Achillin Increases Chemosensitivity to Paclitaxel, Overcoming Resistance and Enhancing Apoptosis in Human Hepatocellular Carcinoma Cell Line Resistant to Paclitaxel (Hep3B/PTX)

 , , , ,
, , , ,
Abstract
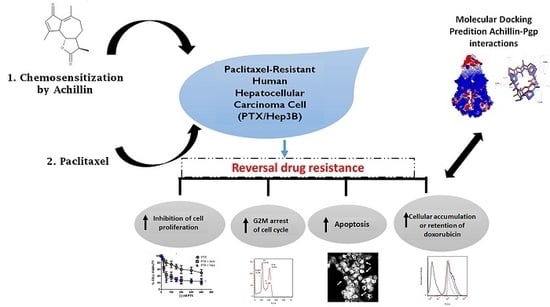
1. Introduction
2. Materials and Methods
2.1. Drugs and Chemicals
2.2. General
2.3. Plant Material
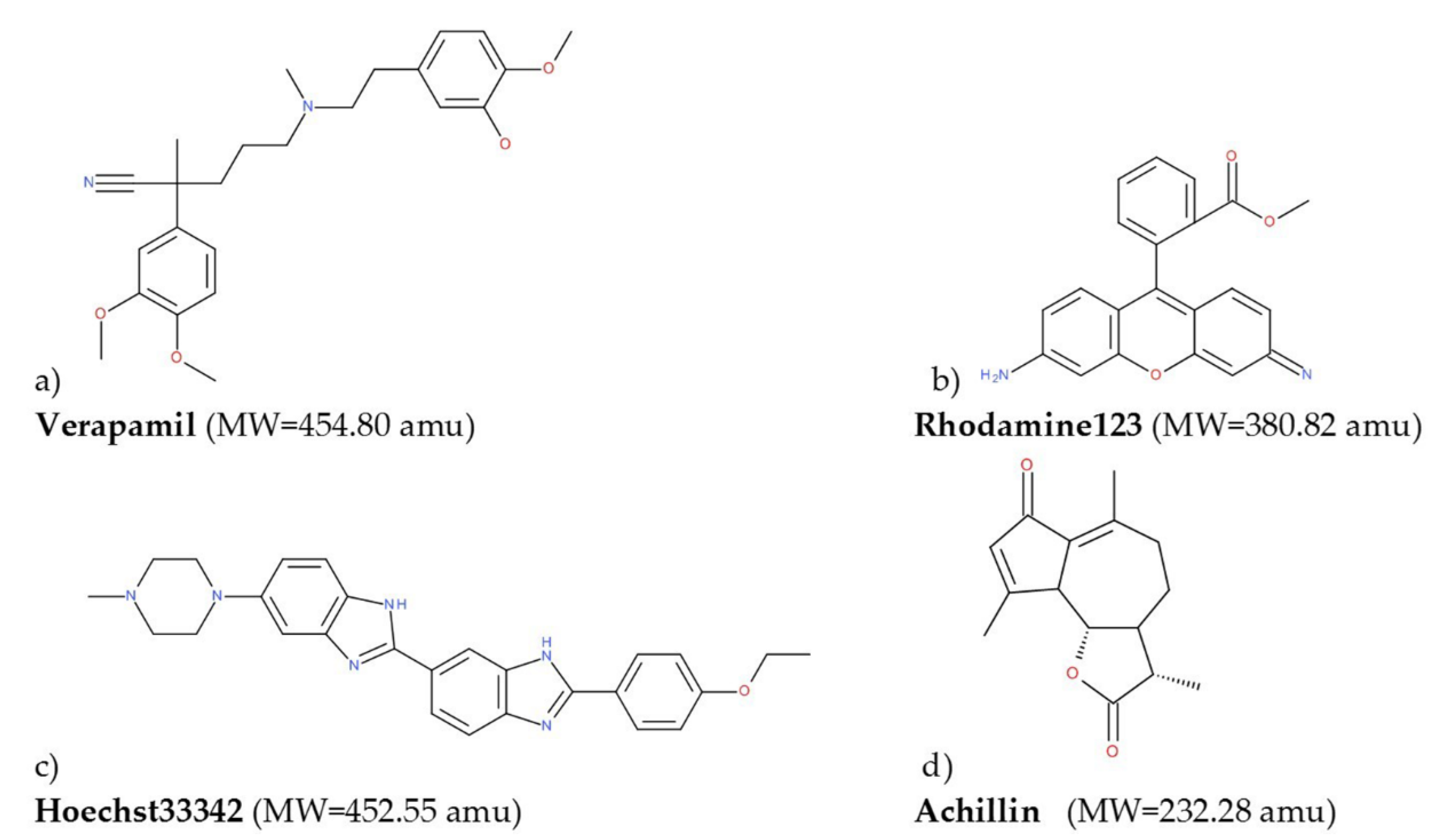
2.4. Isolation of Achillin
2.5. Chemosensitizing Potential to Paclitaxel
Analysis of Combination Effects
2.6. Cellular Doxorubicin Retention Assay
2.7. Cell Cycle Analysis
2.8. Flow Cytometric Apoptosis Assay
2.9. Immunofluorescence
2.10. RT-PCR
2.11. Caspases Activity
2.12. Computational Details
2.12.1. Homology Structural Modeling
2.12.2. Molecular Docking Calculations
2.13. Statistical Analysis
3. Results
3.1. Characterization of Resistant Hep3B/PTX Cells
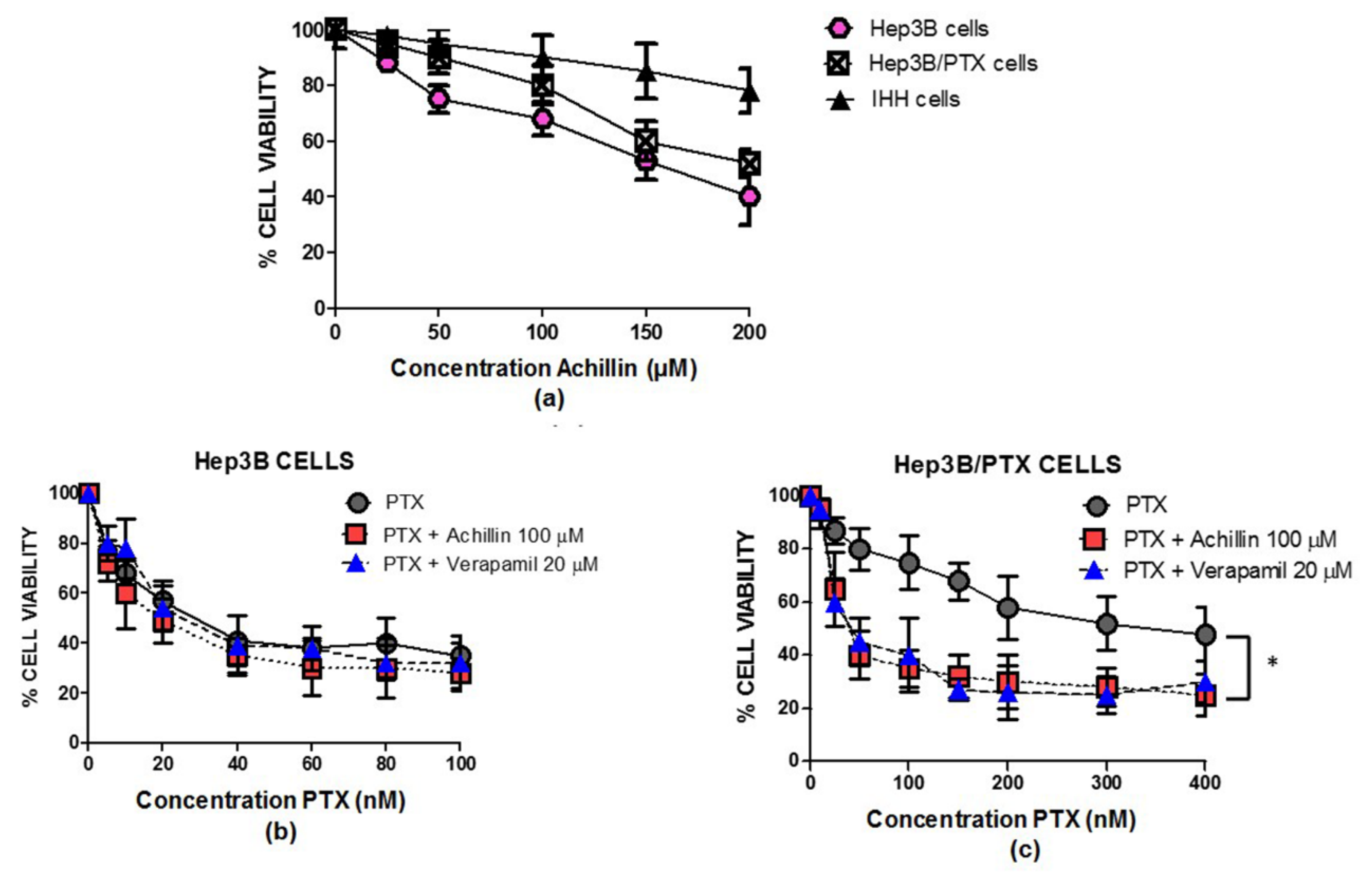
3.2. Achillin Increases the Sensitivity of Hep3B/PTX Cells
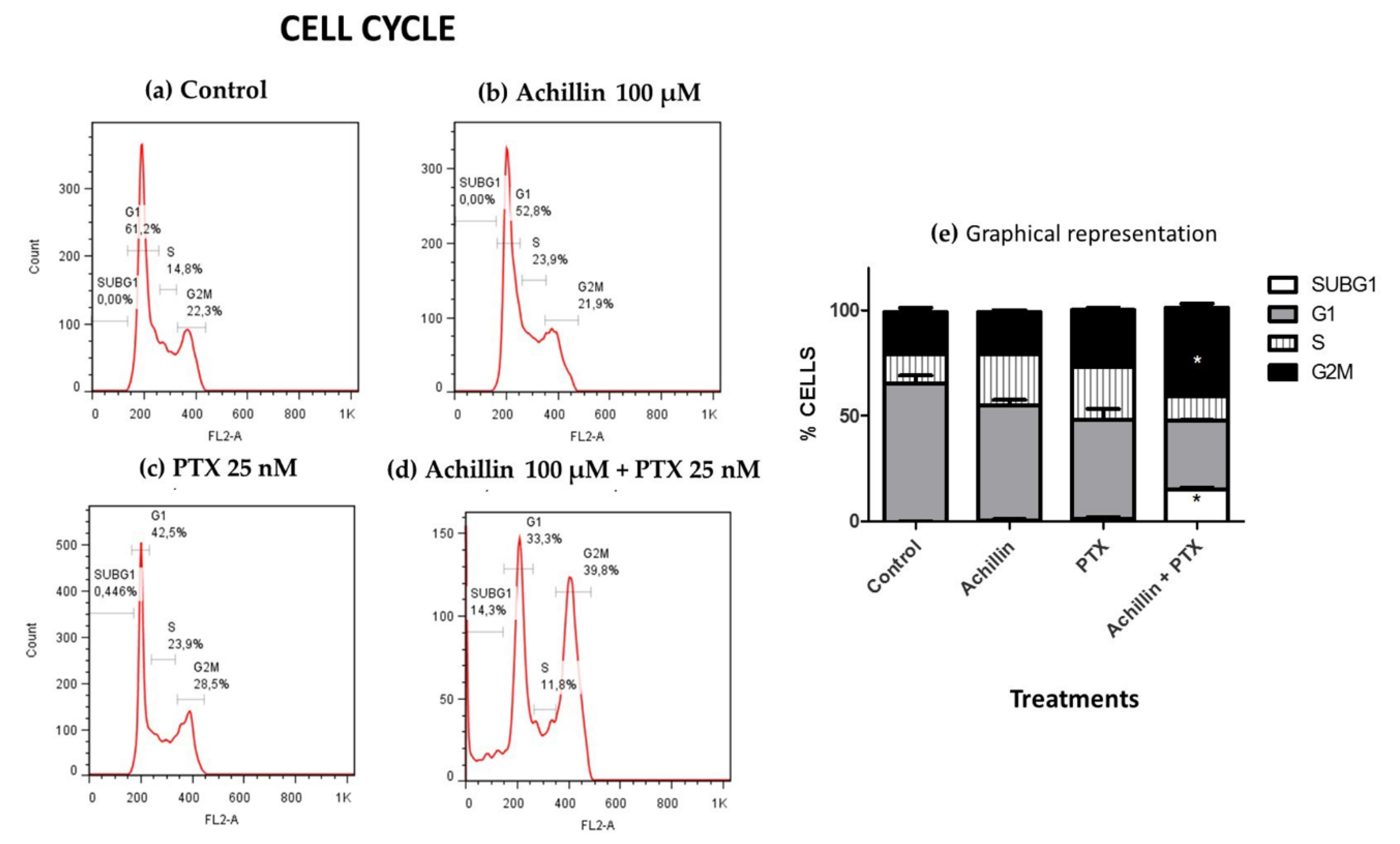
3.3. Cell Cycle Distribution
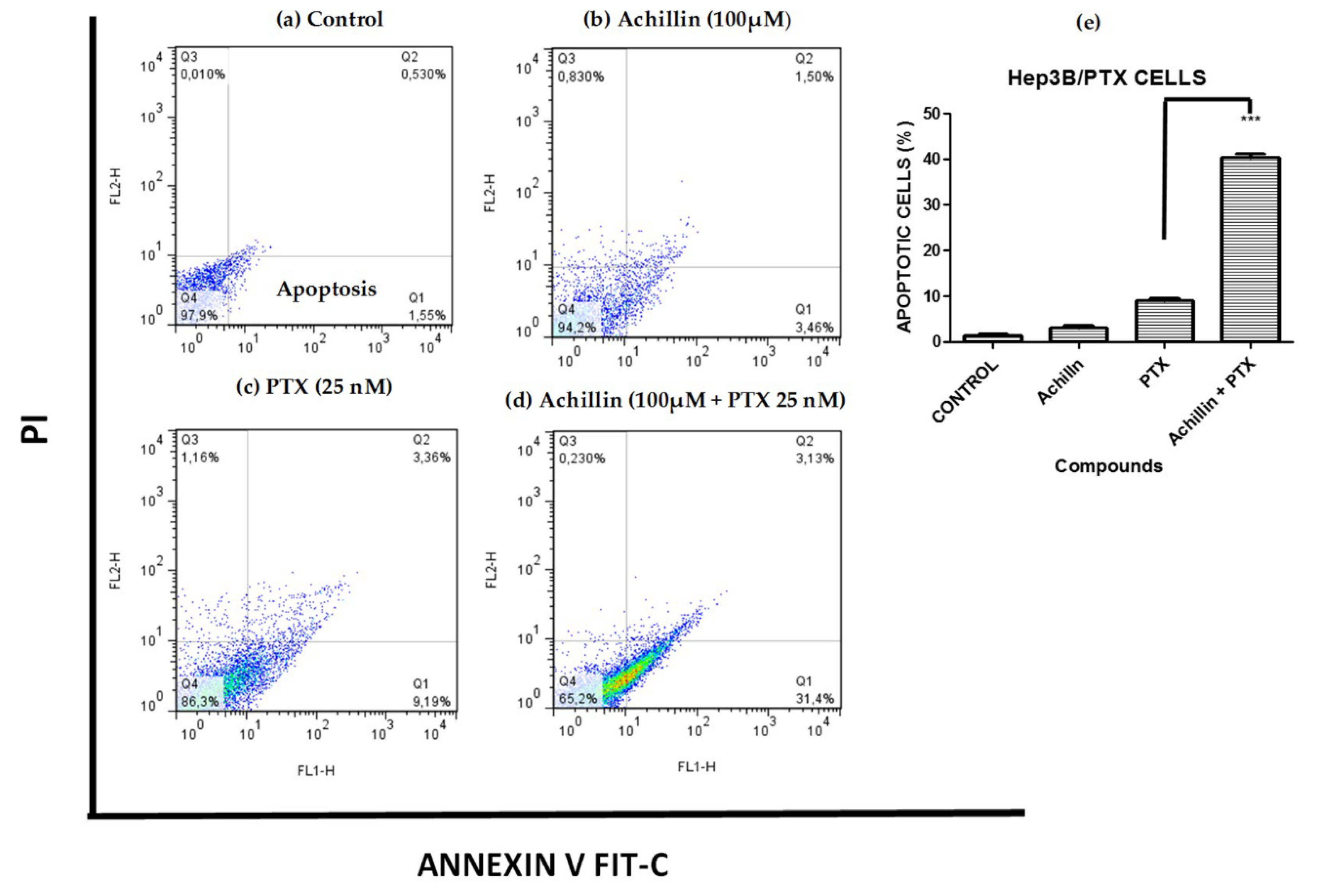
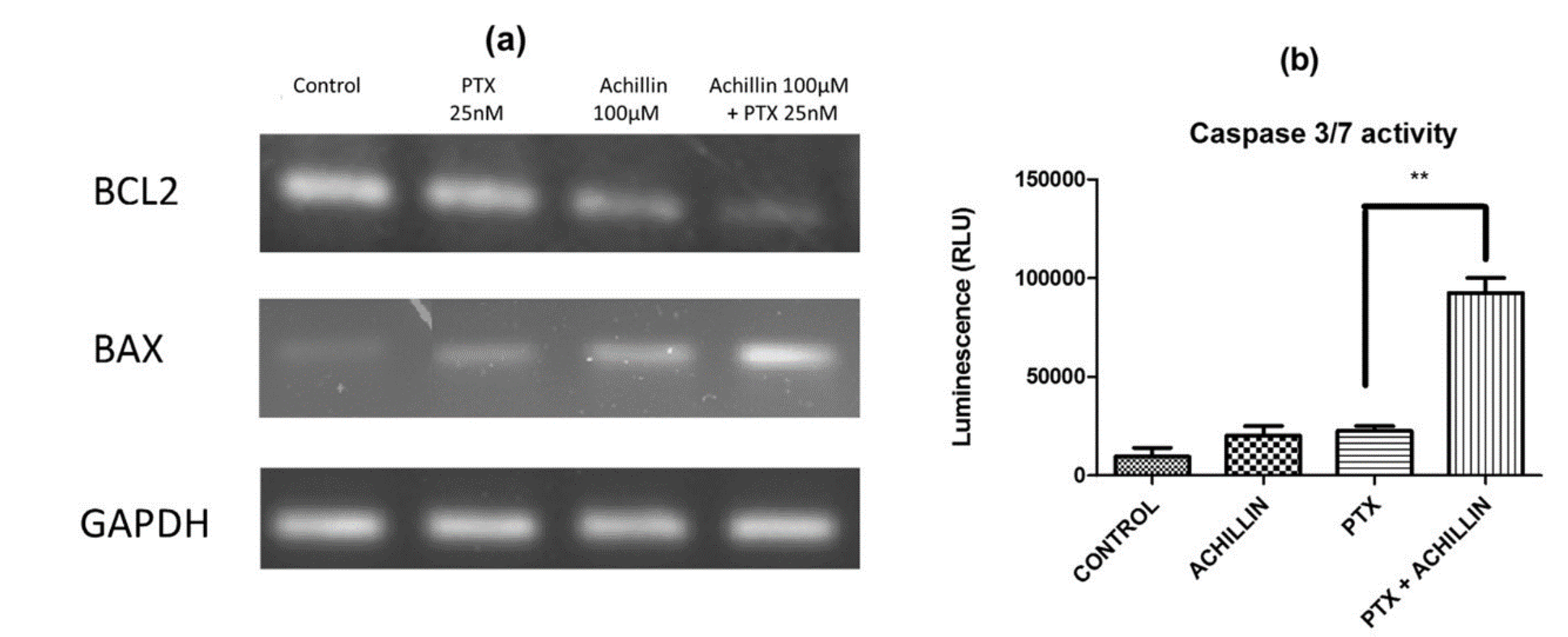
3.4. Effects of Achillin Plus PTX on Cell Death in Hep3B/PTX Cells
3.5. Effect on PTX-Induced Apoptosis
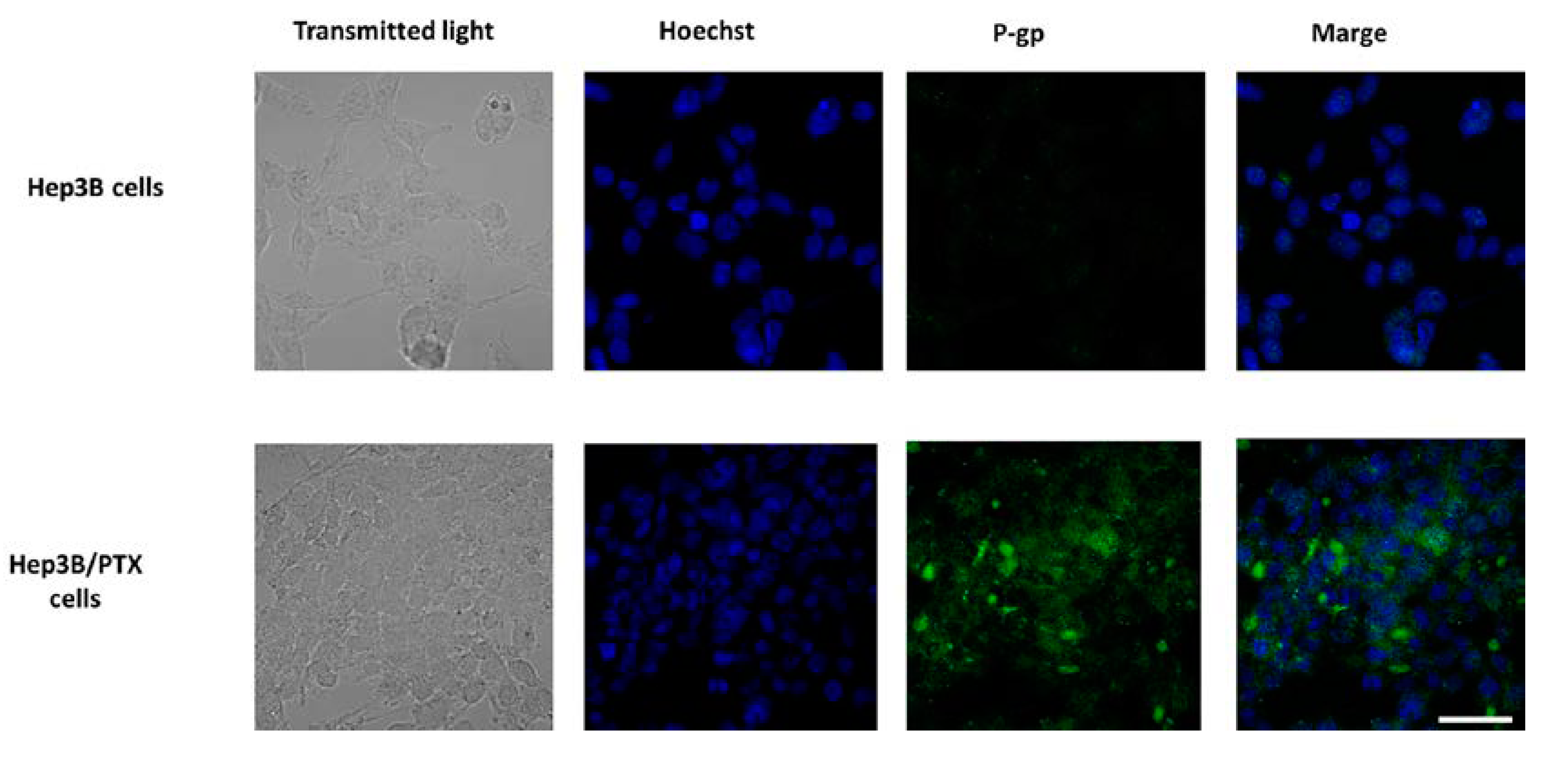
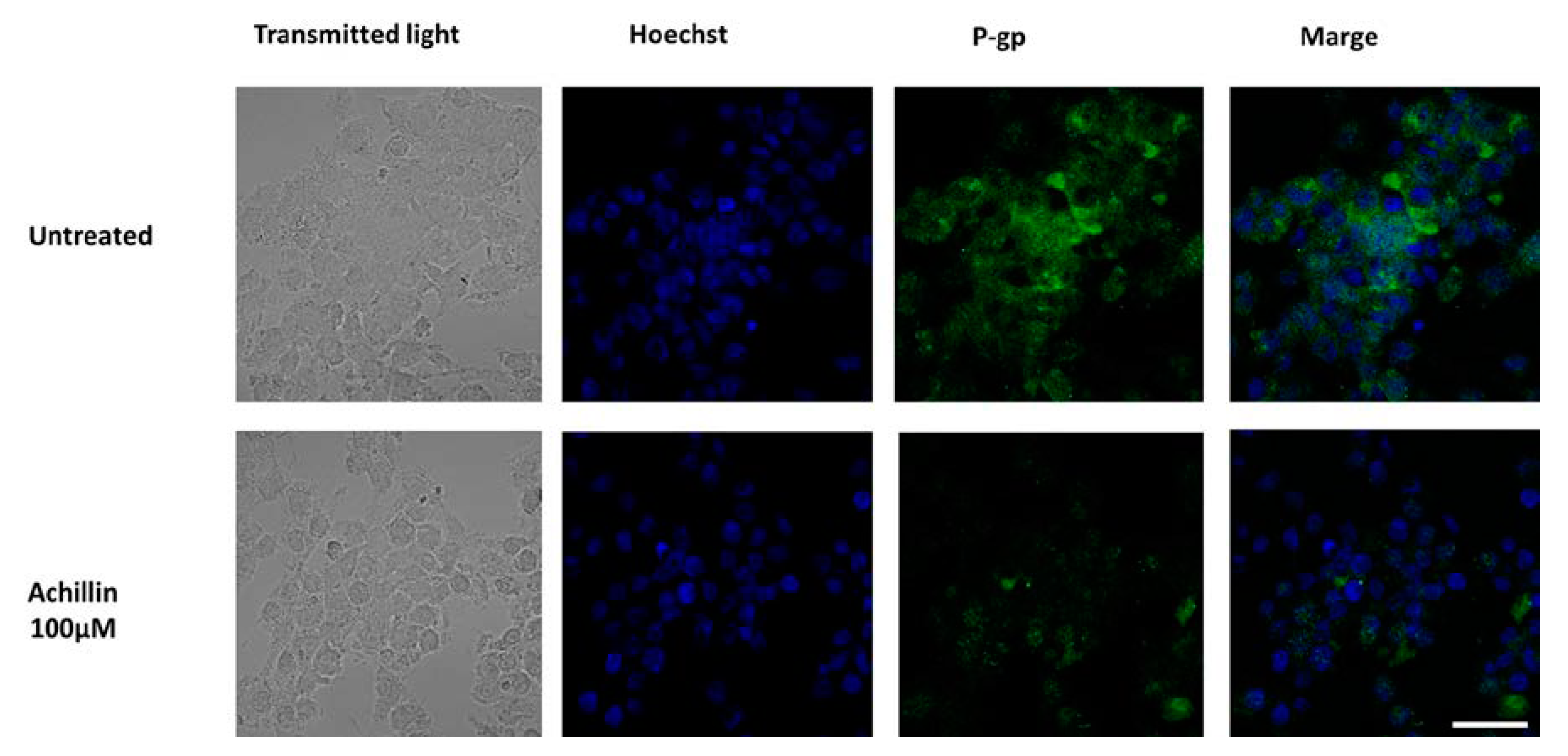
3.6. Effect of Achillin on P-gp Protein Levels in Hep3B/PTX Cells by Immunofluorescence
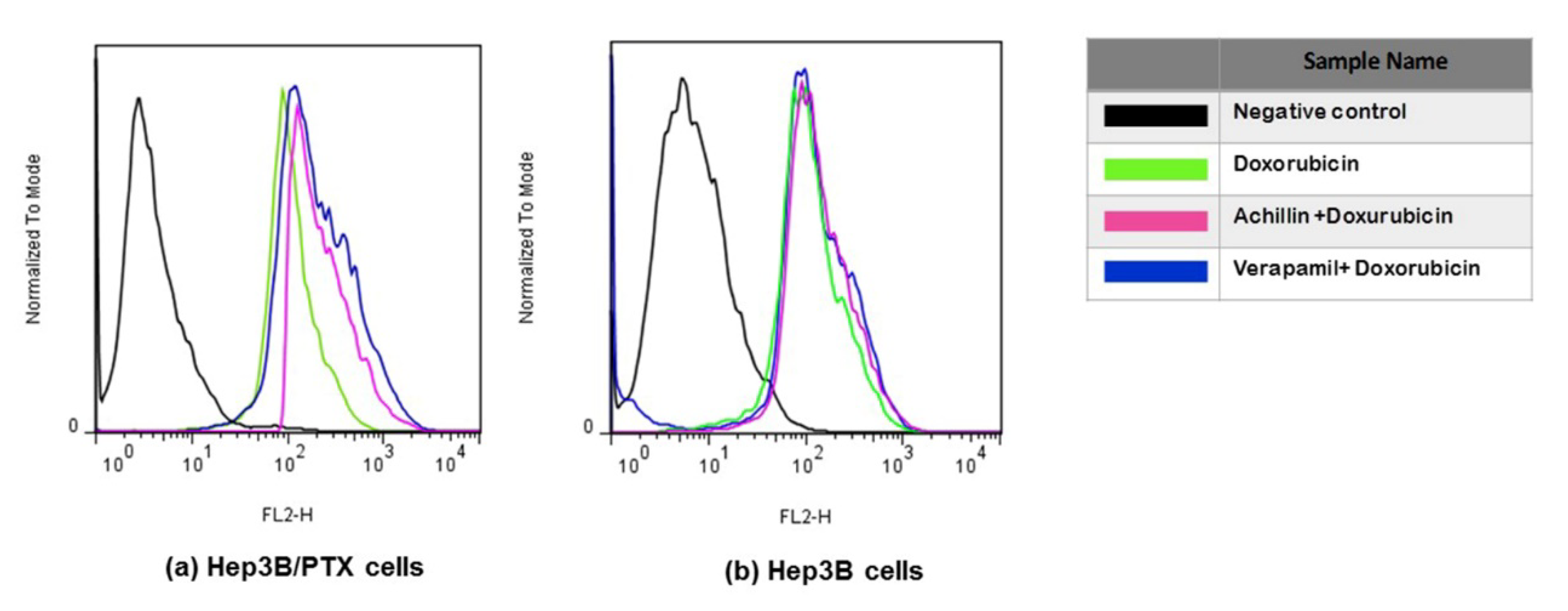
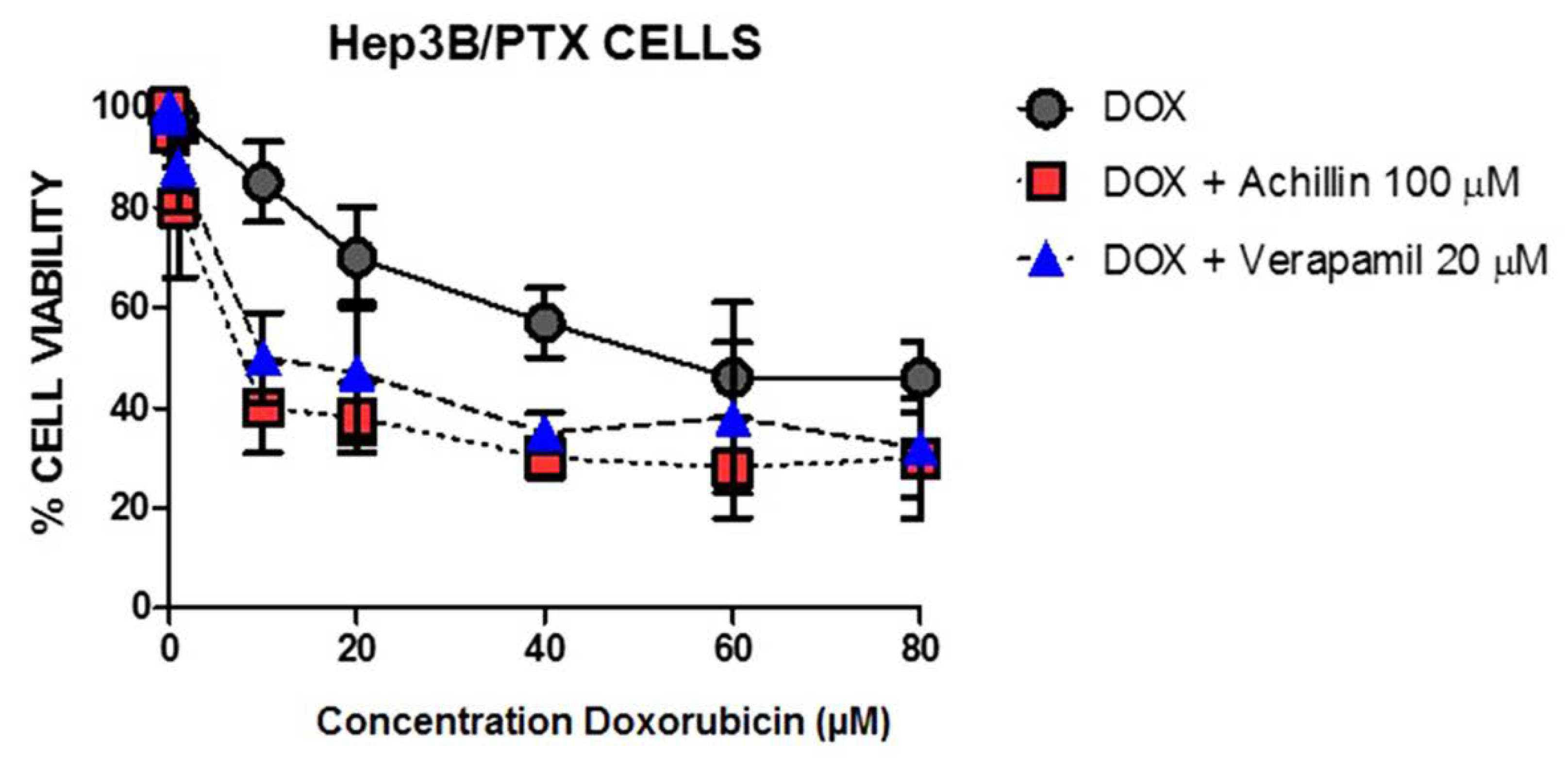
3.7. Effect on Cellular Doxorubicin Retention by Achillin
3.8. Theoretical and Computational Results
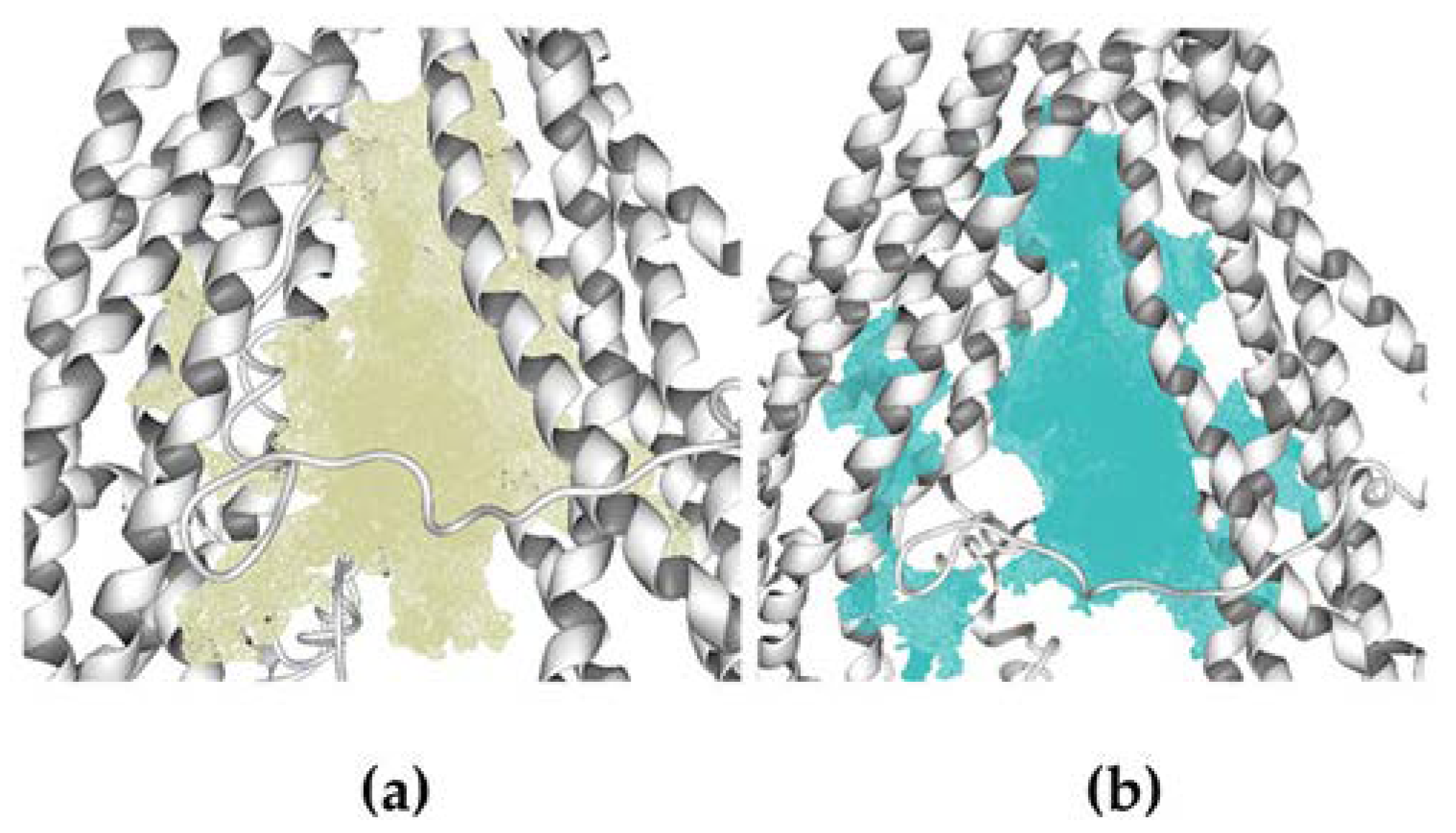
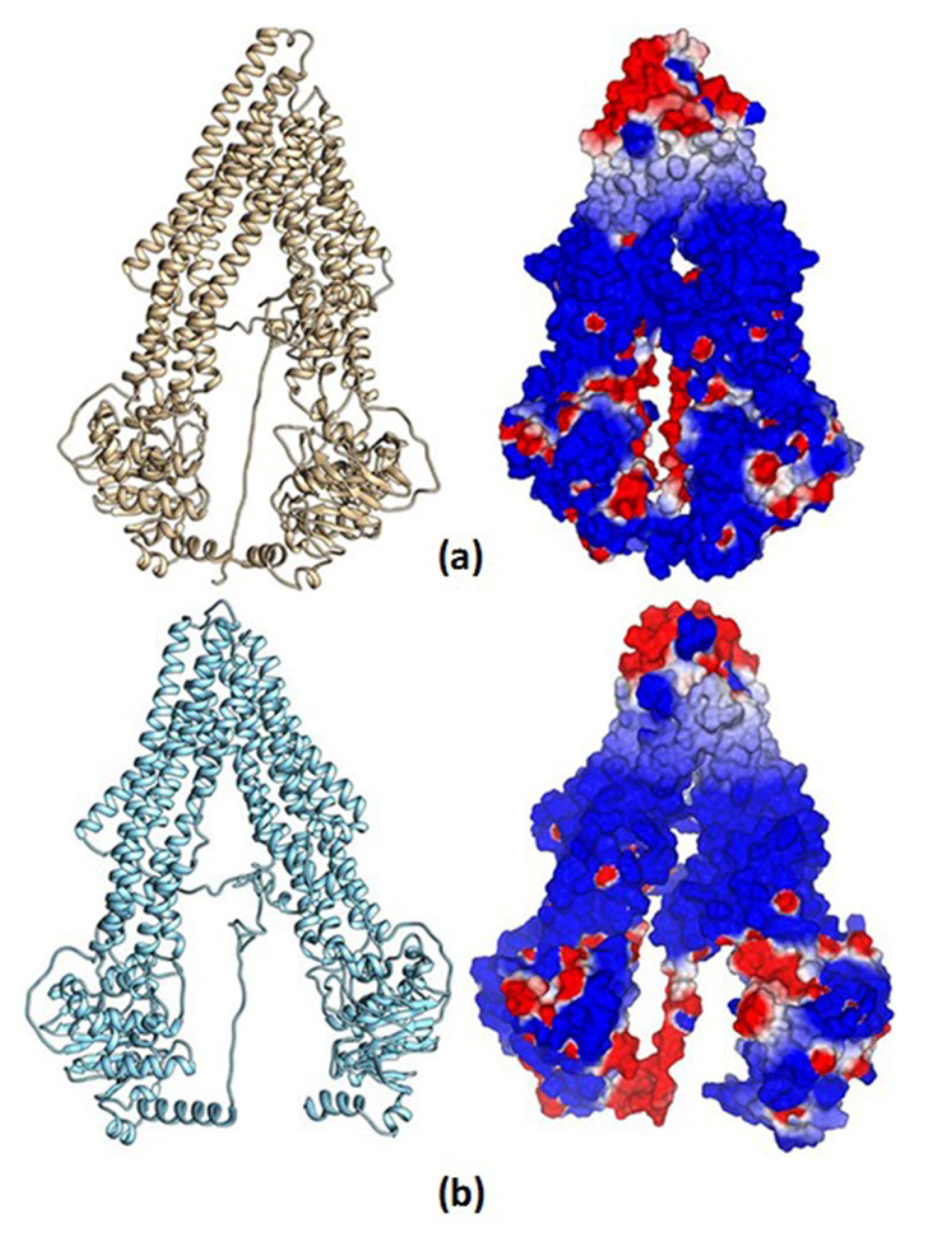
3.8.1. Homology Structural Modeling
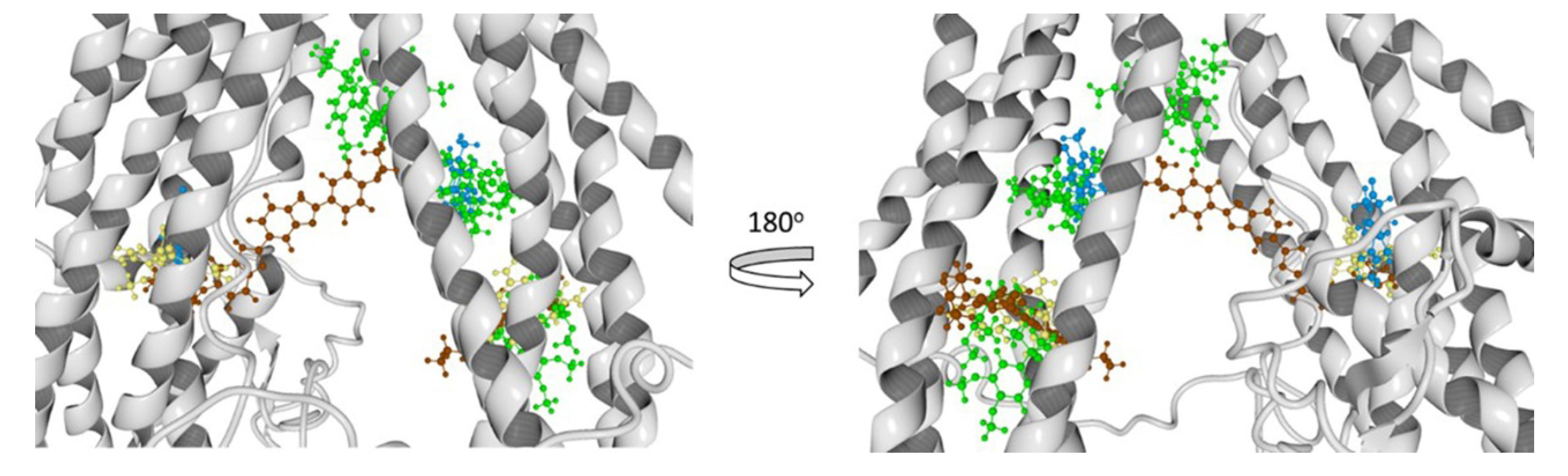
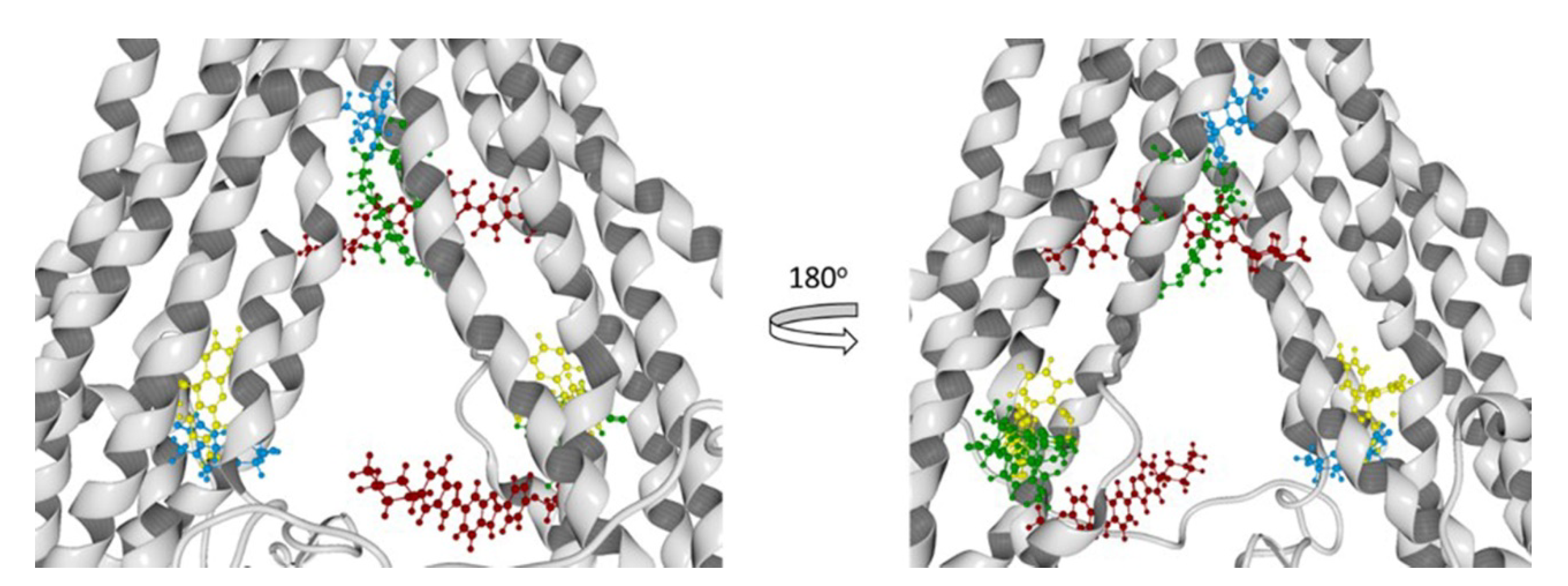
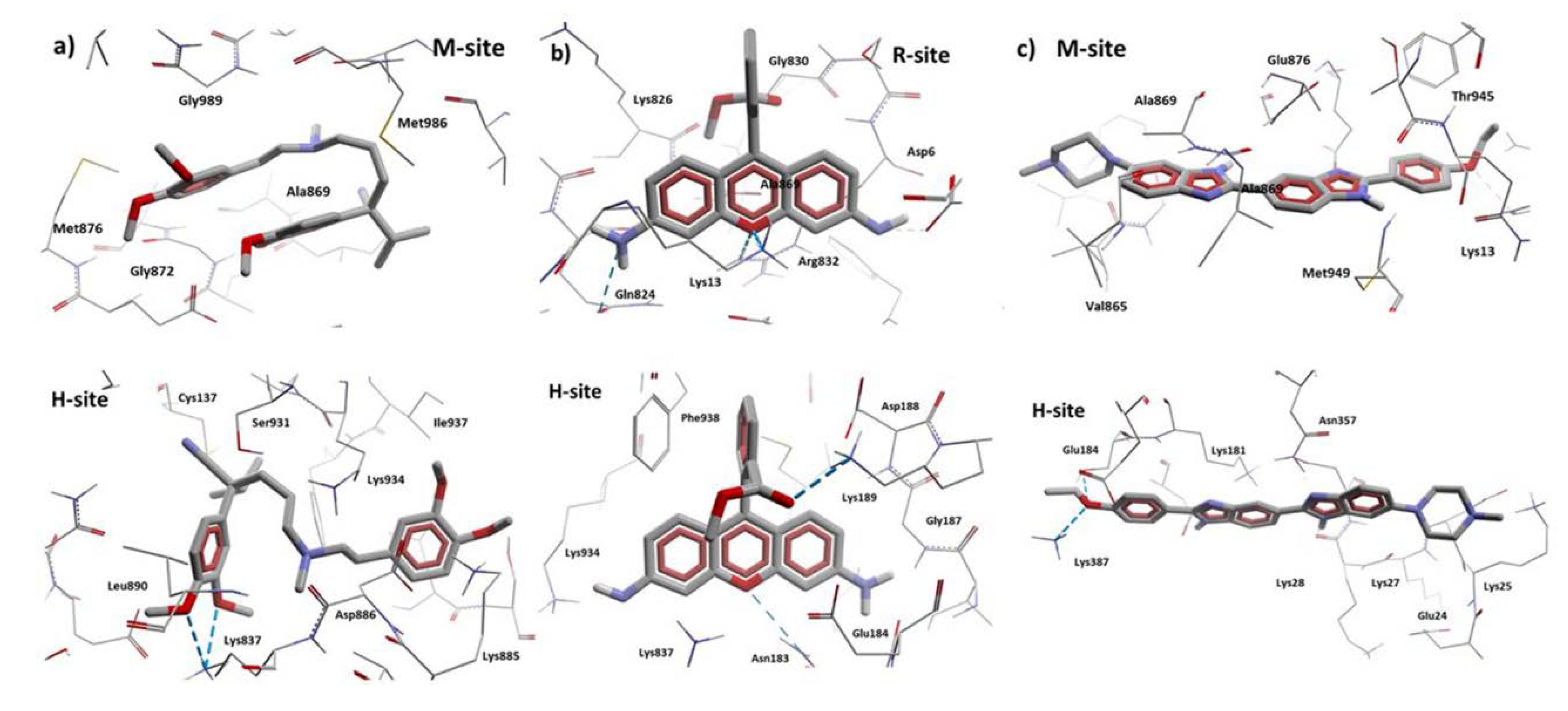
3.8.2. Molecular Docking Calculations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Loong, H.; Yeo, W. Microtubule-targeting agents in oncology and therapeutic potential in hepatocellular carcinoma. Onco Targets Ther. 2014, 7, 575–585. [Google Scholar] [PubMed]
- Nagano, H. Treatment of advanced hepatocellular carcinoma: Intraarterial infusion chemotherapy combined with interferon. Oncology 2010, 78 (Suppl. S1), 142–147. [Google Scholar] [CrossRef]
- Kachalaki, S.; Ebrahimi, M.; Mohamed, K.L.; Mohammadinejad, S.; Baradaran, B. Cancer chemoresistance; biochemical and molecular aspects: A brief overview. Eur. J. Pharm. Sci. 2016, 89, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Pan, C.; Wei, X.; Shi, Y.; Zheng, J.; Lin, X.; Shi, L. lncRNA KRAL reverses 5-fluorouracil resistance in hepatocellular carcinoma cells by acting as a ceRNA against miR-141. Cell Commun. Signal 2018, 16, 47. [Google Scholar] [CrossRef]
- Li, B.; Zhao, S.; Geng, R.; Huo, Z.; Zhang, H. The Sineoculis Homeobox Homolog 1 (SIX1) Gene Regulates Paclitaxel Resistance by Affecting Reactive Oxygen Species and Autophagy in Human Hepatocellular Carcinoma Cell Line HepG2. Med. Sci. Monit. 2018, 24, 2271–2279. [Google Scholar] [CrossRef]
- Huang, H.; Chen, J.; Ding, C.M.; Jin, X.; Jia, Z.M.; Peng, J. LncRNA NR2F1-AS1 regulates hepatocellular carcinoma oxaliplatin resistance by targeting ABCC1 via miR-363. J. Cell. Mol. Med. 2018, 22, 3238–3245. [Google Scholar] [CrossRef]
- Salehan, M.R.; Morse, H.R. DNA damage repair and tolerance: A role in chemotherapeutic drug resistance. Br. J. Biomed. Sci. 2013, 70, 1–40. [Google Scholar] [CrossRef]
- Li, J.; Duan, B.; Guo, Y.; Zhou, R.; Sun, J.; Bie, B.; Yang, S.; Huang, C.; Yang, J.; Li, Z. Baicalein sensitizes hepatocellular carcinoma cells to 5-FU and Epirubicin by activating apoptosis and ameliorating P-glycoprotein activity. Biomed. Pharmacother. 2018, 98, 806–812. [Google Scholar] [CrossRef]
- Dong, J.; Zhai, B.; Sun, W.; Hu, F.; Cheng, H.; Xu, J. Activation of phosphatidylinositol 3-kinase/AKT/snail signaling pathway contributes to epithelial-mesenchymal transition-induced multi-drug resistance to sorafenib in hepatocellular carcinoma cells. PLoS ONE 2017, 21, e0185088. [Google Scholar] [CrossRef]
- Szakacs, G.; Paterson, J.K.; Ludwig, J.A.; Booth-Genthe, C.; Gottesman, M.M. Targeting multidrug resistance in cancer. Nat. Rev. Drug Discov. 2006, 5, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, E.K.; Erba, P.; Mariani, G.; Gomes, C.M. Multidrug resistance in cancer: Its mechanism and its modulation. Drug News Perspect. 2007, 20, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.H. ABC transporters as multidrug resistance mechanisms and the development of chemosensitizers for their reversal. Cancer Cell Int. 2005, 5, 30. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ozben, T. Mechanisms and strategies to overcome multiple drug resistance in cancer. FEBS Lett. 2006, 580, 2903–2909. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Lv, L.; Shen, Y.; Hu, Z.; He, Q.; Chen, X. A nanoparticulate pre-chemosensitizer for efficacious chemotherapy of multidrug resistant breast cancer. Sci. Rep. 2016, 6, 21459. [Google Scholar] [CrossRef]
- Kathawala, R.J.; Gupta, P.; Ashby, C.R.; Chen, Z.-S. The modulation of ABC transporter-mediated multidrug resistance in cancer: A review of the past decade. Drug Resist. Updat 2015, 18, 1–17. [Google Scholar] [CrossRef]
- Gottesman, M.M.; Pastan, I.H. The Role of Multidrug Resistance Efflux Pumps in Cancer: Revisiting a JNCI Publication Exploring Expression of the MDR1 (P-glycoprotein) Gene. J. Natl. Cancer Inst. 2015, 107, djv222. [Google Scholar] [CrossRef]
- Cort, A.; Ozben, T. Natural product modulators to overcome multidrug resistance in cancer. Nutr. Cancer 2015, 67, 411–423. [Google Scholar] [CrossRef]
- Reyes, C.P.; Muñoz-Martinez, F.; Torrecillas, I.R.; Mendoza, C.R.; Gamarro, F.; Bazzocchi, I.L. Biological evaluation, structure-activity relationships, and three-dimensional quantitative structure-activity relationship studies of dihydrob-β-agarofuran sesquiterpenes as modulators of P-glycoprotein dependent multidrug resistance. J. Med. Chem. 2007, 50, 4808–4817. [Google Scholar] [CrossRef]
- Steglich, B.; Mahringer, A.; Li, Y.; Posner, G.H.; Fricker, G.; Efferth, T. Inhibition of P-glycoprotein by two artemisinin derivatives. Nat. Prod. Bioprosp. 2012, 2, 59–64. [Google Scholar] [CrossRef][Green Version]
- Wink, M.; Ashour, M.L.; El-Readi, M.Z. Secondary metabolites from plants inhibiting ABC transporters and reversing resistance of cancer cells and microbes to cytotoxic and antimicrobial agents. Front. Microbiol. 2012, 3, 130. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.L.; Rossi, R.M.; Karnischky, L.; Li, X.; Peterson, D.R.; Howard, D.S.; Jordan, C.T. The sesquiterpene lactone parthenolide induces apoptosis of human acute myelogenous leukemia stem and progenitor cells. Blood 2005, 105, 4163–4169. [Google Scholar] [CrossRef] [PubMed]
- Chadwick, M.; Trewin, H.; Gawthrop, F.; Wagstaff, C. Sesquiterpenoids Lactones: Benefits to Plants and People. Int. J. Mol. Sci. 2013, 14, 12780–12805. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, C.J.; Mehrotra, S.; Sadaria, M.R.; Kumar, S.; Shortle, N.H.; Roman, Y.; Sheridan, C.; Campbell, R.A.; Murry, D.J.; Badve, S.; et al. The sesquiterpene lactone parthenolide in combination with docetaxel reduces metastasis and improves survival in a xenograft model of breast cancer. Mol. Cancer Ther. 2005, 4, 1004–1012. [Google Scholar] [CrossRef] [PubMed]
- Fraga, B.M. Natural sesquiterpenoids. Nat. Prod. Rep. 2011, 28, 1580–1610. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.; Muñoz-Zamora, A.; Nathan, P. Conformational analysis of achillin and leukodin. J. Nat. Prod. 1988, 51, 221–228. [Google Scholar] [CrossRef]
- White, E.H.; Winter, R.E.K. Natural Products from Achillea lanulosa. Tetrahedron Lett. 1963, 4, 137–140. [Google Scholar] [CrossRef]
- Balboul, B.A.; Ahmed, A.A.; Otsuka, H.; Bando, M.; Kido, M.; Takeda, Y. A guaianolide and a germacranolide from Achillea santolina. Phytochemistry 1997, 46, 1045–1049. [Google Scholar] [CrossRef]
- Basu, A.; Saito, K.; Meyer, K.; Ray, R.B. Stellate cell apoptosis by a soluble mediator from immortalized human hepatocytes. Apoptosis 2006, 11, 1391–1400. [Google Scholar] [CrossRef]
- Aszalos, A.; Thompson, K.; Yin, J.J.; Ross, D.D. Combinations of P-glycoprotein blockers, verapamil, PSC833, and cremophor act differently on the multidrug resistance associated protein (MRP) and on P-glycoprotein (P-gP). Anticancer Res. 1999, 19, 1053–1064. [Google Scholar]
- Krstin, S.; Peixoto, H.S.; Wink, M. Combinations of alkaloids affecting different molecular targets with the saponin digitonin can synergistically enhance trypanocidal activity against Trypanosoma brucei brucei. Antimicrob. Agents Chemother. 2015, 59, 7011–7017. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C. Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2000, 9, 40. [Google Scholar] [CrossRef]
- Kim, N.; Shin, J.-M.; No, K.T. In silico study on the interaction between P-glycoprotein and its inhibitors at the drug binding. Bull. Korean Chem. Soc. 2014, 35, 2317–2325. [Google Scholar] [CrossRef][Green Version]
- Syed, S.B.; Arya, H.; Fu, I.H.; Yeh, T.K.; Periyasamy, L.; Hsieh, H.P.; Coumar, M.S. Targeting P-glycoprotein: Investigation of piperine analogs for overcoming drug resistance in cancer. Sci Rep. 2017, 7, 7972. [Google Scholar] [CrossRef]
- Wavefunction, Inc. Spartan’18. Available online: https://www.wavefun.com/ (accessed on 27 November 2017).
- Yang, J.M.; Chen, C.-C. GEMDOCK: A generic evolutionary method for molecular docking. Proteins Struct. Funct. Bioinform. 2004, 55, 288–304. [Google Scholar] [CrossRef]
- Thomsen, R.; Christensen, M.H. A New Technique for High-Accuracy Molecular Docking. J. Med. Chem. 2006, 49, 3315–3321. [Google Scholar] [CrossRef]
- Ferreira, R.J.; Ferreira, M.-J.U.; dos Santos, D.J. Molecular Docking Characterizes Substrate-Binding Sites and Efflux Modulation Mechanisms within P-Glycoprotein. J. Chem. Inf. Model. 2013, 53, 1747–1760. [Google Scholar] [CrossRef]
- Weaver, B.A. How Taxol/paclitaxel kills cancer cells. Mol. Biol. Cell 2014, 25, 2677–2681. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.H.; Kim, C.G.; Lim, Y.; Shin, S.Y.; Lee, Y.H. Curcumin down-regulates the multidrug-resistance mdr1b gene by inhibiting the PI3K/Akt/NF kappa B pathway. Cancer Lett. 2008, 259, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Yang, X.; Zhao, L.; Wang, X.; Liu, L.; Jiao, W.; Shan, B. Celecoxib enhances sensitivity to chemotherapy drugs of T-cell lymphoma. Oncol. Lett. 2018, 15, 4649–4656. [Google Scholar] [CrossRef] [PubMed]
- Follit, C.A.; Brewer, F.K.; Wise, J.G.; Vogel, P.D. In silico identified targeted inhibitors of P-glycoprotein overcome multidrug resistance in human cancer cells in culture. Pharmacol. Res. Perspect. 2015, 3, e00170. [Google Scholar] [CrossRef]
- Wessler, J.D.; Grip, L.T.; Mendell, J.; Giugliano, R.P. The p-glycoprotein transport system and cardiovascular drugs. J. Am. Coll. Cardiol. 2013, 61, 2495–2502. [Google Scholar] [CrossRef]
- Fang, L.J.; Shao, X.T.; Wang, S.; Lu, G.H.; Xu, T.; Zhou, J.Y. Sesquiterpene lactone parthenolide markedly enhances sensitivity of human A549 cells to low-dose oxaliplatin via inhibition of NF-kappaB activation and induction of apoptosis. Planta Med. 2010, 76, 258–264. [Google Scholar] [CrossRef]
- Van, C.; Boot, A.; Corver, W.E.; van Eendenburg, J.D.; Trimbos, B.J.; van Wezel, T. Synergistic effects of the sesquiterpene lactone, EPD, with cisplatin and paclitaxel in ovarian cancer cells. J. Exp. Clin. Cancer Res. 2015, 34, 38. [Google Scholar]
- Li, C.; Wu, H.; Yang, Y.; Liu, J.; Chen, Z. Sesquiterpene lactone 6-O-angeloylplenolin reverses vincristine resistance by inhibiting YB-1 nuclear translocation in colon carcinoma cells. Oncol. Lett. 2018, 15, 9673–9680. [Google Scholar] [CrossRef]
- Nabekura, T. Overcoming multidrug resistance in human cancer cells by natural compounds. Toxins 2010, 2, 1207–1224. [Google Scholar] [CrossRef]
- List, A.F.; Kopecky, K.J.; Willman, C.L.; Head, D.R.; Persons, D.L.; Slovak, M.L.; Dorr, R.; Karanes, C.; Hynes, H.E.; Doroshow, J.H.; et al. Benefit of cyclosporine modulation of drug resistance in patients with poor-risk acute myeloid leukemia: A Southwest Oncology Group study. Blood 2001, 98, 3212–3220. [Google Scholar] [CrossRef]
- Tsuruo, T.; Iida, H.; Tsukagoshi, S.; Sakurai, Y. Overcoming of vincristine resistance in P388 leukemia in vivo and in vitro through enhanced cytotoxicity of vincristine and vinblastine by verapamil. Cancer Res. 1981, 41, 1967–1972. [Google Scholar] [PubMed]
- Ford, J.M. Modulators of multidrug resistance. Preclinical studies. Hematol. Oncol. Clin. N. Am. 1995, 9, 337–361. [Google Scholar] [CrossRef]
- Lampidis, T.J.; Krishan, A.; Planas, L.; Tapiero, H. Reversal of intrinsic resistance to adriamycin in normal cells by verapamil. Cancer Drug Deliv. 1986, 3, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H.; Yu, A.M. ABC transporters in multidrug resistance and pharmacokinetics, and strategies for drug development. Curr. Pharm. Des. 2014, 20, 793–807. [Google Scholar] [CrossRef] [PubMed]
- Yutong, H.; Chuangang, L.; Hong, L.; Molin, L.; Xiaohong, S. Resveratrol-Mediated Reversal of Tumor Multi-Drug Resistance. Curr. Drug Metab. 2014, 15, 703–710. [Google Scholar]
- Cheng, G.; Xie, L. Parthenolide induces apoptosis and cell cycle arrest of human 5637 bladder cancer cells in vitro. Molecules 2011, 16, 6758–6768. [Google Scholar] [CrossRef]
- Dantzic, D.; Noel, P.; Merien, F.; Liu, D.-X.; Lu, J.; Han, H.; McKeage, M.J.; Li, Y. The Effects of Synthetically Modified Natural Compounds on ABC Transporters. Pharmaceutics 2018, 10, 127. [Google Scholar] [CrossRef]
- Jouan, E.; Le, V.M.; Mayati, A.; Denizot, C.; Parmentier, Y.; Fardel, O. Evaluation of P-Glycoprotein Inhibitory Potential Using a Rhodamine 123 Accumulation Assay. Pharmaceutics 2016, 8, 12. [Google Scholar] [CrossRef]
- Loo, T.W.; Bartlett, M.C.; Clarke, D.M. Substrate-induced conformational changes in the transmembrane segments of human P-glycoprotein. J. Biol. Chem. 2003, 278, 13603–13606. [Google Scholar] [CrossRef]
- Wang, Y.; Hao, D.; Stein, W.D.; Yang, L. A kinetic study of Rhodamine123 pumping by P-glycoprotein. Biochim. Biophys. Acta 2006, 1758, 1671–1676. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
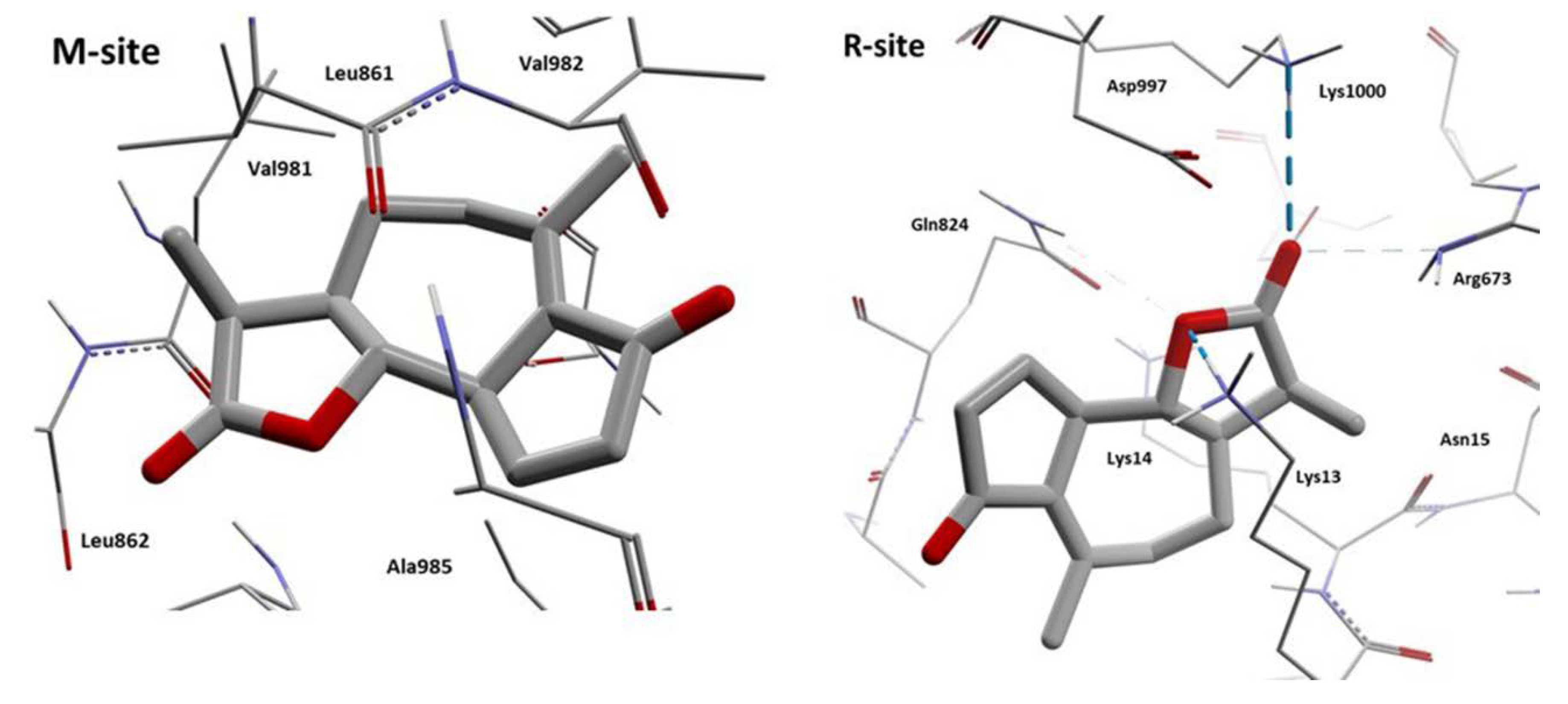{kind=link}
| PTX IC50 nM | |
|---|---|
| PTX/Hep3B | Hep3B |
| 352 ± 12 a | 25 ± 5 a |
| Treatment | PTX aIC50 nM (RRI) b Hep3B Cells | PTX aIC50 nM (RRI) b PTX/Hep3B Cells |
|---|---|---|
| PTX alone | 25 ± 5 | 352 ± 12 a |
| PTX + Achillin 100 µM | 19.5 ± 3 (1.28) | 38 ± 5 (9.1) |
| PTX + Verapamil 20 µM | 21.4 ± 4.2 (1.16) | 33 ± 10 (10) |
| Treatment | IC50 PTX | DRI | CI | Interpretation |
|---|---|---|---|---|
| PTX alone | 0.352 ± 0.012 | 1 | NR | NR |
| PTX + Achillin | - | - | - | - |
| 100 µM (IC20) | 0.038 ± 0.0091 | 9.2 | 0.5 | +++ |
| 160 µM (IC40) | 0.03 ± 0.007 | 7.3 | 0.88 | ++ |
| Molecule | M-site | R-site | H-site |
|---|---|---|---|
| Hoechst 33342 | Thr945, Ala871, Met949, Glu875, Phe942, His61, Ala869, Gly989, Val991, Gln195, Met948, Val873, Gly872, Val865, Ser992, Ala947, Ile870, Val988, Leu843, Gln132, Met876, Ile868, Gln946 | - | Glu29, Lys181, Asp177, Lys25, Asp26, Lys887, Lys28, Asn357, Glu184, Glu24, Lys27, Leu890, Asn183 |
| Verapamil | Met986, Glu875, Met949, Gly989, Ile868, Met876, Ala985, Val982, Gly872, Val988, Ala869, Ala871, Val873, Ser952, Gln990, Leu3, Val865 | - | Asp886, Lys934, Phe938, Ser931, Lys887, Glu184, Ala883, Cys137, Asn183, Leu879, Lys885, Ala935, Ser880, Ile937, Leu890, Asn930, Gln882, Gly141, Asp188, Ser180 |
| Rhodamine123 | - | Lys13, Asp997, Gln824, Ser831, Asp6, Ala823, Phe994, Gly9, Gly827, Leu236, Ile829, Ala828, Leu833, Arg673, Lys14, Gly830, Lys826 | Glu184, Asp188, Lys189, Phe938, Lys877, Lys887, Cys137, Lys934, Asn183, Ile186, Asp886, Ser931, Ala140, Gly187, Val133, Val185, Ala883 |
| Achillin | Leu861, Val982, Phe978, Leu862, Ile868, Ala985, Ser952, Ile847, Thr858, Phe983, Met986, Val865, Val981 | Gln824, Lys13, Lys14, Ala823, Lys826, Asp679, Asp997, Arg673, Lys1000, Asn15, Phe16, Phe17, Ala828, Gly10, Gly827 | - |
| Molecule | M-site | R-site | H-site |
|---|---|---|---|
| Hoechst 33342 | Einter = −81.53 kcal/mol LE = −2.40 kcal/atom | - | Einter = −83.17 kcal/mol LE = −2.45 kcal/atom |
| Verapamil | Einter = −99.18 kcal/mol LE = −3.00 kcal/atom | - | Einter = −110.04 kcal/mol LE = −3.33 kcal/atom |
| Rhodamine123 | - | Einter = −71.97 kcal/mol LE = −2.77 kcal/atom | Einter = −41.22 kcal/mol LE = −1.59 kcal/atom |
| Achillin | Einter = −53.04 kcal/mol LE = −3.12 kcal/atom | Einter = −84.93 kcal/mol LE = −4.99 kcal/atom | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanchez-Carranza, J.N.; González-Maya, L.; Razo-Hernández, R.S.; Salas-Vidal, E.; Nolasco-Quintana, N.Y.; Clemente-Soto, A.F.; García-Arizmendi, L.; Sánchez-Ramos, M.; Marquina, S.; Alvarez, L. Achillin Increases Chemosensitivity to Paclitaxel, Overcoming Resistance and Enhancing Apoptosis in Human Hepatocellular Carcinoma Cell Line Resistant to Paclitaxel (Hep3B/PTX). Pharmaceutics 2019, 11, 512. https://doi.org/10.3390/pharmaceutics11100512
Sanchez-Carranza JN, González-Maya L, Razo-Hernández RS, Salas-Vidal E, Nolasco-Quintana NY, Clemente-Soto AF, García-Arizmendi L, Sánchez-Ramos M, Marquina S, Alvarez L. Achillin Increases Chemosensitivity to Paclitaxel, Overcoming Resistance and Enhancing Apoptosis in Human Hepatocellular Carcinoma Cell Line Resistant to Paclitaxel (Hep3B/PTX). Pharmaceutics. 2019; 11(10):512. https://doi.org/10.3390/pharmaceutics11100512
Chicago/Turabian StyleSanchez-Carranza, Jessica Nayelli, Leticia González-Maya, Rodrigo Said Razo-Hernández, Enrique Salas-Vidal, Ninfa Yaret Nolasco-Quintana, Aldo F. Clemente-Soto, Lucero García-Arizmendi, Mariana Sánchez-Ramos, Silvia Marquina, and Laura Alvarez. 2019. "Achillin Increases Chemosensitivity to Paclitaxel, Overcoming Resistance and Enhancing Apoptosis in Human Hepatocellular Carcinoma Cell Line Resistant to Paclitaxel (Hep3B/PTX)" Pharmaceutics 11, no. 10: 512. https://doi.org/10.3390/pharmaceutics11100512
APA StyleSanchez-Carranza, J. N., González-Maya, L., Razo-Hernández, R. S., Salas-Vidal, E., Nolasco-Quintana, N. Y., Clemente-Soto, A. F., García-Arizmendi, L., Sánchez-Ramos, M., Marquina, S., & Alvarez, L. (2019). Achillin Increases Chemosensitivity to Paclitaxel, Overcoming Resistance and Enhancing Apoptosis in Human Hepatocellular Carcinoma Cell Line Resistant to Paclitaxel (Hep3B/PTX). Pharmaceutics, 11(10), 512. https://doi.org/10.3390/pharmaceutics11100512