
Type W Human Endogenous Retrovirus (HERV-W) Integrations and Their Mobilization by L1 Machinery: Contribution to the Human Transcriptome and Impact on the Host Physiopathology



Abstract
1. Introduction
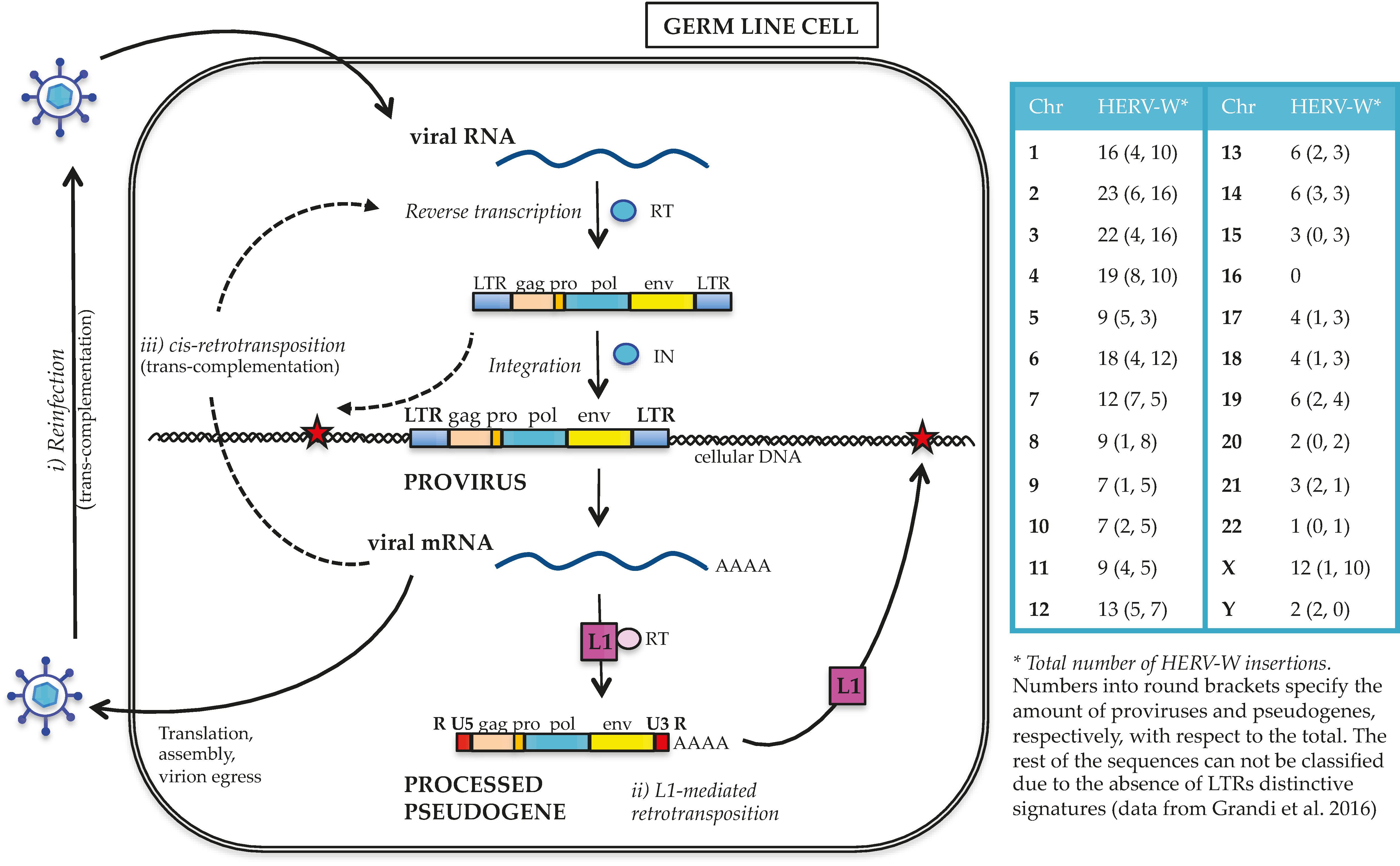
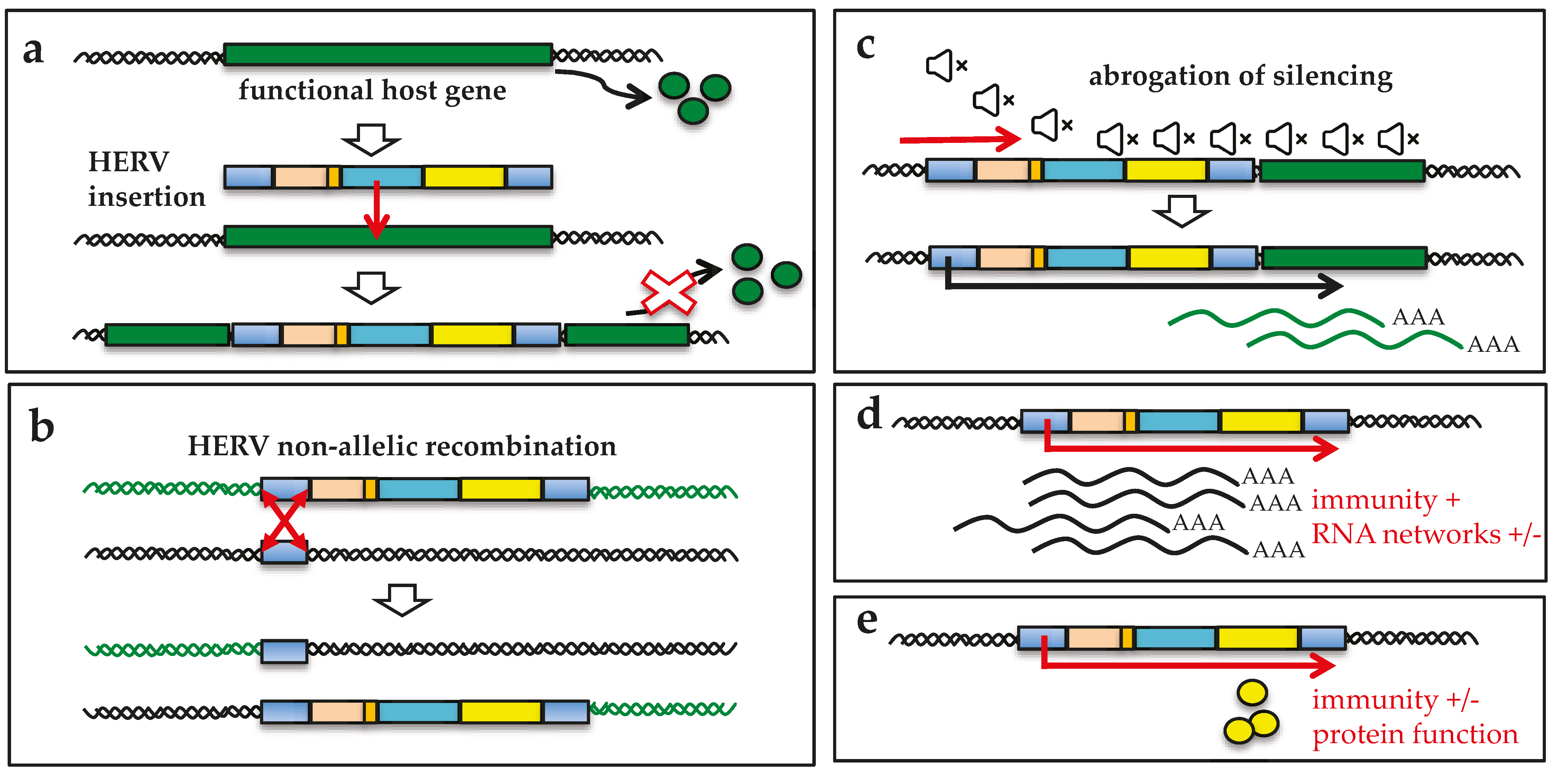
2. HERV-W Group Contribution to the Human Genome
3. HERV-W Placental Expression and Syncytin-1 Env Protein Cooption for Human Physiology
4. General HERV-W Expression in Healthy Tissues Other than the Placenta
5. Syncytin-1 Expression in Placental Pathologies
6. HERV-W Expression in Tumorigenesis and Cancer Progression
7. HERV-W Expression in MS and Other Autoimmune Diseases
7.1. Multiple Sclerosis
7.1.1. Detection of HERV-W/MSRV Nucleic Acids in MS Samples
7.1.2. Presence of HERV-W/MSRV Ags in MS Lesions
7.1.3. Onset of an Immune Response against These Elements
7.1.4. Use of Some Animal Models of MS
7.2. Other Autoimmune Diseases
8. HERV-W Expression in Neurological and Neuropsychiatric Disorders
9. HERV-W Expression in the Presence of Exogenous Infections
9.1. Retroviral Infections
9.2. Herpesviral Infections
9.3. Other Exogenous Infections
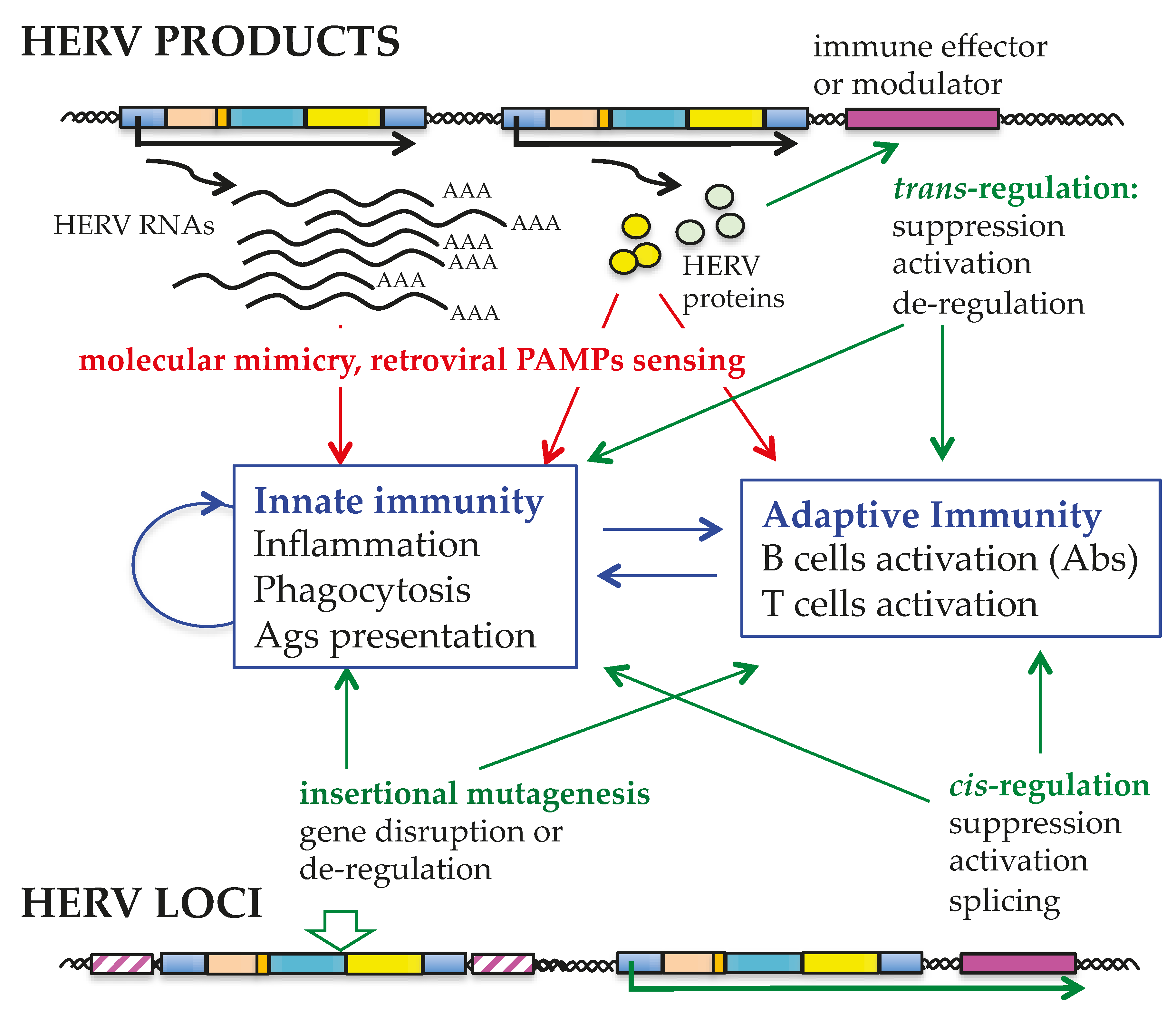
10. The HERV-W Transcriptional Landscape in the Context of Human Physiopathology: Current Needs and Future Perspectives
Supplementary Materials
Acknowledgments
Conflicts of Interest
References
- International Human Genome Sequencing Consortium International Human Genome Sequencing Consortium. Finishing the euchromatic sequence of the human genome. Nature 2004, 431, 931–945. [Google Scholar]
- Cordaux, R.; Batzer, M.A. The impact of retrotransposons on human genome evolution. Nat. Rev. Genet. 2009, 10, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, K. RNA-Mediated Gene Duplication and Retroposons: Retrogenes, LINEs, SINEs, and Sequence Specificity. Int. J. Evol. Biol. 2013, 2013, 424726. [Google Scholar] [CrossRef] [PubMed]
- Bannert, N.; Kurth, R. The evolutionary dynamics of human endogenous retroviral families. Annu. Rev. Genom. Hum. Genet. 2006, 7, 149–173. [Google Scholar] [CrossRef] [PubMed]
- Mager, D.L.; Goodchild, N.L. Homologous recombination between the LTRs of a human retrovirus-like element causes a 5-kb deletion in two siblings. Am. J. Hum. Genet. 1989, 45, 848–854. [Google Scholar] [PubMed]
- Blomberg, J.; Benachenhou, F.; Blikstad, V.; Sperber, G.; Mayer, J. Classification and nomenclature of endogenous retroviral sequences (ERVs): Problems and recommendations. Gene 2009, 448, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Sperber, G.; Airola, T.; Jern, P.; Blomberg, J. Automated recognition of retroviral sequences in genomic data—RetroTector©. Nucleic Acids Res. 2007, 35, 4964–4976. [Google Scholar] [CrossRef] [PubMed]
- Vargiu, L.; Rodriguez-Tomé, P.; Sperber, G.O.; Cadeddu, M.; Grandi, N.; Blikstad, V.; Tramontano, E.; Blomberg, J. Classification and characterization of human endogenous retroviruses; mosaic forms are common. Retrovirology 2016, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Magiorkinis, G.; Belshaw, R.; Katzourakis, A. “There and back again”: Revisiting the pathophysiological roles of human endogenous retroviruses in the post-genomic era. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120504. [Google Scholar] [CrossRef] [PubMed]
- Schön, U.; Seifarth, W.; Baust, C.; Hohenadl, C.; Erfle, V.; Leib-Mösch, C. Cell Type-Specific Expression and Promoter Activity of Human Endogenous Retroviral Long Terminal Repeats. Virology 2001, 279, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Stauffer, Y.; Theiler, G.; Sperisen, P.; Lebedev, Y.; Jongeneel, C.V. Digital expression profiles of human endogenous retroviral families in normal and cancerous tissues. Cancer Immun. Arch. 2004, 4, 2. [Google Scholar]
- Seifarth, W.; Frank, O.; Zeilfelder, U. Comprehensive analysis of human endogenous retrovirus transcriptional activity in human tissues with a retrovirus-specific microarray. J. Virol. 2005, 79, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Flockerzi, A.; Maydt, J.; Frank, O.; Ruggieri, A.; Maldener, E.; Seifarth, W.; Medstrand, P.; Lengauer, T.; Meyerhans, A.; Leib-Mösch, C.; et al. Expression pattern analysis of transcribed HERV sequences is complicated by ex vivo recombination. Retrovirology 2007, 4, 39. [Google Scholar] [CrossRef] [PubMed]
- Hu, L. Endogenous Retroviral RNA Expression in Humans; Uppsala University: Uppsala, Sweden, 2007. [Google Scholar]
- Pérot, P.; Mugnier, N.; Montgiraud, C.; Gimenez, J.; Jaillard, M.; Bonnaud, B.; Mallet, F. Microarray-based sketches of the HERV transcriptome landscape. PLoS ONE 2012, 7, e40194. [Google Scholar]
- Haase, K.; Mösch, A.; Frishman, D. Differential expression analysis of human endogenous retroviruses based on ENCODE RNA-seq data. BMC Med. Genom. 2015, 8, 71. [Google Scholar] [CrossRef] [PubMed]
- Balestrieri, E.; Pica, F.; Matteucci, C.; Zenobi, R.; Sorrentino, R.; Argaw-Denboba, A.; Cipriani, C.; Bucci, I.; Sinibaldi-Vallebona, P. Transcriptional activity of human endogenous retroviruses in human peripheral blood mononuclear cells. Biomed Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Voisset, C.; Weiss, R.A.; Griffiths, D.J. Human RNA “rumor” viruses: The search for novel human retroviruses in chronic disease. Microbiol. Mol. Biol. Rev. 2008, 72, 157–196, table of contents. [Google Scholar] [CrossRef] [PubMed]
- Jern, P.; Coffin, J.M. Effects of Retroviruses on Host Genome Function. Annu. Rev. Genet. 2008, 42, 709–732. [Google Scholar] [CrossRef] [PubMed]
- Christensen, T. Human endogenous retroviruses in neurologic disease. Apmis 2016, 124, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Grandi, N.; Cadeddu, M.; Blomberg, J.; Tramontano, E. Contribution of type W human endogenous retrovirus to the human genome: Characterization of HERV-W proviral insertions and processed pseudogenes. Retrovirology 2016, 13, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Perron, H.; Garson, J.A.; Bedin, F.; Beseme, F.; Paranhos-Baccala, G.; Komurian-Pradel, F.; Mallet, F.; Tuke, P.W.; Voisset, C.; Blond, J.L.; et al. Molecular identification of a novel retrovirus repeatedly isolated from patients with multiple sclerosis. The Collaborative Research Group on Multiple Sclerosis. Proc. Natl. Acad. Sci. USA 1997, 94, 7583–7588. [Google Scholar] [CrossRef] [PubMed]
- Blond, J.L.; Besème, F.; Duret, L.; Bouton, O.; Bedin, F.; Perron, H.; Mandrand, B.; Mallet, F. Molecular characterization and placental expression of HERV-W, a new human endogenous retrovirus family. J. Virol. 1999, 73, 1175–1185. [Google Scholar] [PubMed]
- Blond, J.L.; Lavillette, D.; Cheynet, V.; Bouton, O.; Oriol, G.; Chapel-Fernandes, S.; Mandrand, B.; Mallet, F.; Cosset, F.L. An envelope glycoprotein of the human endogenous retrovirus HERV-W is expressed in the human placenta and fuses cells expressing the type D mammalian retrovirus receptor. J. Virol. 2000, 74, 3321–3329. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Lee, X.; Li, X.; Veldman, G.M.; Finnerty, H.; Racie, L.; Lavallie, E.; Tang, X.; Edouard, P.; Howes, S.; et al. Syncytin is a captive retroviral envelope protein involved. Nature 2000, 403, 785–789. [Google Scholar] [PubMed]
- Cheng, Y.-H. Isolation and Characterization of the Human Syncytin Gene Promoter. Biol. Reprod. 2003, 70, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Mallet, F.; Bouton, O.; Prudhomme, S.; Cheynet, V.; Oriol, G.; Bonnaud, B.; Lucotte, G.; Duret, L.; Mandrand, B. The endogenous retroviral locus ERVWE1 is a bona fide gene involved in hominoid placental physiology. Proc. Natl. Acad. Sci. USA 2004, 101, 1731–1736. [Google Scholar] [CrossRef] [PubMed]
- Bonnaud, B.; Bouton, O.; Oriol, G.; Cheynet, V.; Duret, L.; Mallet, F. Evidence of selection on the domesticated ERVWE1 env retroviral element involved in placentation. Mol. Biol. Evol. 2004, 21, 1895–1901. [Google Scholar] [CrossRef] [PubMed]
- Prudhomme, S.; Oriol, G.; Mallet, F. A retroviral promoter and a cellular enhancer define a bipartite element which controls env ERVWE1 placental expression. J. Virol. 2004, 78, 12157–12168. [Google Scholar] [CrossRef] [PubMed]
- Cheynet, V.; Ruggieri, A.; Oriol, G.; Blond, J.-L.; Boson, B.; Vachot, L.; Verrier, B.; Cosset, F.-L.; Mallet, F. Synthesis, assembly, and processing of the Env ERVWE1/syncytin human endogenous retroviral envelope. J. Virol. 2005, 79, 5585–5593. [Google Scholar] [CrossRef] [PubMed]
- Gimenez, J.; Mallet, F. ERVWE1 (endogenous retroviral family W, Env(C7), member 1). Atlas Genet. Cytogenet. Oncol. Haematol. 2008, 12, 134–148. [Google Scholar] [CrossRef]
- Pavlícek, A.; Paces, J.; Elleder, D. Processed Pseudogenes of Human Endogenous Retroviruses Generated by LINEs: Their Integration, Stability, and Distribution. Genome Res. 2002, 12, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Costas, J. Characterization of the intragenomic spread of the human endogenous retrovirus family HERV-W. Mol. Biol. Evol. 2002, 19, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Beck, C.R.; Garcia-Perez, J.L.; Badge, R.M.; Moran, J.V. LINE-1 Elements in Structural Variation and Disease. Annu. Rev. Genom. Hum. Genet. 2011, 12, 187–215. [Google Scholar] [CrossRef] [PubMed]
- Hancks, D.C.; Kazazian, H.H. Active human retrotransposons: Variation and disease. Curr. Opin. Genet. Dev. 2012, 22, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.R.; Narvaiza, I.; Planegger, R.A.; Weitzman, M.D.; Moran, J.V. APOBEC3A deaminates transiently exposed single-strand DNA during LINE-1 retrotransposition. Elife 2014, 3, e02008. [Google Scholar] [CrossRef] [PubMed]
- Lavialle, C.; Cornelis, G.; Dupressoir, A.; Esnault, C.; Heidmann, O.; Vernochet, C.; Heidmann, T. Paleovirology of “syncytins”, retroviral env genes exapted for a role in placentation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120507. [Google Scholar] [CrossRef] [PubMed]
- Mayer, J.; Meese, E. Human endogenous retroviruses in the primate lineage and their influence on host genomes. Cytogenet. Genome Res. 2005, 110, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Lavie, L.; Maldener, E.; Brouha, B.; Meese, E.U.; Mayer, J. The human L1 promoter: Variable transcription initiation sites and a major impact of upstream flanking sequence on promoter activity. Genome Res. 2004, 14, 2253–2260. [Google Scholar] [CrossRef] [PubMed]
- Voisset, C.; Bouton, O.; Bedin, F.; Duret, L.; Mandrand, B.; Mallet, F.; Paranhos-Baccala, G. Chromosomal distribution and coding capacity of the human endogenous retrovirus HERV-W family. AIDS Res. Hum. Retroviruses 2000, 16, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Perron, C.; Geny, A.; Laurent, C.; Mouriquand, J.; Pellat, J.; Perret, J.; Seigneurin, J. Leptomeningeal cell line from multiple sclerosis with reverse transcriptase activity and viral particles. Res. Virol. 1989, 140, 551–561. [Google Scholar] [CrossRef]
- Perron, H.; Lalande, B.; Gratacap, B.; Laurent, A.; Genoulaz, O.; Geny, C.; Mallaret, M.; Schuller, E.; Stoebner, P.; Seigneurin, J.J.M. Isolation of retrovirus from patients with multiple sclerosis. Lancet 1991, 337, 862–863. [Google Scholar] [CrossRef]
- Knerr, I.; Huppertz, B.; Weigel, C.; Dötsch, J.; Wich, C.; Schild, R.L.; Beckmann, M.W.; Rascher, W. Endogenous retroviral syncytin: Compilation of experimental research on syncytin and its possible role in normal and disturbed human placentogenesis. Mol. Hum. Reprod. 2004, 10, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Frendo, J.-L.; Olivier, D.; Cheynet, V.; Blond, J.-L.; Bouton, O.; Vidaud, M.; Rabreau, M.; Evain-Brion, D.; Mallet, F. Direct involvement of HERV-W Env glycoprotein in human trophoblast cell fusion and differentiation. Mol. Cell. Biol. 2003, 23, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Lavillette, D.; Marin, M.; Ruggieri, A.; Mallet, F.; Cosset, F.L.; Kabat, D. The envelope glycoprotein of human endogenous retrovirus type W uses a divergent family of amino acid transporters/cell surface receptors. J. Virol. 2002, 76, 6442–6452. [Google Scholar] [CrossRef] [PubMed]
- Malassiné, A.; Handschuh, K.; Tsatsaris, V.; Gerbaud, P.; Cheynet, V.; Oriol, G.; Mallet, F.; Evain-Brion, D. Expression of HERV-W Env glycoprotein (syncytin) in the extravillous trophoblast of first trimester human placenta. Placenta 2005, 26, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Li, J.; Wang, F.; Oliver, M.T.; Tipton, T.; Gao, Y.; Jiang, S.-W. Syncytin-1 modulates placental trophoblast cell proliferation by promoting G1/S transition. Cell. Signal. 2013, 25, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Keryer, G.; Alsat, E.; Tasken, K.; Evain-Brion, D. Cyclic AMP-dependent protein kinases and human trophoblast cell differentiation in vitro. J. Cell Sci. 1998, 111, 995–1004. [Google Scholar] [PubMed]
- Huang, Q.; Chen, H.; Wang, F.; Brost, B.C.; Li, J.; Li, Z.; Gao, Y.; Gao, Y.; Jiang, S.W. Reduced syncytin-1 expression in choriocarcinoma BeWo cells activates the calpain1-AIF-mediated apoptosis, implication for preeclampsia. Cell. Mol. Life Sci. 2014, 71, 3151–3164. [Google Scholar] [CrossRef] [PubMed]
- Mangeney, M.; Heidmann, T. Tumor cells expressing a retroviral envelope escape immune rejection in vivo. Proc. Natl. Acad. Sci. USA 1998, 95, 14920–14925. [Google Scholar] [CrossRef] [PubMed]
- Blaise, S.; Mangeney, M.; Heidmann, T. The envelope of Mason-Pfizer monkey virus has immunosuppressive properties. J. Gen. Virol. 2001, 82, 1597–1600. [Google Scholar] [CrossRef] [PubMed]
- Mangeney, M.; Renard, M.; Schlecht-Louf, G.; Bouallaga, I.; Heidmann, O.; Letzelter, C.; Richaud, A.; Ducos, B.; Heidmann, T. Placental syncytins: Genetic disjunction between the fusogenic and immunosuppressive activity of retroviral envelope proteins. Proc. Natl. Acad. Sci. USA 2007, 104, 20534–20539. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, J.M.; Schjenken, J.E.; Clifton, V.L.; Vargas, A.; Barbeau, B.; Lowry, P.; Maiti, K.; Smith, R. The endogenous retroviral envelope protein syncytin-1 inhibits LPS/PHA-stimulated cytokine responses in human blood and is sorted into placental exosomes. Placenta 2012, 33, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Karlsson, H. Expression and regulation of human endogenous retrovirus W elements. Apmis 2016, 124, 52–66. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Kim, H.; Kim, H. Expression of the human endogenous retrovirus HERV-W family in various human tissues and cancer cells. J. Gen. Virol. 2004, 85, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Ahn, K.; Kim, D.S. Quantitative expression of the HERV-W env gene in human tissues. Arch. Virol. 2008, 153, 1587–1591. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Hornung, D.; Kurek, R.; Ostman, H.; Helen, O.; Blomberg, J.; Bergqvist, A. Expression of human endogenous gammaretroviral sequences in endometriosis and ovarian cancer. AIDS Res. Hum. Retroviruses 2006, 22, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Nellåker, C.; Wållgren, U.; Karlsson, H. Molecular beacon-based temperature control and automated analyses for improved resolution of melting temperature analysis using SYBR I Green chemistry. Clin. Chem. 2007, 53, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Nellåker, C.; Li, F.; Uhrzander, F.; Tyrcha, J.; Karlsson, H. Expression profiling of repetitive elements by melting temperature analysis: Variation in HERV-W gag expression across human individuals and tissues. BMC Genom. 2009, 10, 532. [Google Scholar]
- Roland, C.S.; Hu, J.; Ren, C.E.; Chen, H.; Li, J.; Varvoutis, M.S.; Leaphart, L.W.; Byck, D.B.; Zhu, X.; Jiang, S.W. Morphological changes of placental syncytium and their implications for the pathogenesis of preeclampsia. Cell. Mol. Life Sci. 2016, 73, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Vargas, A.; Toufaily, C.; LeBellego, F.; Rassart, É.; Lafond, J.; Barbeau, B. Reduced expression of both syncytin 1 and syncytin 2 correlates with severity of preeclampsia. Reprod. Sci. 2011, 18, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Lee, X.; Keith, J.C.; Stumm, N.; Moutsatsos, I.; McCoy, J.M.; Crum, C.P.; Genest, D.; Chin, D.; Ehrenfels, C.; Pijnenborg, R.; et al. Downregulation of placental syncytin expression and abnormal protein localization in pre-eclampsia. Placenta 2001, 22, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Knerr, I.; Beinder, E.; Rascher, W. Syncytin, a novel human endogenous retroviral gene in human placenta: Evidence for its dysregulation in preeclampsia and HELLP syndrome. Am. J. Obstet. Gynecol. 2002, 186, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Holder, B.S.; Tower, C.L.; Abrahams, V.M.; Aplin, J.D. Syncytin 1 in the human placenta. Placenta 2012, 33, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.-W.; Li, J.; Brost, B.C.; Xia, X.-Y.; Chen, H.B.; Wang, C.-X.; Jiang, S.-W. Decreased expression and altered methylation of syncytin-1 gene in human placentas associated with preeclampsia. Curr. Pharm. Des. 2014, 20, 1796–1802. [Google Scholar] [CrossRef] [PubMed]
- Knerr, I.; Weigel, C.; Linnemann, K.; Dotsch, J.; Meissner, U.; Fusch, C.; Rascher, W. Transcriptional effects of hypoxia on fusiogenic syncytin and its receptor ASCT2 in human cytotrophoblast BeWo cells and in ex vivo perfused placental cotyledons. Am. J. Obs. Gynecol 2003, 189, 583–588. [Google Scholar] [CrossRef]
- Muir, A.; Lever, A.; Moffett, A. Expression and functions of human endogenous retroviruses in the placenta: An update. Placenta 2004, 25 (Suppl. A), S16–S25. [Google Scholar] [CrossRef] [PubMed]
- Frendo, J.L.; Vidaud, M.; Guibourdenche, J.; Luton, D.; Muller, F.; Bellet, D.; Giovagrandi, Y.; Tarrade, A.; Porquet, D.; Blot, P.; et al. Defect of villous cytotrophoblast differentiation into syncytiotrophoblast in Down’s syndrome. J. Clin. Endocrinol. Metab. 2000, 85, 3700–3707. [Google Scholar] [CrossRef] [PubMed]
- Massin, N.; Frendo, J.L.; Guibourdenche, J.; Luton, D.; Giovangrandi, Y.; Muller, F.; Vidaud, M.; Evain-Brion, D. Defect of syncytiotrophoblast formation and human chorionic gonadotropin expression in Down’s syndrome. Placenta 2001, 22, S93–S97. [Google Scholar] [CrossRef] [PubMed]
- Malassiné, A.; Frendo, J.L.; Evain-Brion, D. Trisomy 21—Affected placentas highlight prerequisite factors for human trophoblast fusion and differentiation. Int. J. Dev. Biol. 2010, 54, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Frendo, J.L.; Thérond, P.; Bird, T.; Massin, N.; Muller, F.; Guibourdenche, J.; Luton, D.; Vidaud, M.; Anderson, W.B.; Evain-Brion, D. Overexpression of copper zinc superoxide dismutase impairs human trophoblast cell fusion and differentiation. Endocrinology 2001, 142, 3638–3648. [Google Scholar] [CrossRef] [PubMed]
- Ruebner, M.; Strissel, P.L.; Langbein, M.; Fahlbusch, F.; Wachter, D.L.; Faschingbauer, F.; Beckmann, M.W.; Strick, R. Impaired cell fusion and differentiation in placentae from patients with intrauterine growth restriction correlate with reduced levels of HERV envelope genes. J. Mol. Med. 2010, 88, 1143–1156. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Li, J.; Podratz, K.C.; Tipton, T.; Marzolf, S.; Chen, H.B.; Jiang, S.-W. Hypomethylation and activation of syncytin-1 gene in endometriotic tissue. Curr. Pharm. Des. 2014, 20, 1786–1795. [Google Scholar] [CrossRef] [PubMed]
- Menendez, L.; Benigno, B.B.; McDonald, J.F. L1 and HERV-W retrotransposons are hypomethylated in human ovarian carcinomas. Mol. Cancer 2004, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Strick, R.; Ackermann, S.; Langbein, M.; Swiatek, J.; Schubert, S.W.; Hashemolhosseini, S.; Koscheck, T.; Fasching, P.A.; Schild, R.L.; Beckmann, M.W.; et al. Proliferation and cell-cell fusion of endometrial carcinoma are induced by the human endogenous retroviral Syncytin-1 and regulated by TGF-β. J. Mol. Med. 2007, 85, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Bjerregaard, B.; Holck, S.; Christensen, I.J.; Larsson, L.I. Syncytin is involved in breast cancer-endothelial cell fusions. Cell. Mol. Life Sci. 2006, 63, 1906–1911. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Carballo, D.; Acikelli, A.H.; Klein, J.; Jastrow, H.; Dammann, P.; Wyganowski, T.; Guemues, C.; Gustmann, S.; Bardenheuer, W.; Malak, S.; et al. Therapeutic potential of antiviral drugs targeting chemorefractory colorectal adenocarcinoma cells overexpressing endogenous retroviral elements. J. Exp. Clin. Cancer Res. 2015, 34, 81. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Uzhameckis, D.; Hedborg, F.; Blomberg, J. Dynamic and selective HERV RNA expression in neuroblastoma cells subjected to variation in oxygen tension and demethylation. Apmis 2016, 124, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, Z.C.; Yin, S.J.; Chen, Y.T.; Yu, H.L.; Zeng, J.; Zhang, Q.; Zhu, F. Human endogenous retrovirus W family envelope gene activates the small conductance Ca2+-activated K+ channel in human neuroblastoma cells through CREB. Neuroscience 2013, 247, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Gimenez, J.; Montgiraud, C.; Pichon, J.-P.; Bonnaud, B.; Arsac, M.; Ruel, K.; Bouton, O.; Mallet, F. Custom human endogenous retroviruses dedicated microarray identifies self-induced HERV-W family elements reactivated in testicular cancer upon methylation control. Nucleic Acids Res. 2010, 38, 2229–2246. [Google Scholar] [CrossRef] [PubMed]
- Maliniemi, P.; Vincendeau, M.; Mayer, J.; Frank, O.; Hahtola, S.; Karenko, L.; Carlsson, E.; Mallet, F.; Seifarth, W.; Leib-Mösch, C.; et al. Expression of human endogenous retrovirus-w including syncytin-1 in cutaneous T-cell lymphoma. PLoS ONE 2013, 8, e76281. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Liu, T.; Zhao, Z.; Chen, Y.; Zeng, J.; Liu, S.; Zhu, F. Mutations in 3′-long terminal repeat of HERV-W family in chromosome 7 upregulate syncytin-1 expression in urothelial cell carcinoma of the bladder through interacting with c-Myb. Oncogene 2014, 33, 3947–3958. [Google Scholar] [CrossRef] [PubMed]
- Ruprecht, K.; Mayer, J.; Sauter, M.; Roemer, K.; Mueller-Lantzsch, N. Endogenous retroviruses and cancer. Cell. Mol. Life Sci. 2008, 65, 3366–3382. [Google Scholar] [CrossRef] [PubMed]
- Balada, E.; Ordi-Ros, J.; Vilardell-Tarrés, M. Molecular mechanisms mediated by Human Endogenous Retroviruses (HERVs) in autoimmunity. Rev. Med. Virol. 2009, 19, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Balada, E.; Vilardell-Tarrés, M.; Ordi-Ros, J. Implication of human endogenous retroviruses in the development of autoimmune diseases. Int. Rev. Immunol. 2010, 29, 351–370. [Google Scholar] [CrossRef] [PubMed]
- Trela, M.; Nelson, P.N.; Rylance, P.B. The role of molecular mimicry and other factors in the association of Human Endogenous Retroviruses and autoimmunity. APMIS 2016, 124, 88–104. [Google Scholar] [CrossRef] [PubMed]
- Brodziak, A.; Ziółko, E.; Muc-Wierzgoń, M.; Nowakowska-zajdel, E.; Kokot, T.; Klakla, K. The role of human endogenous retroviruses in the pathogenesis of autoimmune diseases. Med. Sci. Monit. 2012, 18, RA80–RA88. [Google Scholar] [PubMed]
- Volkman, H.E.; Stetson, D.B. The enemy within: Endogenous retroelements and autoimmune disease. Nat. Immunol. 2014, 15, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Hurst, T.P.; Magiorkinis, G. Activation of the innate immune response by endogenous retroviruses. J. Gen. Virol. 2015, 96, 1207–1218. [Google Scholar] [CrossRef] [PubMed]
- Query, C.C.; Keene, J.D. A human autoimmune protein associated with U1 RNA contains a region of homology that is cross-reactive with retroviral p30gag antigen. Cell 1987, 51, 211–220. [Google Scholar] [CrossRef]
- Talal, N.; Flescher, E.; Dang, H. Are endogenous retroviruses involved in human autoimmune disease? J. Autoimmun. 1992, 5 (Suppl. A), 61–66. [Google Scholar] [CrossRef]
- Nelson, P.N.; Lever, A.M.; Bruckner, F.E.; Isenberg, D.A.; Kessaris, N.; Hay, F.C. Polymerase chain reaction fails to incriminate exogenous retroviruses HTLV-I and HIV-1 in rheumatological diseases although a minority of sera cross react with retroviral antigens. Ann. Rheum. Dis. 1994, 53, 749–754. [Google Scholar] [CrossRef] [PubMed]
- Mason, A.L.; Xu, L.; Guo, L.; Garry, R.F. Retroviruses in autoimmune liver disease: Genetic or environmental agents? Arch. Immunol. Ther. Exp. 1999, 47, 289–297. [Google Scholar]
- Sun, B.; Hu, L.; Luo, Z.Y.; Chen, X.P.; Zhou, H.H.; Zhang, W. DNA methylation perspectives in the pathogenesis of autoimmune diseases. Clin. Immunol. 2016, 164, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Antony, J.M.; Deslauriers, A.M.; Bhat, R.K.; Ellestad, K.K.; Power, C. Human endogenous retroviruses and multiple sclerosis: Innocent bystanders or disease determinants? Biochim. Biophys. Acta 2011, 1812, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Lafon, M.; Jouvin-Marche, E.; Marche, P.N.; Perron, H.; Woodland, D.L. Human viral superantigens: To be or not to be transactivated? Trends Immunol. 2002, 23, 238–239. [Google Scholar] [CrossRef]
- Christensen, T. Association of human endogenous retroviruses with multiple sclerosis and possible interactions with herpes viruses. Rev. Med. Virol. 2005, 15, 179–211. [Google Scholar] [CrossRef] [PubMed]
- Perron, H.; Bernard, C.; Bertrand, J.-B.; Lang, A.; Popa, I.; Sanhadji, K.; Portoukalian, J. Endogenous retroviral genes, Herpesviruses and gender in Multiple Sclerosis. J. Neurol. Sci. 2009, 286, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Krone, B.; Grange, J.M. Multiple Sclerosis: Are Protective Immune Mechanisms Compromised by a Complex Infectious Background? Autoimmune Dis. 2011, 2011, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Libbey, J.E.; Cusick, M.F.; Fujinami, R.S. Role of Pathogens in Multiple Sclerosis. Int. Rev. Immunol. 2013, 33, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Morandi, E.; Tarlinton, R.E.; Gran, B. Multiple sclerosis between genetics and infections: Human endogenous retroviruses in monocytes and macrophages. Front. Immunol. 2015, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Alliel, P.M.; Périn, J.-P.; Belliveau, J.; Pierig, R.; Nussbaum, J.-L.; Rieger, F. Multiple sclerosis: Clues on the retroviral hypothesis in the human genome (part 1). Comptes Rendus l’Académie des Sci. Ser. III Sci. la Vie 1998, 321, 495–499. [Google Scholar] [CrossRef]
- Alliel, P.M.; Perin, J.P.; Pierig, R.; Rieger, F. An endogenous retrovirus with nucleic acid sequences similar to those of the multiple sclerosis associated retrovirus at the human T-cell receptor alpha, delta gene locus. Cell. Mol. Biol. (Noisy-le-grand) 1998, 44, 927–931. [Google Scholar]
- Haahr, S.; Sommerlund, M.; Møller-Larsen, A.; Nielsen, R.; Hansen, H. Just another dubious virus in cells from a patient with multiple sclerosis? Lancet 1991, 337, 863–864. [Google Scholar] [CrossRef]
- Perron, H.; Firouzi, R.; Tuke, P.; Garson, J.A.; Michel, M.; Beseme, F.; Bedin, F.; Mallet, F.; Marcel, E.; Seigneurin, J.M.; et al. Cell cultures and associated retroviruses in multiple sclerosis. Acta Neurol. Scand. Suppl. 1997, 169, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Komurian-Pradel, F.; Paranhos-Baccala, G.; Bedin, F.; Ounanian-Paraz, A.; Sodoyer, M.; Ott, C.; Rajoharison, A.; Garcia, E.; Mallet, F.; Mandrand, B.; et al. Molecular cloning and characterization of MSRV-related sequences associated with retrovirus-like particles. Virology 1999, 260, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mameli, G.; Astone, V.; Arru, G.; Marconi, S.; Lovato, L.; Serra, C.; Sotgiu, S.; Bonetti, B.; Dolei, A. Brains and peripheral blood mononuclear cells of multiple sclerosis (MS) patients hyperexpress MS-associated retrovirus/HERV-W endogenous retrovirus, but not Human herpesvirus 6. J. Gen. Virol. 2007, 88, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Mameli, G.; Poddighe, L.; Astone, V.; Delogu, G.; Arru, G.; Sotgiu, S.; Serra, C.; Dolei, A. Novel reliable real-time PCR for differential detection of MSRVenv and syncytin-1 in RNA and DNA from patients with multiple sclerosis. J. Virol. Methods 2009, 161, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Dolei, A.; Perron, H. The multiple sclerosis-associated retrovirus and its HERV-W endogenous family: A biological interface between virology, genetics, and immunology in human physiology and disease. J. Neurovirol. 2009, 15, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Montojo, M.; Dominguez-Mozo, M.; Arias-Leal, A.; Garcia-Martinez, Á.; de las Heras, V.; Casanova, I.; Faucard, R.; Gehin, N.; Madeira, A.; Arroyo, R.; et al. The DNA Copy Number of Human Endogenous Retrovirus-W (MSRV-Type) Is Increased in Multiple Sclerosis Patients and Is Influenced by Gender and Disease Severity. PLoS ONE 2013, 8, e53623. [Google Scholar] [CrossRef] [PubMed]
- Blomberg, J.; Ushameckis, D.; Jern, P. Evolutionary Aspects of Human Endogenous Retroviral Sequences (HERVs) and Disease. In Madame Curie Bioscience Database; Landes Bioscience: Austin, TX, USA, 2000. [Google Scholar]
- Ruprecht, K.; Gronen, F.; Sauter, M.; Best, B.; Rieckmann, P.; Mueller-Lantzsch, N. Lack of immune responses against multiple sclerosis-associated retrovirus/human endogenous retrovirus W in patients with multiple sclerosis. J. Neurovirol. 2008, 14, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, K.; Richter, C.; Backes, C.; Meese, E.; Ruprecht, K.; Mayer, J. Comprehensive analysis of human endogenous retrovirus group HERV-W locus transcription in multiple sclerosis brain lesions by high-throughput amplicon sequencing. J. Virol. 2013, 87, 13837–13852. [Google Scholar] [CrossRef] [PubMed]
- Johnston, J.B.; Silva, C.; Holden, J.; Warren, K.G.; Clark, A.W.; Power, C. Monocyte activation and differentiation augment human endogenous retrovirus expression: Implications for inflammatory brain diseases. Ann. Neurol. 2001, 50, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Dolei, A.; Serra, C.; Mameli, G.; Pugliatti, M.; Sechi, G.; Cirotto, M.C.; Rosati, G.; Sotgiu, S. Multiple sclerosis-associated retrovirus (MSRV) in Sardinian MS patients. Neurology 2002, 58, 471–473. [Google Scholar] [CrossRef] [PubMed]
- Nowak, J.; Januszkiewicz, D.; Pernak, M.; Liwen, I.I.; Zawada, M.; Rembowska, J.; Nowicka, K.; Lewandowski, K.; Hertmanowska, H.; Wender, M. Multiple sclerosis-associated virus-related pol sequences found both in multiple sclerosis and healthy donors are more frequently expressed in multiple sclerosis patients. J. Neurovirol. 2003, 9, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Garson, J.; Tuke, P.; Giraud, P.; Paranhos-Baccala, G.; Perron, H. Detection of virion-associated MSRV-RNA in serum of patients with multiple sclerosis. Lancet 1998, 351, 33. [Google Scholar] [CrossRef]
- Antony, J.M.; van Marle, G.; Opii, W.; Butterfield, D.A.; Mallet, F.; Yong, V.W.; Wallace, J.L.; Deacon, R.M.; Warren, K.; Power, C. Human endogenous retrovirus glycoprotein-mediated induction of redox reactants causes oligodendrocyte death and demyelination. Nat. Neurosci. 2004, 7, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Antony, J.M.; Zhu, Y.; Izad, M.; Warren, K.G.; Vodjgani, M.; Mallet, F.; Power, C. Comparative Expression of Human Endogenous Retrovirus-W Genes in Multiple Sclerosis. AIDS Res. Hum. Retroviruses 2007, 23, 1251–1256. [Google Scholar] [CrossRef] [PubMed]
- Perron, H.; Germi, R.; Bernard, C.; Garcia-Montojo, M.; Deluen, C.; Farinelli, L.; Faucard, R.; Veas, F.; Stefas, I.; Fabriek, B.O.; et al. Human endogenous retrovirus type W envelope expression in blood and brain cells provides new insights into multiple sclerosis disease. Mult. Scler. 2012, 18, 1721–1736. [Google Scholar] [CrossRef] [PubMed]
- Arru, G.; Mameli, G.; Astone, V.; Serra, C.; Huang, Y.-M.; Link, H.; Fainardi, E.; Castellazzi, M.; Granieri, E.; Fernandez, M.; et al. Multiple Sclerosis and HERV-W/MSRV: A Multicentric Study. Int. J. Biomed. Sci. 2007, 3, 292–297. [Google Scholar] [PubMed]
- Zawada, M.; Liwén, I.; Pernak, M.; Januszkiewicz-Lewandowska, D.; Nowicka-Kujawska, K.; Rembowska, J.; Lewandowski, K.; Hertmanowska, H.; Wender, M.; Nowak, J. MSRV pol sequence copy number as a potential marker of multiple sclerosis. Pol. J. Pharmacol. 2003, 55, 869–875. [Google Scholar] [PubMed]
- Perron, H.; Lazarini, F.; Ruprecht, K.; Péchoux-Longin, C.; Seilhean, D.; Sazdovitch, V.; Créange, A.; Battail-Poirot, N.; Sibaï, G.; Santoro, L.; et al. Human endogenous retrovirus (HERV)-W ENV and GAG proteins: Physiological expression in human brain and pathophysiological modulation in multiple sclerosis lesions. J. Neurovirol. 2005, 11, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Brudek, T.; Christensen, T.; Aagaard, L.; Petersen, T.; Hansen, H.J.; Møller-Larsen, A. B cells and monocytes from patients with active multiple sclerosis exhibit increased surface expression of both HERV-H Env and HERV-W Env, accompanied by increased seroreactivity. Retrovirology 2009, 6, 104. [Google Scholar] [CrossRef] [PubMed]
- Van Horssen, J.; Van Der Pol, S.; Nijland, P.; Amor, S.; Perron, H. Human endogenous retrovirus W in brain lesions: Rationale for targeted therapy in multiple sclerosis. Mult. Scler. Relat. Disord. 2016, 8, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Perron, H.; Jouvin-Marche, E.; Michel, M.; Ounanian-Paraz, A.; Camelo, S.; Dumon, A.; Jolivet-Reynaud, C.; Marcel, F.; Souillet, Y.; Borel, E.; et al. Multiple Sclerosis Retrovirus Particles and Recombinant Envelope Trigger an Abnormal Immune Response in Vitro, by Inducing Polyclonal Vβ16 T-Lymphocyte Activation. Virology 2001, 287, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Rolland, A.; Jouvin-Marche, E.; Saresella, M.; Ferrante, P.; Cavaretta, R.; Créange, A.; Marche, P.; Perron, H. Correlation between disease severity and in vitro cytokine production mediated by MSRV (Multiple Sclerosis associated RetroViral element) envelope protein in patients with multiple sclerosis. J. Neuroimmunol. 2005, 160, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Rolland, A.; Jouvin-Marche, E.; Viret, C.; Faure, M.; Perron, H.; Marche, P.N. The Envelope Protein of a Human Endogenous Retrovirus-W Family Activates Innate Immunity through CD14/TLR4 and Promotes Th1-Like Responses. J. Immunol. 2006, 176, 7636–7644. [Google Scholar] [CrossRef] [PubMed]
- Saresella, M.; Rolland, A.; Marventano, I.; Cavarretta, R.; Caputo, D.; Marche, P.; Perron, H.; Clerici, M. Multiple sclerosis-associated retroviral agent (MSRV)-stimulated cytokine production in patients with relapsing-remitting multiple sclerosis. Mult. Scler. 2009, 15, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Mameli, G.; Astone, V.; Khalili, K.; Serra, C.; Sawaya, B.E.; Dolei, A. Regulation of the syncytin-1 promoter in human astrocytes by multiple sclerosis-related cytokines. Virology 2007, 362, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Kremer, D.; Schichel, T.; Förster, M.; Tzekova, N.; Bernard, C.; Van Der Valk, P.; Van Horssen, J.; Hartung, H.P.; Perron, H.; Küry, P. Human endogenous retrovirus type W envelope protein inhibits oligodendroglial precursor cell differentiation. Ann. Neurol. 2013, 74, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Kremer, D.; Förster, M.; Schichel, T.; Göttle, P.; Hartung, H.-P.; Perron, H.; Küry, P. The neutralizing antibody GNbAC1 abrogates HERV-W envelope protein-mediated oligodendroglial maturation blockade. Mult. Scler. 2014, 21, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Madeira, A.; Burgelin, I.; Perron, H.; Curtin, F.; Lang, A.B.; Faucard, R. MSRV envelope protein is a potent, endogenous and pathogenic agonist of human toll-like receptor 4: Relevance of GNbAC1 in multiple sclerosis treatment. J. Neuroimmunol. 2016, 291, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.; Møller-Larsen, A.; Thiel, S.; Brudek, T.; Hansen, T.K.; Christensen, T. Effects of interferon-beta therapy on innate and adaptive immune responses to the human endogenous retroviruses HERV-H and HERV-W, cytokine production, and the lectin complement activation pathway in multiple sclerosis. J. Neuroimmunol. 2009, 215, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Mameli, G.; Serra, C.; Astone, V.; Castellazzi, M.; Poddighe, L.; Fainardi, E.; Neri, W.; Granieri, E.; Dolei, A. Inhibition of multiple-sclerosis-associated retrovirus as biomarker of interferon therapy. J. Neurovirol. 2008, 14, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Mameli, G.; Cossu, D.; Cocco, E.; Frau, J.; Marrosu, M.G.; Niegowska, M.; Sechi, L.A. Epitopes of HERV-Wenv induce antigen-specific humoral immunity in multiple sclerosis patients. J. Neuroimmunol. 2015, 280, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Firouzi, R.; Rolland, A.; Michel, M.; Jouvin-Marche, E.; Hauw, J.; Malcus-Vocanson, C.; Lazarini, F.; Gebuhrer, L.; Seigneurin, J.; Touraine, J.; et al. Multiple sclerosis–associated retrovirus particles cause T lymphocyte–dependent death with brain hemorrhage in humanized SCID mice model. J. Neurovirol. 2003, 9, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Antony, J.M.; Ellestad, K.K.; Hammond, R.; Imaizumi, K.; Mallet, F.; Warren, K.G.; Power, C. The human endogenous retrovirus envelope glycoprotein, syncytin-1, regulates neuroinflammation and its receptor expression in multiple sclerosis: A role for endoplasmic reticulum chaperones in astrocytes. J. Immunol. 2007, 179, 1210–1224. [Google Scholar] [CrossRef] [PubMed]
- Perron, H.; Dougier-Reynaud, H.-L.; Lomparski, C.; Popa, I.; Firouzi, R.; Bertrand, J.-B.; Marusic, S.; Portoukalian, J.; Jouvin-Marche, E.; Villiers, C.L.; et al. Human Endogenous Retrovirus Protein Activates Innate Immunity and Promotes Experimental Allergic Encephalomyelitis in Mice. PLoS ONE 2013, 8, e80128. [Google Scholar] [CrossRef] [PubMed]
- Nellaker, C.; Yao, Y.; Jones-Brando, L.; Mallet, F.; Yolken, R.H.; Karlsson, H. Transactivation of elements in the human endogenous retrovirus W family by viral infection. Retrovirology 2006, 3, 44. [Google Scholar] [CrossRef] [PubMed]
- Laufer, G.; Mayer, J.; Mueller, B.F.; Mueller-Lantzsch, N.; Ruprecht, K. Analysis of transcribed human endogenous retrovirus W env loci clarifies the origin of multiple sclerosis-associated retrovirus env sequences. Retrovirology 2009, 6, 37. [Google Scholar] [CrossRef] [PubMed]
- Roebke, C.; Wahl, S.; Laufer, G.; Stadelmann, C.; Sauter, M.; Mueller-Lantzsch, N.; Mayer, J.; Ruprecht, K. An N-terminally truncated envelope protein encoded by a human endogenous retrovirus W locus on chromosome Xq22.3. Retrovirology 2010, 7, 69. [Google Scholar] [CrossRef] [PubMed]
- Do Olival, G.S.; Faria, T.S.; Nali, L.H.S.; de Oliveira, A.C.P.; Casseb, J.; Vidal, J.E.; Cavenaghi, V.B.; Tilbery, C.P.; Moraes, L.; Fink, M.C.S.; et al. Genomic analysis of ERVWE2 locus in patients with multiple sclerosis: Absence of genetic association but potential role of human endogenous retrovirus type W elements in molecular mimicry with myelin antigen. Front. Microbiol. 2013, 4, 1–7. [Google Scholar] [CrossRef] [PubMed]
- García-Montojo, M.; de la Hera, B.; Varadé, J.; de la Encarnación, A.; Camacho, I.; Domínguez-Mozo, M.; Arias-Leal, A.; García-Martínez, A.; Casanova, I.; Izquierdo, G.; et al. HERV-W polymorphism in chromosome X is associated with multiple sclerosis risk and with differential expression of MSRV. Retrovirology 2014, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Varadé, J.; García-Montojo, M.; de la Hera, B.; Camacho, I.; García-Martnez, M.Á.; Arroyo, R.; Álvarez-Lafuente, R.; Urcelay, E. Multiple sclerosis retrovirus-like envelope gene: Role of the chromosome 20 insertion. BBA Clin. 2015, 3, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.K.; Ellestad, K.K.; Wheatley, B.M.; Warren, R.; Holt, R.A.; Power, C. Age- and disease-dependent HERV-W envelope allelic variation in brain: Association with neuroimmune gene expression. PLoS ONE 2011, 6, e19176. [Google Scholar] [CrossRef] [PubMed]
- Hon, G.M.; Erasmus, R.T.; Matsha, T. Multiple sclerosis-associated retrovirus and related human endogenous retrovirus-W in patients with multiple sclerosis: A literature review. J. Neuroimmunol. 2013, 263, 8–12. [Google Scholar] [CrossRef] [PubMed]
- P. Ryan, F. Human Endogenous Retroviruses in Multiple Sclerosis: Potential for Novel Neuro-Pharmacological Research. Curr. Neuropharmacol. 2011, 9, 360–369. [Google Scholar]
- Curtin, F.; Perron, H.; Kromminga, A.; Porchet, H.; Lang, A.B. Preclinical and early clinical development of GNbAC1, a humanized IgG4 monoclonal antibody targeting endogenous retroviral MSRV-Env protein. MAbs 2015, 7, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Curtin, F.; Perron, H.; Faucard, R.; Porchet, H.; Lang, A.B. Treatment Against Human Endogenous Retrovirus: A Possible Personalized Medicine Approach for Multiple Sclerosis. Mol. Diagn. Ther. 2015, 19, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Gaudin, P.; Ijaz, S.; Tuke, P.W.; Marcel, F.; Paraz, A.; Seigneurin, J.M.; Mandrand, B.; Perron, H.; Garson, J.A. Infrequency of detection of particle-associated MSRV/HERV-W RNA in the synovial fluid of patients with rheumatoid arthritis. Rheumatology (Oxford) 2000, 39, 950–954. [Google Scholar] [CrossRef] [PubMed]
- Bendiksen, S.; Martinez-Zubiavrra, I.; Tümmler, C.; Knutsen, G.; Elvenes, J.; Olsen, E.; Olsen, R.; Moens, U. Human Endogenous Retrovirus W Activity in Cartilage of Osteoarthritis Patients. BioMed Res. Int. 2014, 2014, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Faucard, R.; Madeira, A.; Gehin, N.; Authier, F.-J.; Panaite, P.-A.; Lesage, C.; Burgelin, I.; Bertel, M.; Bernard, C.; Curtin, F.; et al. Human Endogenous Retrovirus and Neuroinflammation in Chronic Inflammatory Demyelinating Polyradiculoneuropathy. EBioMedicine 2016, 6, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Molès, J.P.; Tesniere, A.; Guilhou, J.J. A new endogenous retroviral sequence is expressed in skin of patients with psoriasis. Br. J. Dermatol. 2005, 153, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Hohenadl, C.; Germaier, H.; Walchner, M.; Hagenhofer, M.; Herrmann, M.; Stürzl, M.; Kind, P.; Hehlmann, R.; Erfle, V.; Leib-Mösch, C. Transcriptional activation of endogenous retroviral sequences in human epidermal keratinocytes by UVB irradiation. J. Invest. Dermatol. 1999, 113, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Schanab, O.; Humer, J.; Gleiss, A.; Mikula, M.; Sturlan, S.; Grunt, S.; Okamoto, I.; Muster, T.; Pehamberger, H.; Waltenberger, A. Expression of human endogenous retrovirus K is stimulated by ultraviolet radiation in melanoma. Pigment Cell Melanoma Res. 2011, 24, 656–665. [Google Scholar] [CrossRef] [PubMed]
- De Sousa Nogueira, M.A.; Biancardi Gavioli, C.F.; Pereira, N.Z.; de Carvalho, G.C.; Domingues, R.; Aoki, V.; Sato, M.N. Human endogenous retrovirus expression is inversely related with the up-regulation of interferon-inducible genes in the skin of patients with lichen planus. Arch. Dermatol. Res. 2015, 307, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Weis, S.; Llenos, I.C.; Sabunciyan, S.; Dulay, J.R.; Isler, L.; Yolken, R.; Perron, H. Reduced expression of human endogenous retrovirus (HERV)-W GAG protein in the cingulate gyrus and hippocampus in schizophrenia, bipolar disorder, and depression. J. Neural Transm. 2007, 114, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Diem, O.; Schäffner, M.; Seifarth, W.; Leib-Mösch, C. Influence of antipsychotic drugs on human endogenous retrovirus (HERV) transcription in brain cells. PLoS ONE 2012, 7, e30054. [Google Scholar] [CrossRef] [PubMed]
- Oluwole, S.O.A.; Yao, Y.; Conradi, S.; Kristensson, K.; Karlsson, H. Elevated levels of transcripts encoding a human retroviral envelope protein (syncytin) in muscles from patients with motor neuron disease. Amyotroph. Lateral Scler. 2007, 8, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Jeong, B.-H.; Lee, Y.-J.; Carp, R.I.; Kim, Y.-S. The prevalence of human endogenous retroviruses in cerebrospinal fluids from patients with sporadic Creutzfeldt-Jakob disease. J. Clin. Virol. 2010, 47, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Balestrieri, E.; Arpino, C.; Matteucci, C.; Sorrentino, R.; Pica, F.; Alessandrelli, R.; Coniglio, A.; Curatolo, P.; Rezza, G.; Macciardi, F.; et al. HERVs Expression in Autism Spectrum Disorders. PLoS ONE 2012, 7, e48831. [Google Scholar] [CrossRef] [PubMed]
- Balestrieri, E.; Pitzianti, M.; Matteucci, C.; D’Agati, E.; Sorrentino, R.; Baratta, A.; Caterina, R.; Zenobi, R.; Curatolo, P.; Garaci, E.; et al. Human endogenous retroviruses and ADHD. World J. Biol. Psychiatry 2014, 15, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Deb-Rinker, P.; Klempan, T.A.; O’Reilly, R.L.; Torrey, E.F.; Singh, S.M. Molecular characterization of a MSRV-like sequence identified by RDA from monozygotic twin pairs discordant for schizophrenia. Genomics 1999, 61, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, H.; Bachmann, S.; Schröder, J.; McArthur, J.; Torrey, E.F.; Yolken, R.H. Retroviral RNA identified in the cerebrospinal fluids and brains of individuals with schizophrenia. Proc. Natl. Acad. Sci. USA 2001, 98, 4634–4639. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, H.; Schröder, J.; Bachmann, S.; Bottmer, C.; Yolken, R.H. HERV-W-related RNA detected in plasma from individuals with recent-onset schizophrenia or schizoaffective disorder. Mol. Psychiatry 2003, 9, 12–13. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Schröder, J.; Nellåker, C.; Bottmer, C.; Bachmann, S.; Yolken, R.H.; Karlsson, H. Elevated levels of human endogenous retrovirus-W transcripts in blood cells from patients with first episode schizophrenia. Genes Brain Behav. 2008, 7, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Li, S.; Hu, Y.; Yu, H.; Luo, F.; Zhang, Q.; Zhu, F. Implication of the env gene of the human endogenous retrovirus W family in the expression of BDNF and DRD3 and development of recent-onset schizophrenia. Schizophr. Bull. 2011, 37, 988–1000. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Li, S.; Yan, Q.; Wang, X.; Chen, Y.; Zhou, P.; Lu, M.; Zhu, F. Elevation of Ser9 phosphorylation of GSK3β is required for HERV-W env-mediated BDNF signaling in human U251 cells. Neurosci. Lett. 2016, 627, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Perron, H.; Mekaoui, L.; Bernard, C.; Veas, F.; Stefas, I.; Leboyer, M. Endogenous Retrovirus Type W GAG and Envelope Protein Antigenemia in Serum of Schizophrenic Patients. Biol. Psychiatry 2008, 64, 1019–1023. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, H. GABBR1 has a HERV-W LTR in its regulatory region--a possible implication for schizophrenia. Biol. Direct 2013, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Frank, O.; Giehl, M.; Zheng, C.; Hehlmann, R.; Leib-Mosch, C.; Seifarth, W. Human endogenous retrovirus expression profiles in samples from brains of patients with schizophrenia and bipolar disorders. J. Virol. 2005, 79, 10890–10901. [Google Scholar] [CrossRef] [PubMed]
- Perron, H.; Hamdani, N.; Faucard, R.; Lajnef, M.; Jamain, S.; Daban-Huard, C.; Sarrazin, S.; LeGuen, E.; Houenou, J.; Delavest, M.; et al. Molecular characteristics of Human Endogenous Retrovirus type-W in schizophrenia and bipolar disorder. Transl. Psychiatry 2012, 2, e201. [Google Scholar] [CrossRef] [PubMed]
- Gürtler, C.; Bowie, A.G. Innate immune detection of microbial nucleic acids. Trends Microbiol. 2013, 21, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Melder, D.C.; Pankratz, V.S.; Federspiel, M.J. Evolutionary Pressure of a Receptor Competitor Selects Different Subgroup A Avian Leukosis Virus Escape Variants with Altered Receptor Interactions. J. Virol. 2003, 77, 10504–10514. [Google Scholar] [CrossRef] [PubMed]
- Spencer, T.E.; Mura, M.; Gray, C.A.; Griebel, P.J.; Palmarini, M. Receptor usage and fetal expression of ovine endogenous betaretroviruses: Implications for coevolution of endogenous and exogenous retroviruses. J. Virol. 2003, 77, 749–753. [Google Scholar] [CrossRef] [PubMed]
- An, D.S.; Xie, Y.M.; Chen, I.S. Envelope gene of the human endogenous retrovirus HERV-W encodes a functional retrovirus envelope. J. Virol. 2001, 75, 3488–3489. [Google Scholar] [CrossRef] [PubMed]
- Vincendeau, M.; Göttesdorfer, I.; Schreml, J.M.H.; Wetie, A.G.N.; Mayer, J.; Greenwood, A.D.; Helfer, M.; Kramer, S.; Seifarth, W.; Hadian, K.; et al. Modulation of human endogenous retrovirus (HERV) transcription during persistent and de novo HIV-1 infection. Retrovirology 2015, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- Hurst, T.; Pace, M.; Katzourakis, A.; Phillips, R.; Klenerman, P.; Frater, J.; Magiorkinis, G. Human endogenous retrovirus (HERV) expression is not induced by treatment with the histone deacetylase (HDAC) inhibitors in cellular models of HIV-1 latency. Retrovirology 2016, 13, 10. [Google Scholar] [CrossRef] [PubMed]
- Uleri, E.; Mei, A.; Mameli, G.; Poddighe, L.; Serra, C.; Dolei, A. HIV Tat acts on endogenous retroviruses of the W family and this occurs via Toll-like receptor 4: Inference for neuroAIDS. AIDS 2014, 28, 2659–2670. [Google Scholar] [CrossRef] [PubMed]
- Toufaily, C.; Landry, S.; Leib-Mosch, C.; Rassart, E.; Barbeau, B. Activation of LTRs from different human endogenous retrovirus (HERV) families by the HTLV-1 tax protein and T-cell activators. Viruses 2011, 3, 2146–2159. [Google Scholar] [CrossRef] [PubMed]
- Garrison, K.E.; Jones, R.B.; Meiklejohn, D.A.; Anwar, N.; Ndhlovu, L.C.; Chapman, J.M.; Erickson, A.L.; Agrawal, A.; Spotts, G.; Hecht, F.M.; et al. T cell responses to human endogenous retroviruses in HIV-1 infection. PLoS Pathog. 2007, 3, 1617–1627. [Google Scholar] [CrossRef] [PubMed]
- Perron, H.; Suh, M.; Lalande, B.; Gratacap, B.; Laurent, A.; Stoebner, P.; Seigneurin, J.R. Herpes simplex virus ICP0 and ICP4 immediate early proteins strongly enhance expression of a retrovirus harboured by a leptomeningeal cell line from a patient with multiple sclerosis. J. Gen. Virol. 1993, 74, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Ruprecht, K.; Obojes, K.; Wengel, V.; Gronen, F.; Kim, K.S.; Perron, H.; Schneider-Schaulies, J.; Rieckmann, P. Regulation of human endogenous retrovirus W protein expression by herpes simplex virus type 1: Implications for multiple sclerosis. J. Neurovirol. 2006, 12, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Kwun, H.J.; Kim, H.S.; Jang, K.L. Activation of the human endogenous retrovirus W long terminal repeat by herpes simplex virus type 1 immediate early protein 1. Mol. Cells 2003, 15, 75–80. [Google Scholar] [PubMed]
- Krone, B.; Oeffner, F.; Grange, J.M. Is the risk of multiple sclerosis related to the “biography” of the immune system? J. Neurol. 2009, 256, 1052–1060. [Google Scholar] [CrossRef] [PubMed]
- Mameli, G.; Poddighe, L.; Mei, A.; Uleri, E.; Sotgiu, S.; Serra, C.; Manetti, R.; Dolei, A. Expression and activation by Epstein Barr virus of human endogenous retroviruses-W in blood cells and astrocytes: Inference for multiple sclerosis. PLoS ONE 2012, 7, e44991. [Google Scholar] [CrossRef] [PubMed]
- Assinger, A.; Yaiw, K.-C.; Göttesdorfer, I.; Leib-Mösch, C.; Söderberg-Nauclér, C. Human cytomegalovirus (HCMV) induces human endogenous retrovirus (HERV) transcription. Retrovirology 2013, 10, 132. [Google Scholar] [CrossRef] [PubMed]
- Bergallo, M.; Galliano, I.; Montanari, P.; Gambarino, S.; Mareschi, K.; Ferro, F.; Fagioli, F.; Tovo, P.A.; Ravanini, P. CMV induces HERV-K and HERV-W expression in kidney transplant recipients. J. Clin. Virol. 2015, 68, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Shen, K.; Lin, M.; Chen, P.; Lin, C.; Chang, G.D.; Chen, H. GCMa regulates the syncytin-mediated trophoblastic fusion. J. Biol. Chem. 2002, 277, 50062–50068. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Nellåker, C.; Sabunciyan, S.; Yolken, R.H.; Jones-Brando, L.; Johansson, A.-S.; Owe-Larsson, B.; Karlsson, H. Transcriptional derepression of the ERVWE1 locus following influenza A virus infection. J. Virol. 2014, 88, 4328–4337. [Google Scholar] [CrossRef] [PubMed]
- Ponferrada, V.G.; Mauck, B.S.; Wooley, D.P. The envelope glycoprotein of human endogenous retrovirus HERV-W induces cellular resistance to spleen necrosis virus. Arch. Virol. 2003, 148, 659–675. [Google Scholar] [CrossRef] [PubMed]
- Machnik, G.; Klimacka-Nawrot, E.; Sypniewski, D.; Matczyńska, D.; Gałka, S.; Bednarek, I.; Okopień, B. Porcine endogenous retrovirus (PERV) infection of HEK-293 cell line alters expression of human endogenous retrovirus (HERV-W) sequences. Folia Biol. (Czech Republic) 2014, 60, 35–46. [Google Scholar]
- Sassaman, D.M.; Dombroski, B.A.; Moran, J.V.; Kimberland, M.L.; Naas, T.P.; DeBerardinis, R.J.; Gabriel, A.; Swergold, G.D.; Kazazian, H.H. Many human L1 elements are capable of retrotransposition. Nat. Genet. 1997, 16, 37–43. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | Method | Ref. | Possible Biases a |
|---|---|---|---|
| Blood | Search of Syncytin query in EST data | [11] | Low total HERV EST counts, could not detect HERV-Ws divergent from Syncytin, no information on LTR activity, number of cDNA/EST libraries great variability across tissues, under-representation of poorly expressed genes in small libraries (1) |
| Brain | Search of Syncytin query in EST data | [11] | (1) |
| RT-PCR (gag+, pol+, env+) | [55] | Primers specific for single expressed sequences (placental Syncytin (gag: AF072500, env: AF072506), MSRV clones (pol: AF009668)) could not detect divergent HERV-Ws, no information on full-length HERVs expression and LTR activity, samples amount is poorly representative (2) | |
| Brain (cortex and pons) | env real time qRT-PCR | [56] | Primers specific for placental Syncytin (NM_014590.3) can could not detect env defective or highly divergent HERV-Ws, no information on full-length HERVs expression and LTR activity, samples amount is poorly representative (3) |
| Breast | Search of Syncytin query in EST data | [11] | (1) |
| env real time qRT-PCR | [56] | (3) | |
| Colon | env real time qRT-PCR | [56] | (3) |
| Heart | RT-PCR (gag−, pol−, env+) | [55] | (2) |
| Endometrium | GammaHERV and HERV-W pol-based probe and probe-less real time qPCRs | [57] [14] | Could not detect transcripts defective or highly divergent for pol gene, no information about full-length sequences expression and LTR activity, samples amount is poorly representative (4) |
| Kidney | pol-expression arrays hybridization | [10] | Cross-amplification/hybridization of related HERV groups; could not detect transcripts defective for pol gene, no information about full-length sequences expression and LTR activity, no quantitative information, samples amount is poorly representative (5) |
| RT-PCR (gag−, pol+, env+) | [55] | (2) | |
| Liver | pol-expression arrays hybridization | [10] | (5) |
| RT-PCR (gag−, pol+, env+) | [55] | (2) | |
| env real time qRT-PCR | [56] | (3) | |
| Liver-spleen (fetal) | Search of Syncytin query in EST data | [11] | (1) |
| Lung | RT-PCR (gag−, pol+, env+) | [55] | (2) |
| Ovary | Search of Syncytin query in EST data | [11] | (1) |
| GammaHERV and HERV-W pol-based probe and probe-less real time qPCRs | [57] [14] | (4) | |
| PBMC | pol RT-PCR and env real time PCR | [17] | Low sensitivity and cross-amplification of related HERV groups by RT-PCR degenerate primers, real time PCR primers specific for placental Syncytin (NM_014590.3) could not detect divergent HERV-Ws and transcripts defective for pol/env genes, no information on full-length sequences expression and LTR activity, incomplete characterization of individuals health status |
| Prostate | RT-PCR (gag−, pol+, env+) | [55] | (2) |
| Skel. Muscle | RT-PCR (gag−, pol+, env+) | [55] | (2) |
| Spleen | RT-PCR (gag+, pol+, env+) | [55] | (2) |
| Stomach | env real time qRT-PCR | [56] | (3) |
| Testis | RT-PCR (gag+, pol+, env+) | [55] | (2) |
| Thymus | RT-PCR (gag−, pol+, env+) | [55] | (2) |
| Uterus | RT-PCR (gag−, pol−, env+) | [55] | (2) |
| env real time qRT-PCR | [56] | (3) |
| Locus | Chr:start-end (Strand) a | Type | Genomic Context b | Tissue | Method | Ref. |
|---|---|---|---|---|---|---|
| 2q22.1 | 2:139030735-139031481 (−) | Solo LTR | LTR8 (+) | Testis | Microarray | [15] |
| 2q24.3 | 2:165514421-165516121 (−) | Pseudogene | COBLL1 (−) TCONS_00004484 (−) | Placenta | Microarray | [15] |
| 5q12.1 * | 5:59954322-59962280 (+) | Provirus | DEPDC1B (−) | Placenta | Microarray | [15] |
| 7q21.2 * | 7:92097313:92107506 (−) | Provirus | - | Placenta Testis | Northern blot | [23] [25] |
| 15q21.2 | 15:51552784-51553570 (+) | Solo LTR | CYP19A1 (−) | Placenta | Microarray | [15] |
| Xq21.33 | X:93824238-93824702 (−) | Solo LTR | MER4A (−) | Placenta | Microarray | [15] |
| Tumoral Tissue | Ref. | Method a | Physiol. Expression b | Possible Biases of HERV-W Members Underrepresentation c |
|---|---|---|---|---|
| B cells | [55] * | RT-PCR (gag−, pol−, env+) | [17] ° | Primers specific for single expressed sequences (placental Syncytin-1 gag AF072500 and env AF072506; MSRV clones pol AF009668) could not detect divergent HERV-Ws, no information on full-length HERVs expression and LTR activity, samples amount is poorly representative (2) |
| Bladder | [55] * | RT-PCR (gag−, pol+, env+) | - | (2) |
| Breast | [11] | Search of Syncytin-1 in EST data | [11,56] | Low total HERV EST counts, could not detect HERV-Ws divergent from Syncytin-1, no information on LTR activity, number of cDNA/EST libraries great variability across tissues, under-representation of poorly expressed genes in small libraries (1) |
| [76] * | RT-PCR, real time qRT-PCR, | Specific detection of a Syncytin-1 env portion only, could not detect transcripts divergent/defective for env, no information on full-length sequences expression and LTR activity | ||
| [56] | env real time qRT-PCR | Primers specific for placental Syncytin-1 (NM_014590.3) could not detect env defective or highly divergent HERV-Ws, no information on full-length HERVs expression and LTR activity, samples amount is poorly representative (3) | ||
| [55] * | RT-PCR (gag−, pol+, env+) | (2) | ||
| Brain | [55] * | RT-PCR (gag−, pol+, env+) | [11,55] | (2) |
| Colon | [11] | Search of Syncytin-1 in EST data | [56] | (1) |
| [56] | env real time qRT-PCR | (3) | ||
| [55] * | RT-PCR (gag−, pol+, env+) | (2) | ||
| [77] * | qPCR | Specific detection of a Syncytin-1 env portion only, could not detect transcripts divergent/defective for env, no information on full-length sequences expression and LTR activity | ||
| Endometrium | [75] | qPCR, RT-PCR, NB, WB | [14,57,75] | Specific detection of a small portion of Syncytin-1 env only, samples amount is poorly representative, expression values are highly heterogeneous |
| Esophagus | [55] * | RT-PCR (gag−, pol+, env+) | - | (2) |
| Histiocyte | [55] * | RT-PCR (gag−, pol+, env+) | - | (2) |
| Kidney | [11] | Search of Syncytin-1 in EST data | [10,55] | (1) |
| [55] * | RT-PCR (gag−, pol+, env+) | (2) | ||
| Liver | [56] | env real time qRT-PCR | [10,55,56] | (3) |
| [55] * | RT-PCR (gag−, pol+, env+) | (2) | ||
| Lung | [55] * | RT-PCR (gag−, pol+, env+) | [55] | (2) |
| Neuroblasts | [78] * | pol real time qPCRs | - | Could not detect transcripts defective or highly divergent for pol gene, no information about full-length sequences expression and LTR activity, samples amount is poorly representative (4) |
| Ovary | [74] | Real time qRT-PCR | [57,74] | Primers designed on Syncytin-1 locus (AC000064) could not detect divergent HERV-Ws, samples amount is poorly representative |
| [57] | pol real time qPCRs | (4) | ||
| [55] * | RT-PCR (gag−, pol+, env+) | (2) | ||
| Pancreas | [55] * | RT-PCR (gag−, pol+, env+) | - | (2) |
| Placenta | [11] | Search of Syncytin-1 in EST data | [23,24,25] | (1) |
| Prostate | [55] * | RT-PCR (gag−, pol−, env+) | [55] | (2) |
| Skin | [55] * | RT-PCR (gag−, pol−, env+) | - | (2) |
| Stomach | [56] | env real time qRT-PCR | [56] | (3) |
| [55] * | RT-PCR (gag−, pol+, env+) | (2) | ||
| T-cells | [55] * | RT-PCR (gag−, pol+, env+) | [17] ° | (2) |
| Uterus | [56] | env real time qRT-PCR | [55,56] | (3) |
| [55] * | RT-PCR (gag−, pol+, env+) | (2) |
| Locus | Chr:start-end (Strand) a | Type b | Genomic Context c | Tissue d | LTR e | Method f | Ref. |
|---|---|---|---|---|---|---|---|
| 1q31.2 | 1:192855545-192856320 (−) | LTR | MER21C (−) | Testis | - | MA | [15] |
| 2p24.2 | 2:17520208-17527981 (+) | PV | L3 (−) | Testis | Pro ° | MA, qRT-PCR | [15,80] |
| 2p12 | 2:76098816-76106624 (+) | PV | - | Testis | Pro | MA | [15] |
| 3p12.3 | 3:74921984-74927237 (−) | PG | - | Prostate | - | MA | [15] |
| 3q28 | 3:191376573-191383381 (+) | PG | - | Testis | - | MA | [15] |
| 4p13 | 4:42287455-42294913 (−) | PV | TCONS_00007753 (−) | Testis | Pro ° | MA, qRT-PCR | [15,80] |
| 4q26 | 4:114965536-114972972 (+) | PG | - | Testis | - | MA | [15] |
| 5p13.3 | 5:31109366-31109859 (−) | LTR | - | Ovary | - | MA | [15] |
| 6q21 | 6:106676012-106683689 (+) | PG | ATG5 (−) | Skin T cells | - | MA, qRT-PCR | [81] |
| 7q21.2 | 7:92097313:92107506 (−) | PV | - | Testis* | Pro ° | MA, qRT-PCR | [80] |
| Bladder | Pro | qRT-PCR | [82] | ||||
| Skin T cells | - | MA, qRT-PCR | [81] | ||||
| 7q21.3 | 7:95987661-95988433 (−) | LTR | Alu Sx (−) | Testis | - | MA | [15] |
| 7q31.1b | 7:114019143-114026368 (−) | PG | FOXP2 (+) | Testis | - | MA | [15] |
| 7q36.3 | 7:155177752-155178503 (−) | LTR | BC150495 (+) | Testis | PA | MA | [15] |
| 8q24.13 | 8:125912007-125919468 (−) | PV | - | Prostate | Pro | MA | [15] |
| 13q21.1 | 13:55627766-55635877 (+) | PV | - | Testis | - | MA | [15] |
| 13q21.33 | 13:69795752-69799468 (+) | PV | LINC00383 (+) (Ex) | Testis | Pro ° | MA, qRT-PCR | [80] |
| 16p12.3 | 16:18124951-18125494 (−) | LTR | - | Testis | - | MA | [15] |
| 17q22 | 17:53088886-53095859 (−) | PG | STXBP4 (+) | Testis | - | MA | [15] |
| 21q21.1 | 21:20125060-20132866 (−) | PV | MIR548XHG (−) (Ex) | Testis | - | MA | [15] |
| 21q21.3 | 21:28226756-28234297 (+) | PV | - | Testis | Pro ° | MA, qRT-PCR | [15,80] |
| Xq21.1 | X:82517449-82517774 (−) | LTR | L1 PA11 (+), L1 MA2 (+) | Testis | - | MA | [15] |
| Xq23 | X:113140352-113141135 (−) | LTR | L1 (−), XACT (−) | Testis | Pro ° | MA, qRT-PCR | [80] |
| Xq24 | X:120490096-120490859 (+) | LTR | - | Testis | PA | MA | [15] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grandi, N.; Tramontano, E. Type W Human Endogenous Retrovirus (HERV-W) Integrations and Their Mobilization by L1 Machinery: Contribution to the Human Transcriptome and Impact on the Host Physiopathology. Viruses 2017, 9, 162. https://doi.org/10.3390/v9070162
Grandi N, Tramontano E. Type W Human Endogenous Retrovirus (HERV-W) Integrations and Their Mobilization by L1 Machinery: Contribution to the Human Transcriptome and Impact on the Host Physiopathology. Viruses. 2017; 9(7):162. https://doi.org/10.3390/v9070162
Chicago/Turabian StyleGrandi, Nicole, and Enzo Tramontano. 2017. "Type W Human Endogenous Retrovirus (HERV-W) Integrations and Their Mobilization by L1 Machinery: Contribution to the Human Transcriptome and Impact on the Host Physiopathology" Viruses 9, no. 7: 162. https://doi.org/10.3390/v9070162
APA StyleGrandi, N., & Tramontano, E. (2017). Type W Human Endogenous Retrovirus (HERV-W) Integrations and Their Mobilization by L1 Machinery: Contribution to the Human Transcriptome and Impact on the Host Physiopathology. Viruses, 9(7), 162. https://doi.org/10.3390/v9070162