
A New Orbivirus Isolated from Mosquitoes in North-Western Australia Shows Antigenic and Genetic Similarity to Corriparta Virus but Does Not Replicate in Vertebrate Cells

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Mosquito Collection and Initial Pool Screening
2.3. Parry’s Lagoon Virus Detection and Isolation from Mosquito Homogenates
2.4. Virus Culture
2.5. Electron Microscopy
2.6. Genome Sequencing and Phylogenetic Analysis
2.7. Serological Cross-Reactivity Studies
2.8. Microneutralisation Assay
2.9. Vertebrate Cell Infection Assays
3. Results
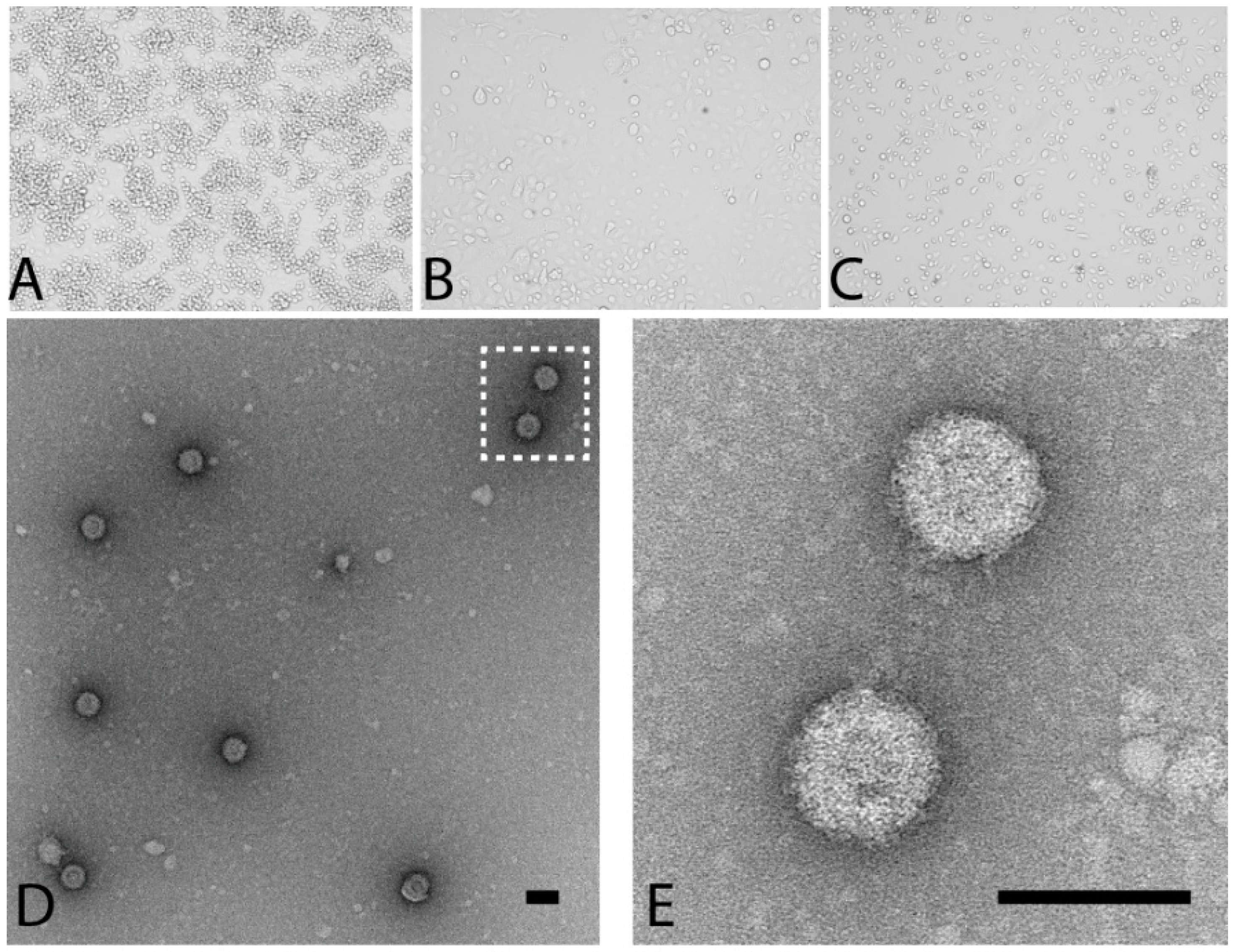
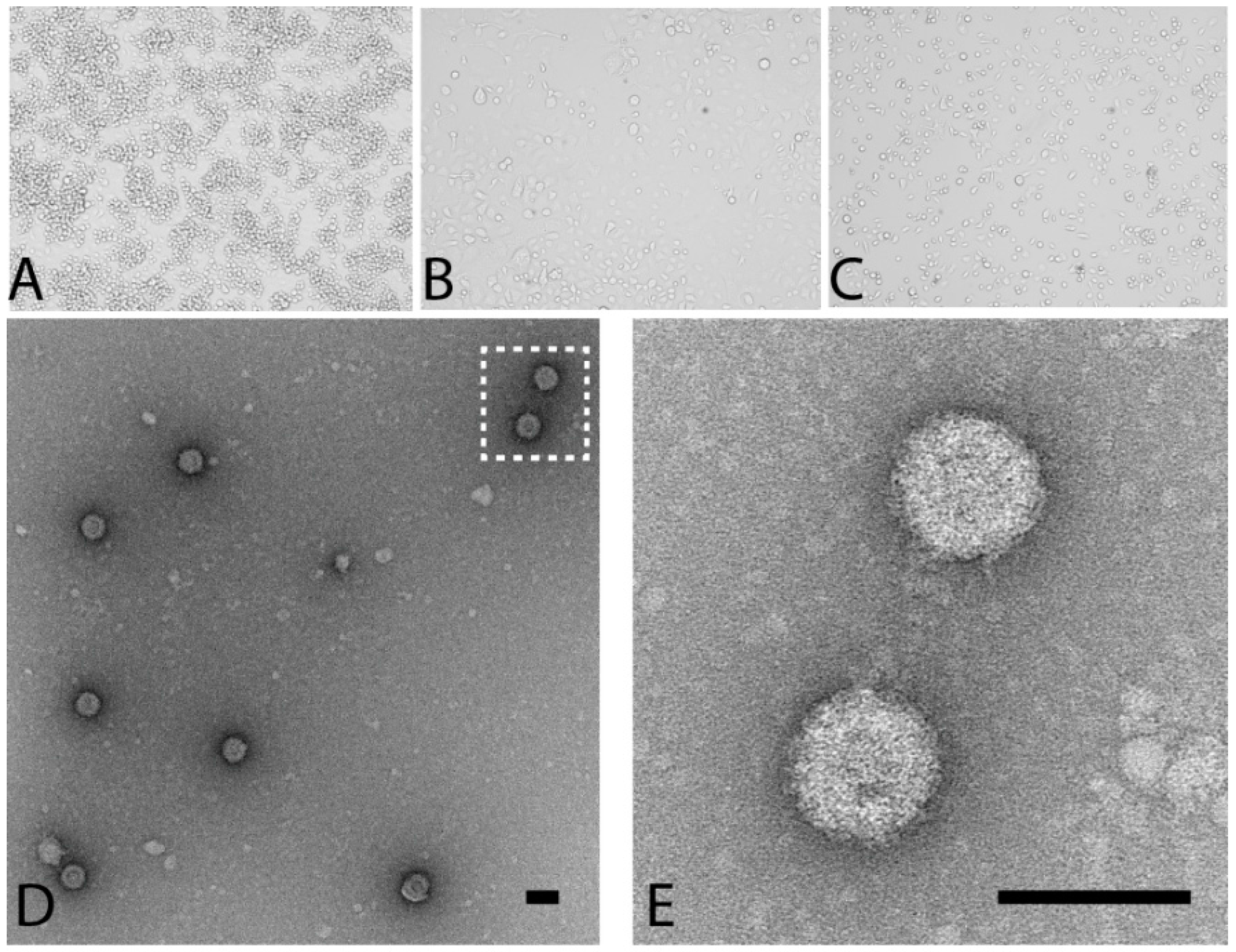
3.1. Detection, Isolation and Culture of the Prototype PLV Isolate
3.2. PLV Virions Display Typical Reovirus Morphology
3.3. Detection of Other PLV Isolates from Mosquitoes Collected in Western Australia
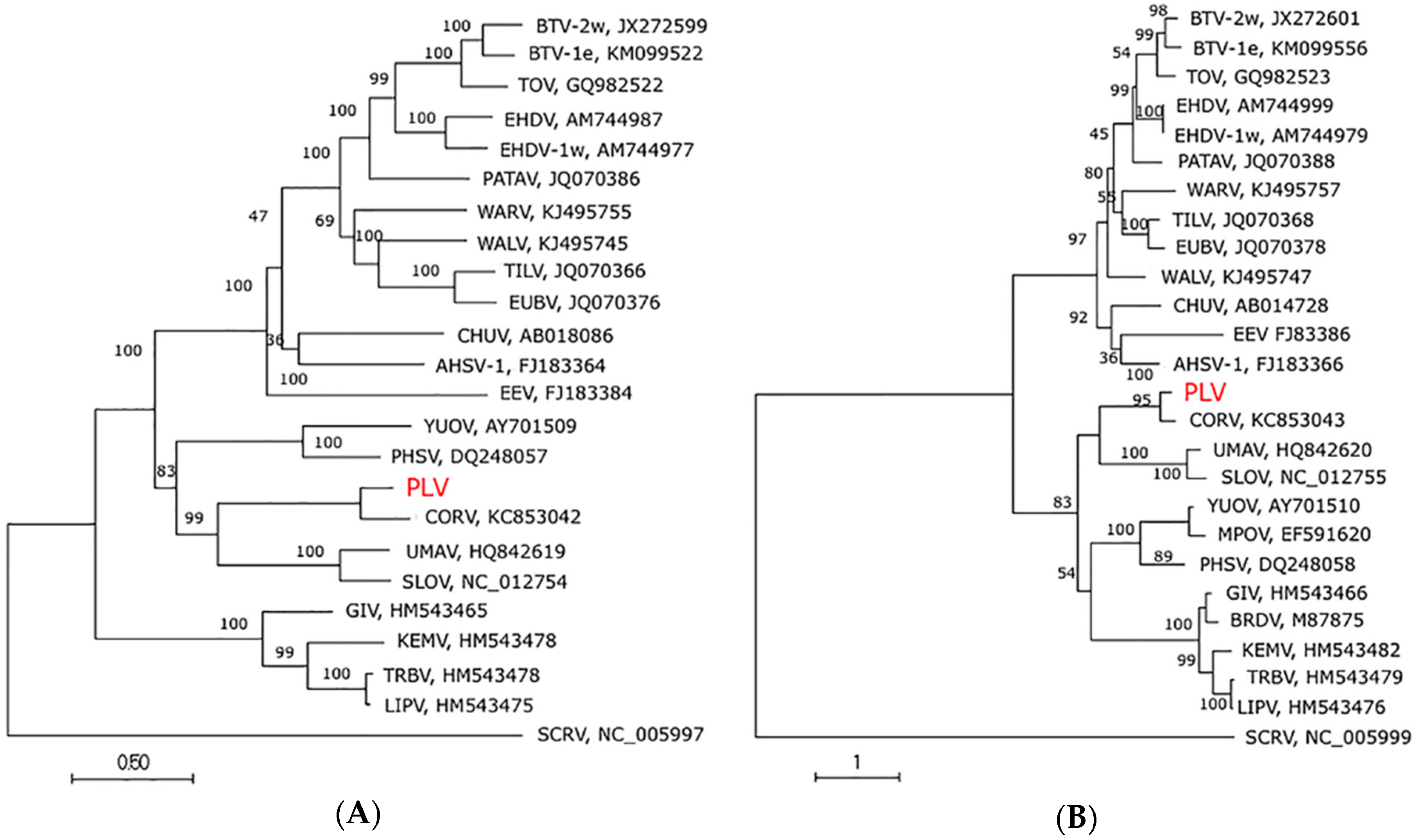
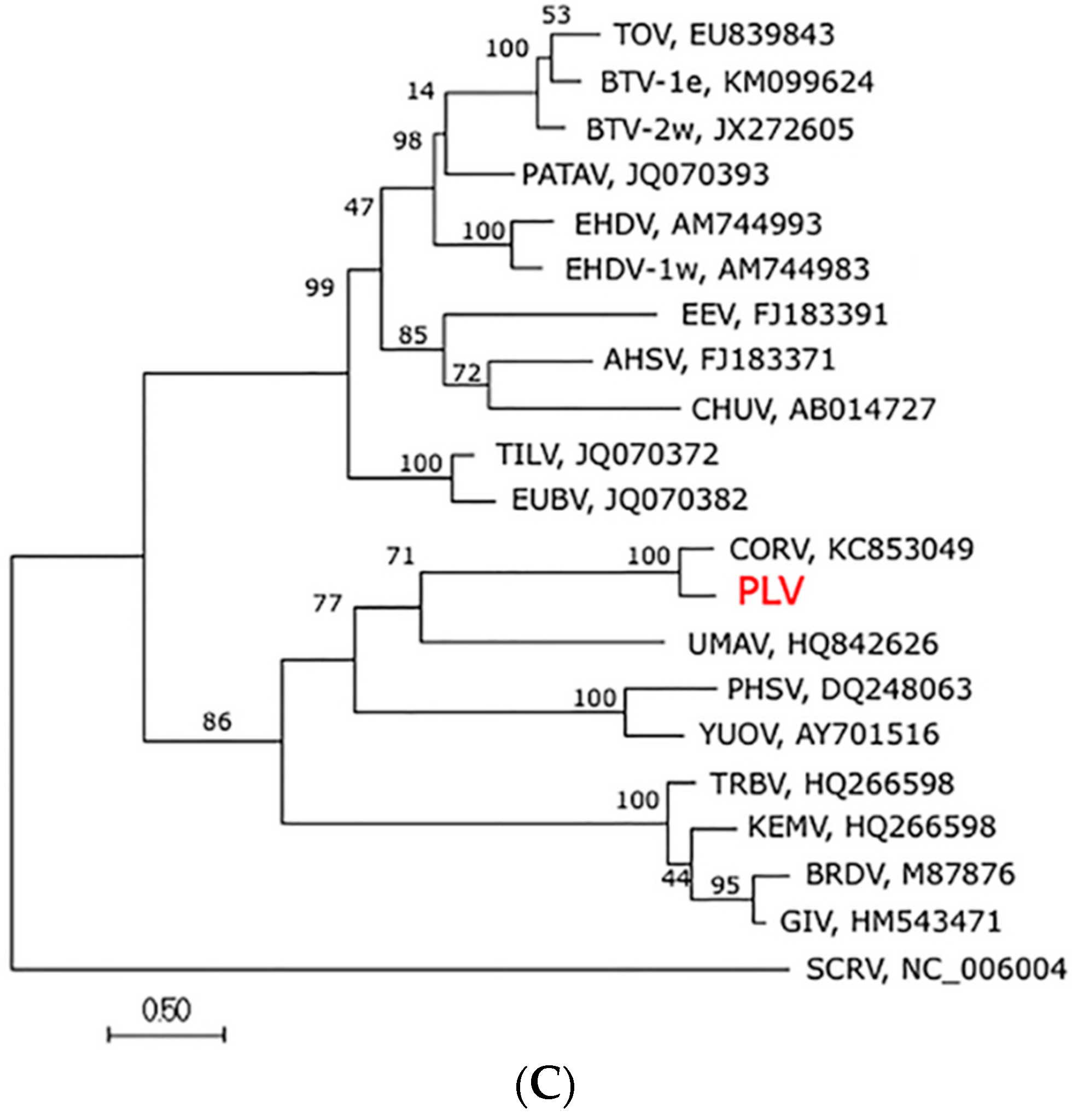
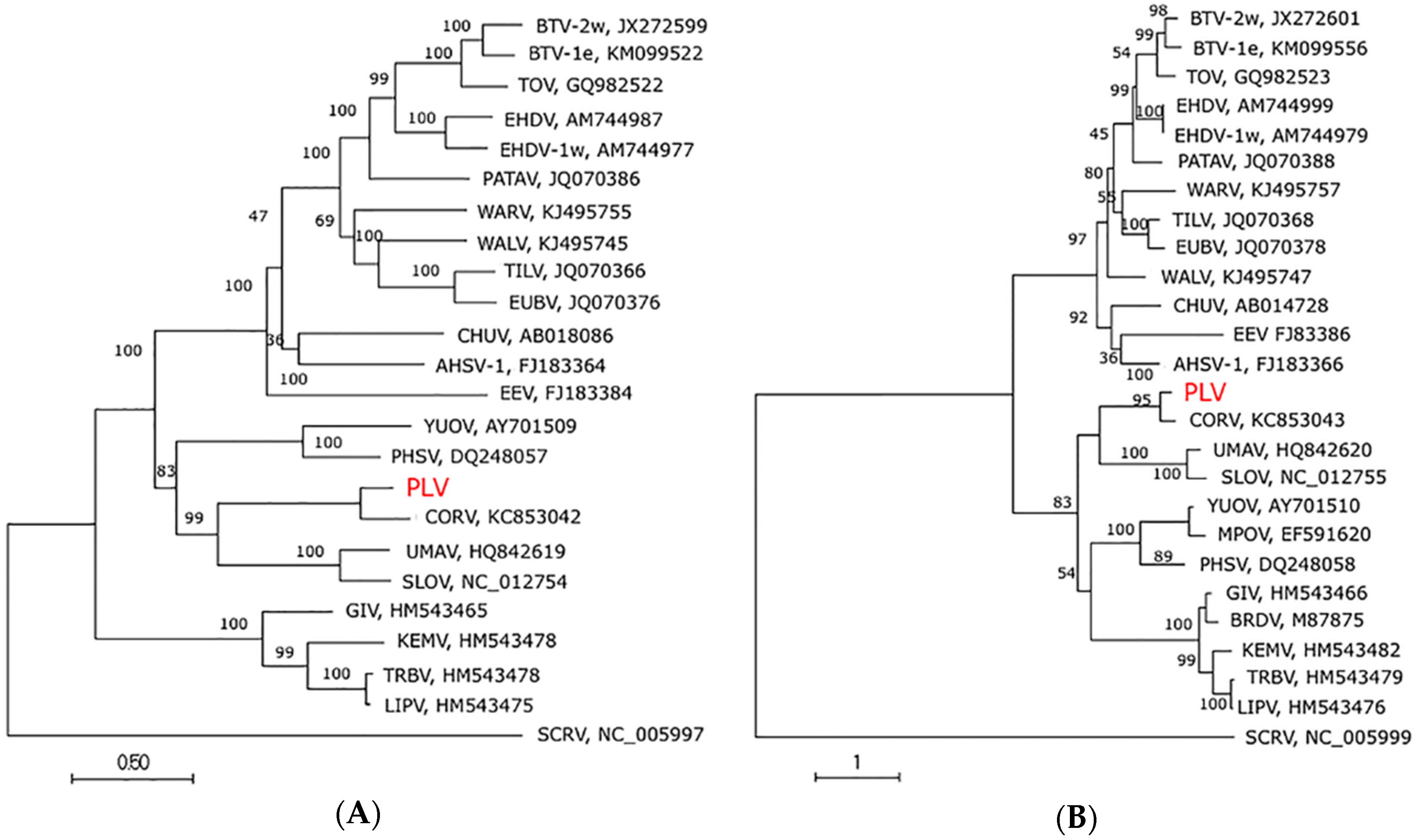
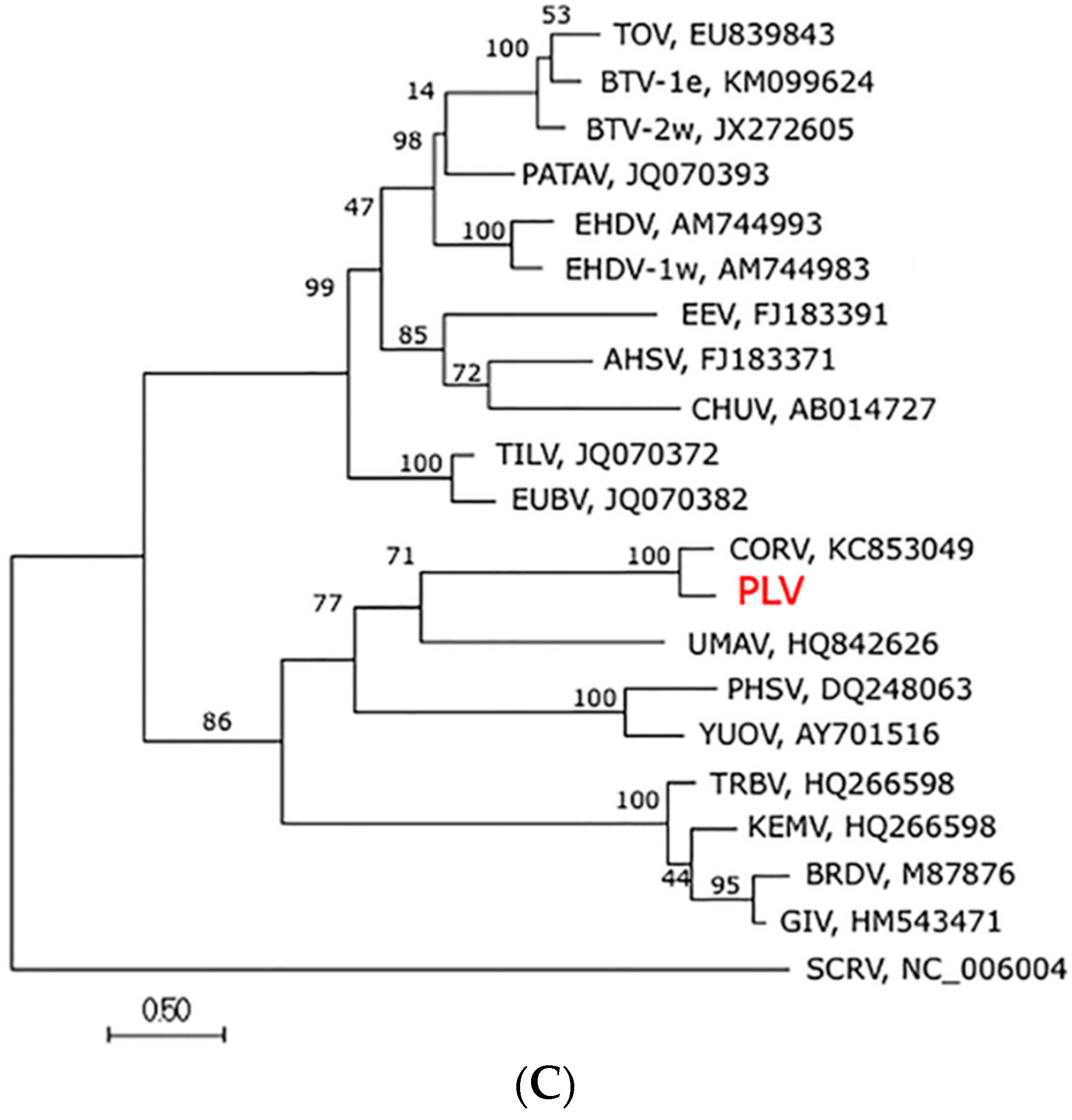
3.4. Genome Sequence, Organisation and Phylogenetics
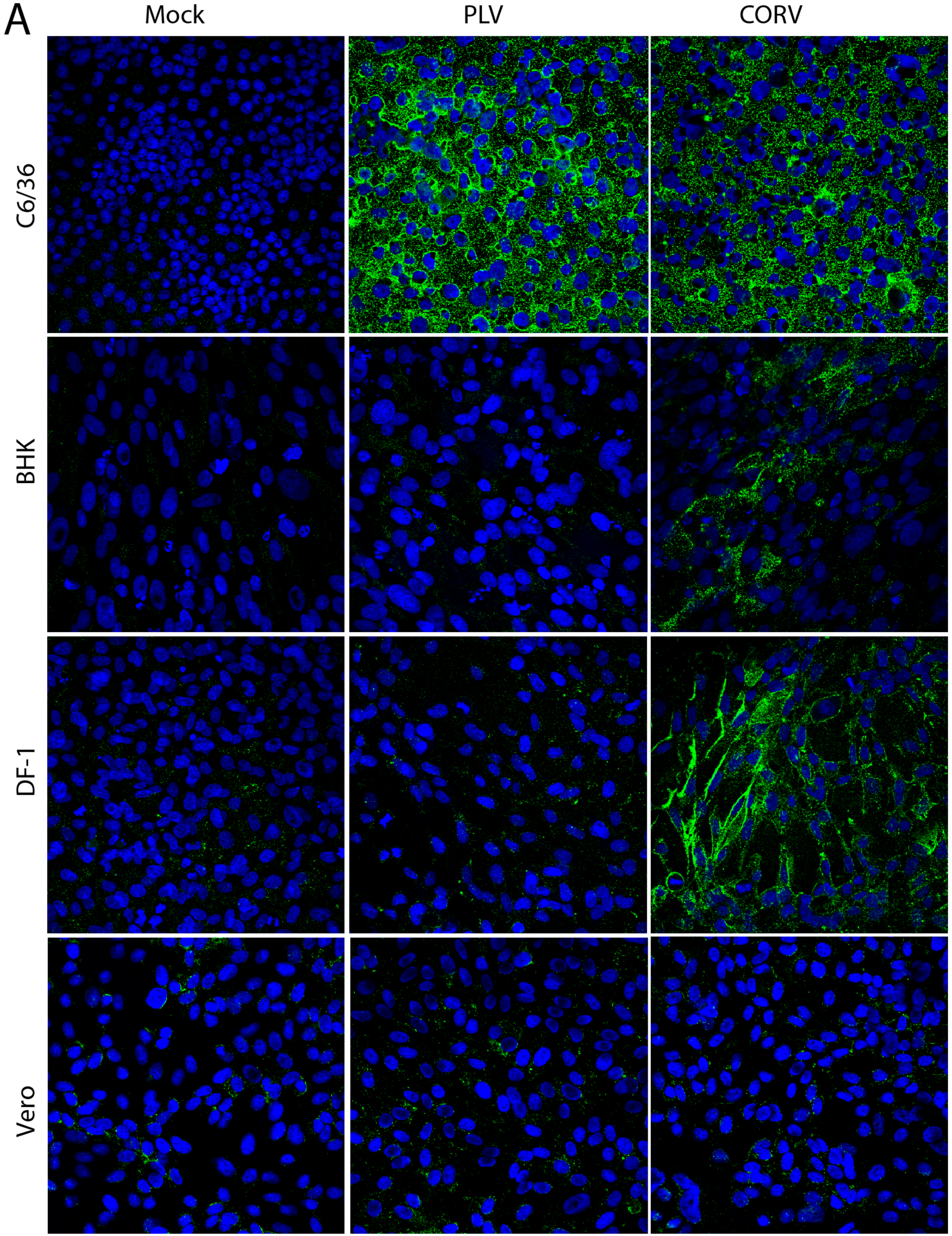
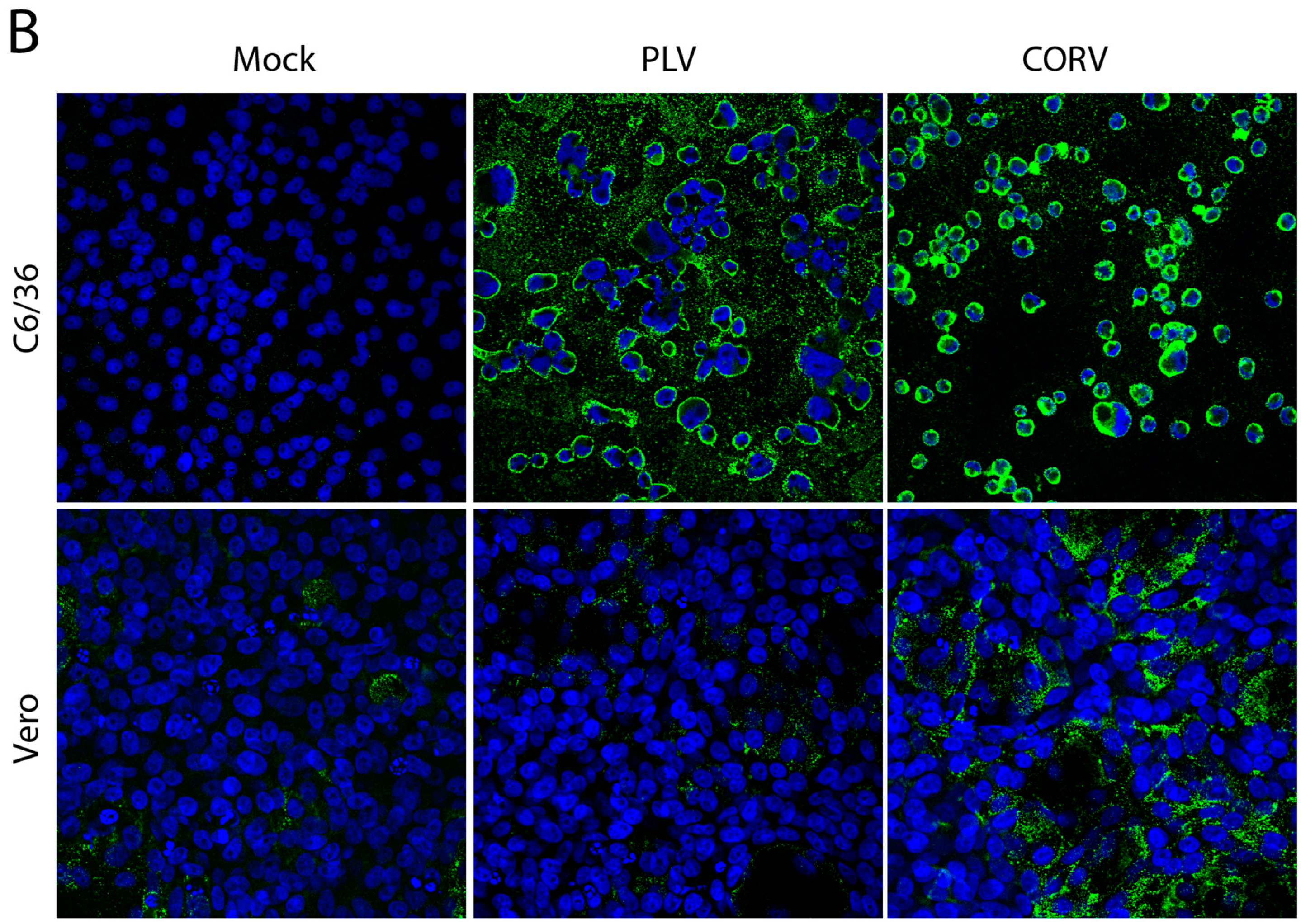
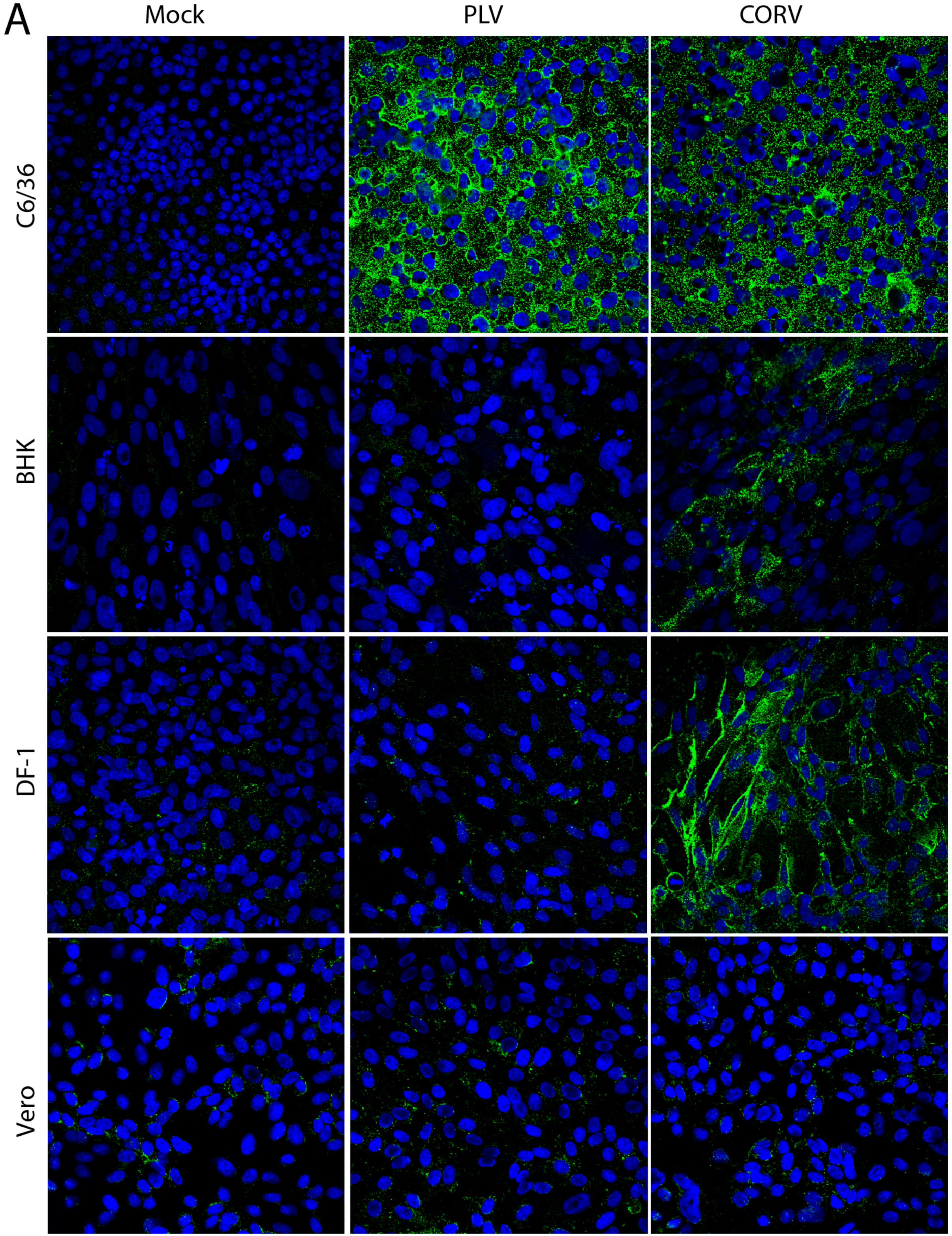
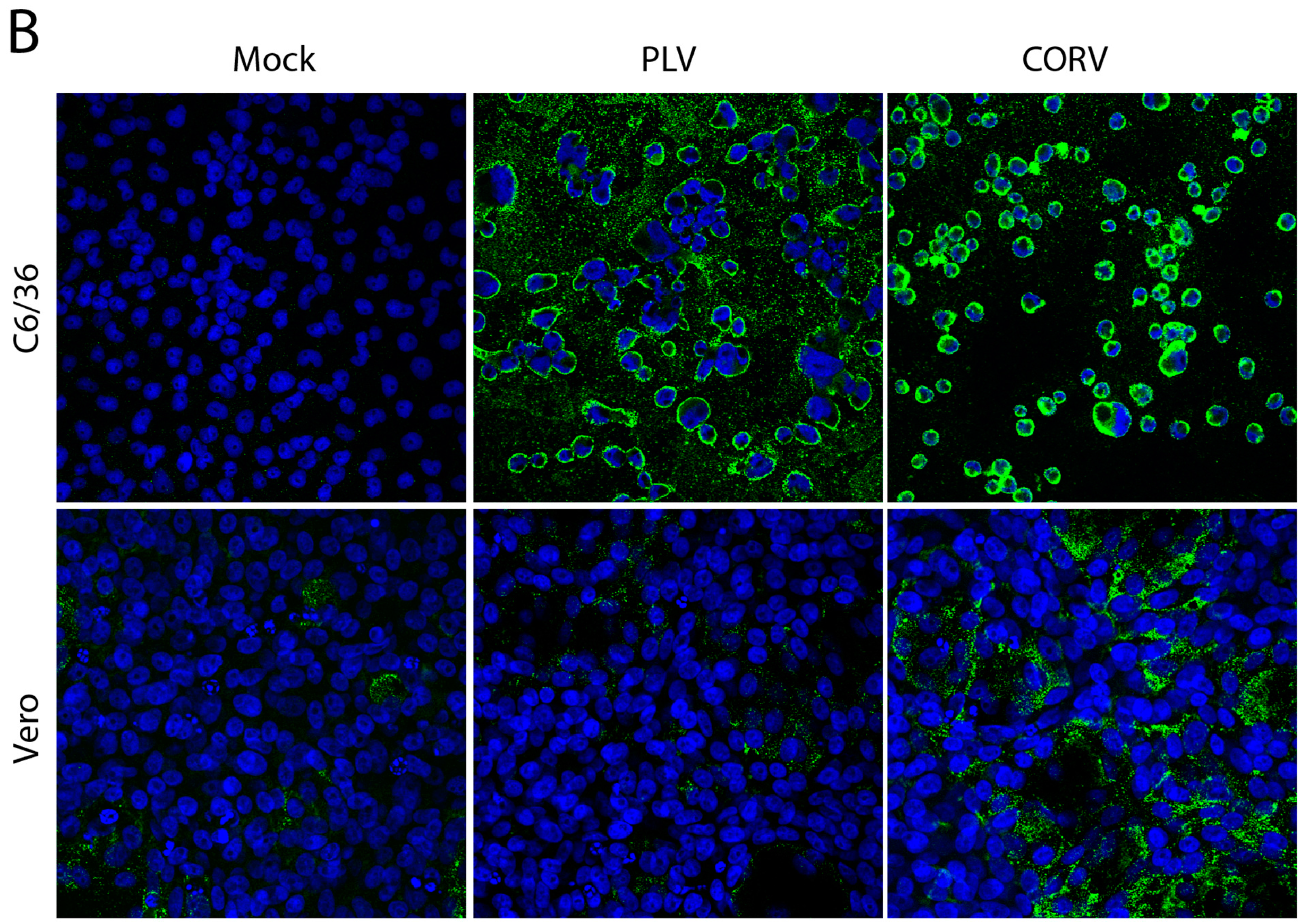
3.5. PLV Does Not Replicate in Vertebrate Cells
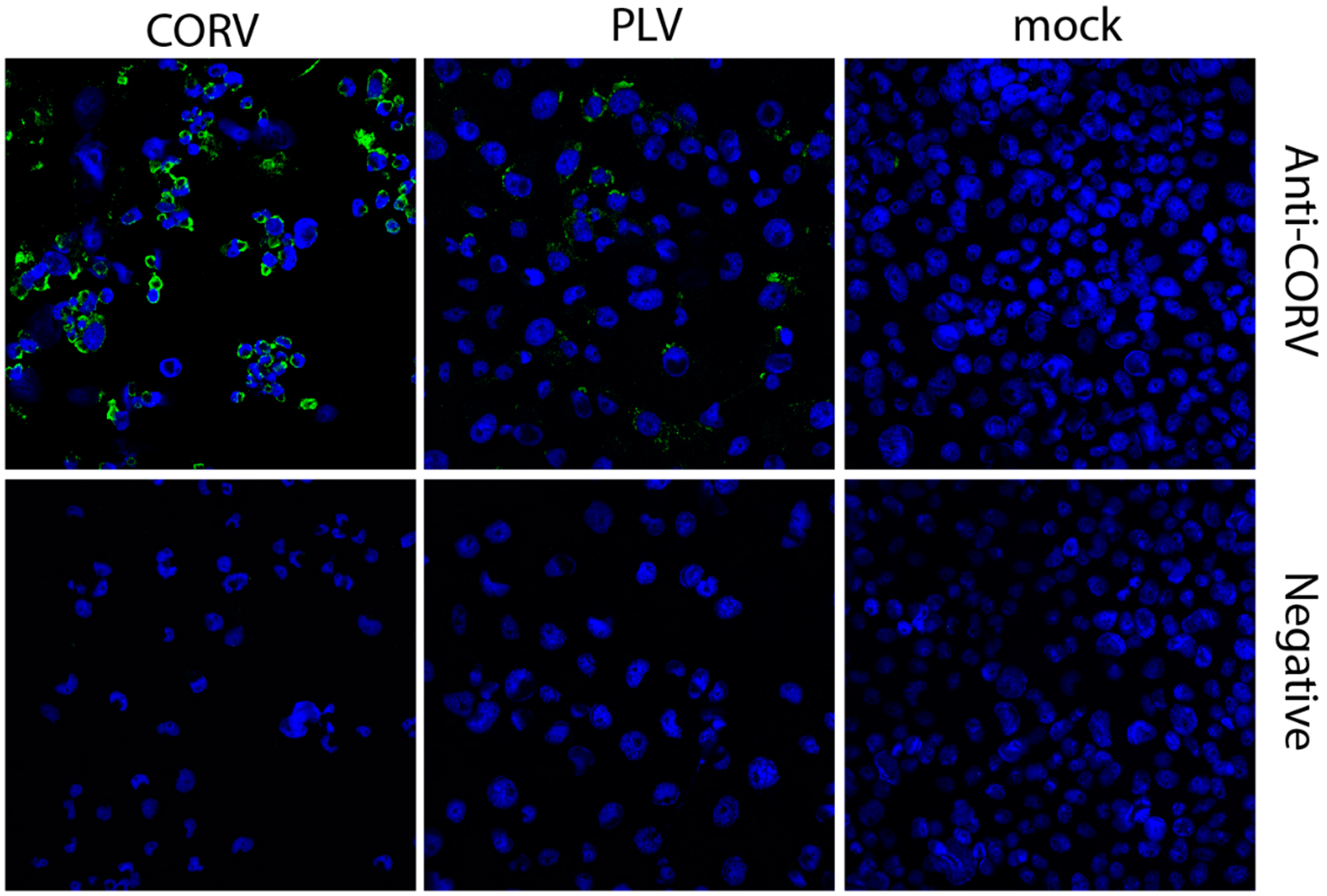
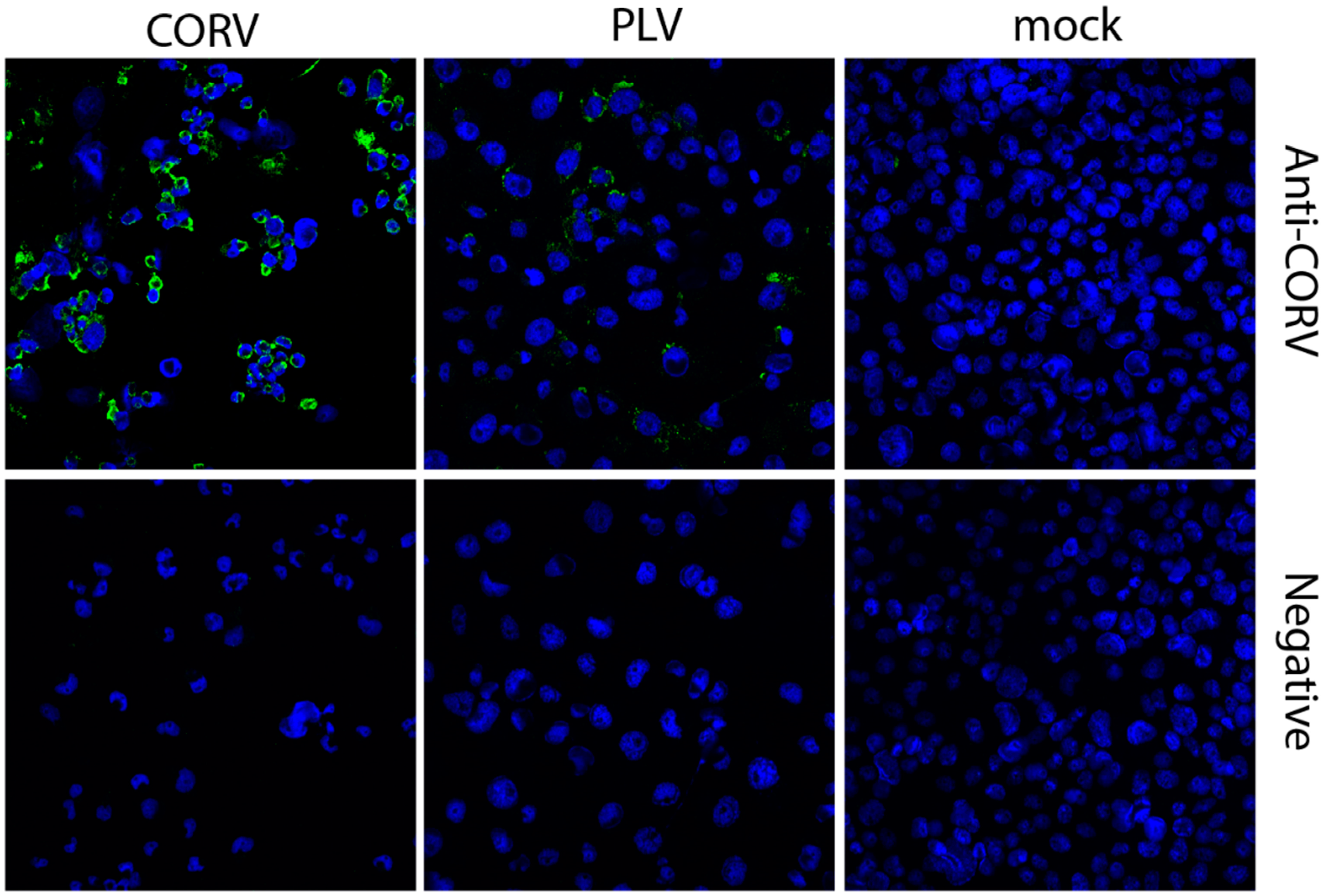
3.6. Corriparta Antiserum Cross-Reacts with and Neutralizes Parry’s Lagoon Virus
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Attoui, H.; Mertens, P.; Becnel, J.; Belaganahalli, S.; Bergoin, M.; Brussaard, C.; Chappell, J.; Ciarlet, M.; del Vas, M.; Dermody, T.; et al. Family: Reoviridae. In Virus Taxonomy Classification and Nomenclature of Viruses, Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier Academic Press: San Diego, CA, USA, 2012. [Google Scholar]
- Attoui, H.; Mendez-Lopez, M.; Rao, S.; Hurtado-Alendes, A.; Lizaraso-Caparo, F.; Jaafar, F.; Samuel, A.; Belhouchet, M.; Pritchard, L.I.; Melville, L.; et al. Peruvian horse sickness virus and Yunnan orbivirus, isolated from vertebrates and mosquitoes in Peru and Australia. Virology 2009, 394, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Coffey, L.; Page, B.; Greninger, A.; Herring, B.; Russell, R.; Doggett, S.; Haniotis, J.; Wang, C.; Deng, X.; Delwart, E.L. Enhanced arbovirus surveillance with deep sequencing: Identification of novel rhabdoviruses and bunyaviruses in australian mosquitoes. Virology 2014, 448, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Cowled, C.; Melville, L.; Weir, R.; Walsh, S.; Gubala, A.; Davis, S.; Boyle, D. Persistent and recrudescent infectin in cattle following natural infection with Middle Point orbivirus. Arch. Virol. 2012, 157, 1161–1165. [Google Scholar] [CrossRef] [PubMed]
- Doherty, R.; Whitehead, R.; Wetters, E.; Gorman, B.; Carley, J. A survey of antibody to 10 arboviruses (Koongol group, Mapputta group and ungrouped) isolated in Queensland. Trans. R. Soc. Trop. Med. Hyg. 1970, 64, 748–753. [Google Scholar] [CrossRef]
- Standfast, H.; Dyce, A.; St. George, T.; Muller, M.; Doherty, R.; Carley, J.; Filippich, C. Isolation of arboviruses ffrom insects collected at Beatrice hill, Northern Territory of Australia, 1974–1976. Aust. J. Biol. Sci. 1984, 37, 351–366. [Google Scholar] [PubMed]
- Cowled, C.; Palacios, G.; Melville, L.; Weir, R.; Walsh, S.; Davis, S.; Gubala, A.; Lipkin, W.; Briese, T.; Boyle, D. Genetic and epidemiological characterization of Stretch Lagoon orbivirus, a novel orbivirus isolated from Culex and Aedes mosquitoes in northern Australia. J. Gen. Virol. 2009, 90, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
- Liehne, P.; Anderson, S.; Stanley, N.; Liehne, C.; Wright, A.; Chan, K.; Leivers, S.; Britten, D.; Hamilton, N. Isolation of Murray Valley encephalitis virus and other arboviruses in the Ord River Valley 1972–1976. Aust. J. Exp. Biol. Med. Sci. 1981, 59, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Agosto, M.; Ivanovic, T.; Nibert, M. Mammalian reovirus, a nonfusogenic nonenveloped virus, forms size-selective pores in a model membrane. Proc. Nalt. Acad. Sci. USA 2006, 103, 16496–16501. [Google Scholar] [CrossRef] [PubMed]
- Cowled, C.; Melville, L.; Weir, R.; Walsh, S.; Hyatt, A.; van Driel, R.; Davis, S.; Gubala, A.; Boyle, D. Genetic and epidemiological characterisation of middle point orbivirus, a novel virus isolated from sentinel cattle in northern Australia. J. Gen. Virol. 2007, 88, 3413–3422. [Google Scholar] [CrossRef] [PubMed]
- Belaganahalli, M.; Maan, S.; Maan, N.; Nomikou, K.; Guimera, M.; Brownlie, J.; Tesh, R.; Attoui, H.; Mertens, P. Full genome sequencing of Corriparta virus, identifies California mosquito pool virus as a member of the Corriparta virus species. PLoS ONE 2013, 8, e70779. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, C.; Hobson-Peters, J.; Yam, A.; Colmant, A.; McLean, B.; Prow, N.; Watterson, D.; Hall-Mendelin, S.; Warrilow, D.; Ng, M.; et al. Viral RNA intermediates as targets for detection and discovery of novel and emerging mosquito-borne viruses. PLoS Negl. Trop. Dis. 2015, 9, e0003629. [Google Scholar] [CrossRef] [PubMed]
- Broom, A.; Lindsay, M.; Plant, A.; Wright, A.; Condon, R.; Mackenzie, J. Epizootic activity of Murray Valley encephalitis virus in an aboriginal community in the southeast region of Western Australia: Results of cross-sectional and longitudinal serologic studies. Am. J. Trop. Med. Hyg. 2002, 67, 319–323. [Google Scholar] [PubMed]
- Liehne, P.; Stanley, N.; Alpers, M.; Liehne, C. Ord River arboviruses—The study site and mosquitoes. Aust. J. Exp. Biol. Med. Sci. 1976, 54, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Johansen, C.; Broom, A.; Lindsay, M.; Avery, V.; Power, S.; Dixon, G.; Sturrock, K.; Maley, F.; Mcfall, S.; Geerlings, K.; et al. Arbovirus and vector surveillance in Western Australia, 2004/05 to 2007/08. Arbovirus Res. Aust. 2009, 10, 76–81. [Google Scholar]
- Broom, A.; Wright, A.; Mackenzie, J.; Lindsay, M.; Robinson, D. Isolation of murray valley encephalitis and ross river viruses from Aedes normanensis (Diptera: Culicidae) in Western Australia. J. Med. Entomol. 1989, 26, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Johansen, C.; van den Hurk, A.; Ritchie, S.; Zborowski, P.; Nisbet, D.; Paru, R.; Bockarie, M.; Macdonald, J.; Drew, A.; Khromykh, T.; et al. Isolation of Japansese encephalitis virus from mosquitoes (Diptera: Culicidae) collected in the Western Province of Papua New Guinea, 1997–1998. Am. J. Trop. Med. Hyg. 2000, 62, 631–638. [Google Scholar] [PubMed]
- Lindsay, M.; Broom, A.; Wright, A.; Johansen, C.; Mackenzie, J. Ross River virus isolations from mosquitoes in arid regions of Western Australia: Implication of vertical transmission as a means of persistence of the virus. Am. J. Trop. Med. Hyg. 1993, 49, 686–696. [Google Scholar] [PubMed]
- Broom, A.; Hall, R.; Johansen, C.; Oliveira, N.; Howard, M.; Lindsay, M.; Kay, B.; Mackenzie, J. Identification of Australian arboviruses in inoculated cell cultures using monoclonal antibodies in ELISA. Pathology 1998, 30, 286–288. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Edgar, R. Muscle: Multiple sequence alignment with high accuracy and high throughout. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A simple, fast and accurate method to estimate large phylogenies by maximum-liklihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Hobson-Peters, J.; Yam, A.; Lu, J.; Setoh, Y.; May, F.; Kurucz, N.; Walsh, S.; Prow, N.; Davis, S.; Weir, R.; et al. A new insect-specific flavivirus from northern Australia suppresses replication of West Nile virus and Murray Valley enephalitis virus in co-infected mosquito cells. PLoS ONE 2013, 8, e56534. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.; Kay, B.; Burgess, G. An enzyme immunoassay to detect Australian flaviviruses and identify the encephalitic subgroup using monoclonal antibodies. Immunol. Cell Biol. 1987, 65, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Jaafar, F.; Belhouchet, M.; Aldrovandi, N.; Tao, S.; Chen, B.; Liang, G.; Tesh, R.; de Micco, P.; Lamballerie, X. Yunnan orbivirus, a new orbivirus species isolated from culex tritaeniorhynchus mosquitoes in China. J. Gen. Virol. 2005, 86, 3409–3417. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Mohd Jaafar, F.; Belhouchet, M.; Biagini, P.; Cantaloube, J.; de Micco, P.; de Lamballerie, X. Expansion of family Reoviridae to include nine-segmented dsRNA viruses: Isolation and characterization of a new virus designated Aedes pseudoscutellaris reovirus assigned to a proposed genus (Dinovernavirus). Virology 2005, 343, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Carley, J.; Standfast, H. Corriparta virus: Properties and multiplication in experimentally-inoculated mosquitoes. Am. J. Epidemiol. 1969, 89, 583–592. [Google Scholar] [PubMed]
- Attoui, H.; Stirling, J.; Munderloh, U.; Billoir, F.; Brookes, S.; Burroughs, J.; di Micco, P.; Mertens, P.; de Lamballerie, X. Complete sequence characterization of the genome of the St Croix River virus, a new orbivirus isolated from cells of Ixodes scapularis. J. Gen. Virol. 2001, 82, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Doherty, R.; Carley, J.; Mackerras, M.; Marks, E. Studies of arthropod-borne virus infections in Queensland III. Isolation and characterisation of virus strains from wild-caught mosquitoes in North Queensland. Aust. J. Exp. Biol. Med. Sci. 1963, 41, 17–40. [Google Scholar] [CrossRef] [PubMed]
- Gumm, I.; Newman, J. The preparation of purified bluetongue virus group antigen for use as a diagnostic reagent. Arch. Virol. 1982, 72, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, H.; Knudson, D. Orbivirus species and speciation: Genetic reassortment between Corriparta Serogroup viruses. Intervirology 1987, 28, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Cybinski, D.; Muller, M. Isolation of arboviruses from cattle and insecs at two sentinel sites in Queensland, Australia, 1979–1985. Aust. J. Zool. 1990, 38, 25–32. [Google Scholar] [CrossRef]
- Lutomiah, J.; Mwandawiro, C.; Magambo, J.; Sang, R. Infection and vertical transmission of Kamiti river virus in laboratory bred Aedes aegypti mosquitoes. J. Insect Sci. 2007, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- McLean, B.; Hobson-Peters, J.; Webb, C.; Watterson, D.; Prow, N.; Nguyen, H.; Hall-Mendelin, S.; Warrilow, D.; Johansen, C.; Jansen, C.; et al. A novel insect-specific flavivirus replicates only in Aedes-derived cells and persists at high prevalence in wild Aedes vigilax populations in Syndey, Australia. Virology 2015, 486, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Saiyasombat, R.; Bolling, B.; Brault, A.; Bartholomay, L.; Blitvich, B. Evidence of efficient transovarial transmission of Culex flavivirus by Culex Pipiens (Diptera: CULICIDAE). J. Med. Entomol. 2011, 48, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Marklewitz, M.; Handrick, S.; Grasse, W.; Kurth, A.; Lukashev, A.; Drosten, C.; Ellerbrok, H.; Leendertz, F.; Pauli, G.; Junglen, S. Gouléako virus isolated from west African mosquitoes constitues a proposed novel genus in the family bunyaviridae. J. Virol. 2011, 85, 9227–9924. [Google Scholar] [CrossRef] [PubMed]
- Marklewitz, M.; Zirkel, F.; Rwego, I.B.; Heidemann, H.; Trippner, P.; Kurth, A.; Kallies, R.; Briese, T.; Lipkin, W.I.; Drosten, C.; et al. Discovery of a unique novel clade of mosquito-associated bunyaviruses. J. Virol. 2013, 87, 12850–12865. [Google Scholar] [CrossRef] [PubMed]
- Auguste, A.; Kaelber, J.; Fokam, E.; Guzman, H.; Carrington, C.; Erasmus, J.; Kamgang, B.; Popov, V.; Jakana, J.; Liu, X.; et al. A newly-isolated reovirus has the simplest genomic and structural organization of any reovirus. J. Virol. 2014, 89, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Chastel, C.; Main, A.; Couatarmanac’h, A.; le Lay, G.; Knudson, D.; Quillien, M.; Beaucournu, J. Isolation of Eyach virus (Reoviridae, Colorado tick fever group) from Ixodes ricinus and I. Ventalloi ticks in France. Arch. Virol. 1984, 82, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Hermanns, K.; Zirkel, F.; Kurth, A.; Drosten, C.; Junglen, S. Cimodo virus belongs to a novel lineage of reoviruses isolated from African mosquitoes. J. Gen. Virol. 2014, 95, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Spear, A.; Sisterson, M.; Stenger, D. Reovirus genomes from plant feeding insects represent a newly discovered lineage with the family Reoviridae. Virus Res. 2012, 163, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Aliota, M.; Jones, S., II; Dupuis, A.; Ciota, A.; Hubalek, Z.; Kramer, L. Characterisation of Rabensburg virus, a Flavivirus closely related to West Nile virus of the Japanese encephalitis group. PLoS ONE 2012, 7, e39387. [Google Scholar] [CrossRef] [PubMed]
- Aliota, M.; Kramer, L. Replication of West Nile virus, rabensburg lineage in mammalian cells is restricted by temperature. Parasites Vectors 2012, 5, 293–301. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pool ID | Mosquito Species | Date Collected | Initial Isolation | Re-Isolation | % Pairwise Identity to Prototype (nt/aa) e | ||
|---|---|---|---|---|---|---|---|
| CPE | CPE | CPE | CPE | ||||
| C6/36 a | Vero b | PS-EK b | C6/36 c | ||||
| Kununurra | |||||||
| K74511 | Culex pullus | April 2011 | + | − | − | + | 99.75/99.25 |
| K74544 | Culex pullus | April 2011 | + | − | − | + | ND |
| K74770 | Culex annulirostris | April 2011 | + | − | − | + | 99.75/99.25 |
| K74775 | Culex species | April 2011 | + | − | − | + | 99.75/99.25 |
| K75023 | Culex annulirostris | April 2011 | + | − | − | + | 99.75/99.25 |
| Parry’s Creek | |||||||
| K71497 | Culex annulirostris | March 2010 | − | − | − | + | 97.47/95.49 |
| K71516 | Culex annulirostris | March 2010 | − | − | − | − | 97.74/95.49 |
| K71520 | Culex annulirostris | March 2010 | − | − | − | − | 97.74/95.49 |
| K71551 | Culex annulirostris | March 2010 | − | − | − | + | 100/100 |
| K71558 | Culex annulirostris | March 2010 | − | − | − | − | 97.74/95.49 |
| K71435 | Culex annulirostris | March 2010 | − | − | − | + | 99.50/98.50 |
| K75533 | Mansonia uniformis | April 2011 | + | − | − | + | 99.75/99.25 |
| Billiluna | |||||||
| K75737 | Aedes normanensis | April 2011 | + | − | − | + | 99.75/99.25 |
| K75749 d | Culex annulirostris | April 2011 | + | + d | + d | + | ND |
| Segment | Protein Encoding | G+C Content (%) | ORF Length (bp) | Predicted Protein Length (aa) | % Pairwise Identity with CORV (nt/aa) |
|---|---|---|---|---|---|
| 1 | VP1 (Pol) | 40.8 | 3873 | 1290 | 78.1/91.9 |
| 2 | VP2 (T2) | 41.8 | 2850 | 949 | 79.9/94.3 |
| 3 | VP3 (OC1) | 42.2 | 2217 | 738 | 74.7/85.8 |
| 4 | NS1 (TuP) | 48.0 | 1770 | 589 | 78.0/89.8 |
| 5 | VP5 (OC2) | 45.3 | 1584 | 527 | 79.1/95.3 |
| 6 | VP4 (Cap) | 44.4 | 1932 | 643 | 78.1/90.4 |
| 7 | NS2 (ViP) | 46.2 | 1116 | 371 | 81.1/92.7 |
| 8 | VP7 (T13) | 46.8 | 1065 | 352 | 79.2/94.3 |
| 9 | VP6 (Hel)/NS4 | 46.0 | 1035 | 344 | 78.0/72.6 |
| 10 | NS3/NS3a | 46.7 | 717 | 238 | 81.2/92.0 |
| Percent Identity a | |||
|---|---|---|---|
| Protein | Virus | CORV | PLV |
| VP2 | CMPV | 92 | 87 |
| CORV | 95 | ||
| VP4 | CMPV | 70 | 71 |
| CORV | 90 | ||
| VP5 | CMPV | 83 | 84 |
| CORV | 95 | ||
| VP7 | CMPV | 92 | 94 |
| CORV | 94 | ||
| VP6 | CMPV | 39 | 42 |
| CORV | 73 | ||
| NS4 | CMPV | 56 | 58 |
| CORV | 77 | ||
| Virus | CORV Antiserum 1 (B97) | CORV Antiserum 2 (CSIRO109) | Negative |
|---|---|---|---|
| CORV | 80 | 320 | <10 |
| PLV | 20 | >1280 | <10 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harrison, J.J.; Warrilow, D.; McLean, B.J.; Watterson, D.; O’Brien, C.A.; Colmant, A.M.G.; Johansen, C.A.; Barnard, R.T.; Hall-Mendelin, S.; Davis, S.S.; et al. A New Orbivirus Isolated from Mosquitoes in North-Western Australia Shows Antigenic and Genetic Similarity to Corriparta Virus but Does Not Replicate in Vertebrate Cells. Viruses 2016, 8, 141. https://doi.org/10.3390/v8050141
Harrison JJ, Warrilow D, McLean BJ, Watterson D, O’Brien CA, Colmant AMG, Johansen CA, Barnard RT, Hall-Mendelin S, Davis SS, et al. A New Orbivirus Isolated from Mosquitoes in North-Western Australia Shows Antigenic and Genetic Similarity to Corriparta Virus but Does Not Replicate in Vertebrate Cells. Viruses. 2016; 8(5):141. https://doi.org/10.3390/v8050141
Chicago/Turabian StyleHarrison, Jessica J., David Warrilow, Breeanna J. McLean, Daniel Watterson, Caitlin A. O’Brien, Agathe M.G. Colmant, Cheryl A. Johansen, Ross T. Barnard, Sonja Hall-Mendelin, Steven S. Davis, and et al. 2016. "A New Orbivirus Isolated from Mosquitoes in North-Western Australia Shows Antigenic and Genetic Similarity to Corriparta Virus but Does Not Replicate in Vertebrate Cells" Viruses 8, no. 5: 141. https://doi.org/10.3390/v8050141
APA StyleHarrison, J. J., Warrilow, D., McLean, B. J., Watterson, D., O’Brien, C. A., Colmant, A. M. G., Johansen, C. A., Barnard, R. T., Hall-Mendelin, S., Davis, S. S., Hall, R. A., & Hobson-Peters, J. (2016). A New Orbivirus Isolated from Mosquitoes in North-Western Australia Shows Antigenic and Genetic Similarity to Corriparta Virus but Does Not Replicate in Vertebrate Cells. Viruses, 8(5), 141. https://doi.org/10.3390/v8050141