Zika Virus Exhibits Lineage-Specific Phenotypes in Cell Culture, in Aedes aegypti Mosquitoes, and in an Embryo Model

 ,
, 
Abstract
:
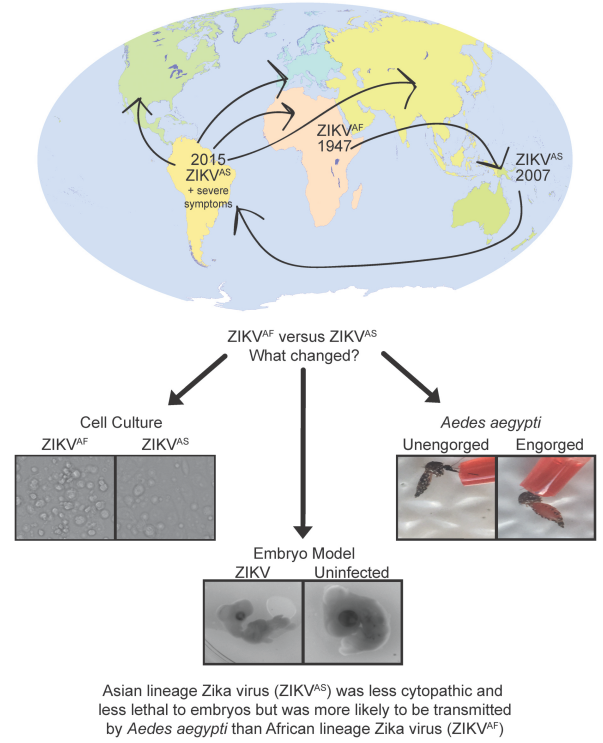
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Zika Virus Isolates
2.3. Determination of Viral Titers
2.4. Viral Replication Curves
2.5. Cell Metabolic Activity/Viability
2.6. Human Interferon-Beta 1A Response
2.7. Intracellular STAT2 Abundance
2.8. Ae. aegypti Rearing and Colony Maintenance
2.9. ZIKV Infection in Ae. aegypti Mosquitoes
2.10. ZIKV Inoculations in Chicken Embryos
2.11. Statistical Analysis
3. Results
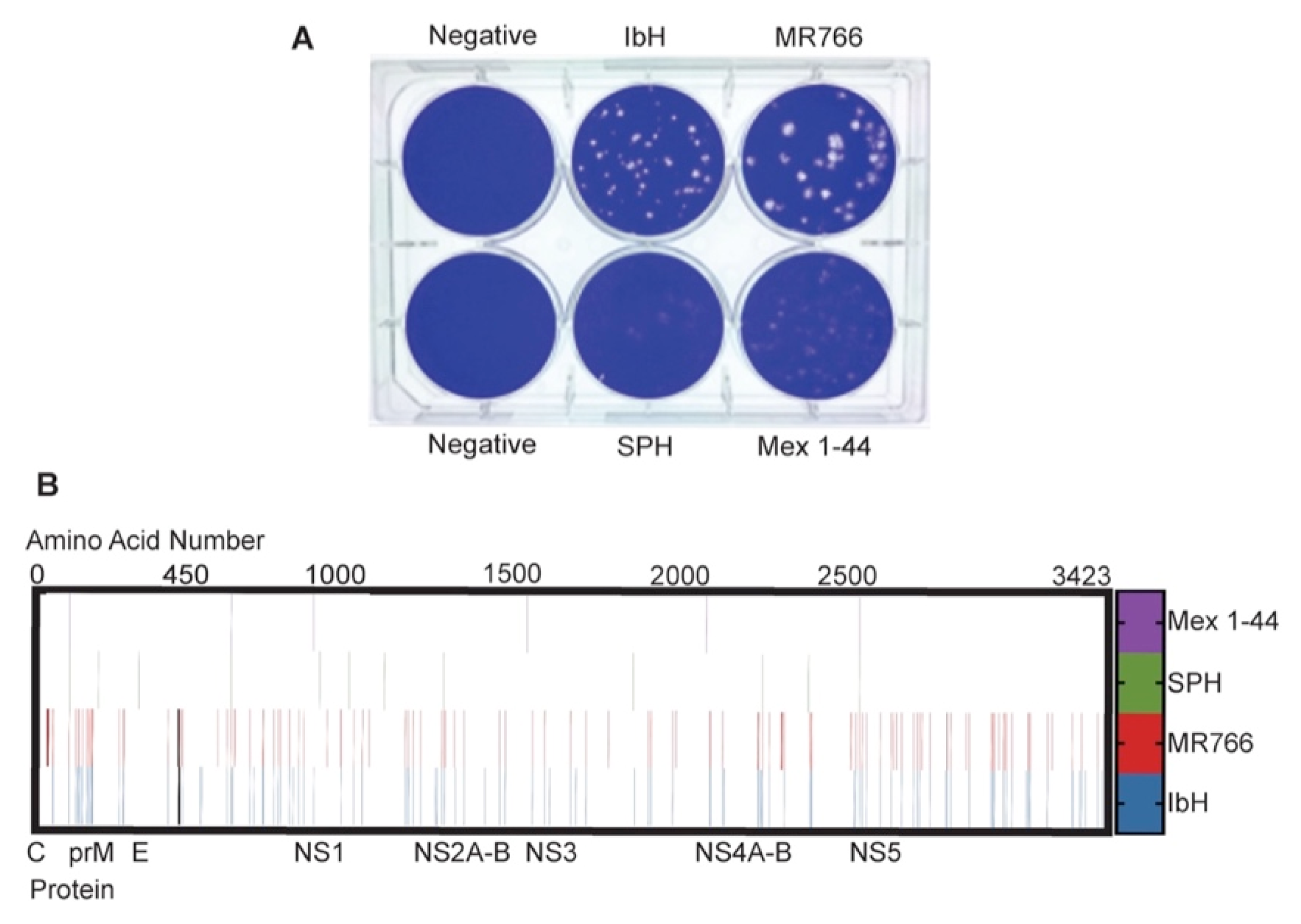
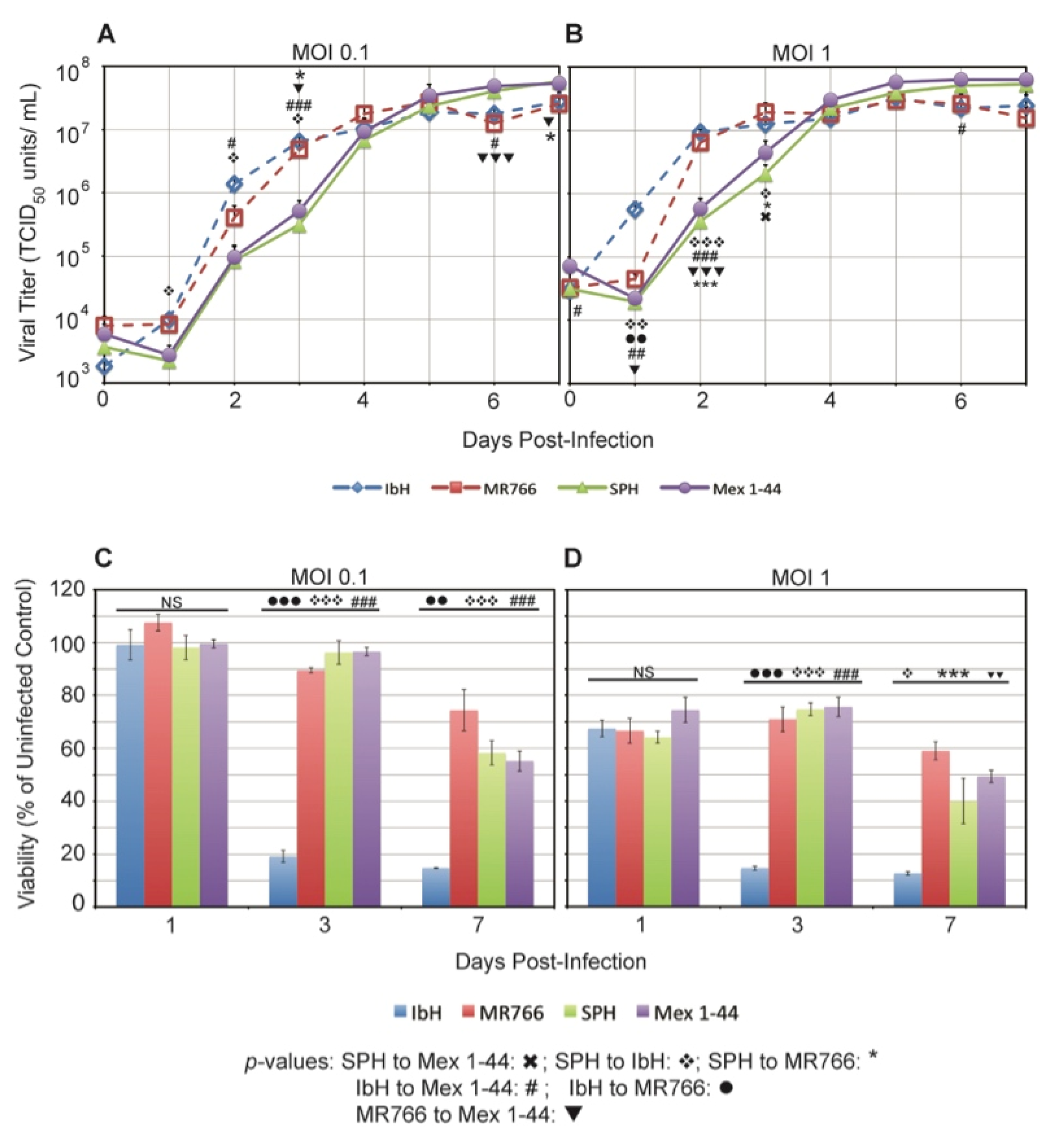
3.1. ZIKV Plaque Morphology Varies Based on Lineage
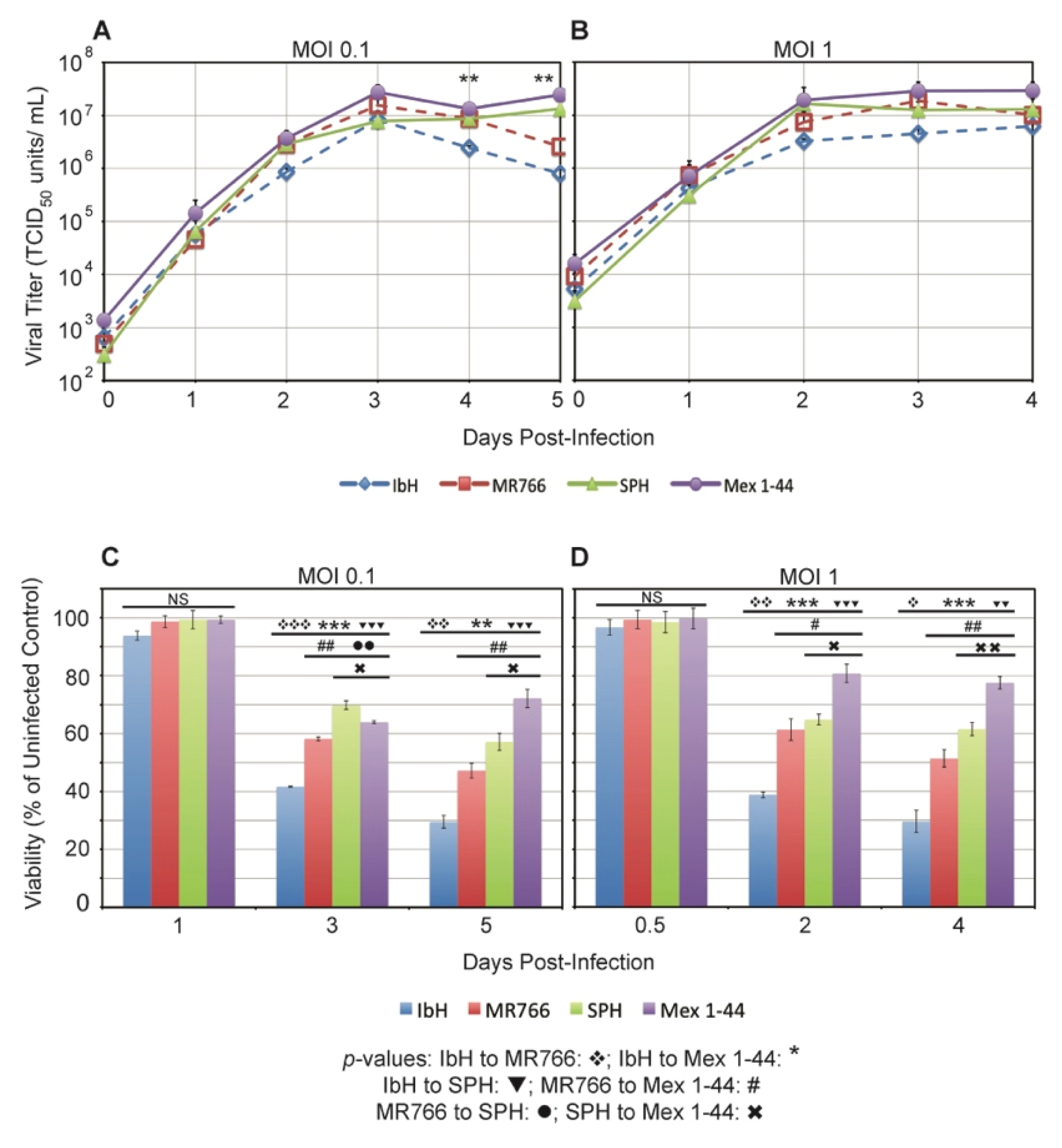
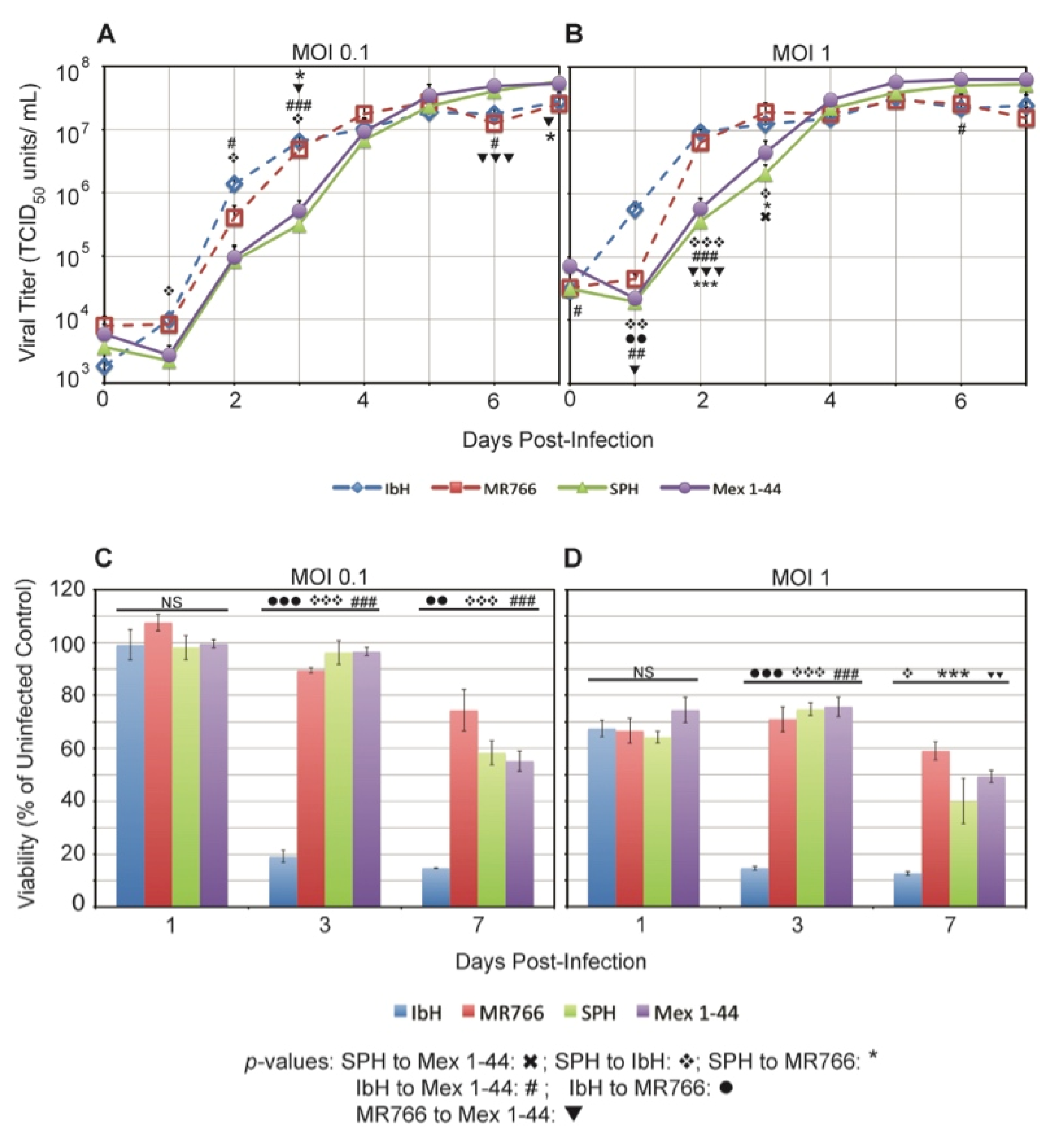
3.2. ZIKV Lineage Impacts Vero Cell Viability
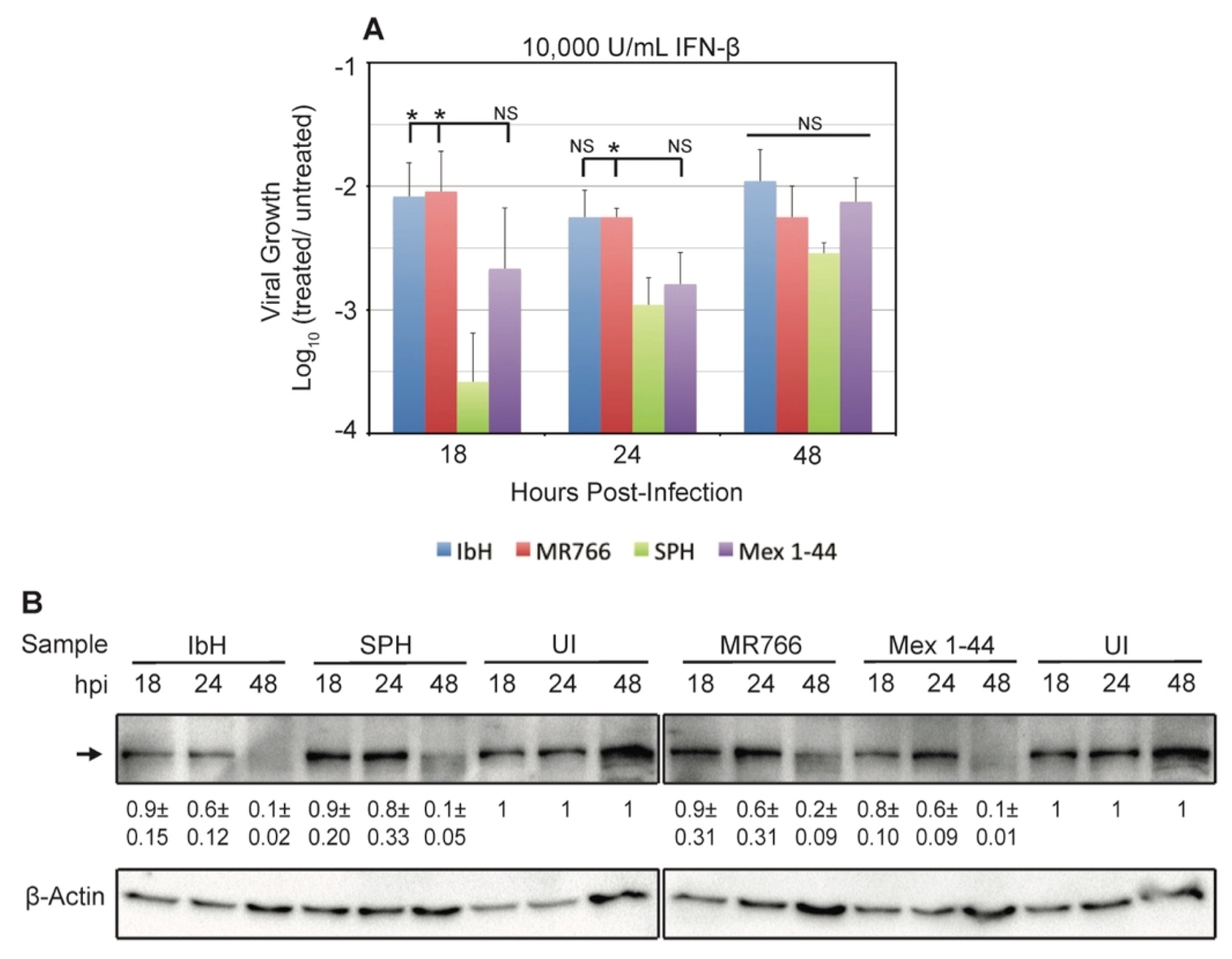
3.3. ZIKV Response to Human IFN-β Treatment Varies by Isolate
3.4. ZIKV Lineage Impacts Growth Kinetics in C6/36 Mosquito Cells
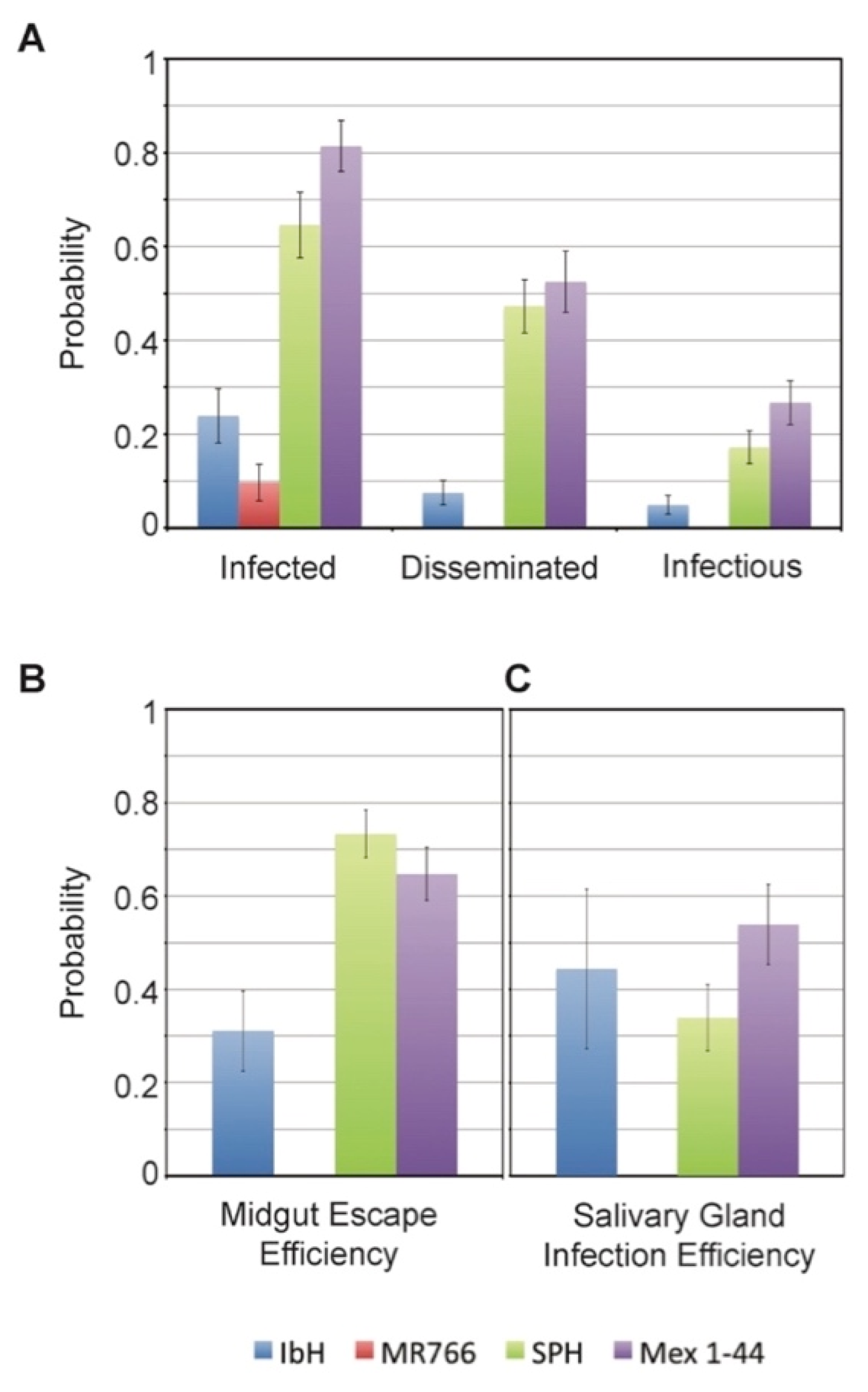
3.5. Vector Competence of Aedes aegypti Depends on ZIKV Lineage
3.6. ZIKV Lineages Cause Differential Mortalities in Chicken Embryos
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dick, G.W.; Kitchen, S.F.; Haddow, A.J. Zika virus. I. Isolations and serological specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Haddow, A.J.; Williams, M.C.; Woodall, J.P.; Simpson, D.I.; Goma, L.K. Twelve isolations of zika virus from aedes (stegomyia) africanus (theobald) taken in and above a uganda forest. Bull. World Health Organ. 1964, 31, 57–69. [Google Scholar] [PubMed]
- Armstrong, P.; Hennessey, M.; Adams, M.; Cherry, C.; Chiu, S.; Harrist, A.; Kwit, N.; Lewis, L.; McGuire, D.O.; Oduyebo, T.; et al. Travel-associated zika virus disease cases among US Residents—United States, January 2015–February 2016. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 286–289. [Google Scholar] [CrossRef] [PubMed]
- Oster, A.M.; Russell, K.; Stryker, J.E.; Friedman, A.; Kachur, R.E.; Petersen, E.E.; Jamieson, D.J.; Cohn, A.C.; Brooks, J.T. Update: Interim guidance for prevention of sexual transmission of zika virus—United States, 2016. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 323–325. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.R.; Chen, T.H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika virus outbreak on yap island, federated states of micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef] [PubMed]
- Musso, D.; Nilles, E.J.; Cao-Lormeau, V.M. Rapid spread of emerging zika virus in the pacific area. Clin. Microbiol. Infect. 2014, 20, O595–O596. [Google Scholar] [CrossRef] [PubMed]
- Shragai, T.; Tesla, B.; Murdock, C.; Harrington, L.C. Zika and chikungunya: Mosquito-borne viruses in a changing world. Ann. N. Y. Acad. Sci. 2017, 1399, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Ledermann, J.P.; Guillaumot, L.; Yug, L.; Saweyog, S.C.; Tided, M.; Machieng, P.; Pretrick, M.; Marfel, M.; Griggs, A.; Bel, M.; et al. Aedes hensilli as a potential vector of chikungunya and zika viruses. PLoS Negl. Trop. Dis. 2014, 8, e3188. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Costa, F.; Garcia-Blanco, M.A.; Ko, A.I.; Ribeiro, G.S.; Saade, G.; Shi, P.Y.; Vasilakis, N. Zika virus: History, emergence, biology, and prospects for control. Antivir. Res. 2016, 130, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Gugliemini, O.; Harber, S.; Harrison, A.; Houle, L.; Ivory, J.; Kersten, S.; Khan, R.; Kim, J.; LeBoa, C.; et al. Environmental and social change drive the explosive emergence of zika virus in the Americas. PLoS Negl. Trop. Dis. 2017, 11, e0005135. [Google Scholar] [CrossRef] [PubMed]
- Pettersson JH-O, E.V.; Seligman, S.J.; Lundkvist, Å.; Falconar, A.K.; Gaunt, M.W.; Musso, D.; Nougairède, A.; Charrel, R.; Gould, E.A.; de Lamballerie, X. How did zika virus emerge in the pacific islands and Latin America? mBio 2016, 7, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Haddow, A.D.; Schuh, A.J.; Yasuda, C.Y.; Kasper, M.R.; Heang, V.; Huy, R.; Guzman, H.; Tesh, R.B.; Weaver, S.C. Genetic characterization of zika virus strains: Geographic expansion of the Asian lineage. PLoS Negl. Trop. Dis. 2012, 6, e1477. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Shi, J.; Wang, J.; Tang, S.; Wang, H.; Hu, Z.; Deng, F. Phylogenetic analysis revealed the central roles of two african countries in the evolution and worldwide spread of zika virus. Virol. Sin. 2016, 31, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Gao, Y.; Han, G.Z. Zika virus: Two or three lineages? Trends Microbiol. 2016, 24, 521–522. [Google Scholar] [CrossRef] [PubMed]
- Faria, N.R.; Azevedo Rdo, S.; Kraemer, M.U.; Souza, R.; Cunha, M.S.; Hill, S.C.; Theze, J.; Bonsall, M.B.; Bowden, T.A.; Rissanen, I.; et al. Zika virus in the Americas: Early epidemiological and genetic findings. Science 2016, 352, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Herrera, B.B.; Chang, C.A.; Hamel, D.J.; Mboup, S.; Ndiaye, D.; Imade, G.; Okpokwu, J.; Agbaji, O.; Bei, A.K.; Kanki, P.J. Continued transmission of zika virus in humans in West Africa, 1992–2016. J. Infect. Dis 2017, 215, 1546–1550. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.H.; Duval, C.W. Studies upon the etiology of dengue fever: I. Experimental transmission to the lower animal. J. Exp. Med. 1924, 40, 817–833. [Google Scholar] [CrossRef] [PubMed]
- CDC. Chikungunya Virus: Symptoms, Diagnosis, & Treatment. Available online: https://www.cdc.gov/chikungunya/symptoms/index.html (accessed on 11 April 2017).
- Moulin, E.; Selby, K.; Cherpillod, P.; Kaiser, L.; Boillat-Blanco, N. Simultaneous outbreaks of dengue, chikungunya and zika virus infections: Diagnosis challenge in a returning traveller with nonspecific febrile illness. New Microbes New Infect. 2016, 11, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Schuler-Faccini, L.; Ribeiro, E.M.; Feitosa, I.M.; Horovitz, D.D.; Cavalcanti, D.P.; Pessoa, A.; Doriqui, M.J.; Neri, J.I.; Neto, J.M.; Wanderley, H.Y.; et al. Possible association between zika virus infection and microcephaly—Brazil, 2015. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 59–62. [Google Scholar] [CrossRef] [PubMed]
- CDC. CDC Concludes zika Causes Microcephaly and Other Birth Defects. Available online: https://www.cdc.gov/media/releases/2016/s0413-zika-microcephaly.html (accessed on 24 October 2016).
- Rasmussen, S.A.; Jamieson, D.J.; Honein, M.A.; Petersen, L.R. Zika virus and birth defects—Reviewing the evidence for causality. N. Engl. J. Med. 2016, 374, 1981–1987. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Huang, X.Y.; Liu, Z.Y.; Zhang, F.; Zhu, X.L.; Yu, J.Y.; Ji, X.; Xu, Y.P.; Li, G.; Li, C.; et al. A single mutation in the PRM protein of zika virus contributes to fetal microcephaly. Science 2017, 358, 933–936. [Google Scholar] [CrossRef] [PubMed]
- Brasil, P.; Sequeira, P.C.; Freitas, A.D.; Zogbi, H.E.; Calvet, G.A.; de Souza, R.V.; Siqueira, A.M.; de Mendonca, M.C.; Nogueira, R.M.; de Filippis, A.M.; et al. Guillain-barre syndrome associated with zika virus infection. Lancet 2016, 387, 1482. [Google Scholar] [CrossRef]
- Broutet, N.; Krauer, F.; Riesen, M.; Khalakdina, A.; Almiron, M.; Aldighieri, S.; Espinal, M.; Low, N.; Dye, C. Zika virus as a cause of neurologic disorders. N. Engl. J. Med. 2016, 374, 1506–1509. [Google Scholar] [CrossRef] [PubMed]
- Morrison, T.E.; Diamond, M.S. Animal models of zika virus infection, pathogenesis, and immunity. J. Virol. 2017, 91, e00009-17. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.R.; Quicke, K.M.; Maddur, M.S.; O'Neal, J.T.; McDonald, C.E.; Fedorova, N.B.; Puri, V.; Shabman, R.S.; Pulendran, B.; Suthar, M.S. Zika virus antagonizes type I interferon responses during infection of human dendritic cells. PLoS Pathog. 2017, 13, e1006164. [Google Scholar] [CrossRef] [PubMed]
- Grant, A.; Ponia, S.S.; Tripathi, S.; Balasubramaniam, V.; Miorin, L.; Sourisseau, M.; Schwarz, M.C.; Sanchez-Seco, M.P.; Evans, M.J.; Best, S.M.; et al. Zika virus targets human STAT2 to inhibit type I interferon signaling. Cell. Host Microbe 2016, 19, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Hamel, R.; Dejarnac, O.; Wichit, S.; Ekchariyawat, P.; Neyret, A.; Luplertlop, N.; Perera-Lecoin, M.; Surasombatpattana, P.; Talignani, L.; Thomas, F.; et al. Biology of zika virus infection in human skin cells. J. Virol. 2015, 89, 8880–8896. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, M.A.; Yunusov, D.; Balaraman, V.; Alexenko, A.P.; Yabe, S.; Verjovski-Almeida, S.; Schust, D.J.; Franz, A.W.; Sadovsky, Y.; Ezashi, T.; et al. Vulnerability of primitive human placental trophoblast to zika virus. Proc. Natl. Acad. Sci. USA 2017, 114, E1587–E1596. [Google Scholar] [CrossRef] [PubMed]
- McGrath, E.L.; Rossi, S.L.; Gao, J.; Widen, S.G.; Grant, A.C.; Dunn, T.J.; Azar, S.R.; Roundy, C.M.; Xiong, Y.; Prusak, D.J.; et al. Differential responses of human fetal brain neural stem cells to zika virus infection. Stem Cell Rep. 2017, 8, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Simonin, Y.; Loustalot, F.; Desmetz, C.; Foulongne, V.; Constant, O.; Fournier-Wirth, C.; Leon, F.; Molès, J.-P.; Goubaud, A.; Lemaitre, J.-M.; et al. Zika virus strains potentially display different infectious profiles in human neural cells. EBioMedicine 2016, 12, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Mossenta, M.; Marchese, S.; Poggianella, M.; Slon Campos, J.L.; Burrone, O.R. Role of N-glycosylation on zika virus E protein secretion, viral assembly and infectivity. Biochem. Biophys. Res. Commun. 2017, 492, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Aliota, M.T.; Dudley, D.M.; Newman, C.M.; Mohr, E.L.; Gellerup, D.D.; Breitbach, M.E.; Buechler, C.R.; Rasheed, M.N.; Mohns, M.S.; Weiler, A.M.; et al. Heterologous protection against Asian zika virus challenge in rhesus macaques. PLoS Negl. Trop. Dis. 2016, 10, e0005168. [Google Scholar] [CrossRef] [PubMed]
- Beasley, D.W.; Whiteman, M.C.; Zhang, S.; Huang, C.Y.; Schneider, B.S.; Smith, D.R.; Gromowski, G.D.; Higgs, S.; Kinney, R.M.; Barrett, A.D. Envelope protein glycosylation status influences mouse neuroinvasion phenotype of genetic lineage 1 West Nile virus strains. J. Virol. 2005, 79, 8339–8347. [Google Scholar] [CrossRef] [PubMed]
- Hamel, R.; Ferraris, P.; Wichit, S.; Diop, F.; Talignani, L.; Pompon, J.; Garcia, D.; Liegeois, F.; Sall, A.A.; Yssel, H.; et al. African and Asian zika virus strains differentially induce early antiviral responses in primary human astrocytes. Infect. Genet. Evol. 2017, 49, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Roundy, C.M.; Azar, S.R.; Rossi, S.L.; Huang, J.H.; Leal, G.; Yun, R.; Fernandez-Salas, I.; Vitek, C.J.; Paploski, I.A.; Kitron, U.; et al. Variation in Aedes aegypti mosquito competence for zika virus transmission. Emerg. Infect. Dis. 2017, 23, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Chouin-Carneiro, T.; Vega-Rua, A.; Vazeille, M.; Yebakima, A.; Girod, R.; Goindin, D.; Dupont-Rouzeyrol, M.; Lourenco-de-Oliveira, R.; Failloux, A.B. Differential susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to zika virus. PLoS Negl. Trop. Dis. 2016, 10, e0004543. [Google Scholar] [CrossRef] [PubMed]
- Hart, C.E.; Roundy, C.M.; Azar, S.R.; Huang, J.H.; Yun, R.; Reynolds, E.; Leal, G.; Nava, M.R.; Vela, J.; Stark, P.M.; et al. Zika virus vector competency of mosquitoes, gulf coast, United States. Emerg. Infect. Dis. 2017, 23, 559–560. [Google Scholar] [CrossRef] [PubMed]
- Lazear, H.M.; Govero, J.; Smith, A.M.; Platt, D.J.; Fernandez, E.; Miner, J.J.; Diamond, M.S. A mouse model of zika virus pathogenesis. Cell. Host Microbe 2016, 19, 720–730. [Google Scholar] [CrossRef] [PubMed]
- Cugola, F.R.; Fernandes, I.R.; Russo, F.B.; Freitas, B.C.; Dias, J.L.; Guimaraes, K.P.; Benazzato, C.; Almeida, N.; Pignatari, G.C.; Romero, S.; et al. The Brazilian zika virus strain causes birth defects in experimental models. Nature 2016, 534, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Koide, F.; Goebel, S.; Snyder, B.; Walters, K.B.; Gast, A.; Hagelin, K.; Kalkeri, R.; Rayner, J. Development of a zika virus infection model in cynomolgus macaques. Front. Microbiol 2016, 7, 2028. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.M. Studies on certain viruses isolated in the tropics of Africa and South America; their growth and behavior in the embryonated hen egg. J. Immunol. 1952, 68, 473–494. [Google Scholar] [PubMed]
- Goodfellow, F.T.; Tesla, B.; Simchick, G.; Zhao, Q.; Hodge, T.; Brindley, M.A.; Stice, S.L. Zika virus induced mortality and microcephaly in chicken embryos. Stem Cells Dev. 2016, 25, 1691–1697. [Google Scholar] [CrossRef] [PubMed]
- Guerbois, M.; Fernandez-Salas, I.; Azar, S.R.; Danis-Lozano, R.; Alpuche-Aranda, C.M.; Leal, G.; Garcia-Malo, I.R.; Diaz-Gonzalez, E.E.; Casas-Martinez, M.; Rossi, S.L.; et al. Outbreak of zika virus infection, Chiapas state, Mexico, 2015, and first confirmed transmission by Aedes aegypti mosquitoes in the Americas. J. Infect. Dis. 2016, 214, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, M.A. Determination of 50% endpoint titer using a simple formula. World J. Virol. 2016, 5, 85–86. [Google Scholar] [CrossRef] [PubMed]
- Ebel, G.D.; Fitzpatrick, K.A.; Lim, P.Y.; Bennett, C.J.; Deardorff, E.R.; Jerzak, G.V.; Kramer, L.D.; Zhou, Y.; Shi, P.Y.; Bernard, K.A. Nonconsensus West Nile virus genomes arising during mosquito infection suppress pathogenesis and modulate virus fitness in vivo. J. Virol. 2011, 85, 12605–12613. [Google Scholar] [CrossRef] [PubMed]
- Emeny, J.M.; Morgan, M.J. Regulation of the interferon system: Evidence that vero cells have a genetic defect in interferon production. J. Gen. Virol. 1979, 43, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Osada, N.; Kohara, A.; Yamaji, T.; Hirayama, N.; Kasai, F.; Sekizuka, T.; Kuroda, M.; Hanada, K. The genome landscape of the African green monkey kidney-derived vero cell line. DNA Res. 2014, 21, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Frumence, E.; Roche, M.; Krejbich-Trotot, P.; El-Kalamouni, C.; Nativel, B.; Rondeau, P.; Missé, D.; Gadea, G.; Viranaicken, W.; Desprès, P. The south pacific epidemic strain of zika virus replicates efficiently in human epithelial A549 cells leading to IFN-β production and apoptosis induction. Virology 2016, 493, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Aliota, M.T.; Caine, E.A.; Walker, E.C.; Larkin, K.E.; Camacho, E.; Osorio, J.E. Characterization of lethal zika virus infection in AG129 mice. PLoS Negl. Trop. Dis. 2016, 10, e0004682. [Google Scholar] [CrossRef] [PubMed]
- Dowall, S.D.; Graham, V.A.; Rayner, E.; Atkinson, B.; Hall, G.; Watson, R.J.; Bosworth, A.; Bonney, L.C.; Kitchen, S.; Hewson, R. A susceptible mouse model for zika virus infection. PLoS Negl. Trop. Dis. 2016, 10, e0004658. [Google Scholar] [CrossRef] [PubMed]
- Miner, J.J.; Cao, B.; Govero, J.; Smith, A.M.; Fernandez, E.; Cabrera, O.H.; Garber, C.; Noll, M.; Klein, R.S.; Noguchi, K.K.; et al. Zika virus infection during pregnancy in mice causes placental damage and fetal demise. Cell 2016, 165, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Yockey, L.J.; Varela, L.; Rakib, T.; Khoury-Hanold, W.; Fink, S.L.; Stutz, B.; Szigeti-Buck, K.; van den Pol, A.; Lindenbach, B.D.; Horvath, T.L.; et al. Vaginal exposure to zika virus during pregnancy leads to fetal brain infection. Cell 2016, 166, 1247–1256.e4. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Jordan, J.L.; Laurent-Rolle, M.; Ashour, J.; Martinez-Sobrido, L.; Ashok, M.; Lipkin, W.I.; Garcia-Sastre, A. Inhibition of α/β interferon signaling by the NS4B protein of flaviviruses. J. Virol. 2005, 79, 8004–8013. [Google Scholar] [CrossRef] [PubMed]
- Daffis, S.; Suthar, M.S.; Gale, M., Jr.; Diamond, M.S. Measure and countermeasure: Type I IFN (IFN-α/β) antiviral response against West Nile virus. J. Innate Immun. 2009, 1, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. 1951. Dev. Dyn. 1992, 195, 231–272. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.; Balasubramaniam, V.R.; Brown, J.A.; Mena, I.; Grant, A.; Bardina, S.V.; Maringer, K.; Schwarz, M.C.; Maestre, A.M.; Sourisseau, M.; et al. A novel zika virus mouse model reveals strain specific differences in virus pathogenesis and host inflammatory immune responses. PLoS Pathog. 2017, 13, e1006258. [Google Scholar] [CrossRef] [PubMed]
- Widman, D.G.; Young, E.; Yount, B.L.; Plante, K.S.; Gallichotte, E.N.; Carbaugh, D.L.; Peck, K.M.; Plante, J.; Swanstrom, J.; Heise, M.T.; et al. A reverse genetics platform that spans the zika virus family tree. MBio 2017, 8, e02014-16. [Google Scholar] [CrossRef] [PubMed]
- Pompon, J.; Morales-Vargas, R.; Manuel, M.; Huat Tan, C.; Vial, T.; Hao Tan, J.; Sessions, O.M.; Vasconcelos, P.D.C.; Ng, L.C.; Misse, D. A zika virus from America is more efficiently transmitted than an Asian virus by Aedes aegypti mosquitoes from Asia. Sci Rep. 2017, 7, 1215. [Google Scholar] [CrossRef] [PubMed]
- Hanna, S.L.; Pierson, T.C.; Sanchez, M.D.; Ahmed, A.A.; Murtadha, M.M.; Doms, R.W. N-linked glycosylation of West Nile virus envelope proteins influences particle assembly and infectivity. J. Virol. 2005, 79, 13262–13274. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Leang, S.K.; Davidson, A.; Lobigs, M. Both E protein glycans adversely affect dengue virus infectivity but are beneficial for virion release. J. Virol. 2010, 84, 5171–5180. [Google Scholar] [CrossRef] [PubMed]
- Donald, C.L.; Brennan, B.; Cumberworth, S.L.; Rezelj, V.V.; Clark, J.J.; Cordeiro, M.T.; Freitas de Oliveira Franca, R.; Pena, L.J.; Wilkie, G.S.; da Silva Filipe, A.; et al. Full genome sequence and sfRNA interferon antagonist activity of zika virus from Recife, Brazil. PLoS Negl. Trop. Dis. 2016, 10, e0005048. [Google Scholar] [CrossRef] [PubMed]
- Cumberworth, S.L.; Clark, J.J.; Kohl, A.; Donald, C.L. Inhibition of type I interferon induction and signalling by mosquito-borne flaviviruses. Cell. Microbiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Manokaran, G.; Finol, E.; Wang, C.; Gunaratne, J.; Bahl, J.; Ong, E.Z.; Tan, H.C.; Sessions, O.M.; Ward, A.M.; Gubler, D.J.; et al. Dengue subgenomic RNA binds TRIM25 to inhibit interferon expression for epidemiological fitness. Science 2015, 350, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, S.; Starmer, W.T. Possible roles of new mutations shared by Asian and American zika viruses. Mol. Biol. Evol. 2017, 34, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Blaney, J.E., Jr.; Sathe, N.S.; Goddard, L.; Hanson, C.T.; Romero, T.A.; Hanley, K.A.; Murphy, B.R.; Whitehead, S.S. Dengue virus type 3 vaccine candidates generated by introduction of deletions in the 3′ untranslated region (3′-UTR) or by exchange of the DENV-3 3′-UTR with that of DENV-4. Vaccine 2008, 26, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Weger-Lucarelli, J.; Ruckert, C.; Chotiwan, N.; Nguyen, C.; Garcia Luna, S.M.; Fauver, J.R.; Foy, B.D.; Perera, R.; Black, W.C.; Kading, R.C.; et al. Vector competence of American mosquitoes for three strains of zika virus. PLoS Negl. Trop. Dis. 2016, 10, e0005101. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, J.; Du, S.; Shan, C.; Nie, K.; Zhang, R.; Li, X.F.; Zhang, R.; Wang, T.; Qin, C.F.; et al. Evolutionary enhancement of zika virus infectivity in Aedes aegypti mosquitoes. Nature 2017, 545, 482–486. [Google Scholar] [PubMed]
- Karpala, A.J.; Bagnaud-Baule, A.; Goossens, K.E.; Lowenthal, J.W.; Bean, A.G. Ontogeny of the interferon system in chickens. J. Reprod. Immunol. 2012, 94, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Liniger, M.; Summerfield, A.; Zimmer, G.; McCullough, K.C.; Ruggli, N. Chicken cells sense influenza A virus infection through MDA5 and CARDIF signaling involving lgp2. J. Virol. 2012, 86, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Jordan, J.L.; Fredericksen, B.L. How flaviviruses activate and suppress the interferon response. Viruses 2010, 2, 676–691. [Google Scholar] [CrossRef] [PubMed]
- Nasirudeen, A.M.; Wong, H.H.; Thien, P.; Xu, S.; Lam, K.P.; Liu, D.X. RIG-I, MDA5 and TLR3 synergistically play an important role in restriction of dengue virus infection. PLoS Negl. Trop. Dis. 2011, 5, e926. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Genbank Accession Number | First Reference | Lineage | Isolation Location | Isolate Source | Passage Number in This Study |
|---|---|---|---|---|---|---|
| MR766 | LC002520 | Dick et al., 1952 [1] | African | Uganda, 1947 | Rhesus macaque | >100 |
| IbH 30656 | KU963574 | Haddow et al., 2012 [12] | African | Nigeria, 1968 | Human patient | >25 |
| SPH | KU321639.1 | Faria et al., 2016 [15] | Asian | Brazil, 2015 | Human patient | <10 |
| Mex 1-44 | KX856011.1 | Guerbois et al., 2016 [45] | Asian | Mexico, 2015 | Aedes aegypti | <10 |
| Probability of Infection | Probability of Dissemination | Probability of Infectiousness | Efficiency of Midgut Escape | Efficiency of Salivary Gland Invasion | |
|---|---|---|---|---|---|
| ZIKV isolate | F = 34.77 d.f. = 3 p < 0.0001 | F = 14.71 d.f. = 3 p < 0.0001 | F = 8.64 d.f. = 3 p < 0.0001 | F = 6.00 d.f. = 3 p = 0.001 | F = 1.794 d.f. = 2 p = 0.171 |
| Tukey HSD | All pairwise comparisons, p < 0.0001 | Mex 1-44 vs. SPH, N.S.; all remaining pairwise comparisons, p < 0.0001 | IbH vs. MR766, N.S.; Mex 1-44 vs. SPH, N.S.; all remaining pairwise comparisons, p < 0.0001 | Mex 1-44 vs. SPH, N.S.; all remaining pairwise comparisons, p < 0.0001 | All pairwise comparisons are N.S. |
| Isolate | Trial | Blood Meal Titer (log10 Plaque Forming Units (PFU)/mL) | Total Blood-Fed (n) | Percent Infected Bodies (n) | Percent Infected Heads (n) | Percent Infected Saliva (n) |
|---|---|---|---|---|---|---|
| IbH 30656 | 1 | 6.0 | 48 | 22.92 (11) | 9.09 (1) | 0.00 (0) |
| 2 | 5.9 | 31 | 6.45 (2) | 50.00 (1) | 0.00 (0) | |
| 3 | 5.9 | 39 | 46.15 (18) | 44.44 (8) | 50.00 (4) | |
| MR766 | 1 | 6.2 | 46 | 8.70 (4) | 0.00 (0) | 0.00 (0) |
| 2 | 6.2 | 47 | 8.51 (4) | 0.00 (0) | 0.00 (0) | |
| SPH | 1 | 5.8 | 36 | 58.33 (21) | 52.38 (11) | 18.18 (2) |
| 2 | 5.8 | 37 | 70.27 (26) | 92.73 (24) | 45.83 (11) | |
| 3 | 6.6 | 43 | 65.12 (28) | 71.43 (20) | 35.00 (7) | |
| Mex 1-44 | 1 | 6.8 | 47 | 93.62 (44) | 81.82 (36) | 50.00 (18) |
| 2 | 6.7 | 43 | 62.79 (27) | 37.04 (10) | 60.00 (6) |
| Isolate | Infection | Injection Titer (Viral Particles/Egg) |
|---|---|---|
| IbH 30656 | Low dose | 7 |
| High dose | 72 | |
| MR766 | Low dose | 5 |
| High dose | 54 | |
| SPH | Low dose | 9 |
| High dose | 94 | |
| Mex 1-44 | Low dose | 17 |
| High dose | 168 |
| Isolate | Infection | Percent ZIKV-Positive Embryos | Average Titer (ZIKV-Positive Embryos) |
|---|---|---|---|
| IbH 30656 | Low dose | 85.71 | 8.86 × 107 |
| High dose | 94.44 | 7.58 × 108 | |
| MR766 | Low dose | 69.23 | 1.42 × 108 |
| High dose | 100 | 2.92 × 109 | |
| SPH | Low dose | 22.22 | 2.49 × 108 |
| High dose | 44.44 | 9.42 × 108 | |
| Mex 1-44 | Low dose | 30 | 6.50 × 106 |
| High dose | 88.89 | 8.78 × 107 |
| Low Titer Injection p-Values | High Titer Injection p-Values | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| IbH | MR766 | SPH | Mex 1-44 | IbH | MR766 | SPH | Mex 1-44 | ||
| MR766 | 0.36 | MR766 | 0.5521 | ||||||
| SPH | 0.0194 | 0.2595 | SPH | 0.0001 | 0.0001 | ||||
| Mex 1-44 | 0.0339 | 0.3275 | 0.778 | Mex 1-44 | 0.0519 | 0.0245 | 0.0437 | ||
| Sham | 0.0001 | 0.0036 | 0.0435 | 0.0215 | Sham | 0.0001 | 0.0001 | 0.0001 | 0.0001 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Willard, K.A.; Demakovsky, L.; Tesla, B.; Goodfellow, F.T.; Stice, S.L.; Murdock, C.C.; Brindley, M.A. Zika Virus Exhibits Lineage-Specific Phenotypes in Cell Culture, in Aedes aegypti Mosquitoes, and in an Embryo Model. Viruses 2017, 9, 383. https://doi.org/10.3390/v9120383
Willard KA, Demakovsky L, Tesla B, Goodfellow FT, Stice SL, Murdock CC, Brindley MA. Zika Virus Exhibits Lineage-Specific Phenotypes in Cell Culture, in Aedes aegypti Mosquitoes, and in an Embryo Model. Viruses. 2017; 9(12):383. https://doi.org/10.3390/v9120383
Chicago/Turabian StyleWillard, Katherine A., Leah Demakovsky, Blanka Tesla, Forrest T. Goodfellow, Steven L. Stice, Courtney C. Murdock, and Melinda A. Brindley. 2017. "Zika Virus Exhibits Lineage-Specific Phenotypes in Cell Culture, in Aedes aegypti Mosquitoes, and in an Embryo Model" Viruses 9, no. 12: 383. https://doi.org/10.3390/v9120383
APA StyleWillard, K. A., Demakovsky, L., Tesla, B., Goodfellow, F. T., Stice, S. L., Murdock, C. C., & Brindley, M. A. (2017). Zika Virus Exhibits Lineage-Specific Phenotypes in Cell Culture, in Aedes aegypti Mosquitoes, and in an Embryo Model. Viruses, 9(12), 383. https://doi.org/10.3390/v9120383