
Clinical Implications of Antiviral Resistance in Influenza

Abstract
:1. Introduction

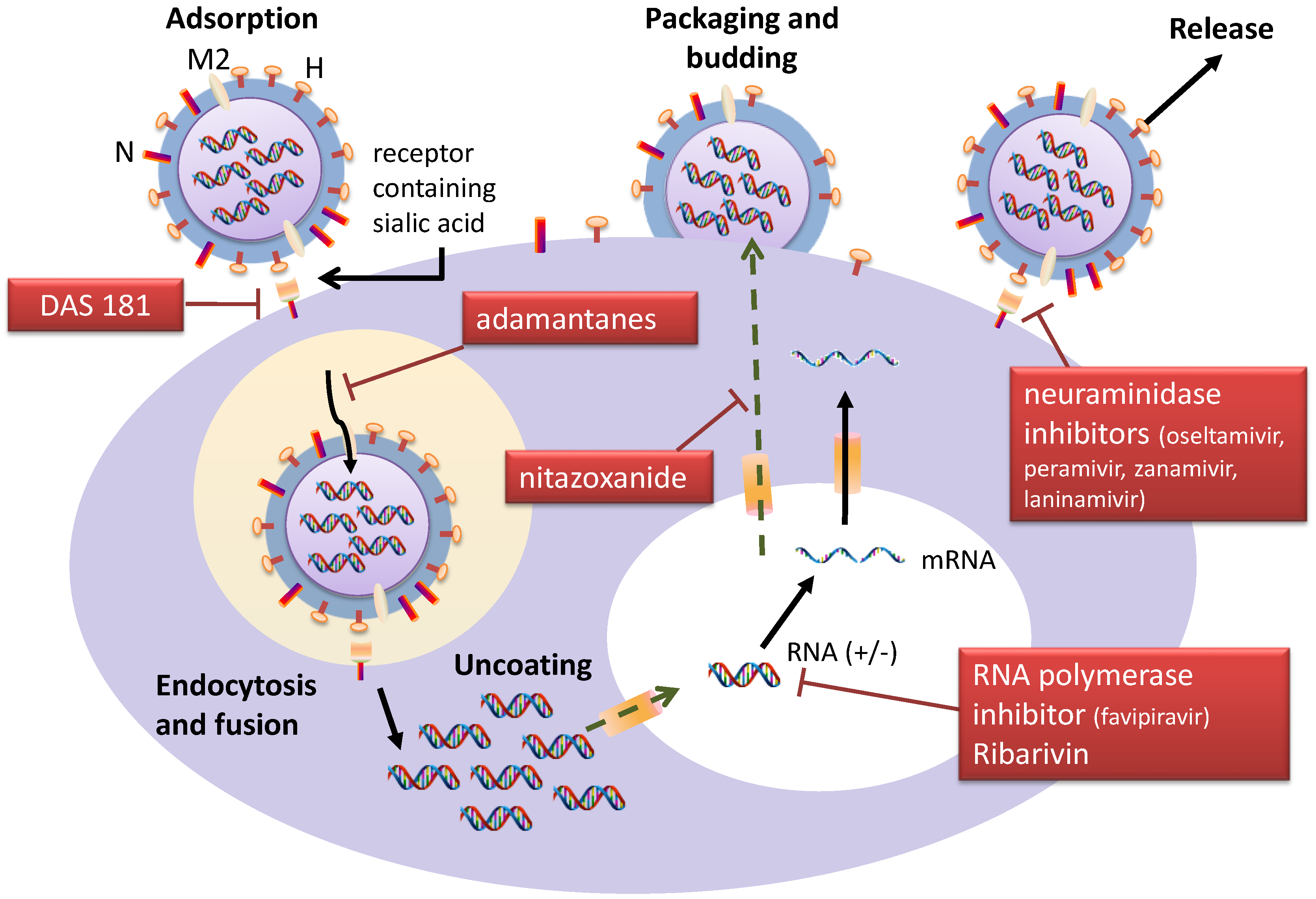{kind=link}
| Route of Administration | Availability | Indications | |
|---|---|---|---|
| Neuraminidase inhibitors | |||
| oseltamivir | oral | commercially available | Treatment of influenza A and B age ≥14 days (adults: dose reduction if CrCl <60 ml/min; children: tiered weight-based regimen if <40 kg) |
| Chemoprophylaxis age ≥1 year (with half-treatment dose) (a) | |||
| zanamivir | inhalational | commercially available | Treatment of influenza A and B age ≥7 years (contraindicated in underlying respiratory diseases, e.g., chronic obstructive airway disease (COPD), asthma) (b) |
| Chemoprophylaxis age ≥5 years (with half-treatment dose) | |||
| intravenous | limited (to a compassionate use program) | N.A. (under Phase III clinical trial) | |
| peramivir | intravenous | limited (USA, Japan, Korea, China) | Treatment of influenza A & B age ≥18 years (single dose application) (c) |
| laninamivir | inhalational | limited (only in Japan) | Treatment of influenza A and B adults and children (b) (single dose application) |
| Chemoprophylaxis age ≥10 years (with half-treatment dose) | |||
| M2-inhibitors | |||
| amantadine/rimantadine | oral | commercially available | Not recommended due to resistance in nearlyall circulating influenza A (H3N2, H1N1pdm09) and B virus strains |
| Polymerase inhibitor | |||
| Favipiravir | oral | limited (only in Japan) | Treatment of “novel or re-emerging” influenza virus infections age ≥18 years (under Phase III clinical trial) |
| Neuraminidase Inhibitors | Adamantanes | ||
|---|---|---|---|
| Oseltamivir | Zanamivir | Amantadine, Rimantadine | |
| A/H3N2 | <3% | rare | >99% |
| R292K, E119V (a) | S31N (b) | ||
| A/H1N1 (2007-08) | >99% | rare | rare |
| H275Y (c) | |||
| A/H1N1pdm09 | <3% | rare | >99% |
| H275Y (c) | S31N | ||
| B | rare | rare | 100% |
| I221V/T (d) | |||
| A/H5N1 | rare | rare | variable (b) |
| H275Y (c) | |||
| Oseltamivir | Zanamivir | Oseltamivir | |
| A/H7N9 | data limited | rare | >99% |
| R294K (e) | |||
2. Seasonal Influenza Viruses A/H3N2, A/H1N1, B
3. Pandemic Influenza Virus, A/H1N1pdm09
4. Avian Influenza Viruses, A/H5N1, A/H7N9
5. Detection and Clinical Management
| Key Elements |
|---|
1. Assessing for risk factors:
|
2. High-index of suspicion:
|
3. Testing for resistance:
|
4. Infection control:
|
5. Treatment:
|
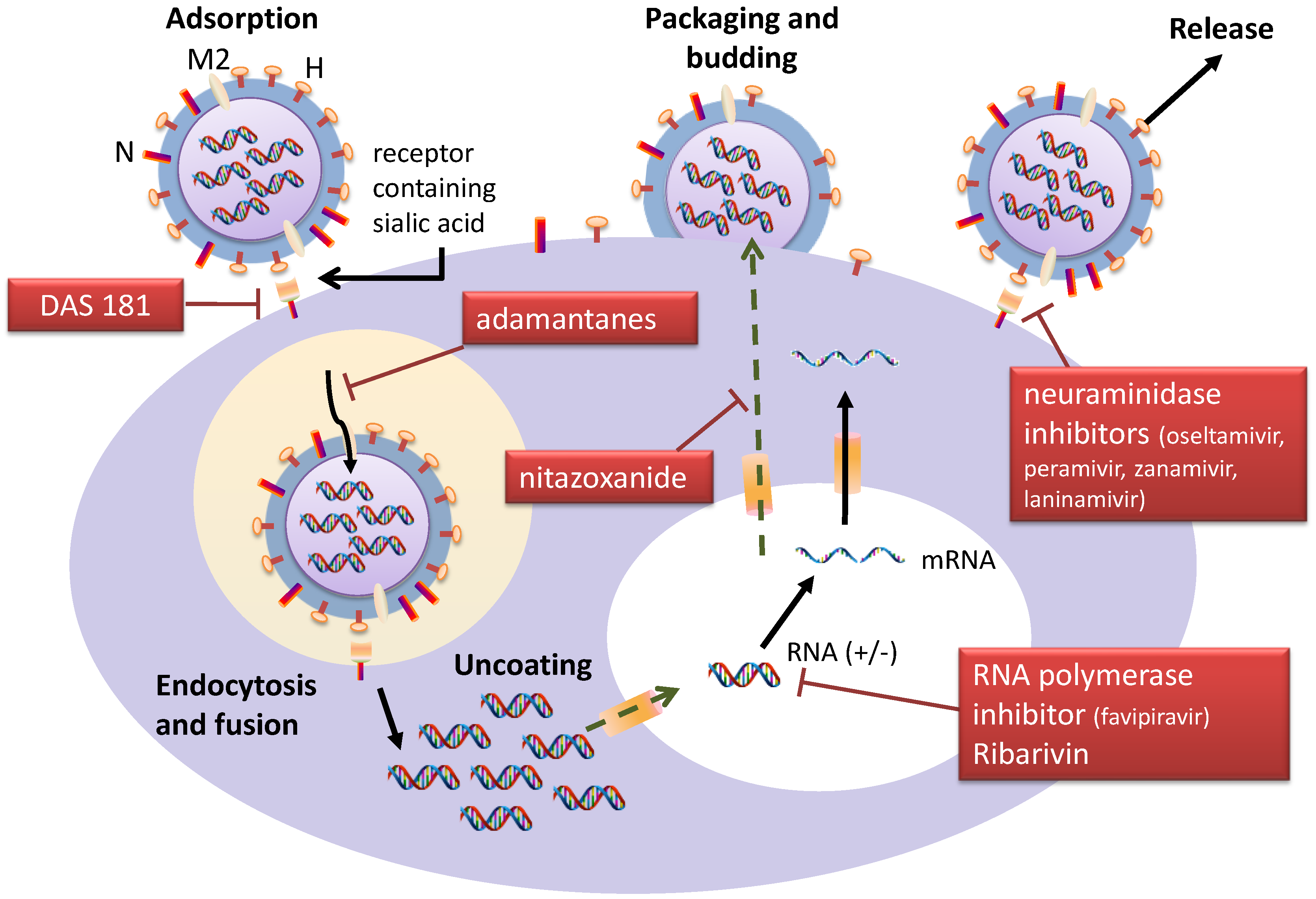
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lee, N.; Ison, M.G. Diagnosis, management and outcomes of adults hospitalized with influenza. Antivir. Ther. 2012, 17, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Choi, K.W.; Chan, P.K.; Hui, D.S.; Lui, G.C.; Wong, B.C.K.; Wong, R.Y.K.; Sin, W.Y.; Hui, W.M.; Ngai, K.L.K.; et al. Outcomes of adults hospitalised with severe influenza. Thorax 2010, 65, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Leo, Y.S.; Cao, B.; Chan, P.K.; Kyaw, W.M.; Uyeki, T.M.; Tam, W.W.S.; Cheung, C.S.K.; Yung, I.M.H.; Li, H.; et al. Neuraminidase inhibitors, superinfection and corticosteroids affect survival of influenza patients. Eur. Respir. J. 2015, 45, 1642–1652. [Google Scholar] [CrossRef] [PubMed]
- Dunning, J.; Baillie, J.K.; Cao, B.; Hayden, F.G. Antiviral combinations for severe influenza. Lancet Infect. Dis. 2014, 14, 1259–1270. [Google Scholar] [CrossRef]
- De Jong, M.D.; Ison, M.G.; Monto, A.S.; Metev, H.; Clark, C.; O’Neil, B.; Elder, J.; McCullough, A.; Collis, P.; Sheridan, W.P.; et al. Evaluation of intravenous peramivir for treatment of influenza in hospitalized patients. Clin. Infect. Dis. 2014, 59, e172–e285. [Google Scholar] [CrossRef] [PubMed]
- McKimm-Breschkin, J.L. Influenza neuraminidase inhibitors: Antiviral action and mechanisms of resistance. Influ. Other Respir. Viruses 2013, 7 (Suppl. 1), 25–36. [Google Scholar] [CrossRef] [PubMed]
- Takashita, E.; Meijer, A.; Lackenby, A.; Gubareva, L.; Rebelo-de-Andrade, H.; Besselaar, T.; Fry, A.; Gregory, V.; Leang, S.; Huang, W.; et al. Global update on the susceptibility of human influenza viruses to neuraminidase inhibitors, 2013–2014. Antivir. Res. 2015, 117, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Whitley, R.J.; Boucher, C.A.; Lina, B.; Nguyen-Van-Tam, J.S.; Osterhaus, A.; Schutten, M.; Monto, A.S. Global assessment of resistance to neuraminidase inhibitors, 2008–2011: The Influenza Resistance Information Study (IRIS). Clin. Infect. Dis. 2013, 56, 1197–1205. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, I.; Democratis, J.; Lackenby, A.; McNally, T.; Smith, J.; Pareek, M.; Ellis, J.; Bermingham, A.; Nicholson, K.; Zambon, M. Neuraminidase inhibitor resistance after oseltamivir treatment of acute influenza A and B in children. Clin. Infect. Dis. 2009, 48, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Ciancio, B.C.; Meerhoff, T.J.; Kramarz, P.; Bonmarin, I.; Borgen, K.; Boucher, C.A.; Buchholz, U.; Buda, S.; Dijkstra, F.; Dudman, S.; et al. Oseltamivir-resistant influenza A(H1N1) viruses detected in Europe during season 2007–2008 had epidemiologic and clinical characteristics similar to co-circulating susceptible A(H1N1) viruses. EuroSurveill 2009, 14, S1–S8. [Google Scholar]
- Dharan, N.J.; Gubareva, L.V.; Meyer, J.J.; Okomo-Adhiambo, M.; McClinton, R.C.; Marshall, S.A.; St George, K.; Epperson, S.; Brammer, L.; Klimov, A.I.; et al. Infections with oseltamivir-resistant influenza A(H1N1) virus in the United States. JAMA 2009, 301, 1034–1041. [Google Scholar] [CrossRef] [PubMed]
- Hurt, A.C. The epidemiology and spread of drug resistant human influenza viruses. Curr. Opin. Virol. 2014, 8, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Meijer, A.; Lackenby, A.; Hungnes, O.; Lina, B.; van der Werf, S.; Schweiger, B.; Opp, M.; Paget, J.; van-de-Kassteele, J.; Hay, A.; et al. Oseltamivir-resistant influenza virus A (H1N1), Europe, 2007–2008 season. Emerg. Infect. Dis. 2009, 15, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Baz, M.; Abed, Y.; Simon, P.; Hamelin, M.E.; Boivin, G. Effect of the neuraminidase mutation H274Y conferring resistance to oseltamivir on the replicative capacity and virulence of old and recent human influenza A(H1N1) viruses. J. Infect. Dis. 2010, 201, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Govorkova, E.A.; Bahl, J.; Zaraket, H.; Baranovich, T.; Seiler, P.; Prevost, K.; Webster, R.G.; Webby, R.J. Epistatic interactions between neuraminidase mutations facilitated the emergence of the oseltamivir-resistant H1N1 influenza viruses. Nat. Commun. 2014, 5, e5029. [Google Scholar] [CrossRef] [PubMed]
- Gooskens, J.; Jonges, M.; Claas, E.C.; Meijer, A.; Van den Broek, P.J.; Kroes, A.M. Morbidity and mortality associated with nosocomial transmission of oseltamivir-resistant influenza A(H1N1) virus. JAMA 2009, 301, 1042–1046. [Google Scholar] [CrossRef] [PubMed]
- Kelso, A.; Hurt, A.C. The ongoing battle against influenza: Drug-resistant influenza viruses: Why fitness matters. Nat. Med. 2012, 18, 1470–1471. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.C.; Lee, N.; Lui, G.C.; Ngai, K.K.; Wong, R.Y.; Choi, K.W.; Chan, P.K. Comparisons of oseltamivir-resistant (H275Y) and concurrent oseltamivir-susceptible seasonal influenza A(H1N1) virus infections in hospitalized adults, 2008–2009. Influ. Other Respir. Viruses 2013, 7, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Kawai, N.; Ikematsu, H.; Hirotsu, N.; Maeda, T.; Kawashima, T.; Tanaka, O.; Yamauchi, S.; Kawamura, K.; Matsuura, S.; Nishimura, M.; et al. Clinical effectiveness of oseltamivir and zanamivir for treatment of influenza A virus subtype H1N1 with the H274Y mutation: A Japanese, multicenter study of the 2007–2008 and 2008–2009 influenza seasons. Clin. Infect. Dis. 2009, 49, 1828–1835. [Google Scholar] [CrossRef] [PubMed]
- Kiso, M.; Mitamura, K.; Sakai-Tagawa, Y.; Shiraishi, K.; Kawakami, C.; Kimura, K.; Hayden, F.G.; Sugaya, N.; Kawaoka, Y. Resistant influenza A viruses in children treated with oseltamivir: Descriptive study. Lancet 2004, 364, 759–765. [Google Scholar] [CrossRef]
- Tamura, D.; Sugaya, N.; Ozawa, M.; Takano, R.; Ichikawa, M.; Yamazaki, M.; Kawakami, C.; Shimizu, H.; Uehara, R.; Kiso, M.; et al. Frequency of drug-resistant viruses and virus shedding in pediatric influenza patients treated with neuraminidase inhibitors. Clin. Infect. Dis. 2011, 52, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Eshaghi, A.; Shalhoub, S.; Rosenfeld, P.; Li, A.; Higgins, R.R.; Stogios, P.J.; Savchenko, A.; Bastien, N.; Li, Y.; Rotstein, C.; et al. Multiple influenza A (H3N2) mutations conferring resistance to neuraminidase inhibitors in a bone marrow transplant recipient. Antimicrob. Agents Chemother. 2014, 58, 7188–7197. [Google Scholar] [CrossRef] [PubMed]
- Gooskens, J.; Jonges, M.; Claas, E.C.; Meijer, A.; Kroes, A.C. Prolonged influenza virus infection during lymphocytopenia and frequent detection of drug-resistant viruses. J. Infect. Dis. 2009, 199, 1435–1441. [Google Scholar] [CrossRef] [PubMed]
- Hurt, A.C.; Leang, S.K.; Tiedemann, K.; Butler, J.; Mechinaud, F.; Kelso, A.; Downie, P.; Barr, I.G. Progressive emergence of an oseltamivir-resistant A(H3N2) virus over two courses of oseltamivir treatment in an immunocompromised paediatric patient. Influ. Other Respir. Viruses 2013, 7, 904–908. [Google Scholar] [CrossRef] [PubMed]
- Okomo-Adhiambo, M.; Demmler-Harrison, G.J.; Deyde, V.M.; Sheu, T.G.; Xu, X.; Klimov, A.I.; Gubareva, L.V. Detection of E119V and E119I mutations in influenza A (H3N2) viruses isolated from an immunocompromised patient: Challenges in diagnosis of oseltamivir resistance. Antimicrob. Agents Chemother. 2010, 54, 1834–1841. [Google Scholar] [CrossRef] [PubMed]
- Kawai, N.; Ikematsu, H.; Iwaki, N.; Maeda, T.; Kanazawa, H.; Kawashima, T.; Tanaka, O.; Yamauchi, S.; Kawamura, K.; Nagai, T.; et al. A comparison of the effectiveness of zanamivir and oseltamivir for the treatment of influenza A and B. J. Infect. 2008, 56, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Chan, P.K.; Hui, D.S.; Rainer, T.H.; Wong, E.; Choi, K.W.; Lui, G.C.; Wong, B.C.; Wong, R.Y.; Lam, W.Y.; et al. Viral loads and duration of viral shedding in adult patients hospitalized with influenza. J. Infect. Dis. 2009, 200, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Sugaya, N.; Mitamura, K.; Yamazaki, M.; Tamura, D.; Ichikawa, M.; Kimura, K.; Kawakami, C.; Kiso, M.; Ito, M.; Hatakeyama, S.; et al. Lower clinical effectiveness of oseltamivir against influenza B contrasted with influenza A infection in children. Clin. Infect. Dis. 2007, 44, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Hui, D.S.; Zuo, Z.; Ngai, K.L.; Lui, G.C.; Wo, S.K.; Tam, W.W.; Chan, M.C.; Wong, B.C.; Wong, R.Y.; et al. A prospective intervention study on higher-dose oseltamivir treatment in adults hospitalized with influenza A and B infections. Clin. Infect. Dis. 2013, 57, 1511–1519. [Google Scholar] [CrossRef] [PubMed]
- Ison, M.G.; Hui, D.S.; Clezy, K.; O’Neil, B.J.; Flynt, A.; Collis, P.J.; Simon, T.J.; Alexander, W.J. A clinical trial of intravenous peramivir compared with oral oseltamivir for the treatment of seasonal influenza in hospitalized adults. Antivir. Ther. 2013, 18, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.; Moore, Z.; Lee, N.; McKenna, J.; Bishop, A.; Fleischauer, A.; Springs, C.B.; Nguyen, H.T.; Sheu, T.G.; Sleeman, K.; et al. A cluster of patients infected with I221V influenza B virus variants with reduced oseltamivir susceptibility—North Carolina and South Carolina, 2010–2011. J. Infect. Dis. 2013, 207, 966–973. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, S.; Sugaya, N.; Ito, M.; Yamazaki, M.; Ichikawa, M.; Kimura, K.; Kiso, M.; Shimizu, H.; Kawakami, C.; Koike, K.; et al. Emergence of influenza B viruses with reduced sensitivity to neuraminidase inhibitors. JAMA 2007, 297, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Sleeman, K.; Huang, W.; Nguyen, H.T.; Levine, M.; Cheng, Y.; Li, X.; Tan, M.; Xing, X.; Xu, X.; et al. Neuraminidase inhibitor susceptibility testing of influenza type B viruses in China during 2010 and 2011 identifies viruses with reduced susceptibility to oseltamivir and zanamivir. Antivir. Res. 2013, 97, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Farrukee, R.; Leang, S.K.; Butler, J.; Lee, R.T.; Maurer-Stroh, S.; Tilmanis, D.; Sullivan, S.; Mosse, J.; Barr, I.G.; Hurt, A.C.; et al. Influenza viruses with B/Yamagata- and B/Victoria-like neuraminidases are differentially affected by mutations that alter antiviral susceptibility. J. Antimicrob. Chemother. 2015. [Google Scholar] [CrossRef] [PubMed]
- Leang, S.K.; Deng, Y.M.; Shaw, R.; Caldwell, N.; Iannello, P.; Komadina, N.; Buchy, P.; Chittaganpitch, M.; Dwyer, D.E.; Fagan, P.; et al. Influenza antiviral resistance in the Asia-Pacific region during 2011. Antivir. Res. 2013, 97, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Okomo-Adhiambo, M.; Fry, A.M.; Su, S.; Nguyen, H.T.; Elal, A.A.; Negron, E.; Hand, J.; Garten, R.J.; Barnes, J.; Xiyan, X.; et al. Oseltamivir-resistant influenza A(H1N1)pdm09 viruses, United States, 2013–2014. Emerg. Infect. Dis. 2015, 21, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Calatayud, L.; Lackenby, A.; Reynolds, A.; McMenamin, J.; Phin, N.F.; Zambon, M.; Pebody, R. Oseltamivir-resistant pandemic (H1N1) 2009 virus infection in England and Scotland, 2009–2010. Emerg. Infect. Dis. 2011, 17, 1807–1815. [Google Scholar] [CrossRef] [PubMed]
- Hurt, A.C.; Chotpitayasunondh, T.; Cox, N.J.; Daniels, R.; Fry, A.M.; Gubareva, L.V.; Hayden, F.G.; Hui, D.S.; Hungnes, O.; Lackenby, A.; et al. Antiviral resistance during the 2009 influenza A H1N1 pandemic: Public health, laboratory, and clinical perspectives. Lancet Infect. Dis. 2012, 12, 240–248. [Google Scholar] [CrossRef]
- Inoue, M.; Barkham, T.; Leo, Y.S.; Chan, K.P.; Chow, A.; Wong, C.W.; Lee, T.C.; Maurer-Stroh, S.; Lin, R.; Lin, C. Emergence of oseltamivir-resistant pandemic (H1N1) 2009 virus within 48 hours. Emerg. Infect. Dis. 2010, 16, 1633–1636. [Google Scholar] [CrossRef] [PubMed]
- Ghedin, E.; Holmes, E.C.; DePasse, J.V.; Pinilla, L.T.; Fitch, A.; Hamelin, M.E.; Papenburg, J.; Boivin, G. Presence of oseltamivir-resistant pandemic A/H1N1 minor variants before drug therapy with subsequent selection and transmission. J. Infect. Dis. 2012, 206, 1504–1511. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.; Galiano, M.; Lackenby, A.; Abdelrahman, T.; Barnes, R.; Evans, M.R.; Fegan, C.; Froude, S.; Hastings, M.; Knapper, S.; et al. Evidence of person-to-person transmission of oseltamivir-resistant pandemic influenza A(H1N1)2009 virus in a hematology unit. J. Infect. Dis. 2011, 203, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Tramontana, A.R.; George, B.; Hurt, A.C.; Doyle, J.S.; Langan, K.; Reid, A.B.; Harper, J.M.; Thursky, K.; Worth, L.J.; Dwyer, D.E.; et al. Oseltamivir resistance in adult oncology and hematology patients infected with pandemic (H1N1) 2009 virus, Australia. Emerg. Infect. Dis. 2010, 16, 1068–1075. [Google Scholar] [CrossRef] [PubMed]
- Baz, M.; Abed, Y.; Papenburg, J.; Bouhy, X.; Hamelin, M.E.; Boivin, G. Emergence of oseltamivir-resistant pandemic H1N1 virus during prophylaxis. N. Engl. J. Med. 2009, 361, 2296–2297. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Oseltamivir-resistant 2009 pandemic influenza A (H1N1) virus infection in two summer campers receiving prophylaxis—North Carolina, 2009. Morb. Mortal. Wkly. Rep. 2009, 58, 969–972. [Google Scholar]
- Moghadas, S.M.; Bowman, C.S.; Röst, G.; Fisman, D.N.; Wu, J. Post-exposure prophylaxis during pandemic outbreaks. BMC Med. 2009, 7, e73. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Meetings of the WHO working group on surveillance of influenza antiviral susceptibility—Geneva, November 2011 and June 2012. Wkly. Epidemiol. Rec. 2012, 87, 369–374. [Google Scholar]
- Storms, A.D.; Gubareva, L.V.; Su, S.; Wheeling, J.T.; Okomo-Adhiambo, M.; Pan, C.Y.; Reisdorf, E.; St George, K.; Myers, R.; Wotton, J.T.; et al. Oseltamivir-resistant pandemic (H1N1) 2009 virus infections, United States, 2010–2011. Emerg. Infect. Dis. 2012, 18, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Hurt, A.C.; Hardie, K.; Wilson, N.J.; Deng, Y.M.; Osbourn, M.; Gehrig, N.; Kelso, A. Community transmission of oseltamivir-resistant A(H1N1)pdm09 influenza. N. Engl. J. Med. 2011, 365, 2541–2542. [Google Scholar] [CrossRef] [PubMed]
- Takashita, E.; Ejima, M.; Itoh, R.; Miura, M.; Ohnishi, A.; Nishimura, H.; Odagiri, T.; Tashiro, M. A community cluster of influenza A(H1N1)pdm09 virus exhibiting cross-resistance to oseltamivir and peramivir in Japan, November to December 2013. Euro Surveill. 2014, 19, e20666. [Google Scholar] [CrossRef]
- Wang, B.; Taylor, J.; Ratnamohan, M.; McPhie, K.; Kesson, A.; Dixit, R.; Booy, R.; Hurt, A.; Saksena, N.; Dwyer, D.E. Frequency of oseltamivir resistance in Sydney, during the Newcastle outbreak of community transmitted oseltamivir-resistant influenza A(H1N1)pdm09 virus, Australia, June to August 2011. Euro Surveill. 2012, 17, 20210. [Google Scholar] [PubMed]
- Boivin, G. Detection and management of antiviral resistance for influenza viruses. Influ. Other Respir. Viruses 2013, 7 (Suppl. 3), 18–23. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.; Hooper, K.A.; Petrie, S.; Lee, R.; Maurer-Stroh, S.; Reh, L.; Guarnaccia, T.; Baas, C.; Xue, L.; Vitesnik, S.; et al. Estimating the fitness advantage conferred by permissive neuraminidase mutations in recent oseltamivir-resistant A(H1N1)pdm09 influenza viruses. PLoS Pathog. 2014, 10, e1004065. [Google Scholar] [CrossRef] [PubMed]
- Abed, Y.; Pizzorno, A.; Boivin, G. Therapeutic activity of intramuscular peramivir in mice infected with a recombinant influenza A/WSN/33 (H1N1) virus containing the H275Y neuraminidase mutation. Antimicrob. Agents Chemother. 2012, 56, 4375–4380. [Google Scholar] [CrossRef] [PubMed]
- Birnkrant, D.; Cox, E. The Emergency Use Authorization of peramivir fortreatment of 2009 H1N1 influenza. N. Engl. J. Med. 2009, 361, 2204–2207. [Google Scholar] [CrossRef] [PubMed]
- Renaud, C.; Pergam, S.A.; Polyak, C.; Jain, R.; Kuypers, J.; Englund, J.A.; Corey, L.; Boeckh, M.J. Early emergence of an H275Y mutation in a hematopoietic cell transplant recipient treated with intravenous peramivir. Transpl. Infect. Dis. 2010, 12, 513–517. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Trujillo, A.A.; Sheu, T.G.; Levine, M.; Mishin, V.P.; Shaw, M.; Ades, E.W.; Klimov, A.I.; Fry, A.M.; Gubareva, L.V. Analysis of influenza viruses from patients clinically suspected of infection with an oseltamivir resistant virus during the 2009 pandemic in the United States. Antivir. Res. 2012, 93, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Tamura, D.; DeBiasi, R.L.; Okomo-Adhiambo, M.; Mishin, V.P.; Campbell, A.P.; Loechelt, B.; Wiedermann, B.L.; Fry, A.M.; Gubareva, L.V. Emergence of multidrug resistant influenza A(H1N1)pdm09 virus variants in an immunocompromised child treated with oseltamivir and zanamivir. J. Infect. Dis. 2015. [Google Scholar] [CrossRef] [PubMed]
- Van der Vries, E.; Stelma, F.F.; Boucher, C.A. Emergence of a multidrug-resistant pandemic influenza A (H1N1) virus. N. Engl. J. Med. 2010, 363, 1381–1382. [Google Scholar] [CrossRef] [PubMed]
- Govorkova, E.A.; Baranovich, T.; Seiler, P.; Armstrong, J.; Burnham, A.; Guan, Y.; Peiris, M.; Webby, R.J.; Webster, R.G. Antiviral resistance among highly pathogenic influenza A (H5N1) viruses isolated worldwide in 2002–2012 shows need for continued monitoring. Antivir. Res. 2013, 98, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Nguyen, T.; Mishin, V.P.; Sleeman, K.; Balish, A.; Jones, J.; Creanga, A.; Marjuki, H.; Uyeki, T.M.; Nguyen, D.H.; et al. Antiviral susceptibility of highly pathogenic avian influenza A(H5N1) viruses isolated from poultry, Vietnam, 2009–2011. Emerg. Infect. Dis. 2013, 19, 1963–1971. [Google Scholar] [CrossRef] [PubMed]
- Adisasmito, W.; Chan, P.K.; Lee, N.; Oner, A.F.; Gasimov, V.; Aghayev, F.; Zaman, M.; Bamgboye, E.; Dogan, N.; Coker, R.; et al. Effectiveness of antiviral treatment in human influenza A(H5N1) infections: Analysis of a Global Patient Registry. J. Infect. Dis. 2010, 202, 1154–1160. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.K.; Lee, N.; Zaman, M.; Adisasmito, W.; Coker, R.; Hanshaoworakul, W.; Gasimov, V.; Oner, A.F.; Dogan, N.; Tsang, O.; et al. Determinants of antiviral effectiveness in influenza virus A subtype H5N1. J. Infect. Dis. 2012, 206, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- De Jong, M.D.; Tran, T.T.; Truong, H.K.; Vo, M.H.; Smith, G.J.; Nguyen, V.C. Oseltamivir resistance during treatment of influenza A (H5N1) infection. N. Engl. J. Med. 2005, 353, 2667–2672. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.N.; Lu, H.Z.; Cao, B.; Du, B.; Shang, H.; Gan, J.H.; Lu, S.H.; Yang, Y.D.; Fang, Q.; Shen, Y.Z.; et al. Clinical findings in 111 cases of influenza A (H7N9) virus infection. N. Engl. J. Med. 2013, 368, 2277–2285. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Lu, S.; Song, Z.; Wang, W.; Hao, P.; Li, J.; Zhang, X.; Yen, H.L.; Shi, B.; Li, T.; et al. Association between adverse clinical outcome in human disease caused by novel influenza A H7N9 virus and sustained viral shedding and emergence of antiviral resistance. Lancet 2013, 381, 2273–2279. [Google Scholar] [CrossRef]
- Sleeman, K.; Guo, Z.; Barnes, J.; Shaw, M.; Stevens, J.; Gubareva, L.V. R292K substitution and drug susceptibility of influenza A(H7N9) viruses. Emerg. Infect. Dis. 2013, 19, 1521–1524. [Google Scholar] [CrossRef] [PubMed]
- Marjuki, H.; Mishin, V.P.; Chesnokov, A.P.; De La Cruz, J.A.; Davis, C.T.; Villanueva, J.M.; Fry, A.M.; Gubareva, L.V. Neuraminidase Mutations Conferring Resistance to Oseltamivir in Influenza A(H7N9) Viruses. J. Virol. 2015, 89, 5419–5426. [Google Scholar] [CrossRef] [PubMed]
- Hai, R.; Schmolke, M.; Leyva-Grado, V.H.; Thangavel, R.R.; Margine, I.; Jaffe, E.L.; Krammer, F.; Solórzano, A.; García-Sastre, A.; Palese, P.; et al. Influenza A(H7N9) virus gains neuraminidase inhibitor resistance without loss of in vivo virulence or transmissibility. Nat. Commun. 2013, 4, 2854. [Google Scholar] [CrossRef] [PubMed]
- Yen, H.L.; Zhou, J.; Choy, K.T.; Sia, S.F.; Teng, O.; Ng, I.H.; Fang, V.J.; Hu, Y.; Wang, W.; Cowling, B.J.; et al. The R292K mutation that confers resistance to neuraminidase inhibitors leads to competitive fitness loss of A/Shanghai/1/2013 (H7N9) influenza virus in ferrets. J. Infect. Dis. 2014, 210, 1900–1908. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Fry, A.M.; Gubareva, L.V. Neuraminidase inhibitor resistance in influenza viruses and laboratory testing methods. Antivir. Ther. 2012, 17, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Pozo, F.; Lina, B.; Andrade, H.R.; Enouf, V.; Kossyvakis, A.; Broberg, E.; Daniels, R.; Lackenby, A.; Meijer, A.; Community Network of Reference Laboratories for Human Influenza in Europe. Guidance for clinical and public health laboratories testing for influenza virus antiviral drug susceptibility in Europe. J. Clin. Virol. 2013, 57, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Chan, P.K.; Wong, C.K.; Wong, K.T.; Choi, K.W.; Joynt, G.M.; Lam, P.; Chan, M.C.; Wong, B.C.; Lui, G.C.; et al. Viral clearance and inflammatory response patterns in adults hospitalized for pandemic 2009 influenza A(H1N1) virus pneumonia. Antivir. Ther. 2011, 16, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Sheu, T.G.; Mishin, V.P.; Klimov, A.I.; Gubareva, L.V. Assessment of pandemic and seasonal influenza A (H1N1) virus susceptibility to neuraminidase inhibitors in three enzyme activity inhibition assays. Antimicrob. Agents Chemother. 2010, 54, 3671–3677. [Google Scholar] [CrossRef] [PubMed]
- Ngai, K.L.; Lam, W.Y.; Lee, N.; Leung, T.F.; Hui, D.S.; Chan, P.K. Allele-specific conventional reverse-transcription polymerase chain reaction as a screening assay for discriminating influenza a H1N1 (H275Y) oseltamivir-resistant and wild-type viruses. J. Med. Virol. 2010, 82, 1295–1298. [Google Scholar] [CrossRef] [PubMed]
- Tamura, D.; Okomo-Adhiambo, M.; Mishin, V.P.; Guo, Z.; Xu, X.; Villanueva, J.; Fry, A.M.; Stevens, J.; Gubareva, L.V. Application of a seven-target pyrosequencing assay to improve the detection of neuraminidase inhibitor-resistant Influenza A(H3N2) viruses. Antimicrob. Agents Chemother. 2015, 59, 2374–2479. [Google Scholar] [CrossRef] [PubMed]
- Jonges, M.; Welkers, M.R.; Jeeninga, R.E.; Meijer, A.; Schneeberger, P.; Fouchier, R.A.; De Jong, M.D.; Koopmans, M. Emergence of the virulence-associated PB2 E627K substitution in a fatal human case of highly pathogenic avian influenza virus A(H7N7) infection as determined by Illumina ultra-deep sequencing. J. Virol. 2014, 88, 1694–1702. [Google Scholar] [CrossRef] [PubMed]
- Téllez-Sosa, J.; Rodríguez, M.H.; Gómez-Barreto, R.E.; Valdovinos-Torres, H.; Hidalgo, A.C.; Cruz-Hervert, P.; Luna, R.S.; Carrillo-Valenzo, E.; Ramos, C.; García-García, L.; et al. Using high-throughput sequencing to leverage surveillance of genetic diversity and oseltamivir resistance: A pilot study during the 2009 influenza A(H1N1) pandemic. PLoS ONE 2013, 8, e67010. [Google Scholar] [CrossRef] [PubMed]
- Fraaij, P.L.; van der Vries, E.; Beersma, M.F.; Riezebos-Brilman, A.; Niesters, H.G.; Van der Eijk, A.A.; de Jong, M.D.; Reis Miranda, D.; Horrevorts, A.M.; Ridwan, B.U.; et al. Evaluation of the antiviral response to zanamivir administered intravenously for treatment of critically ill patients with pandemic influenza A (H1N1) infection. J. Infect. Dis. 2011, 204, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Gaur, A.H.; Bagga, B.; Barman, S.; Hayden, R.; Lamptey, A.; Hoffman, J.M.; Bhojwani, D.; Flynn, P.M.; Tuomanen, E.; Webby, R. Intravenous zanamivir for oseltamivir-resistant 2009 H1N1 influenza. N. Engl. J. Med. 2010, 362, 88–89. [Google Scholar]
- Marty, F.M.; Man, C.Y.; van der Horst, C.; Francois, B.; Garot, D.; Mánez, R.; Thamlikitkul, V.; Lorente, J.A.; Alvarez-Lerma, F.; Brealey, D.; et al. Safety and pharmacokinetics of intravenous zanamivir treatment in hospitalized adults with influenza: An open-label, multicenter, single-arm, phase II study. J. Infect. Dis. 2014, 209, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, A.; Yates, P.J.; Murayama, M.; Soutome, T.; Furukawa, H. Evaluation of safety and efficacy of intravenous zanamivir in the treatment of hospitalized Japanese patients with influenza: An open-label, single-arm study. Antivir. Ther. 2014. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, A.; Chang, S.C.; Kim, M.J.; Chu, D.W.; Ohashi, Y.; MARVEL Study Group. Long-acting neuraminidase inhibitor laninamivir octanoate versus oseltamivir for treatment of influenza: A double-blind, randomized, noninferiority clinical trial. Clin. Infect. Dis. 2010, 51, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Kakuta, M.; Kubo, S.; Tanaka, M.; Tobiume, S.; Tomozawa, T.; Yamashita, M. Efficacy of a single intravenous administration of laninamivir (an active metabolite of laninamivir octanoate) in an influenza virus infection mouse model. Antivir. Res. 2013, 100, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Duval, X.; van der Werf, S.; Blanchon, T.; Mosnier, A.; Bouscambert-Duchamp, M.; Tibi, A.; Enouf, V.; Charlois-Ou, C.; Vincent, C.; Andreoletti, L.; et al. Efficacy of oseltamivir-zanamivir combination compared to each monotherapy for seasonal influenza: A randomized placebo-controlled trial. PLoS Med. 2010, 7, e1000362. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, J.T.; Hoopes, J.D.; Smee, D.F.; Prichard, M.N.; Driebe, E.M.; Engelthaler, D.M.; Le, M.H.; Keim, P.S.; Spence, R.P.; Went, G.T. Triple combination of oseltamivir , amantadine and ribavirin displays a synergistic activity against multiple influenza virus strains In vitro. Antimicrob. Agents Chemother. 2009, 53, 4115–4126. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.; Englund, J.A.; Nguyen, J.T.; Pukrittayakamee, S.; Lindegardh, N.; Tarning, J.; Tambyah, P.A.; Renaud, C.; Went, G.T.; de Jong, M.D.; et al. Combination therapy with amantadine, oseltamivir and ribavirin for influenza A infection: Safety and pharmacokinetics. Antivir. Ther. 2013, 18, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.Y.; Young Suh, G.; Huh, J.W.; Kim, S.H.; Kim, M.J.; Kim, Y.S.; Kim, H.R.; Ryu, Y.J.; Han, M.S.; Ko, Y.G.; et al. Triple-combination antiviral drug for pandemic H1N1 influenza virus infection in critically ill patients on mechanical ventilation. Antimicrob. Agents Chemother. 2011, 55, 5703–5709. [Google Scholar] [CrossRef] [PubMed]
- Furuta, Y.; Gowen, B.B.; Takahashi, K.; Shiraki, K.; Smee, D.F.; Barnard, D.L. Favipiravir (T-705), a novel viral RNA polymerase inhibitor. Antivir. Res. 2013, 100, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Kiso, M.; Takahashi, K.; Sakai-Tagawa, Y.; Shinya, K.; Sakabe, S.; Le, Q.M.; Ozawa, M.; Furuta, Y.; Kawaoka, Y. T-705 (favipiravir) activity against lethal H5N1 influenza A viruses. Proc. Natl. Acad. Sci. USA 2010, 107, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, J.I.; Lee, I.; Lee, S.; Hwang, M.W.; Bae, J.Y.; Heo, J.; Kim, D.; Jang, S.I.; Kim, H.; et al. Combination effects of peramivir and favipiravir against oseltamivir-resistant 2009 pandemic influenza A (H1N1) infection in mice. PLoS ONE 2014, 9, e101325. [Google Scholar] [CrossRef] [PubMed]
- Sleeman, K.; Mishin, V.P.; Deyde, V.M.; Furuta, Y.; Klimov, A.I.; Gubareva, L.V. In vitro antiviral activity of favipiravir (T-705) against drug-resistant influenza and 2009 A(H1N1) viruses. Antimicrob. Agents Chemother. 2010, 54, 2517–2524. [Google Scholar] [CrossRef] [PubMed]
- Smee, D.F.; Hurst, B.L.; Wong, M.H.; Bailey, K.W.; Tarbet, E.B.; Morrey, J.D.; Furuta, Y. Effects of the combination of favipiravir (T-705) and oseltamivir on influenza A virus infections in mice. Antimicrob. Agents Chemother. 2010, 54, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Tarbet, E.B.; Maekawa, M.; Furuta, Y.; Babu, Y.S.; Morrey, J.D.; Smee, D.F. Combinations of favipiravir and peramivir for the treatment of pandemic influenza A/California/04/2009 (H1N1) virus infections in mice. Antivir. Res. 2012, 94, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Moss, R.B.; Hansen, C.; Sanders, R.L.; Hawley, S.; Li, T.; Steigbigel, R.T. A phase II study of DAS181, a novel host directed antiviral for the treatment of influenza infection. J. Infect. Dis. 2012, 206, 1844–1851. [Google Scholar] [CrossRef] [PubMed]
- Marjuki, H.; Mishin, V.P.; Chesnokov, A.P.; De La Cruz, J.A.; Fry, A.M.; Villanueva, J.; Gubareva, L.V. An investigational antiviral drug, DAS181, effectively inhibits replication of zoonotic influenza A virus subtype H7N9 and protects mice from lethality. J. Infect. Dis. 2014, 210, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Chalkias, S.; Mackenzie, M.R.; Gay, C.; Dooley, C.; Marty, F.M.; Moss, R.B.; Li, T.; Routh, R.L.; Walsh, S.R.; Tan, C.S. DAS181 treatment of hematopoietic stem cell transplant patients with parainfluenza virus lung disease requiring mechanical ventilation. Transpl. Infect. Dis. 2014, 16, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, J.F. Nitazoxanide: A first-in-class broad-spectrum antiviral agent. Antivir. Res. 2014, 110, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Haffizulla, J.; Hartman, A.; Hoppers, M.; Resnick, H.; Samudrala, S.; Ginocchio, C.; Bardin, M.; Rossignol, J.F.; US Nitazoxanide Influenza Clinical Study Group. Effect of nitazoxanide in adults and adolescents with acute uncomplicated influenza: A double-blind, randomised, placebo-controlled, phase 2b/3 trial. Lancet Infect. Dis. 2014, 14, 609–618. [Google Scholar] [CrossRef]
- Belardo, G.; Cenciarelli, O.; La Frazia, S.; Rossignol, J.F.; Santoro, M.G. Synergistic effect of nitazoxanide with neuraminidase inhibitors against influenza A viruses In vitro. Antimicrob. Agents Chemother. 2015, 59, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Byrn, R.A.; Jones, S.M.; Bennett, H.B.; Bral, C.; Clark, M.P.; Jacobs, M.D.; Kwong, A.D.; Ledeboer, M.W.; Leeman, J.R.; McNeil, C.F.; et al. Preclinical activity of VX-787, a first-in-class, orally bioavailable inhibitor of the influenza virus polymerase PB2 subunit. Antimicrob. Agents Chemother. 2015, 59, 1569–1582. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, T.C.M.; Chan, M.C.W.; Lee, N. Clinical Implications of Antiviral Resistance in Influenza. Viruses 2015, 7, 4929-4944. https://doi.org/10.3390/v7092850
Li TCM, Chan MCW, Lee N. Clinical Implications of Antiviral Resistance in Influenza. Viruses. 2015; 7(9):4929-4944. https://doi.org/10.3390/v7092850
Chicago/Turabian StyleLi, Timothy C. M., Martin C. W. Chan, and Nelson Lee. 2015. "Clinical Implications of Antiviral Resistance in Influenza" Viruses 7, no. 9: 4929-4944. https://doi.org/10.3390/v7092850
APA StyleLi, T. C. M., Chan, M. C. W., & Lee, N. (2015). Clinical Implications of Antiviral Resistance in Influenza. Viruses, 7(9), 4929-4944. https://doi.org/10.3390/v7092850