
The Immune Response to Astrovirus Infection

Biology Department, Drake University, Des Moines, IA 50311, USA
Viruses 2017, 9(1), 1; https://doi.org/10.3390/v9010001
Submission received: 30 November 2016
/
Revised: 20 December 2016
/
Accepted: 28 December 2016
/
Published: 30 December 2016
(This article belongs to the Special Issue Astroviruses)
Abstract
:Astroviruses are one of the leading causes of pediatric gastroenteritis worldwide and are clinically importantly pathogens in the elderly and immunocompromised populations. Although the use of cell culture systems and small animal models have enhanced our understanding of astrovirus infection and pathogenesis, little is known about the immune response to astrovirus infection. Studies from humans and animals suggest that adaptive immunity is important in restricting classic and novel astrovirus infections, while studies from animal models and cell culture systems suggest that an innate immune system plays a role in limiting astrovirus replication. The relative contribution of each arm of the immune system in restricting astrovirus infection remains unknown. This review summarizes our current understanding of the immune response to astrovirus infection and highlights some of the key questions that stem from these studies. A full understanding of the immune response to astrovirus infection is required to be able to treat and control astrovirus-induced gastroenteritis.
1. Introduction
Astroviruses (AstV) are small, nonenveloped, RNA viruses that are a major cause of gastroenteritis in infants, immunocompromised people, and the elderly, and they also cause disease in mammals and birds [1,2,3,4,5,6,7,8,9]. Despite the disease burden, little is known about the immune response to astrovirus infection. Human clinical studies have demonstrated that an antibody-mediated response may be responsible for limiting astrovirus infection and clinical disease. Recent work using small animal models and cell culture systems have revealed an important role in the innate immune response in restricting astrovirus replication and pathogenesis. This review will summarize the current knowledge of the innate and adaptive immune responses to astrovirus infection using studies of humans, small animal models, and cell culture systems and will discuss how astroviruses evade the immune system. This review will also highlight the increasing reports of astroviruses as possible causes of central nervous system disease, especially in immunocompromised individuals. Finally, we will conclude with unanswered questions, future studies, and how the use of a newly developed mouse model can enhance our understanding of the immune response to astrovirus infection, and how these responses play a role in astrovirus-induced disease.
2. Astrovirus Infection
Astrovirus infection begins by binding to an unidentified receptor(s) on epithelial cells in the intestine after fecal-oral transmission [10,11] and enters cells via clathrin-mediated endocytosis [12]. After acidification of the endosome, endosome rupture, [12] and viral uncoating, two nonstructural proteins are translated from the single stranded, positive-sense genomic RNA from open reading frames (ORFs) 1a and 1b [13,14]. These polyproteins are cleaved into the non-structural proteins required for transcription and viral replication. Negative-strand RNA is produced from the genomic strand, and transcription of the negative-strand yields the genomic and subgenomic RNA [15]. The structural proteins encoded in the third ORF (ORF2) are expressed from the subgenomic RNA [13,14]. New astrovirus particles have been observed on double membranes likely serving as the site for replication and assembly [16,17,18,19,20]. After assembly, the progeny virions exit the cell through a non-lytic mechanism promoted by caspase activation [21,22].
Although astroviruses are considered gastrointestinal pathogens, viral RNA and infectious viral particles have been recovered from extraintestinal organs in both animals and humans (Table 1). In the current small animal model, turkey poults, turkey astrovirus type 2 (TAstV2) was found in bursa, thymus, spleen, kidney, liver, skeletal muscle, marrow, pancreas and plasma by reverse transcription polymerase chain reaction (RT-PCR), immunofluorescence, and virus isolation [6]. However, these tissues were negative by in situ hybridization using a riboprobe to the capsid gene, suggesting that the virus only replicates in the intestines [6,23]. Whether astrovirus localization to these extra-intestinal tissues leads to pathogenesis remains to be directly determined, and astroviruses have been implicated as a cause of neurological and other diseases. Examples in animals implicate astroviruses as the cause of hepatitis in ducks, causing encephalomyelitis in minks [24,25,26] and encephalitis in cattle [27,28,29]. Identification of astroviruses found in extra-intestinal tissues in humans will be discussed later in this review.
3. Adaptive Immunity
The innate immune response provides signals to recruit the adaptive immune response, which controls viral infection at later times during an infection. This response is pathogen specific and has a memory component that induces a more rapid and robust response following a second infection with the same pathogen. The adaptive immune response contains two arms: the humoral, or antibody-mediated/B cell response, and the cell-mediated response, which involves antigen-specific cytotoxic T cells. Although the findings between the current turkey model and the newly emerging mouse model yielded conflicting results, the reports showing the biphasic age distribution as well as immunocompromised individuals have formed a general conclusion that the adaptive immune system is a major component in controlling astrovirus disease (reviewed in [39]).
Early studies in the turkey poult model found little adaptive immune responses after TAstV-2 infection [40]. Koci et al. found very low levels of IgG in the serum and IgA in the bile 21 dpi [40]. Additionally, the CD4 to CD8 T cell ratios did not change at 5, 9, and 16 dpi when comparing TAstV-2 infected poults to uninfected poults [40], demonstrating that TAstV-2 infection is a poor inducer of adaptive immune responses.
Studies using the newly emerging mouse model and clinical studies in humans demonstrate that the adaptive immune response is key in controlling astrovirus infection and disease. Experimental infection of recombination activating gene 1 (rag1) gene knockout mice (Rag1−/−), which lack mature B cells or T cells [41], had higher (2 logs) levels of murine astrovirus (MuAstV) RNA shed in their feces compared to wild type mice at 14 dpi [42]. The kidneys, liver, and mesenteric lymph nodes were positive for astrovirus RNA, whereas infected wild type mice were negative for astrovirus RNA in these organs [42]. Rag1−/− mice also contained 3–4 logs higher astrovirus RNA levels in the intestines compared to wild type controls [42]. These data indicate that the adaptive immune response is important in controlling astrovirus infection. Similar to the studies with interferon (IFN)-α receptor knockout (IFNaR−/−) mice by Marvin et al., the Rag1−/− mice were positive for astrovirus prior to experimental inoculation [42]. Therefore, a complete understanding in the adaptive immune response, and how it limits astrovirus replication, cannot be determined until MuAstV-negative Rag1−/− mice are generated.
Studies in immunocompetent humans also show that 70% of healthy adults have antibodies against astroviruses [43], indicating that an adaptive immune response is mounted in humans. In earlier clinical studies using human volunteers, the majority of subjects that had no or only mild clinical signs had developed anti-astrovirus antibodies [5,44]. Conversely, subjects that exhibited more severe disease did not develop anti-astrovirus antibodies [44]. Other recent studies have found astrovirus antibodies against astroviruses of other animal species in the absence of disease [45,46]. For example, humans that work closely with turkeys develop antibodies to TAstV [45].
Antibodies that recognize the spike domains on the human astrovirus spike, the domain of the capsid involved in astrovirus binding [47], and neutralized virus activity have been identified [48,49]. However, the development of anti-HAstV therapies has been hampered by the gap in knowledge of neutralizing antibodies epitopes on HAstV surfaces. Bogdanoff et al. mapped the neutralizing epitopes on the HAstV-2 with a neutralizing monoclonal antibody to the spike domain, which prevented spike binding to Caco2 cells [47]. The solving of the crystal structures of HAstV-1 and HAstV-8 capsids and spike domains, and the HAstV-2 and TAstV-2 spike domains can advance our understanding of anti-astrovirus antibody binding [47,50,51,52,53]. These structures are crucial for future studies to develop vaccines and antibody therapy prevention and treatment of astrovirus disease.
Increasing reports of astrovirus infection in extra-intestinal tissues in immunocompromised patients highlight the role of the adaptive immune response during astrovirus infection. Several astrovirus isolates have been identified as potential causes of encephalitis in immunocompromised human patients (Table 1). For example, astrovirus RNA was found in the central nervous system of a child with X-linked agammaglobulinemia who died of encephalitis [35], and have been thought to be the cause of encephalopathy after bone marrow or hematopoietic stem cell transplantation (HSCT) [36,37]. Astrovirus has also been detected in nasopharyngeal secretions and serum and plasma of severe immunodeficiency (SCID) patients after HSCT [32], and detection of astrovirus in the blood has been associated with fever [33]. As the number of reports of astrovirus detection in extra-intestinal tissues increase, how the immune systems limits spread of astrovirus to extra-intestinal tissue will be an important question for future studies.
Few studies have demonstrated a role for T cell-mediated control of astrovirus infection. Molberg et al. recovered astrovirus-specific CD4+ and CD8+ T cells from astrovirus-stimulated biopsies taken from the duodena of patients that had histologically normal intestines [54]. Wood et al. found chronic rotavirus and astrovirus infection in two children with T cell immunodeficiency [55]. Considering the fact that CD4+ T cells are essential in B cell maturation and antibody specificity; it is reasonable to suspect that T cells play a role in the immune response to astrovirus infection.
4. Innate Immune Responses
The innate immune system is the first line of defense against an invading pathogen. In vitro studies in cell culture models and in vivo animal models implicate that innate immune responses may be important in controlling astrovirus infection and replication. Although knowledge of the innate immune response to astrovirus infection is still in its infancy, we will discuss the known responses, or lack of, including histological changes, nitric oxide production, and type I IFN production in astrovirus replication and pathogenesis (summarized in Figure 1).
4.1. Histological Changes
Unlike other enteric pathogens, such as rotavirus and enteropathogenic Escherichia coli, which cause inflammation and cell death [56,57], astrovirus infection causes only mild histological changes. Although turkey poults infected with TAstV2 had watery diarrhea by 24 h post infection (hpi), this diarrhea was not associated with inflammation and pathology, as few lesions and little cell death were observed in the intestinal villi [4,6]. In gnotobiotic lambs and calves infected with ovine astrovirus or bovine astrovirus, only mild destruction of the villi was observed with few observations of dying enterocytes [19,58]. These animal results are similar to what is seen in humans, with little destruction of the intestinal villi after astrovirus infection [59].
4.2. Nitric Oxide Production
Nitric oxide is a key mediator of the innate immune system and is involved in pathogenesis and the control of infectious pathogens. Studies by Koci et al. demonstrate that TAstV-2 infection stimulates nitrite production, which limits viral replication [40]. Splenocytes from infected turkey poults made more nitrite after lipopolysaccharide (LPS) stimulation after splenocytes from uninfected poults [40]. TAstV infection increased nitrite levels after in vivo infection [40] in isolated avian macrophages and a chicken macrophage cell line (HD11) [40,60]. Treatment with nitric oxide synthase (NOS) inhibitors increased TAstV-2 replication in vitro and in vivo, demonstrating for the first time a role in the innate immune system in controlling astrovirus infection [40]. Importantly, turkey inducible nitric oxide synthase (iNOS) was shown to be turned on in intestinal epithelial cells where astrovirus replication was occurring, suggesting that these cells are capable of mounting their own defense against astrovirus infection [61].
4.3. Type I IFN
Two recent, independent studies explored the type I IFN response during human astrovirus infection, providing the most comprehensive knowledge about an innate immune response to astrovirus infection to date [62,63]. Although each study used different astrovirus serotypes, both Marvin et al. (using HAstV-1) and Guix et al. (using HAstV-4) found that infection induced a small, but significant increase in IFN-β mRNA and protein levels at 24 hpi [62,63]. This increase in IFN-β mRNA levels was not seen after infection with either UV inactivation of the virus or HAstV1 recombinant capsid, indicating that this type I IFN response is dependent on viral replication [62,63]. The transcription factor IFN regulatory factor (IRF)-3, which, when phosphorylated, translocates into the nucleus to promote transcription of the IFN-β promoter, was located in the cytoplasm in infected cells at 12 hpi but was localized in the nucleus at 24–48 hpi [63]. Many RNA viruses, such as coronaviruses and rotaviruses, encode IFN antagonists [64,65,66,67]. However, Guix et al. found that HAstV-4 infection did not block polyinosinic:polycytidylic acid (polyI:C)-induced type I IFN levels, suggesting that HAstV-4 does not encode a protein that exhibits interferon–antagonist activity [63].
To determine if type I IFN limited astrovirus infection and replication, both groups pretreated the human intestinal adenocarcinoma cell line Caco2 with recombinant type I IFN prior to astrovirus infection. Guix et al. pretreated with a single dose of IFN alpha (IFN-α) and saw a signification reduction in new capsid protein synthesis and mRNA levels at all HAstV-4 multiplicity of infection (MOIs) used [63]. They also saw a 2-fold increase in viral RNA and infectious virion levels when inhibiting TANK-binding kinase 1 (TBK1), which lies upstream of IRF-3 [63]. Marvin et al. pretreated Caco2 cells with increasing amounts of IFN-β and saw a dose-dependent reduction in new capsid protein synthesis, and, importantly, found that the amount of IFN-β produced during astrovirus infection is sufficient to decrease infectious progeny virion levels, as treatment with an IFN-β neutralizing antibody during infection increased HAstV-1 titers by 2.5 log-units [62]. Marvin et al. also saw decreased levels of positive-strand RNA levels at 8 and 16 hpi in Caco2 cells pretreated with IFN-β [62]. This decrease was not observed at early time points (1 and 4 hpi), suggesting that IFN-β pretreatment does not decrease HAstV-1 entry [62].
In addition to limiting viral replication, both studies asked if type I IFN influences astrovirus-induced barrier permeability. Astrovirus infection has been shown to increase epithelial permeability, which correlated with the disruption of tight junction proteins and sodium malabsorption [3,4,68]. Guix et al. concluded a positive correlation between IFN-β RNA production and barrier permeability during HAstV-4 infection [63]. Marvin et al. demonstrated that type I IFN may be important in protecting the epithelial barrier during HAstV-1 infection. Pretreatment of Caco2 cells with IFN-β reduced the HAstV-1-induced decrease in transepithelial electrical resistance (TER) and decreased the transmigration of an inert dye (flux) from the apical side of the cell monolayer to the basolateral side [62]. Additionally, HAstV-1 infection in the presence of an IFN-β neutralizing antibody saw increased levels of flux compared to isotype control [62]. Although the permeability studies were performed in the absence of trypsin, and therefore only one round of replication occurred, it is possible that the varying levels of unprocessed viral particles (lower levels with IFN-β pretreatment and higher levels in the presence of an IFN-β neutralizing antibody compared to control cells) could contribute to the differences in barrier permeability since astrovirus-induced barrier permeability is independent of viral replication [68].
The effects of type I IFN signaling in astrovirus replication, clearance, and barrier permeability have been examined in vivo using IFNaR-/- knockout mice. Oral gavage of wild-type C57BL/6 mice with MuAstV-positive fecal filtrates results in productive infection, viral shedding, and increased intestinal permeability [62]. However, the IFNaR−/− mice used in the study were found to already be MuAstV-positive in their colony. The viral titers shed in the stool of wild-type mice peaked at 6 to 11 dpi, shedding similar levels as the IFNaR−/− mice. However, wild-type mice titers rapidly decreased after 11 dpi and cleared the infection by 53 dpi, compared to IFNaR−/− mice that shed virus at high levels (>106 copies/μg RNA) at 53 dpi. These data suggest that type I IFN signaling is important for viral clearance in vivo [62]. IFNaR−/− mice also had increased intestinal permeability compared to MuAstV-infected wild-type mice, which correlated with their in vitro observations, suggesting a role for type I IFN in protecting the epithelial barrier during infection, although MuAstV-free IFNaR−/− mice would need to be generated to definitively determine the role of type I IFN signaling in intestinal permeability [62].
Overall, the studies of Guix et al. and Marvin et al. demonstrate that the type I IFN system can limit astrovirus replication in vitro and in vivo and protects against astrovirus-induced barrier permeability. How astrovirus replication induces type I IFN production, and why induction is only seen late in infection and at low levels remains unknown. Additionally, how type I IFN protects against astrovirus-induced barrier permeability remains to be resolved.
4.4. Additional Innate Immune Responses
Several observations in animals have described other innate immune responses during astrovirus infection. Astroviruses have been found in macrophages of lambs and described in calves with cells around Peyer’s patches [58,69]. However, whether these cells are activated in response to astrovirus infection have yet to be determined. TAstV-2 infection induces active TGF-β, an immunosuppressive cytokine that may explain why there are limited histological changes after infection [70], in the serum of infected turkey poults [6]. Whether TGF-β levels play a role in astrovirus replication and/or suppression of the immune response has yet to be determined.
Finally, a study in mice found a role for the signal transducer and activator of transcription 1 (STAT1) transcription factor, a factor known for controlling viral infection [71], in limiting MuAstV replication [42]. STAT1 knockout mice (STAT1−/−) shed 10-fold more viral copies in the feces 14 dpi compared to wild-type mice and had viral RNA in the spleen and the mesenteric lymph nodes, while viral RNA was not detected in those organs of wild-type mice [42]. Like type I IFN signaling, STAT1 has a role in limiting mouse astrovirus infection, but the mechanism(s) remains unknown. It is possible that type I IFN signaling leads to STAT1-mediated transcription of antiviral proteins that can inhibit astrovirus replication.
5. Astrovirus Evasion and Suppression of the Immune Response
Multiple studies have concluded that astrovirus may evade the immune system by preventing complement activation [72,73,74,75]. The complement system is a fundamental component of the immune response that detects pathogens through one of three pathways, the classical, the mannose-binding lectin (MBL) and the alternative pathways. Once activated, it can eliminate pathogens, regulate the inflammatory response, and help shape the adaptive immune response. The HAstV-1 capsid protein inhibits the classical and lectin pathways by binding to key initiator molecules C1q and MBL, preventing subsequent downstream activation [73,74,75]. Considering a critical role of complement is initiating an inflammatory response and the observations that astroviruses do not cause inflammation highly suggests that astrovirus inhibition of complement activation is a key player in evading the immune system and suppressing the inflammatory response. Indeed, Tam et al. found only low levels of complement-mediated NF-κB activation upon infection with HAstV compared to adenovirus and human papillomavirus virus-like particles, suggesting that HAstV has strategies to evade detection by complement factor C3 [72]. Tam et al. also determined that intracellular sensing of C3 activated the interferon regulatory factor 3 transcription pathways, resulting in a robust secretion of IFN-β [72]. HAstV ability to evade detection by C3, which is involved in triggering an interferon response, may be why Guix et al. and Marvin et al. detected low levels of type I IFN at later times post infection. Future studies involving HAstV mutants that do not bind complement molecules will delineate the role(s) of suppression of complement activation during astrovirus infection.
Studies in turkey poults suggest that astrovirus infection suppresses the immune system, making the host more susceptible to subsequent infections [60,76]. Turkey macrophage infection in vitro reduced macrophage viability and intracellular killing of a subsequent Escherichia coli (E. coli) challenge compared to uninfected macrophages [60]. In vivo experiments showed that macrophages isolated from TAstV-infected poults had a percentage of phagocytic macrophages compared to macrophages isolated from uninfected poults [60]. These macrophages had less interleukin (IL)-1 and IL-6 activity and had fewer E. coli per macrophage compared to macrophages isolated from uninfected poults [60]. TAstV-infected poults also recruited almost half the number of Sephadex-elicited inflammatory cells to the abdominal cavity compared to uninfected poults [60]. These results correlate with an increased number of E. coli in the spleens of TAstV-infected poults [60], suggesting that TAstV infection decreases macrophage viability and function, rendering the host more susceptible to secondary bacterial, and possibly viral, infections.
6. Conclusions
In summary, although our understanding of the immune responses to astrovirus infection has progressed over the years, further studies are required to fully characterize the cellular and molecular factors that mediate the immune response to astrovirus infection, including identifying the correlates of protection that inhibit viral replication and pathogenesis. Future areas of inquiry include the absence of a potent inflammatory response following the rupture of endosomal membranes [12], as seen with other viral-induced membrane ruptures [77,78] and determination of how astrovirus particles exit cells through a non-lytic mechanism [21]. Additionally, an improved understanding of the mechanisms underlying viral inhibition of the inflammatory response and how type I IFN limits astrovirus-induced barrier permeability are critical for the development of therapeutics and treatment protocols. The development of a mouse model will be invaluable in understanding the viral replication kinetics and innate and adaptive immune responses to astrovirus infection in mammals. A full understanding of the immune response to astrovirus infection is necessary if we are to control and prevent astrovirus diseases.
Acknowledgments
I thank Anita Zahs and Bryan Kaplan for providing helpful comments and discussion on the manuscript and acknowledge funding from Drake University.
Conflicts of Interest
The author declares no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Zimmerman, C.M.; Bresee, J.S.; Parashar, U.D.; Riggs, T.L.; Holman, R.C.; Glass, R.I. Cost of diarrhea-associated hospitalizations and outpatient visits in an insured population of young children in the United States. Pediatr. Infect. Dis. J. 2001, 20, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.E.; Mitchell, D.K. Role of astroviruses in childhood diarrhea. Curr. Opin. Pediatr. 2000, 12, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Nighot, P.K.; Moeser, A.; Ali, R.A.; Blikslager, A.T.; Koci, M.D. Astrovirus infection induces sodium malabsorption and redistributes sodium hydrogen exchanger expression. Virology 2010, 401, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Meliopoulos, V.A.; Marvin, S.A.; Freiden, P.; Moser, L.A.; Nighot, P.; Ali, R.; Blikslager, A.; Reddivari, M.; Heath, R.J.; Koci, M.D.; et al. Oral administration of astrovirus capsid protein is sufficient to induce acute diarrhea in vivo. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, J.B.; Lee, T.W.; Craig, J.W.; Reed, S.E. Astrovirus infection in volunteers. J. Med. Virol. 1979, 3, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Koci, M.D.; Moser, L.A.; Kelley, L.A.; Larsen, D.; Brown, C.C.; Schultz-Cherry, S. Astrovirus induces diarrhea in the absence of inflammation and cell death. J. Virol. 2003, 77, 11798–11808. [Google Scholar] [CrossRef] [PubMed]
- Glass, R.I.; Noel, J.; Mitchell, D.; Herrmann, J.E.; Blacklow, N.R.; Pickering, L.K.; Dennehy, P.; Ruiz-Palacios, G.; de Guerrero, M.L.; Monroe, S.S. The changing epidemiology of astrovirus-associated gastroenteritis: A review. Arch. Virol. Suppl. 1996, 12, 287–300. [Google Scholar] [PubMed]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals—Molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Pinto, R.M.; Guix, S. Human astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef] [PubMed]
- Midthun, K.; Greenberg, H.B.; Kurtz, J.B.; Gary, G.W.; Lin, F.Y.; Kapikian, A.Z. Characterization and seroepidemiology of a type 5 astrovirus associated with an outbreak of gastroenteritis in Marin County, California. J. Clin. Microbiol. 1993, 31, 955–962. [Google Scholar] [PubMed]
- Kurtz, J.B.; Lee, T.W.; Parsons, A.J. The action of alcohols on rotavirus, astrovirus and enterovirus. J. Hosp. Infect. 1980, 1, 321–325. [Google Scholar] [CrossRef]
- Mendez, E.; Munoz-Yanez, C.; Sanchez-San Martin, C.; Aguirre-Crespo, G.; Banos-Lara Mdel, R.; Gutierrez, M.; Espinosa, R.; Acevedo, Y.; Arias, C.F.; Lopez, S. Characterization of human astrovirus cell entry. J. Virol. 2014, 88, 2452–2460. [Google Scholar] [CrossRef] [PubMed]
- Willcocks, M.M.; Carter, M.J. Identification and sequence determination of the capsid protein gene of human astrovirus serotype 1. FEMS Microbiol. Lett. 1993, 114, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Monroe, S.S.; Jiang, B.; Stine, S.E.; Koopmans, M.; Glass, R.I. Subgenomic RNA sequence of human astrovirus supports classification of astroviridae as a new family of RNA viruses. J. Virol. 1993, 67, 3611–3614. [Google Scholar] [PubMed]
- Jang, S.Y.; Jeong, W.H.; Kim, M.S.; Lee, Y.M.; Lee, J.I.; Lee, G.C.; Paik, S.Y.; Koh, G.P.; Kim, J.M.; Lee, C.H. Detection of replicating negative-sense RNAs in Caco-2 cells infected with human astrovirus. Arch. Virol. 2010, 155, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- Monceyron, C.; Grinde, B.; Jonassen, T.O. Molecular characterisation of the 3′-end of the astrovirus genome. Arch. Virol. 1997, 142, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Mendez, E.; Aguirre-Crespo, G.; Zavala, G.; Arias, C.F. Association of the astrovirus structural protein VP90 with membranes plays a role in virus morphogenesis. J. Virol. 2007, 81, 10649–10658. [Google Scholar] [CrossRef] [PubMed]
- Guix, S.; Caballero, S.; Bosch, A.; Pinto, R.M. C-terminal NSP1a protein of human astrovirus colocalizes with the endoplasmic reticulum and viral RNA. J. Virol. 2004, 78, 13627–13636. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.W.; Angus, K.W.; Snodgrass, D.R. Ultrastructure of the small intestine in astrovirus-infected lambs. J. Gen. Virol. 1980, 49, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Denison, M.R. Seeking membranes: Positive-strand RNA virus replication complexes. PLoS Biol. 2008, 6, e270. [Google Scholar] [CrossRef] [PubMed]
- Banos-Lara Mdel, R.; Mendez, E. Role of individual caspases induced by astrovirus on the processing of its structural protein and its release from the cell through a non-lytic mechanism. Virology 2010, 401, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Mendez, E.; Salas-Ocampo, E.; Arias, C.F. Caspases mediate processing of the capsid precursor and cell release of human astroviruses. J. Virol. 2004, 78, 8601–8608. [Google Scholar] [CrossRef] [PubMed]
- Behling-Kelly, E.; Schultz-Cherry, S.; Koci, M.; Kelley, L.; Larsen, D.; Brown, C. Localization of astrovirus in experimentally infected turkeys as determined by in situ hybridization. Vet. Pathol. 2002, 39, 595–598. [Google Scholar] [CrossRef] [PubMed]
- Gough, R.E.; Borland, E.D.; Keymer, I.F.; Stuart, J.C. An outbreak of duck hepatitis type II in commercial ducks. Avian Pathol. 1985, 14, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Gavier-Widen, D.; Brojer, C.; Dietz, H.H.; Englund, L.; Hammer, A.S.; Hedlund, K.O.; Hard af Segerstad, C.; Nilsson, K.; Nowotny, N.; Puurula, V.; et al. Investigations into shaking mink syndrome: An encephalomyelitis of unknown cause in farmed mink (mustela vison) kits in Scandinavia. J. Vet. Diagn. Investig. 2004, 16, 305–312. [Google Scholar]
- Blomstrom, A.L.; Widen, F.; Hammer, A.S.; Belak, S.; Berg, M. Detection of a novel astrovirus in brain tissue of mink suffering from shaking mink syndrome by use of viral metagenomics. J. Clin. Microbiol. 2010, 48, 4392–4396. [Google Scholar] [CrossRef] [PubMed]
- Seuberlich, T.; Wuthrich, D.; Selimovic-Hamza, S.; Drogemuller, C.; Oevermann, A.; Bruggmann, R.; Bouzalas, I. Identification of a second encephalitis-associated astrovirus in cattle. Emerg. Microbes Infect. 2016, 5, e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Diab, S.; McGraw, S.; Barr, B.; Traslavina, R.; Higgins, R.; Talbot, T.; Blanchard, P.; Rimoldi, G.; Fahsbender, E.; et al. Divergent astrovirus associated with neurologic disease in cattle. Emerg. Infect. Dis. 2013, 19, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Bouzalas, I.G.; Wuthrich, D.; Walland, J.; Drogemuller, C.; Zurbriggen, A.; Vandevelde, M.; Oevermann, A.; Bruggmann, R.; Seuberlich, T. Neurotropic astrovirus in cattle with nonsuppurative encephalitis in Europe. J. Clin. Microbiol. 2014, 52, 3318–3324. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Pan, M.; Wang, X.; Xu, Y.; Xie, X.; Knowles, N.J.; Yang, H.; Zhang, D. Complete sequence of a duck astrovirus associated with fatal hepatitis in ducklings. J. Gen. Virol. 2009, 90, 1104–1108. [Google Scholar] [CrossRef] [PubMed]
- Brnic, D.; Prpic, J.; Keros, T.; Roic, B.; Staresina, V.; Jemersic, L. Porcine astrovirus viremia and high genetic variability in pigs on large holdings in Croatia. Infect. Genet. Evol. 2013, 14, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Wunderli, W.; Meerbach, A.; Gungor, T.; Berger, C.; Greiner, O.; Caduff, R.; Trkola, A.; Bossart, W.; Gerlach, D.; Schibler, M.; et al. Astrovirus infection in hospitalized infants with severe combined immunodeficiency after allogeneic hematopoietic stem cell transplantation. PLoS ONE 2011, 6, e27483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtz, L.R.; Wylie, K.M.; Sodergren, E.; Jiang, Y.; Franz, C.J.; Weinstock, G.M.; Storch, G.A.; Wang, D. Astrovirus MLB2 viremia in febrile child. Emerg. Infect. Dis. 2011, 17, 2050–2052. [Google Scholar] [CrossRef] [PubMed]
- Cordey, S.; Vu, D.L.; Schibler, M.; L’Huillier, A.G.; Brito, F.; Docquier, M.; Posfay-Barbe, K.M.; Petty, T.J.; Turin, L.; Zdobnov, E.M.; et al. Astrovirus MLB2, a new gastroenteric virus associated with meningitis and disseminated infection. Emerg. Infect. Dis. 2016, 22, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Quan, P.L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Firth, C.; Palacios, G.; Baisre-De-Leon, A.; Paddock, C.D.; et al. Astrovirus encephalitis in boy with X-linked agammaglobulinemia. Emerg. Infect. Dis. 2010, 16, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Naccache, S.N.; Peggs, K.S.; Mattes, F.M.; Phadke, R.; Garson, J.A.; Grant, P.; Samayoa, E.; Federman, S.; Miller, S.; Lunn, M.P.; et al. Diagnosis of neuroinvasive astrovirus infection in an immunocompromised adult with encephalitis by unbiased next-generation sequencing. Clin. Infect. Dis. 2015, 60, 919–923. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.R.; Morfopoulou, S.; Hubb, J.; Emmett, W.A.; Ip, W.; Shah, D.; Brooks, T.; Paine, S.M.; Anderson, G.; Virasami, A.; et al. Astrovirus VA1/HMO-C: An increasingly recognized neurotropic pathogen in immunocompromised patients. Clin. Infect. Dis. 2015, 60, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Cordey, S.; Brito, F.; Vu, D.L.; Turin, L.; Kilowoko, M.; Kyungu, E.; Genton, B.; Zdobnov, E.M.; D’Acremont, V.; Kaiser, L. Astrovirus VA1 identified by next-generation sequencing in a nasopharyngeal specimen of a febrile Tanzanian child with acute respiratory disease of unknown etiology. Emerg. Microbes Infect. 2016, 5, e67. [Google Scholar] [CrossRef] [PubMed]
- Koci, M.D. Immunity and resistance to astrovirus infection. Viral Immunol. 2005, 18, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Koci, M.D.; Kelley, L.A.; Larsen, D.; Schultz-Cherry, S. Astrovirus-induced synthesis of nitric oxide contributes to virus control during infection. J. Virol. 2004, 78, 1564–1574. [Google Scholar] [CrossRef] [PubMed]
- Mombaerts, P.; Iacomini, J.; Johnson, R.S.; Herrup, K.; Tonegawa, S.; Papaioannou, V.E. Rag-1-deficient mice have no mature B and T lymphocytes. Cell 1992, 68, 869–877. [Google Scholar] [CrossRef]
- Yokoyama, C.C.; Loh, J.; Zhao, G.; Stappenbeck, T.S.; Wang, D.; Huang, H.V.; Virgin, H.W.; Thackray, L.B. Adaptive immunity restricts replication of novel murine astroviruses. J. Virol. 2012, 86, 12262–12270. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, J.; Lee, T. Astrovirus gastroenteritis age distribution of antibody. Med. Microbiol. Immunol. 1978, 166, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.K. Astrovirus gastroenteritis. Pediatr. Infect. Dis. J. 2002, 21, 1067–1069. [Google Scholar] [CrossRef] [PubMed]
- Meliopoulos, V.A.; Kayali, G.; Burnham, A.; Oshansky, C.M.; Thomas, P.G.; Gray, G.C.; Beck, M.A.; Schultz-Cherry, S. Detection of antibodies against turkey astrovirus in humans. PLoS ONE 2014, 9, e96934. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.A.; Small, C.T.; Freiden, P.; Feeroz, M.M.; Matsen, F.A.T.; San, S.; Hasan, M.K.; Wang, D.; Jones-Engel, L.; Schultz-Cherry, S. Non-human primates harbor diverse mammalian and avian astroviruses including those associated with human infections. PLoS Pathog. 2015, 11, e1005225. [Google Scholar] [CrossRef] [PubMed]
- Bogdanoff, W.A.; Campos, J.; Perez, E.I.; Yin, L.; Alexander, D.L.; DuBois, R.M. Structure of a human astrovirus capsid—antibody complex and mechanistic insights into virus neutralization. J. Virol. 2016, in press. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Fauquier, A.; Carrascosa, A.L.; Carrascosa, J.L.; Otero, A.; Glass, R.I.; Lopez, J.A.; San Martin, C.; Melero, J.A. Characterization of a human astrovirus serotype 2 structural protein (VP26) that contains an epitope involved in virus neutralization. Virology 1994, 201, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Bass, D.M.; Upadhyayula, U. Characterization of human serotype 1 astrovirus-neutralizing epitopes. J. Virol. 1997, 71, 8666–8671. [Google Scholar] [PubMed]
- York, R.L.; Yousefi, P.A.; Bogdanoff, W.; Haile, S.; Tripathi, S.; DuBois, R.M. Structural, mechanistic, and antigenic characterization of the human astrovirus capsid. J. Virol. 2015, 90, 2254–2263. [Google Scholar] [CrossRef] [PubMed]
- Toh, Y.; Harper, J.; Dryden, K.A.; Yeager, M.; Arias, C.F.; Mendez, E.; Tao, Y.J. Crystal structure of the human astrovirus capsid protein. J. Virol. 2016, 90, 9008–9017. [Google Scholar] [CrossRef] [PubMed]
- DuBois, R.M.; Freiden, P.; Marvin, S.; Reddivari, M.; Heath, R.J.; White, S.W.; Schultz-Cherry, S. Crystal structure of the avian astrovirus capsid spike. J. Virol. 2013, 87, 7853–7863. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Dong, L.; Mendez, E.; Tao, Y. Crystal structure of the human astrovirus capsid spike. Proc. Natl. Acad. Sci. USA 2011, 108, 12681–12686. [Google Scholar] [CrossRef] [PubMed]
- Molberg, O.; Nilsen, E.M.; Sollid, L.M.; Scott, H.; Brandtzaeg, P.; Thorsby, E.; Lundin, K.E. CD4+ T cells with specific reactivity against astrovirus isolated from normal human small intestine. Gastroenterology 1998, 114, 115–122. [Google Scholar] [CrossRef]
- Wood, D.J.; David, T.J.; Chrystie, I.L.; Totterdell, B. Chronic enteric virus infection in two T-cell immunodeficient children. J. Med. Virol. 1988, 24, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, C.A.; Acosta, O. Inflammatory and oxidative stress in rotavirus infection. World J. Virol. 2016, 5, 38–62. [Google Scholar] [CrossRef] [PubMed]
- Crane, J.K.; Majumdar, S.; Pickhardt, D.F., 3rd. Host cell death due to enteropathogenic Escherichia coli has features of apoptosis. Infect. Immun. 1999, 67, 2575–2584. [Google Scholar] [PubMed]
- Woode, G.N.; Pohlenz, J.F.; Gourley, N.E.; Fagerland, J.A. Astrovirus and breda virus infections of dome cell epithelium of bovine ileum. J. Clin. Microbiol. 1984, 19, 623–630. [Google Scholar] [PubMed]
- Sebire, N.J.; Malone, M.; Shah, N.; Anderson, G.; Gaspar, H.B.; Cubitt, W.D. Pathology of astrovirus associated diarrhoea in a paediatric bone marrow transplant recipient. J. Clin. Pathol. 2004, 57, 1001–1003. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, M.A.; Saif, Y.M.; Heggen-Peay, C.L.; Edens, F.W.; Havenstein, G.B. Induction of functional defects in macrophages by a poult enteritis and mortality syndrome-associated turkey astrovirus. Avian Dis. 2001, 45, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Meyerhoff, R.R.; Nighot, P.K.; Ali, R.A.; Blikslager, A.T.; Koci, M.D. Characterization of turkey inducible nitric oxide synthase and identification of its expression in the intestinal epithelium following astrovirus infection. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Marvin, S.A.; Huerta, C.T.; Sharp, B.; Freiden, P.; Cline, T.D.; Schultz-Cherry, S. Type I interferon response limits astrovirus replication and protects against increased barrier permeability in vitro and in vivo. J. Virol. 2015, 90, 1988–1996. [Google Scholar] [CrossRef] [PubMed]
- Guix, S.; Perez-Bosque, A.; Miro, L.; Moreto, M.; Bosch, A.; Pinto, R.M. Type I interferon response is delayed in human astrovirus infections. PLoS ONE 2015, 10, e0123087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopecky-Bromberg, S.A.; Martinez-Sobrido, L.; Frieman, M.; Baric, R.A.; Palese, P. Severe acute respiratory syndrome coronavirus open reading frame (ORF) 3b, ORF 6, and nucleocapsid proteins function as interferon antagonists. J. Virol. 2007, 81, 548–557. [Google Scholar] [CrossRef] [PubMed]
- Graff, J.W.; Ettayebi, K.; Hardy, M.E. Rotavirus NSP1 inhibits NFκB activation by inducing proteasome-dependent degradation of β-TrCP: A novel mechanism of IFN antagonism. PLoS Pathog. 2009, 5, e1000280. [Google Scholar] [CrossRef] [PubMed]
- Clementz, M.A.; Chen, Z.; Banach, B.S.; Wang, Y.; Sun, L.; Ratia, K.; Baez-Santos, Y.M.; Wang, J.; Takayama, J.; Ghosh, A.K.; et al. Deubiquitinating and interferon antagonism activities of coronavirus papain-like proteases. J. Virol. 2010, 84, 4619–4629. [Google Scholar] [CrossRef] [PubMed]
- Barro, M.; Patton, J.T. Rotavirus NSP1 inhibits expression of type I interferon by antagonizing the function of interferon regulatory factors IRF3, IRF5, and IRF7. J. Virol. 2007, 81, 4473–4481. [Google Scholar] [CrossRef] [PubMed]
- Moser, L.A.; Carter, M.; Schultz-Cherry, S. Astrovirus increases epithelial barrier permeability independently of viral replication. J. Virol. 2007, 81, 11937–11945. [Google Scholar] [CrossRef] [PubMed]
- Snodgrass, D.R.; Angus, K.W.; Gray, E.W.; Menzies, J.D.; Paul, G. Pathogenesis of diarrhoea caused by astrovirus infections in lambs. Arch. Virol. 1979, 60, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Letterio, J.J.; Roberts, A.B. Regulation of immune responses by TGF-β. Annu. Rev. Immunol. 1998, 16, 137–161. [Google Scholar] [CrossRef] [PubMed]
- Durbin, J.E.; Hackenmiller, R.; Simon, M.C.; Levy, D.E. Targeted disruption of the mouse Stat1 gene results in compromised innate immunity to viral disease. Cell 1996, 84, 443–450. [Google Scholar] [CrossRef]
- Tam, J.C.; Bidgood, S.R.; McEwan, W.A.; James, L.C. Intracellular sensing of complement C3 activates cell autonomous immunity. Science 2014, 345, 1256070. [Google Scholar] [CrossRef] [PubMed]
- Hair, P.S.; Gronemus, J.Q.; Crawford, K.B.; Salvi, V.P.; Cunnion, K.M.; Thielens, N.M.; Arlaud, G.J.; Rawal, N.; Krishna, N.K. Human astrovirus coat protein binds C1q and MBL and inhibits the classical and lectin pathways of complement activation. Mol. Immunol. 2010, 47, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Gronemus, J.Q.; Hair, P.S.; Crawford, K.B.; Nyalwidhe, J.O.; Cunnion, K.M.; Krishna, N.K. Potent inhibition of the classical pathway of complement by a novel C1q-binding peptide derived from the human astrovirus coat protein. Mol. Immunol. 2010, 48, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Bonaparte, R.S.; Hair, P.S.; Banthia, D.; Marshall, D.M.; Cunnion, K.M.; Krishna, N.K. Human astrovirus coat protein inhibits serum complement activation via C1, the first component of the classical pathway. J. Virol. 2008, 82, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, M.A.; Yu, M.; Saif, Y.M. A novel “small round virus” inducing poult enteritis and mortality syndrome and associated immune alterations. Avian Dis. 2000, 44, 275–283. [Google Scholar] [CrossRef] [PubMed]
- McGuire, K.A.; Barlan, A.U.; Griffin, T.M.; Wiethoff, C.M. Adenovirus type 5 rupture of lysosomes leads to cathepsin B-dependent mitochondrial stress and production of reactive oxygen species. J. Virol. 2011, 85, 10806–10813. [Google Scholar] [CrossRef] [PubMed]
- Barlan, A.U.; Griffin, T.M.; McGuire, K.A.; Wiethoff, C.M. Adenovirus membrane penetration activates the NLRP3 inflammasome. J. Virol. 2011, 85, 146–155. [Google Scholar] [CrossRef] [PubMed]
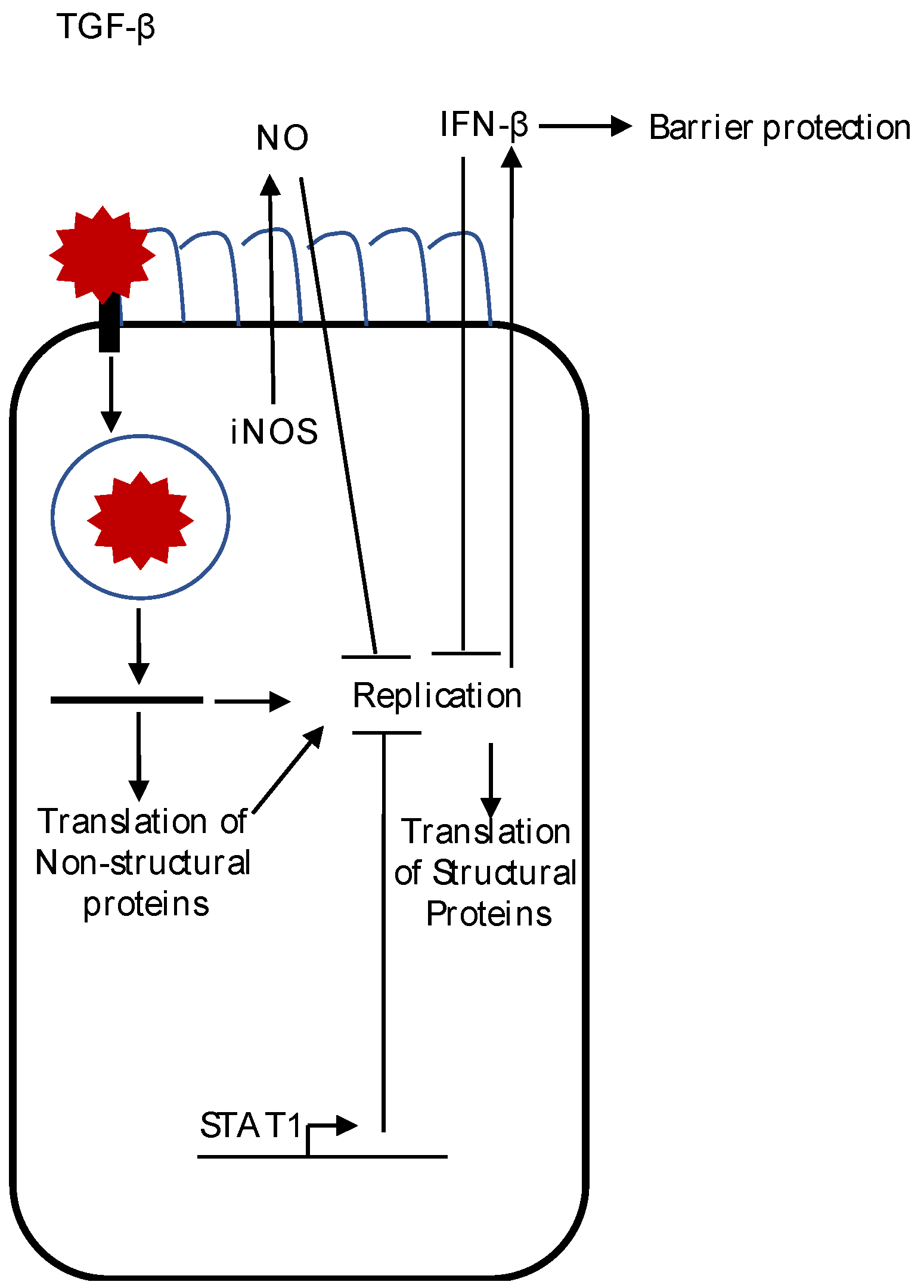
Figure 1.
A summary of the innate immune response to astrovirus infection. After binding and entry into the cell, viral uncoating of the astrovirus genome occurs. Replication induces IFN-β production, which suppresses viral replication and translation of the structural protein, and decreases astrovirus-induced barrier permeability. Synthesis of inducible nitric oxide synthase (iNOS) protein also occurs during astrovirus infection of epithelial cells. Although replication is not needed to induce iNOS in macrophages, whether or not iNOS induction occurs during astrovirus infection of epithelial cells is unknown. Active transforming growth factor beta (TGF-β) levels are increased after astrovirus infection, but the role of TGF-β after astrovirus infection is not known. The presence of signal transducer and activator of transcription 1 (STAT1) decreases astrovirus replication, but the mechanism(s) remain to be determined.
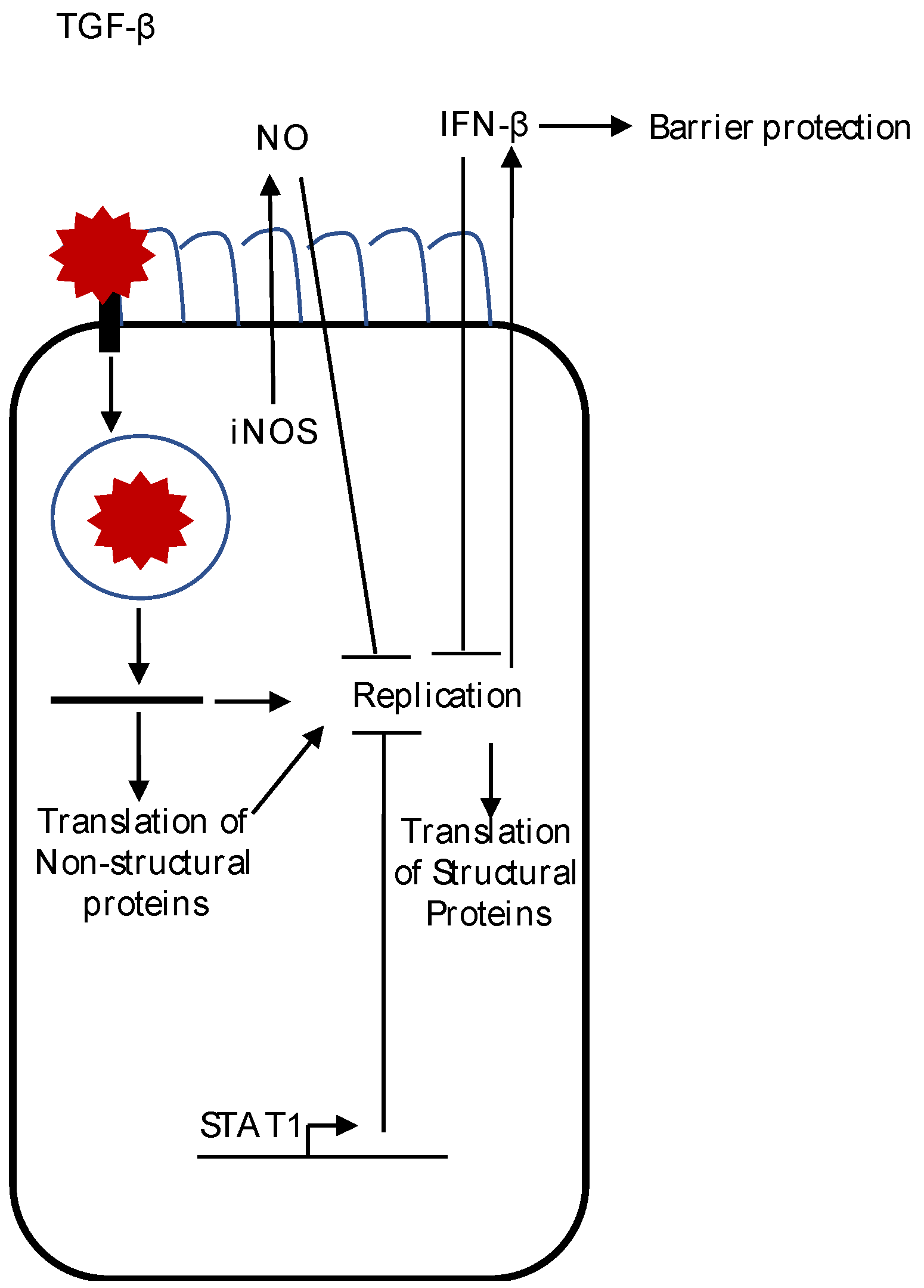
Figure 1.
A summary of the innate immune response to astrovirus infection. After binding and entry into the cell, viral uncoating of the astrovirus genome occurs. Replication induces IFN-β production, which suppresses viral replication and translation of the structural protein, and decreases astrovirus-induced barrier permeability. Synthesis of inducible nitric oxide synthase (iNOS) protein also occurs during astrovirus infection of epithelial cells. Although replication is not needed to induce iNOS in macrophages, whether or not iNOS induction occurs during astrovirus infection of epithelial cells is unknown. Active transforming growth factor beta (TGF-β) levels are increased after astrovirus infection, but the role of TGF-β after astrovirus infection is not known. The presence of signal transducer and activator of transcription 1 (STAT1) decreases astrovirus replication, but the mechanism(s) remain to be determined.

{kind=link}
| Animal/Human | Tissue(s) | Method(s) of Detection | Reference |
|---|---|---|---|
| Turkey | Bursa, Thymus, Spleen, Kidney, Liver, Skeletal Muscle, Bone Marrow, Pancreas, Plasma | RT-PCR, immunofluorescence, infectious virus isolation | [6,23] |
| Duck | Liver | RT-PCR | [30] |
| Cow | Brain | RT-PCR, Sequencing | [27,28,29] |
| Mink | Brain | Sequencing | [26] |
| Pig | Blood | RT-PCR, Sequencing | [31] |
| Human | Blood | RT-PCR, Sequencing | [32,33,34] |
| Human | Cerebrospinal Fluid | Sequencing | [34] |
| Human | Urine | Sequencing | [34] |
| Human | Brain | Sequencing, Immunohistochemistry | [35,36,37] |
| Human | Nasopharyngeal swab | RT-PCR, Sequencing | [32] |
| Human | Pharyngeal swab | RT-PCR, Sequencing | [32,38] |
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Marvin, S.A. The Immune Response to Astrovirus Infection. Viruses 2017, 9, 1. https://doi.org/10.3390/v9010001
AMA Style
Marvin SA. The Immune Response to Astrovirus Infection. Viruses. 2017; 9(1):1. https://doi.org/10.3390/v9010001
Chicago/Turabian StyleMarvin, Shauna A. 2017. "The Immune Response to Astrovirus Infection" Viruses 9, no. 1: 1. https://doi.org/10.3390/v9010001
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.