harmala | — | — | — | — | 1A1↓ 1A2↓ 2C9↓ 2C19↓ 2D6↓ 2E1↓ | — | — | A↓ B↓ | — | ↓ | 2↑S | — | — | — | — | [124,126,294,295,296] |
| 9-Methyl-9B-Carboline * | P. harmala | — | — | — | — | 3A4↓ 2D6S↓ | — | — | A↓ B ↓ | — | — | — | 0 | — | — | — | [127,296,297] |
| Casticin * | V. agnus-castus | — | — | — | — | 3A4↓ 2C9↓ | — | — | — | — | — | — | — | — | — | — | [298] |
| Catalpol * | R. glutinosa | — | — | — | — | 3A4↑↓ 2C9↑↓ 2E1↓ | — | ↓ | — | — | — | — | ↑ | — | — | — | [292,299,300,301] |
| Catechin * | C. rotundus L. bicolor M. chamomilla T. cacao | — | — | — | — | 1A2 ↓ 3A4 ↓ 2C9 ↓ | — | — | — | — | — | — | ↓ | — | 47A1↓ 47A2↓ 22A1↓ 22A2↓ 21A3↓ 21A6↓ | — | [302,303] |
| Chebulagic acid * | T. chebula P. emblica | — | — | — | 2↓ | — | — | — | — | — | — | — | ↓ | — | — | — | [304,305] |
| Chlorogenic acid * | F. vulgare H. perforatum I. paraguariensis M. chamomilla M. lucida R. officinalis | — | ↑ | — | 2↓ | 1A2↓ 3A4o 2E1↓ | — | ↓ | B↓ | — | — | — | — | ↓ | 22A6↓ 22A8↓ | — | [306,307,308,309,310,311] |
| 1,8-Cineole (eucalyptol) * | F. vulgare S. lavandulaefolia R. officinalis | — | — | — | — | 3A4S | — | — | — | — | — | — | S | — | — | — | [312] |
| Cinnamaldehyde * | C. wilsonii | — | ↑ | — | 2↓ | 1A2↓ 2E1↓ | — | — | BS | Ant | — | — | ↓ | — | — | — | [75,313,314,315,316,317] |
| Coniferyl ferulate * | A. sinensis | — | — | — | — | 3A4↑ 2D6↑ | — | — | — | — | — | — | ↓ | — | — | — | [246,318] |
| Convolamine * | C. pluricaulis | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Conypododiol | A. adscendens | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Coptisine | C. chinensis | — | — | — | — | 1A2↓ 3A4↑ 2C9↑ 2D6↓ | — | — | A↓ B↓ | — | — | — | S↑ | — | 22A1S↓ 22A2S↓ | — | [272,273,274,319,320] |
| p-Coumaric acid * | B. rufescens M. lucida | — | ↑ | — | 2↓ | — | — | — | — | — | — | — | — | — | — | — | [321,322] |
| Corilagin * | T. chebula P. emblica | — | — | — | — | — | — | — | — | — | — | 2↑ | — | — | — | 2B4↑ | [323] |
| Crocetin * | C. sativus | — | — | — | — | — | — | — | — | Ant | — | 1↓ 2↓ | S | — | — | — | [324,325,326] |
| Crocin * | C. sativus | — | — | — | 1↓ 2↓ | 3A4↓ 3A5↓ 3A7↓ 2B6↓ | — | — | A↓ B↓ | — | — | 1↓ 2↓ | ↓ | — | — | — | [327,328,329] |
| Curcumin * | C. longa | — | — | — | — | 1A2↓ 3A4↓ 2B6↓ 2C9↓ 2D6↓ | — | — | — | — | ↑↓ | 1↓ 2↓ | ↓ | — | 21A6↓ 21A8↓ | 1A1↓ 1A6↓ 1A9↓ | [247,330,331,332,333,334,335,336,337,338,339] |
| Cyanidin-3-O-β-arabinose * | V. angustifolium | — | — | — | — | 1A2↓ 3A4↓ | — | — | — | — | — | — | — | — | — | — | [340] |
| Cyanidin-3-O-β-galactoside * | V. angustifolium | — | — | — | — | 1A2↓ 3A4↓ 2C9↓ | — | — | — | — | S | — | 0 | — | — | — | [341,342] |
| Cyanidin-3-O-β-glucoside * | E. amoenum V. angustifolium V. uliginosum L. | — | — | — | 2↑ | 3A4↓ 2C9↓ | — | — | — | — | S | — | — | ↓ | 21A9↓ | — | [341,343,344,345] |
| Cyclomicrophylline A | B. papillosa | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Cycloprotobuxine-C | B. papillosa | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Cyclovirobuxeine-A | B. papillosa | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Cynatroside A | C. atratum | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | |
| Cynatroside B | C. atratum | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Daidzein * | L. bicolor M. lucida P. lobata | — | — | — | — | 1A1S 1A2S 3A4↓ | — | — | — | — | ↓S | 1↑ | ↑ | — | — | 1A1S 1A9S | [245,285,346,347,348,349] |
| Decursin * | A. gigas | — | — | — | — | 1A1↓ 1A2↓ | — | — | — | — | — | 2↓ | ↓ | — | — | — | [247,350] |
| Decursinol angelate * | A. gigas | — | — | — | — | 1A1↓ 1A2↓ 3A4S 2C19S | — | — | — | — | — | — | S | — | — | — | [350,351,352] |
| Delphinidin * | V. uliginosum L. | — | — | — | — | 3A4↓ 2B6↓ 2C9↓ | — | — | — | — | ↓ | — | — | — | 21A8↓ | — | [341,344,353] |
| Delphinidin-3-O-β-galactoside * | V. angustifolium | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Diosgenin * | D. polystachya D. oppositifolia T. foenum-graecum | — | — | — | 2↑ | 3A4↓ | — | — | — | — | — | — | S | — | — | — | [354,355,356] |
| Ellagic acid * | T. chebula P. emblica | — | — | — | — | 1A1↑↓ 2B6↓ 2E1↓ | — | — | — | — | ↓ | — | S | — | 22A6↓ 22A11↓ | — | [357,358,359,360,361,362] |
| Ent-kaurane * | C. tonkinensis | — | ↑ | — | 2↓ | — | — | — | — | — | — | — | ↓ | — | — | — | [363,364,365] |
| Epicatechin gallate (ECG) * | C. sinensis | — | — | — | — | 1A1↓ 1A2↓ 3A4↓ | — | — | — | — | — | — | ↓ | — | 21A6S↓ 21A8↓ | — | [366,367,368,369] |
| Epigallocatechin-3-gallate (EGCG) * | C. sinensis | — | — | — | — | 1A1↓ 1A2↓ 3A4↓ 3A5↓ 2B6↓ 2C8↓ 2C9↓ 2C19↓ 2D6↓ 2E1↓ | — | — | — | — | ↓ | — | S↓ | — | 21A6↓ 21A8S↓ 21A9↓ 22A1↓ 22A2↓ 47A1↓ 47A2↓ | — | [248,302,303,345,367,370,371,372,373,374,375,376] |
| Epinorgalantamine | N. confusus G. woronowii | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Eugenol * | C. wilsonii R. acori R. officinalis | — | ↓ | — | 2↓ | 1A1↓ 2E1S | — | 0 | A↓ | ↓ | — | — | 0↓ | — | — | 1A1↑ | [77,247,377,378,379,380,381,382,383] |
| Ferulic acid * | A. sinensis B. rufescens | — | — | — | — | 1A1↑ 1A2↑S 3A4↑S 2B6↑S 2C8↑S 2C9↑S 2C19↑S 2D6↑S | — | — | B↓ | Ant | — | — | ↓ | — | — | 1A1S 2B7S | [246,384,385,386,387] |
| Forskolin * | C. forskohlii | — | ↓ | — | 2↓ | 3A4S↑ 2C↑ 2B6↑ | ↑ | — | — | ↑ | — | — | s↓ | — | — | — | [388,389,390,391,392,393,394,395,396,397] |
| Galantamine * | G. nivalis G. woronowii L. radiata N. confusus P. illyricum | — | — | — | — | 3A4S 0 2D6S | — | — | — | — | — | — | ↓ | — | — | 1A1S | [398,399,400,401,402] |
| 11-Hydroxy Galantamine * | N. poeticus | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Gallic acid * | M. pruriens P. emblica T. chebula | — | — | — | 2↓ | 3A4↓ 2D6↓ | — | — | A↓ | — | — | — | ↓ | — | 21A8↓ | 2B7↑ | [403,404,405,406,407,408,409] |
| Genistein * | L. bicolor M. lucida M. pruriens | — | ↑ | — | 2↓ | 1A1↓ 1A2↓S 3A4↑ 2C8↓ 2C9↓ 2D6↓ | — | ↓ | A↓ B↓ | ↓ | ↓S | 1↓ 2↓S | ↓ | — | 21A6↓S | — | [218,285,369,374,409,410,411,412,413,414,415] |
| Gingerol * | A. melegueta Z. officinale Rosc. | — | — | — | — | 3A4↓ 2C9↓ 2C19↓ | — | — | — | — | — | — | ↓ | — | — | — | [416,417] |
| Ginkgolide A * | G. biloba | — | — | — | 2↓ | 3A4↓ 2C9↓ | — | — | — | ↓ | — | — | ↑ | ↓ | 6A2↓ 22A6↓ 22A8↓ | — | [218,281,418,419,420] |
| Ginkgolide B * | G. biloba | — | — | — | — | 3A4↓ 2C9↓ | — | — | — | — | — | — | ↑ | ↓ | 6A2↓ 22A6↓ 22A8↓ | — | [218,281,418] |
| Ginsenoside Rb * | P. ginseng | — | — | — | — | 1A1↑ 1A2↓ 3A4↓ 2C9↓ | — | — | — | — | ↓ | — | — | — | 21A8S | — | [421,422,423] |
| Ginsenoside Rc * | P. ginseng | — | — | — | — | 3A4↓ 2C9 ↓ | — | — | — | — | — | — | — | — | 21A8S | — | [421,423] |
| Ginsenoside Rd * | P. ginseng | — | — | — | — | 3A4↓ 2C9↓ 2C19↓ 2D6↓ | — | — | — | — | ↓ | — | — | — | 21A8S | — | [421,423] |
| Ginsenoside Rg * | P. ginseng | — | — | — | — | 1A1↑ 1A2↓ 3A4↓ 2C9↓ 2D6↓ | — | — | — | — | S | — | — | — | 21A8S | — | [421,422,423] |
| 1,2,3,4,6-Penta-O-galloyl-β-d-glucose * | T. chebula | — | — | — | 2↓ | — | — | — | — | — | — | — | ↓ | — | — | — | [424,425] |
| Glycyrrhetinic acid * | G. glabra | — | — | — | — | — | — | — | — | — | — | — | — | — | 21A9↓ | — | [426] |
| Glycyrrhizin * | G. glabra | — | — | S | — | 3A4↓ 2D6↓ | — | — | — | — | — | — | ↑↓ | — | 21A8↓ | — | [247,257,427,428,429] |
| Haloxysterol A *, B *, C *, D * | H. recurvum | — | — | — | 2↓ | — | — | — | — | — | — | — | — | — | — | — | [430] |
| Harpagide * | S. buergeriana | — | — | — | — | 3A40 | — | — | — | — | — | — | — | — | — | — | [431] |
| 8-O-E-p-Methoxy Cinnamoyl Harpagide | S. buergeriana | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| E-Harpagoside * | S. buergeriana | — | — | — | 1↓0 2↓0 | 3A40 | — | — | — | — | — | — | ↓ | — | — | — | [431,432,433,434] |
| Huperzine A * | H. serrata | — | — | — | — | 1A2↑ 3A4↑0 | — | — | — | — | — | — | S | — | — | — | [435,436,437] |
| Hyperforin * | H. perforatum | ↓ | — | — | 1↓ 20 | 2D6↓0 2C9↓ 3A4↓ 3A5↓ 1A2↓ 2C19↓ | — | — | A0 | Ant | ↓ | 2↑ | ↑↓ | — | 21A6↓ 21A8↓ 21A9↓ | 1A1↑↑ | [98,124,202,278,438,439,440,441,442,443,444,445,446,447,448,449] |
| Hypericin * | H. perforatum | — | ↑ | — | 2D6↓0 2C9↓ 3A4↓ 1A2↓ 2C19↓ | — | — | A↓0 B↓0 | Ant | ↓ | 1↑ | ↑ | — | 21A9↓S | 1A6↓ | [102,124,278,336,442,443,444,447,450,451,452,453,454,455] | |
| Hyperoside * | B. rufescens C. japonica H. perforatum P. pterocarpum | — | — | — | — | 2D6↓ 2C9↓ | — | — | — | — | — | — | S | — | — | — | [289,456] |
| Isoliquiritigenin * | G. uralensis | — | — | — | — | 1A2↓ 3A4↓ 2C9↓ 2C19↓ | — | — | — | — | — | — | — | — | — | — | [457] |
| Isorhamnetin * | G. biloba R. nigrum | — | — | — | — | 1A2↓ 3A4↓ | — | — | — | — | — | 2↓ | S↑↓ | — | 21A8↓ | — | [200,247,249,458,459] |
| Isosarcodine | S. saligna | — | — | — | 2↓ | — | — | — | — | — | — | — | — | — | — | — | [430] |
| p-Kaempferol * | A. melegueta C. pluricaulis C. japonica C. rotundus H. perforatum I. paraguariensis | — | — | — | — | 1A2↓ 3A4↓ 2C9↓ 2C19↓ | — | — | — | Ant | — | — | ↓S | — | 21A8↓ 21A9↓ 22A4↓ 22A8↓ | 1A1↑ | [200,218,250,251,374,460,461,462,463] |
| Kaempferol-3-glucoside * | C. ternatea C. pluricaulis | — | — | — | — | — | — | — | — | — | — | — | — | — | 21A6↓ 21A8↓ | — | [345,463] |
| 11α-Hydroxy-O-methylleucotamine | P. illyricum | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| N-3-methoxybenzyl-linoleamide (macamide) * | L. meyenii | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Levodopa * | M. pruriens | — | — | — | — | — | — | — | — | Ant | — | — | S | — | — | — | [464,465] |
| Luteolin * | L. bicolor M. chamomilla M. officinalis P. emblica R. officinalis | — | ↑ | — | 2↓ | 1A2↓ 3A4↓ 3A50 2B6↓ 2C8↓ 2C9↓ 2C19↓ 2D6↓ 2E1↓ | — | ↓ | A↓ B↓ | — | ↓ | — | — | — | — | 1A1↓S 1A9↓ 2B7↓ | [276,466,467,468,469,470,471,472] |
| Lycorine | G. woronowii P. illyricum | — | — | — | — | 3A40 | — | — | — | — | — | — | 0 | — | — | — | [342,473] |
| Malvidin 3-galactoside * | V. angustifolium | — | — | — | — | 3A4↓ 2C9↓ | — | — | — | — | ↓S | — | ↓ | — | 21A6↑ | — | [341,353,474] |
| Malvidin 3-glucoside * | V. angustifolium V. uliginosum L. | — | — | — | — | 3A4↓ 2C9↓ | — | — | — | — | ↓ | — | ↓ | — | 21A6↑ | — | [341,353,474] |
| Malvidin-3-O-β-arabinose * | V. angustifolium | — | ↑ | — | — | — | — | — | — | - | — | — | — | — | — | — | [475] |
| Mangiferin * | M. indica | — | — | — | — | 1A1↓ 1A2↓ 3A4↓ 2C8↓ 2B6↓ 2D6↓ | — | — | — | — | — | — | ↓ | — | — | 1A1↓ 1A9↓ 2B7↓ | [198,476,477] |
| Methylliberine * | C. arabica | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Mucunadine * | M. pruriens | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Mucunine * | M. pruriens | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Myricetin * | M. peregrina R. nigrum | — | — | — | — | 1A2↓ 3A4↓ 2D6↓ | — | — | — | — | ↓ | 1↓ | ↓S | — | 21A6↓ | — | [218,247,248,276,478] |
| Naringenin | L. bicolor M. lucida | — | ↑ | — | 2↓ | 1A2↓0 3A4↓ 2B60 2C9↓ 2C19↓ 2D60 2E10 | — | ↓ | A↓ B↓ | — | ↓ | — | ↓ | — | 22A6↓ 21A6↓ 21A8↓ 21A9↓ | 1A1↑ 1A3↑ 1A6↑ 1A9↑ 2B4↑ 2B7↑ | [200,218,247,253,479,480,481,482,483,484,485,486,487,488] |
| Naringin * | M. lucida | — | ↑ | — | 2↓ | 3A4↓↑ 1A2↓ | — | ↓ | A↓ | — | ↑ | — | ↓ | — | 21A9↓ | — | [200,253,338,483,487,489,490,491,492,493] |
| N,N-dimethyl buxapapine | B. papillosa | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Nobiletin * | C. reticulata | — | — | — | 2↓ | 1A1↓↑ 1A2↓↑ 3A4↓ | — | — | A↓ B↓ | — | ↓ | 1↓ | ↓ | — | 21A8↓ 21A9↓ | — | [208,247,337,487,494,495,496,497] |
| Oleanolic acid * | M. lucida R. officinalis | — | ↑ | — | 2↑ | 1A2↓ 3A4↓ | — | ↓ | A↓ | —- | ↓ | 1↓ | 0 | — | — | — | [276,498,499,500,501,502,503] |
| α-Onocerin * | L. clavatum | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| p-Synephrine * | C. aurantium | — | — | — | — | 1A20 3A4↓ 2D60 2E10 | — | — | AS BS | — | — | — | ↓ | — | — | — | [79,504,505,506] |
| Paeonol * | P. lactiflora | — | — | — | 2↓ | 1A2S | — | — | A↓ B↓ | — | ↓ | — | S | — | — | — | [507,508,509,510,511] |
| Palmatine | C. chinensis C. speciosa | — | — | — | 2↓ | 1A1↓ 1A2S 3A4↑ 2D6S | — | — | A↓ | — | — | — | ↑ | — | — | — | [274,512,513,514,515] |
| Peonidin-3-O-β-arabinose * | V. angustifolium | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| d-Pinitol | B. rufescens | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| 15-Methyl Pinusolidic acid * | P. orientalis | — | — | — | — | — | — | — | — | A | — | — | — | — | — | — | [133] |
| Phyllanthin * | P. emblica | — | — | — | — | 1A20 3A4↓ 2C90 2D60 2E10 | — | — | — | — | — | 20 | ↓ | — | — | — | [516,517] |
| Phytol | M. lucida | — | ↑ | — | 2↓ | — | — | ↓ | — | — | — | — | — | — | — | — | [518,519,520] |
| α-Pinene * | R. officinalis S. lavandulaefolia | — | — | — | 2↓ | 2B6↓ | — | — | — | Ant | — | — | — | — | — | — | [521,522,523] |
| Piperine * | P. nigrum | — | — | — | — | 1A2↑ 3A4↓ 2C9↓ 2E1↓ | — | — | — | — | ↓ | — | ↓ | — | 22A2↓ | 1A1↓ | [524,525,526] |
| Polygalaxanthone III * | P. tenuifolia | — | — | — | — | 2E1↓ | — | — | — | — | — | — | — | — | — | — | [527] |
| Protocatechuic acid * | A. oxyphylla G. biloba M. officinalis R. officinalis V. vinifera | — | — | — | — | 1A1↓ 1A2↓ 2C9↓ 2E1↓ | — | — | — | — | — | — | — | — | — | — | [245,528] |
| Prurienidine * | M. pruriens | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Prurienine * | M. pruriens | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Puerarin * | P. lobata | — | ↑ | — | 2↓ | 1A1↑ 1A2↑ 3A4↓ 2B6↓ 2C9↓ 2D6↓ 2E1↓ | — | ↓ | — | ↓ | — | — | ↑ | — | — | 1A1↑ | [529,530,531,532,533,534,535,536,537] |
| Quercetin * | A. melegueta C. sativus C. rotundus H. perforatum I. paraguariensis L. meyenii M. lucida P. emblica R. nigrum S. costus V. uliginosum L. | — | — | — | — | 1A1↓ 1A2↓ 3A4↓ 2C8↓ 2C9↓ 2C19↓ 2D6↓ | — | — | A↓ | Ant | ↑S | 1↓ 2↓ | ↓S | — | 21A6↓ 21A8↓ 21A9↓ 22A6↓ | 1A1↑ 1A6↑ | [24,200,218,247,289,338,374,462,487,488,538,539,540,541,542] |
| Quercetin-3-O-β-d-glucuronide | P. pterocarpum | — | — | — | 2↓ | 3A4↓ | — | — | — | — | — | 2↓ | ↓ | — | 21A8↓ 21A9↓ | 1A1↓ | [539,543] |
| Resveratrol * | V. vinifera | — | — | — | — | 1A1↓S 1A2↓ 3A4↓ 2C9↓ 2C19↓ 2D6↓ | ↑ | ↓ | A↓ | ↓ | ↑S | 2S | ↓ | — | 21A8S | — | [218,298,338,544,545,546,547,548,549,550,551] |
| Rosarin * | R. rosea | — | — | — | — | 1A2↓ 3A4↓ 2C9↓ 2D6↓ | — | — | A0 B0 | — | — | — | — | — | — | — | [552,553] |
| Rosavin * | R. rosea | — | — | — | — | 1A2↓ 3A4↓ 2C9↓ 2D6↓ | — | — | A0 B0 | — | — | — | — | — | — | — | [552,553] |
| Rosin * | R. rosea | — | — | — | — | 1A2↓ 3A4↓ 2C9↓ 2D6↓ | — | — | A0 B0 | — | — | — | — | — | — | — | [552,553] |
| Rosmarinic acid * | M. officinalis M. spicata R. officinalis | — | — | — | — | 3A4↓ 2C9↓ 2C19↓ 2D6↓ 2E1↓ | — | — | — | — | ↓ | — | ↓ | — | — | 1A1↓ | [416,554,555] |
| Rutin * | H. perforatum L. bicolor M. chamomilla M. flexuosa M. lucida M. peregrina V. uliginosum L. | — | ↑ | — | 2↓ | 1A1↑ 3A4↓ | — | ↓ | B↓ | — | ↓ | — | ↓ | — | 21A6↓ 21A8↓ 21A9↓↑ | — | [24,248,253,345,426,556,557,558,559,560,561,562,563] |
| Safranal * | C. sativus | — | — | — | — | 2B6↑ | — | — | — | — | ↓ | — | ↓ | — | — | — | [247,328,564] |
| Salidroside * | R. rosea | ↓ | — | — | 1↓ 2↓ | 1A2↑ 3A4↓ 2B6↑ 2C9↑ | — | — | A↓ B↓ | ↑ | — | 1↓ | ↓ | ↓ | — | — | [565,566,567,568,569,570] |
| Salvianolic acid * | M. spicata S. miltiorrhiza | — | — | — | 2↓ | 1A2↓ 3A4↓ | — | — | — | — | ↑ | — | ↓ | — | 21A8S | — | [571,572,573,574] |
| Sanguinine | E. grandiflora P. illyricum | — | — | — | — | 3A40 | — | — | — | — | — | — | — | — | — | — | [399] |
| Sarcocine | S. saligna | — | — | — | — | — | — | — | — | — | — | — | — | — | 15A1S 15A2S | — | [575] |
| Sarcodine | S. saligna | — | — | — | 2↓ | — | — | — | — | — | — | — | — | — | — | — | [430] |
| Sarcorine | S. saligna | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Sarsalignenone | S. saligna | — | — | — | 2↓ | — | — | — | — | — | — | — | — | — | — | — | [430] |
| Sarsasapogenin * | A. racemosus | — | — | — | — | — | — | — | A↓ B↓ | — | — | — | — | — | 21A6S | — | [369,576] |
| Schisandrin B * | S. chinensis | — | — | — | — | 3A4↓ 3A5↓ 2B6↑ 2E1S | — | — | — | — | — | — | ↓ | — | 21A6↑ | — | [577,578,579,580,581] |
| Shatavarin IV * | A. adscendens A. racemosus | — | — | — | — | — | — | — | A↓ B↓ | — | — | — | — | — | — | — | [582] |
| Shogaol * | A. melegueta Z. officinale Rosc. | — | — | — | 1↓ 2↓ | 1A2↓ 2C9↓ 2C19↓ | — | — | — | ↓ | — | — | — | — | — | 1A1S 1A3S 2B7S | [583,584,585,586] |
| Sitoindosides VII-X * | W. somnifera | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| β-Sitosterol * | A. lancea C. pluricaulis M. peregrina M. pruriens | — | ↑ | — | 2↓ | 3A40 3A50 2C190 | — | ↓ | — | — | ↓ | — | 0 | — | — | — | [587,588,589,590,591,592] |
| Spirofornabuxine | B. hyrcana | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Stigmasterol * | A. lancea | — | — | — | — | 3A4↓ 3A5↓ | — | — | — | — | — | — | ↓ | — | — | — | [590,593] |
| Syringin * | E. senticosus | - | - | - | 2↓ | 2C9↓ 2E1↓ | — | — | — | — | — | — | S | — | — | — | [93,594] |
| Tannic acid * | T. chebula | — | — | — | 2↓ | 1A2↓ 3A4↓ 2B6↓ | — | — | — | — | — | 1↓ 2↓ | ↓ | — | 21A6S↓ | — | [333,369,595,596,597,598] |
| Tanshinone I * | S. miltiorrhiza | — | — | — | — | 1A1↑ 1A2↑↓ 2C9↓ 2E1↓ | — | — | — | — | ↓ | — | S↓ | — | — | 1A1o 1A30 1A90 2B70 | [24,599,600,601,602] |
| Tanshinone IIA * | S. miltiorrhiza | — | — | — | 2↓ | 1A1↑ 1A2↑↓ 3A4↑↓ 2C9↓ 2E1↓ | — | — | — | — | ↓ | — | ↓ | — | — | 1A1o 1A30 1A90 2B70 | [24,599,600,601,603] |
| Tanshinone IIB * | S.miltiorrhiza | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Tenuifolin * | P.tenuifolia | — | — | — | 2↓ | — | — | — | — | — | — | — | — | — | — | — | [604] |
| Tenuifoliside B * | P. tenuifolia | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Tenuifoliside C * | P. tenuifolia | — | — | — | — | 2E1↓ | — | — | — | — | — | — | — | — | — | — | [527] |
| Theacrine * | C. arabica T. grandiflorum | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Theobromine * | T. cacao | — | — | — | — | 1A2S 3A40 2C90 2C190 2D60 | — | — | — | — | — | — | 0 | — | — | — | [218,605] |
| Timosaponin AIII * | A. asphodeloides | — | ↑ | — | 2↓ | — | — | — | — | — | — | — | ↓ | — | — | — | [606] |
| Trigonelline * | T. foenum-graecum | — | — | — | — | 1A2↓ 3A40 2B60 2C80 2C190 2D6↓ 2E10 | — | ↑ | — | — | ↑ | — | — | — | — | — | [607,608] |
| Ursolic acid * | R. officinalis | — | — | — | 2↓ | 1A2↓ 3A4↓S 2C8↓ 2C9↓S 2C19↓ 2D6↓ | — | — | A↓ | — | ↓ | 10 | — | — | 21A6↓ 21A8↓ 21A9↓ 22A8↓ | — | [276,409,501,609,610,611,612,613] |
| Vaganine | S. saligna | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| (+)-α-Viniferin * | C. chamiague | — | — | — | — | 3A4S | — | — | — | — | — | — | S | — | — | — | [218] |
| Vinpocetine * | V. minor | — | — | — | — | 3A4S | — | — | — | — | — | — | — | — | — | — | [614] |
| Withaferin A * | W. somnifera | — | — | — | — | 1A1↓ | — | — | — | — | — | — | — | — | — | — | [615] |
| Withanolide A * | W. somnifera | — | — | — | — | 1A1↓ | — | — | — | — | — | — | — | — | — | — | [615] |
| Withanolide B * | W. somnifera | — | — | — | — | 1A1↓ | — | — | — | — | — | — | — | — | — | — | [615] |
| Z-Ligustilide * | A. sinensis | — | — | — | — | 1A1↓ 3A4↑ 2D6↑ | — | — | — | — | — | — | — | — | — | — | [246,616] |
| Bioactive | Drug | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Pr | Al | Se | Mt | Di | At | Me | Ta | Cl | |
| Apigenin | x | x | x | x | x | x | x | x | x |
| α-Asarone | x | x | x | - | x | x | x | x | x |
| β-Asarone | x | x | x | ▲ | x | x | - | x | x |
| Bacoside A | x | x | x | - | x | x | ▼ | x | x |
| Bacoside B | x | x | x | - | x | x | ▼ | x | x |
| Berberine | x | x | x | x | x | x | - | x | x |
| Biapigenin | x | x | x | - | x | x | x | x | x |
| Bilobalide | x | x | x | - | x | x | x | x | x |
| Biochanin A | x | x | x | - | x | x | x | x | x |
| Caffeic acid | x | x | x | x | x | x | ▼ | x | x |
| β-Carbolines (Harmine) | x | x | x | x | x | x | ▼ | - | x |
| 9-Methyl-9B-Carboline | x | x | x | - | x | x | - | x | x |
| Catalpol | x | x | x | - | x | x | - | x | x |
| Catechin | x | x | x | x | x | x | x | x | x |
| Chlorogenic acid | x | - | x | ▲ | x | x | ▼ | - | x |
| Cinnamaldehyde | x | x | x | ▲ | x | x | x | x | x |
| Coptisine | x | x | x | x | x | ▲ | x | x | x |
| Crocin | x | x | x | - | x | x | ▼ | x | x |
| Curcumin | x | x | x | x | x | x | ▼ | x | x |
| Cyanidin-3-O-β-glucoside | x | x | x | - | x | x | - | x | x |
| Daidzein | x | x | x | x | x | x | - | x | x |
| Decursin | x | - | x& | - | x | x | - | x | x |
| Delphinidin | x | x | x | x | x | x | ▼ | x | x |
| Ellagic acid | ▼ | - | x | x | x | x | ▼ | - | x |
| ECG | x | x | x | - | x | x | - | x | x |
| EGCG | x | x | x | x | x | x | x | x | x |
| Eugenol | ▼ | - | x | ◄► | ▼ | x | x | - | x |
| Ferulic acid | x | x | x | - | x | x | x | x | x |
| Forskolin | x | x | x | ▲ | x | x | x | x | x |
| Gallic acid | x | x | x | - | x | x | - | x | x |
| Genistein | x | x | x | x | x | x | x | x | x |
| Gingerol | x | x | x | - | x | x | ▼ | x | x |
| Ginkgolide A | x | x | x | - | x | x | ▼ | x | x |
| Ginkgolide B | x | x | x | - | x | x | ▼ | x | x |
| Ginsenoside Rb | x | x | x | x | x | x | - | x | x |
| Ginsenoside Rd | x | x | x | x | x | x | ▼ | x | x |
| Ginsenoside Rg | x | x | x | - | x | x | - | x | x |
| Glycyrrhizin | x | x | x | - | x | x | - | x | x |
| Hyperforin | x | x | x | x | x | x | x | x | x |
| Hypericin | x | x | x | x | x | x | x | x | x |
| Isoliquiritigenin | x | x | x | - | x | x | ▼ | x | x |
| Isorhamnetin | x | x | x | - | x | x | - | x | x |
| p-Kaempferol | x | x | x | - | x | x | x | x | x |
| Luteolin | x | x | x | x | x | x | ▼ | x | x |
| Malvidin 3-galactoside | x | x | x | x | x | x | - | x | x |
| Malvidin 3-glucoside | x | x | x | x | x | x | - | x | x |
| Mangiferin | x | x | x | - | x | x | ▼ | x | x |
| Myricetin | x | x | x | x | x | x | - | x | x |
| Naringenin | x | x | x | x | x | x | ▼ | x | x |
| Naringin | x | x | x | x | x | x | - | x | x |
| Nobiletin | x | x | x | x | x | x | - | x | x |
| Oleanolic acid | x | x | x | x | x | x | - | x | x |
| Paeonol | ▼ | - | x | x | x | x | - | - | - |
| Palmatine | x | x | x | - | x | x | - | x | x |
| Piperine | x | x | x | x | x | x | x | x | x |
| Protocatechuic acid | x | x | x | - | x | ◄► | - | - | x |
| Puerarin | x | x | x | ▲ | x | x | ▼ | x | x |
| Quercetin | x | x | x | x | x | x | x | x | x |
| Quercetin-3-O-β-d-glucuronide | x | x | x | - | x | x | - | x | x |
| Resveratrol | x | x | x | x | x | x | x | x | x |
| Rosarin | x | x | x | - | x | x | - | x | x |
| Rosavin | x | x | x | - | x | x | - | x | x |
| Rosin | x | x | x | - | x | x | - | x | x |
| Rosmarinic acid | x | x | x | x | x | x | ▼ | x | x |
| Rutin | x | x | x | x | x | x | - | x | x |
| Salidroside | x | x | x | - | x | x | x | x | x |
| Salvianolic acid | x | x | x | x | x | x | - | x | x |
| Schisandrin B | x | x | x | - | x | x | ▲ | x | x |
| Shogaol | x | x | x | - | x | ▼ | x | - | x |
| β-Sitosterol | - | - | x# | x | x | x | - | - | - |
| Tannic acid | x | x | x | - | x | x | ▼ | x | x |
| Tanshinone I | x | x | x | x | x | x | - | x | x |
| Tanshinone IIA | x | x | x | x | x | x | - | x | x |
| Trigonelline | x | - | x | x | x | x | - | - | x |
| Ursolic acid | x | x | x | x | x | x | ▼ | x | x |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Auxtero, M.D.; Chalante, S.; Abade, M.R.; Jorge, R.; Fernandes, A.I. Potential Herb–Drug Interactions in the Management of Age-Related Cognitive Dysfunction. Pharmaceutics 2021, 13, 124. https://doi.org/10.3390/pharmaceutics13010124
Auxtero MD, Chalante S, Abade MR, Jorge R, Fernandes AI. Potential Herb–Drug Interactions in the Management of Age-Related Cognitive Dysfunction. Pharmaceutics. 2021; 13(1):124. https://doi.org/10.3390/pharmaceutics13010124
Chicago/Turabian StyleAuxtero, Maria D., Susana Chalante, Mário R. Abade, Rui Jorge, and Ana I. Fernandes. 2021. "Potential Herb–Drug Interactions in the Management of Age-Related Cognitive Dysfunction" Pharmaceutics 13, no. 1: 124. https://doi.org/10.3390/pharmaceutics13010124
APA StyleAuxtero, M. D., Chalante, S., Abade, M. R., Jorge, R., & Fernandes, A. I. (2021). Potential Herb–Drug Interactions in the Management of Age-Related Cognitive Dysfunction. Pharmaceutics, 13(1), 124. https://doi.org/10.3390/pharmaceutics13010124