
The Iminosugar UV-4 is a Broad Inhibitor of Influenza A and B Viruses ex Vivo and in Mice

Abstract
:1. Introduction
2. Materials and Methods
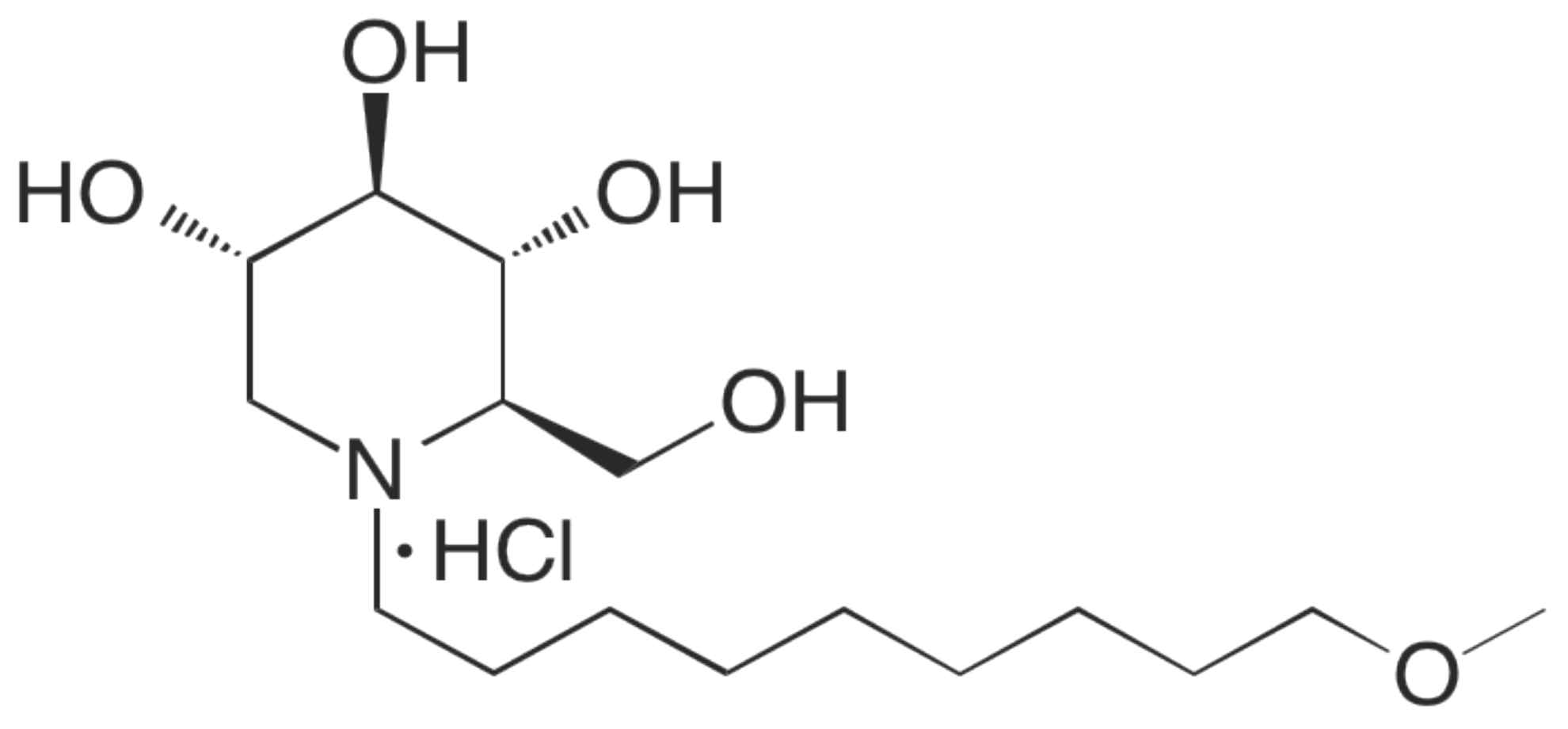
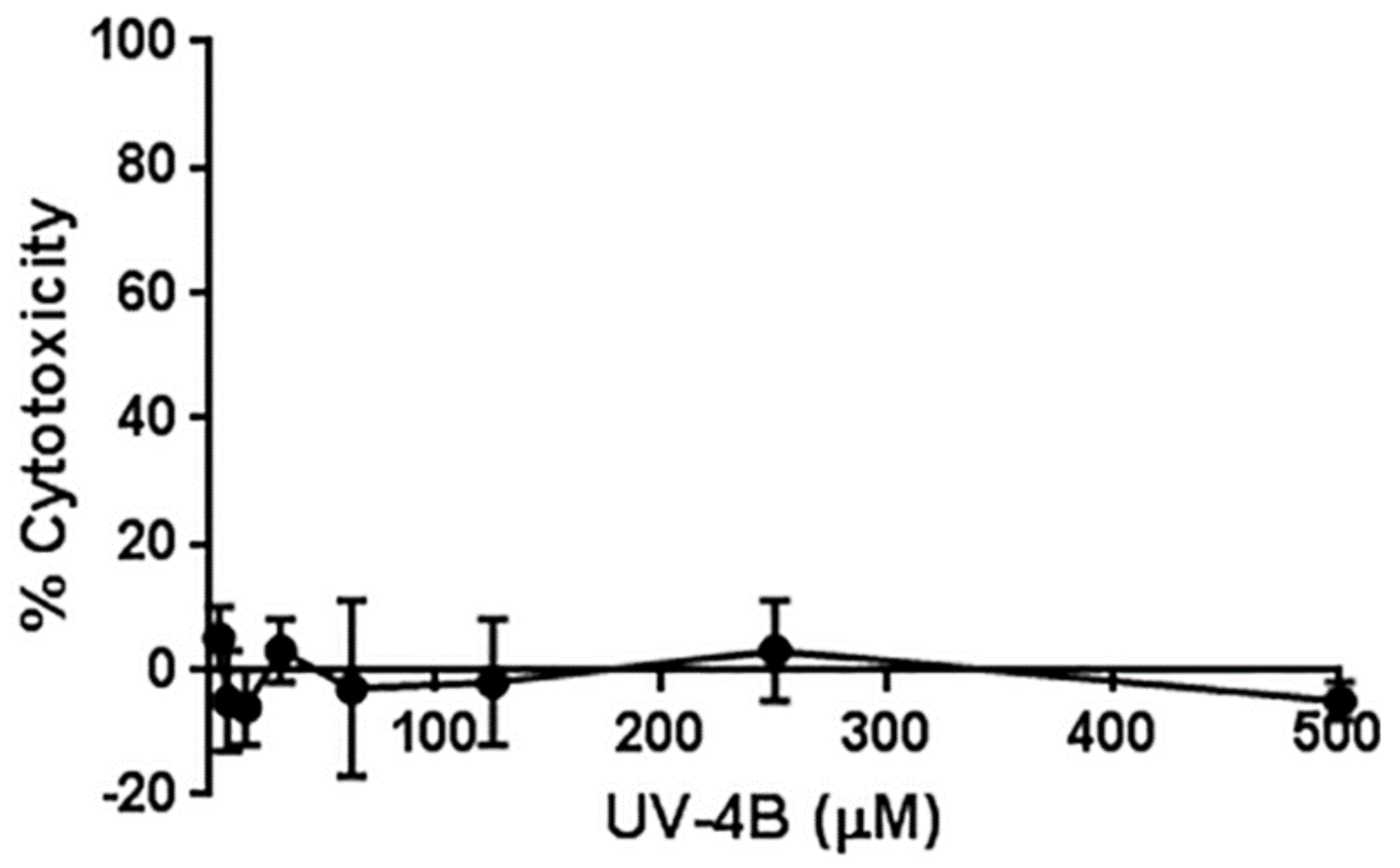
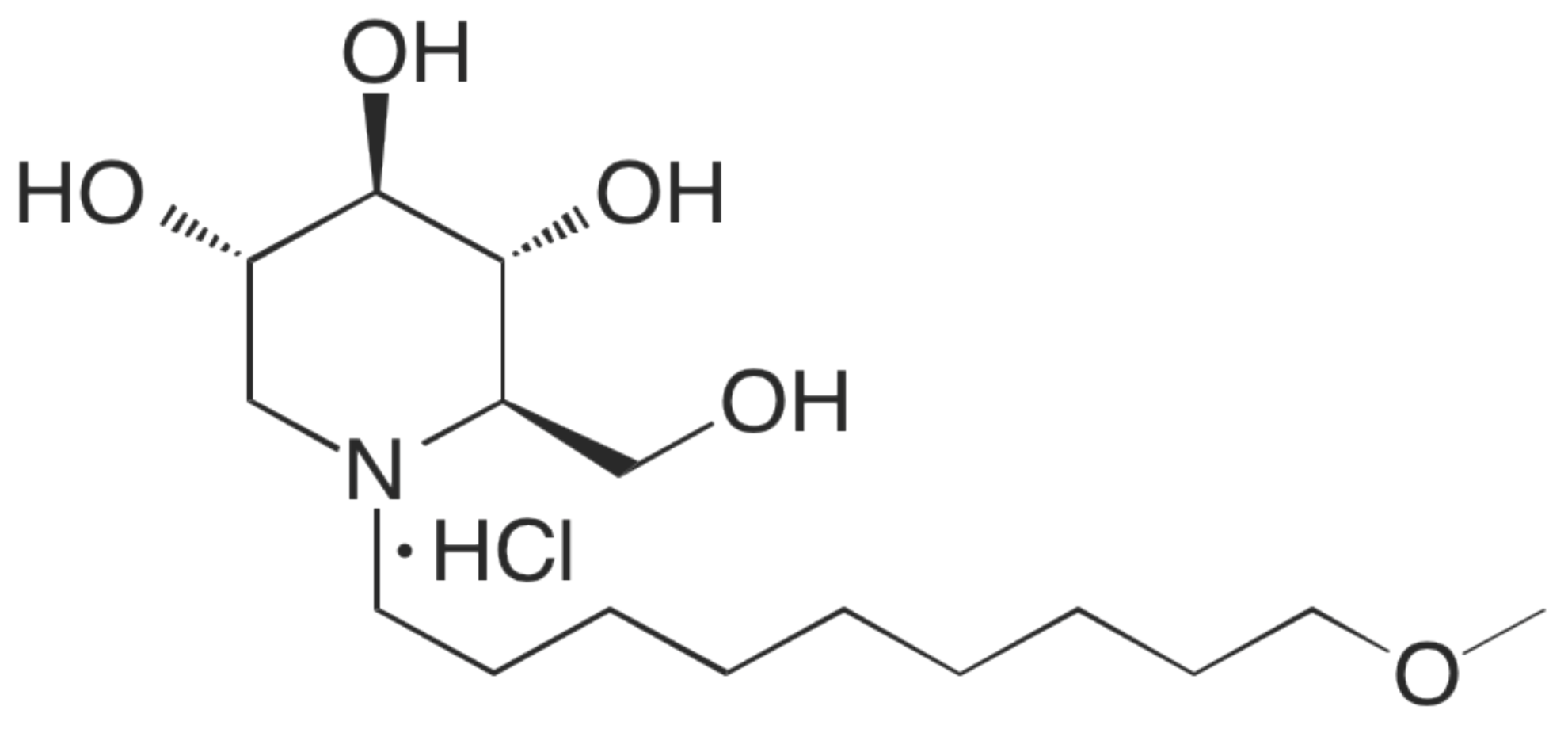
2.1. Cytotoxicity Assessment of UV-4B
2.2. Antiviral Activity in Primary Differentiated Normal Human Bronchial Epithelial (dNHBE) Cells
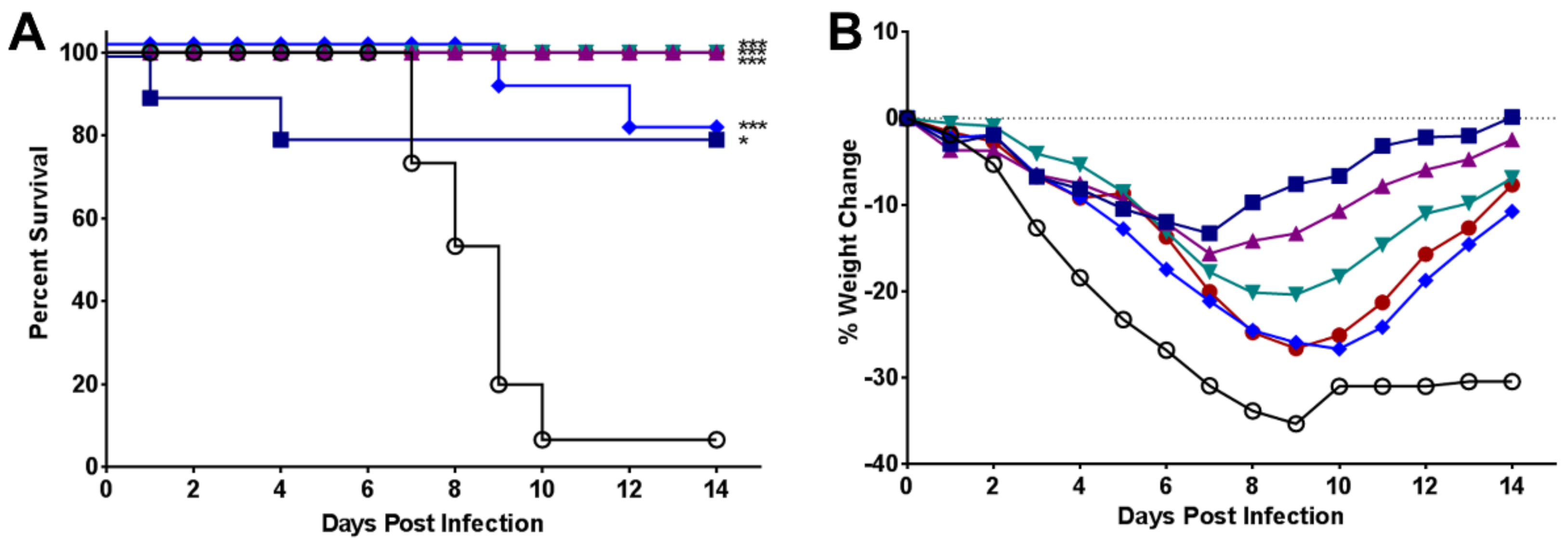
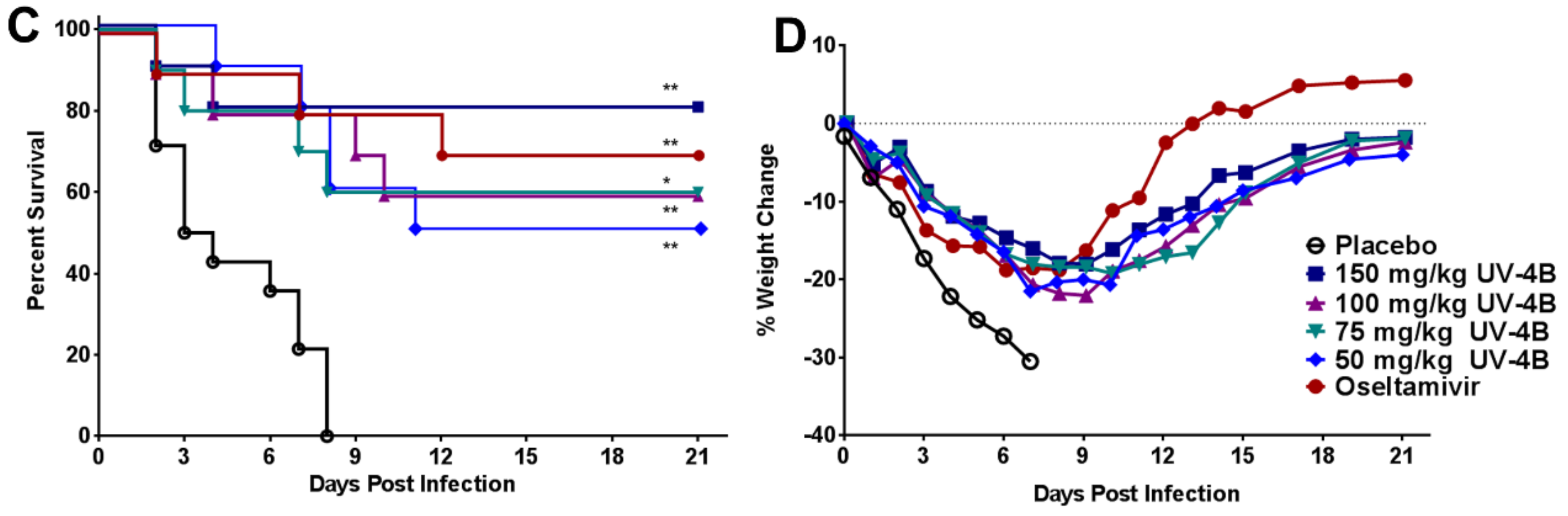
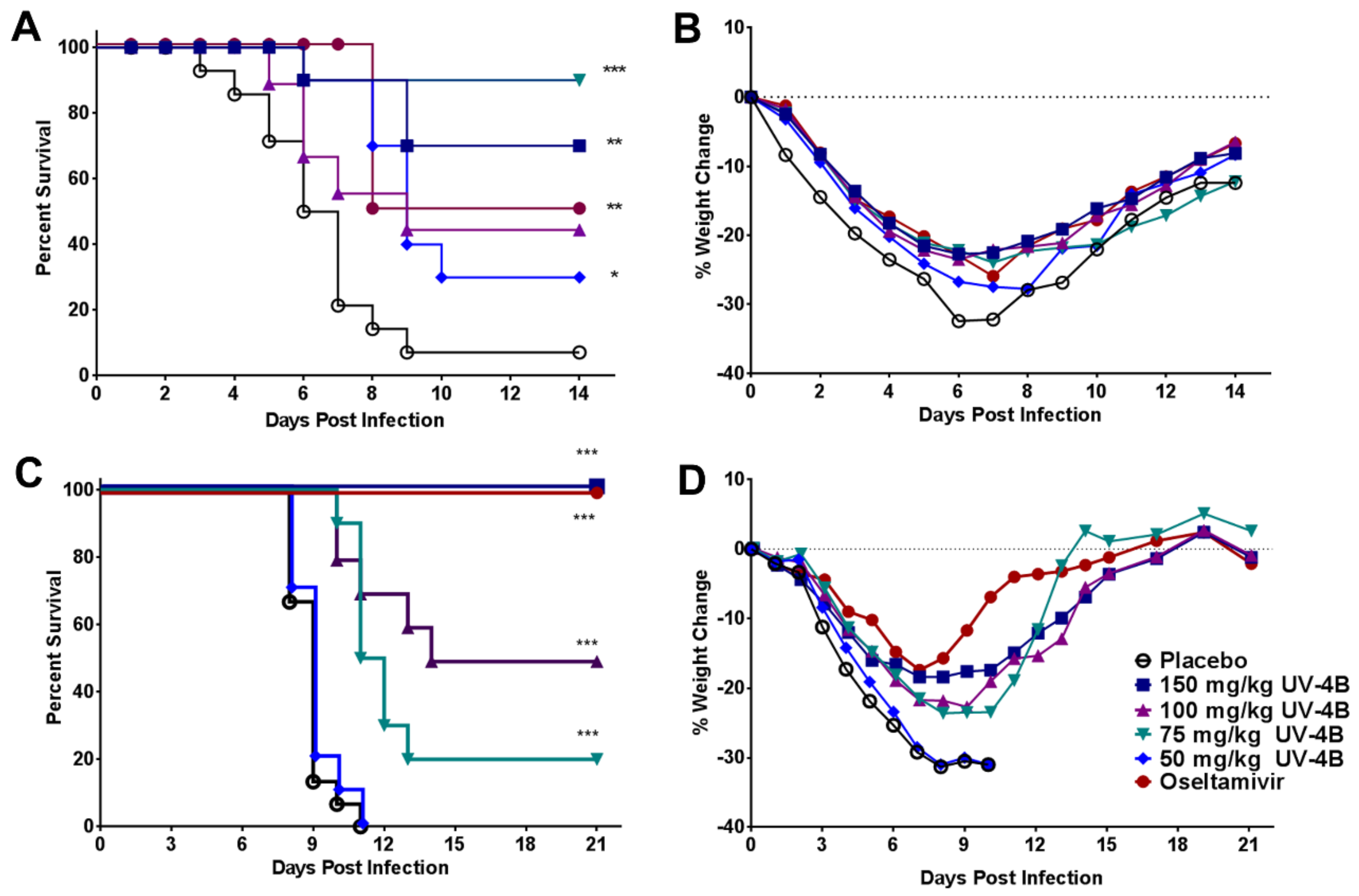
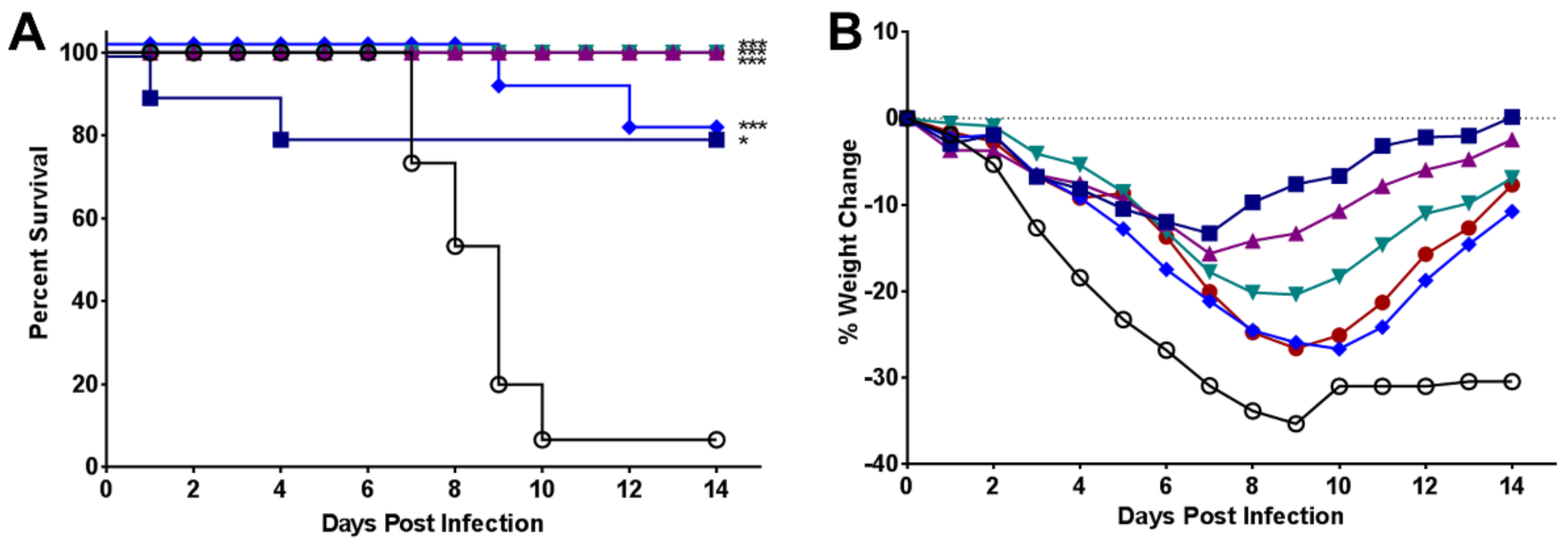
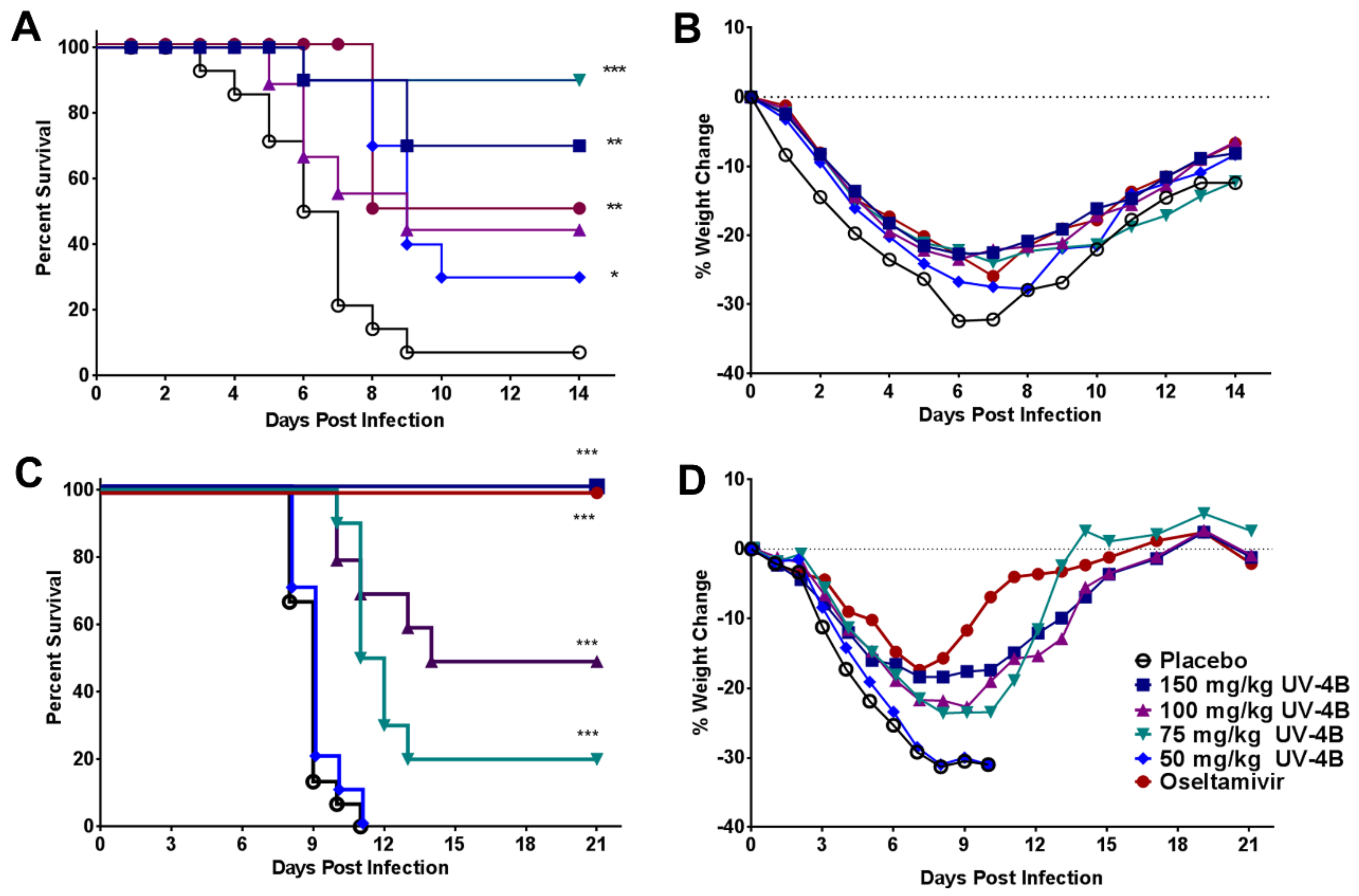
2.3. Efficacy of UV-4B in Lethal Mouse Models
3. Results
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Influenza (seasonal), Fact Sheet no211, March 2014. Available online: http://www.who.int/mediacentre/factsheets/fs211/en/ (accessed on 18 August 2015).
- Watanabe, T.; Watanabe, S.; Maher, E.A.; Neumann, G.; Kawaoka, Y. Pandemic potential of avian influenza a (h7n9) viruses. Trends Microbiol. 2014, 22, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Pascua, P.N.; Choi, Y.K. Zoonotic infections with avian influenza a viruses and vaccine preparedness: A game of “mix and match”. Clin. Exp. Vaccine Res. 2014, 3, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Loregian, A.; Mercorelli, B.; Nannetti, G.; Compagnin, C.; Palu, G. Antiviral strategies against influenza virus: Towards new therapeutic approaches. Cell. Mol. Life Sci. 2014, 71, 3659–3683. [Google Scholar] [CrossRef] [PubMed]
- Hurt, A.C.; Hardie, K.; Wilson, N.J.; Deng, Y.M.; Osbourn, M.; Leang, S.K.; Lee, R.T.; Iannello, P.; Gehrig, N.; Shaw, R.; et al. Characteristics of a widespread community cluster of h275y oseltamivir-resistant a(h1n1)pdm09 influenza in Australia. J. Infect. Dis. 2012, 206, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Zumla, A.; Memish, Z.A.; Maeurer, M.; Bates, M.; Mwaba, P.; Al-Tawfiq, J.A.; Denning, D.W.; Hayden, F.G.; Hui, D.S. Emerging novel and antimicrobial-resistant respiratory tract infections: New drug development and therapeutic options. Lancet Infect. Dis. 2014, 14, 1136–1149. [Google Scholar] [CrossRef]
- Lee, S.M.; Yen, H.L. Targeting the host or the virus: Current and novel concepts for antiviral approaches against influenza virus infection. Antivir. Res. 2012, 96, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Zitzmann, N.; Block, T.; Methta, A.; Rudd, P.; Burton, D.; Wilson, I.; Platt, F.; Butters, T.; Dwek, R.A. Glycosylation: Disease targets and therapy. Adv. Exp. Med. Biol. 2005, 564, 1–2. [Google Scholar] [PubMed]
- Chang, J.; Block, T.M.; Guo, J.T. Antiviral therapies targeting host er alpha-glucosidases: Current status and future directions. Antivir. Res. 2013, 99, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Dalziel, M.; Crispin, M.; Scanlan, C.N.; Zitzmann, N.; Dwek, R.A. Emerging principles for the therapeutic exploitation of glycosylation. Science 2014, 343, 1235681. [Google Scholar] [CrossRef] [PubMed]
- Helenius, A.; Aebi, M. Roles of n-linked glycans in the endoplasmic reticulum. Annu. Rev. Biochem. 2004, 73, 1019–1049. [Google Scholar] [CrossRef] [PubMed]
- Stavale, E.J.; Vu, H.; Sampath, A.; Ramstedt, U.; Warfield, K.L. In vivo therapeutic protection against influenza a (h1n1) oseltamivir-sensitive and resistant viruses by the iminosugar uv-4. PLoS ONE 2015, 10, e0121662. [Google Scholar] [CrossRef] [PubMed]
- Perry, S.T.; Buck, M.D.; Plummer, E.M.; Penmasta, R.A.; Batra, H.; Stavale, E.J.; Warfield, K.L.; Dwek, R.A.; Butters, T.D.; Alonzi, D.S.; et al. An iminosugar with potent inhibition of dengue virus infection in vivo. Antivir. Res 2013, 98, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Warfield, K.L.; Plummer, E.; Sayce, A.C.; Alonzi, D.; Tang, W.; Tyrrell, B.E.; Hill, M.L.; Caputo, A.T.; Killingbeck, S.S.; Beatty, P.R.; et al. Inhibition of endoplasmic reticulum glucosidases is required for in vitro and in vivo dengue antiviral activity by the iminosugar uv-4. Antivir. Res. 2016. [Google Scholar] [CrossRef]
- Triana-Baltzer, G.B.; Babizki, M.; Chan, M.C.; Wong, A.C.; Aschenbrenner, L.M.; Campbell, E.R.; Li, Q.X.; Chan, R.W.; Peiris, J.S.; Nicholls, J.M.; et al. Das181, a sialidase fusion protein, protects human airway epithelium against influenza virus infection: An in vitro pharmacodynamic analysis. J. Antimicrob. Chemother. 2010, 65, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.S.; Chertow, D.S.; Moyer, J.E.; Suzich, J.; Sandouk, A.; Dorward, D.W.; Logun, C.; Shelhamer, J.H.; Taubenberger, J.K. Validation of normal human bronchial epithelial cells as a model for influenza a infections in human distal trachea. J. Histochem. Cytochem. 2015, 63, 312–328. [Google Scholar] [CrossRef] [PubMed]
- Smee, D.F.; Barnard, D.L. Methods for evaluation of antiviral efficacy against influenza virus infections in animal models. Methods Mol. Biol. 2013, 1030, 407–425. [Google Scholar] [PubMed]
- Smee, D.F.; Wong, M.H.; Bailey, K.W.; Sidwell, R.W. Activities of oseltamivir and ribavirin used alone and in combination against infections in mice with recent isolates of influenza a (h1n1) and b viruses. Antivir. Chem. Chemother. 2006, 17, 185–192. [Google Scholar] [CrossRef]
- Julander, J.G.; Kesler, K.; Van Wettere, A.J.; Morrey, J.D.; Smee, D.F. The use of plethysmography in determining the severity of lung pathology in a mouse model of minimally lethal influenza virus infection. Antivir. Res. 2014, 108, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Yamaguchi, I. Effect of glycosylation and glucose trimming inhibitors on the influenza a virus glycoproteins. J. Vet. Med. Sci. 2000, 62, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.T.; Hori, H.; Saul, R.; Sanford, B.A.; Molyneux, R.J.; Elbein, A.D. Castanospermine inhibits the processing of the oligosaccharide portion of the influenza viral hemagglutinin. Biochemistry 1983, 22, 3975–3984. [Google Scholar] [CrossRef] [PubMed]
- Karaivanova, V.K.; Luan, P.; Spiro, R.G. Processing of viral envelope glycoprotein by the endomannosidase pathway: Evaluation of host cell specificity. Glycobiology 1998, 8, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Datema, R.; Romero, P.A.; Rott, R.; Schwarz, R.T. On the role of oligosaccharide trimming in the maturation of sindbis and influenza virus. Arch. Virol. 1984, 81, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Romero, P.A.; Datema, R.; Schwarz, R.T. N-methyl-1-deoxynojirimycin, a novel inhibitor of glycoprotein processing, and its effect on fowl plague virus maturation. Virology 1983, 130, 238–242. [Google Scholar] [CrossRef]
- Hussain, S.; Miller, J.L.; Harvey, D.J.; Gu, Y.; Rosenthal, P.B.; Zitzmann, N.; McCauley, J.W. Strain-specific antiviral activity of iminosugars against human influenza a viruses. J. Antimicrob. Chemother. 2015, 70, 136–152. [Google Scholar] [CrossRef] [PubMed]
- Nct02061358: Randomized, Double-Blind, Placebo-Controlled, Parallel Group, Single-Ascending Dose Study to Determine the Safety, Tolerability and Pharmacokinetics of uv-4b Solution Administered Orally in Healthy Subjects. Available online: https://clinicaltrials.gov/ct2/show/NCT02061358 (accessed on 11 August 2015).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Strain | MOI | UV-4B IC90 | Control | IC90 |
|---|---|---|---|---|
| A/California/07/09 (H1N1) | 10−2 CCID50/cell | >320 µM a,e | Oseltamivir | 0.36 µM |
| A/California/12/2012 (H1N1) | 10−4 CCID50/cell | 320 µM c | Ribavirin | <13 µM b |
| 10−3.5 CCID50/cell | 219 µM e | 4.7 µM | ||
| A/Victoria/3/75 (H3N2) | 10−4 CCID50/cell | 440 µM c | Ribavirin | <13 µM b |
| 10−3.5 CCID50/cell | 483 µM e | 5.4 µM | ||
| A/Texas/50/2012 (H3N2) | 10−5 CCID50/cell | 82 µM d | Ribavirin | <13 µM b |
| B/Brisbane/60/2008 | 10−2 CCID50/cell | 200 µM e | Oseltamivir | 3.0 µM |
| B/Florida/4/2006 | 10−2 CCID50/cell | 150 µM e | Oseltamivir | 3.4 µM |
| B/Massachusetts/2/2012 | 10−4 CCID50/cell | 209µM c | Ribavirin | <13 µMb |
| 10−3.5 CCID50/cell | 245 µM c | 4.2 µM | ||
| B/Malaysia/2506/2004 | 10−4 CCID50/cell | >500 µM a,c | Ribavirin | <13 µMb |
| 10−3.5 CCID50/cell | >500 µM a,e | 4.8 µM |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Warfield, K.L.; Barnard, D.L.; Enterlein, S.G.; Smee, D.F.; Khaliq, M.; Sampath, A.; Callahan, M.V.; Ramstedt, U.; Day, C.W. The Iminosugar UV-4 is a Broad Inhibitor of Influenza A and B Viruses ex Vivo and in Mice. Viruses 2016, 8, 71. https://doi.org/10.3390/v8030071
Warfield KL, Barnard DL, Enterlein SG, Smee DF, Khaliq M, Sampath A, Callahan MV, Ramstedt U, Day CW. The Iminosugar UV-4 is a Broad Inhibitor of Influenza A and B Viruses ex Vivo and in Mice. Viruses. 2016; 8(3):71. https://doi.org/10.3390/v8030071
Chicago/Turabian StyleWarfield, Kelly L., Dale L. Barnard, Sven G. Enterlein, Donald F. Smee, Mansoora Khaliq, Aruna Sampath, Michael V. Callahan, Urban Ramstedt, and Craig W. Day. 2016. "The Iminosugar UV-4 is a Broad Inhibitor of Influenza A and B Viruses ex Vivo and in Mice" Viruses 8, no. 3: 71. https://doi.org/10.3390/v8030071
APA StyleWarfield, K. L., Barnard, D. L., Enterlein, S. G., Smee, D. F., Khaliq, M., Sampath, A., Callahan, M. V., Ramstedt, U., & Day, C. W. (2016). The Iminosugar UV-4 is a Broad Inhibitor of Influenza A and B Viruses ex Vivo and in Mice. Viruses, 8(3), 71. https://doi.org/10.3390/v8030071