
Apigenin Restricts FMDV Infection and Inhibits Viral IRES Driven Translational Activity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Compound and Antibodies
2.2. Cells and Viruses
2.3. Plasmids Construction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence 5'→3' |
|---|---|
| IRES-GFP-F | GGGAATTCCCACAACTGAGAAAACTCG |
| IRES-GFP-R | CCTCGAGCTACTTGTACAGCTCGTCC |
| Over-R | CGCCCTTGCTCACCATGGATTTAGT |
| Over-F | ATGGTGAGCAAGGGCGAGGAGCTGT |
| GFP-F | GCAGAAGAACGGCATCAAGG |
| GFP-R | GTGCTCAGGTAGTGGTTGTC |
| GAPDH-F | TCATGACCACAGTCCATGCC |
| GAPDH-R | GGATGACCTTGCCCACAGCC |
| 3D-F | GAACACATTCTTTACACCAGGAT |
| 3D-R | CATATCTTTGCCAATCAACATCAG |
2.4. Cytotoxicity Assay
2.6. Time-of-Addition Assay
2.7. Titration of Virus
2.8. Western Blotting Assay
2.9. Semi-Quantitative PCR and Quantitative Real-Time PCR
2.10. Statistics
3. Results
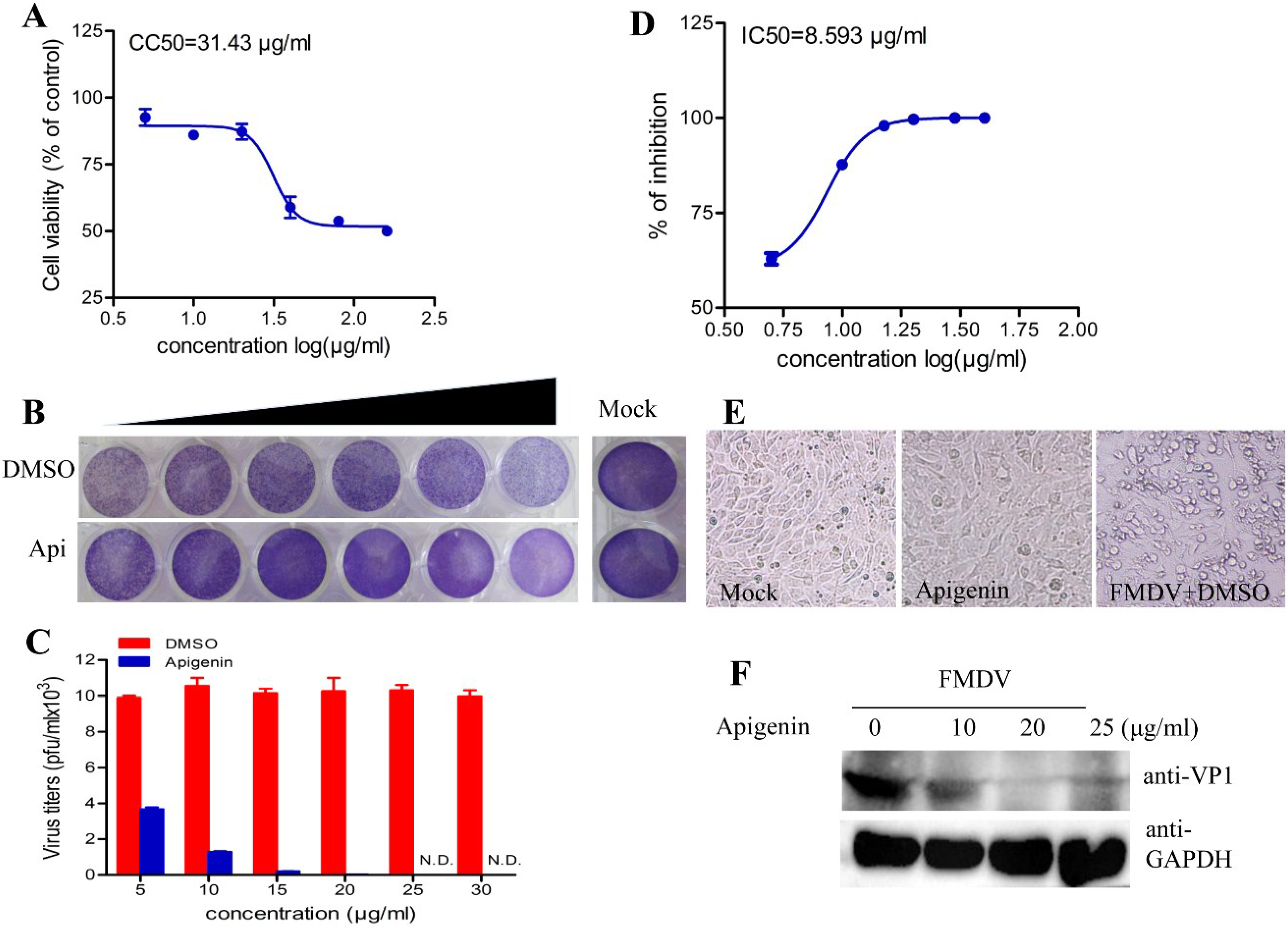
3.1. Apigenin Inhibits FMDV Infection
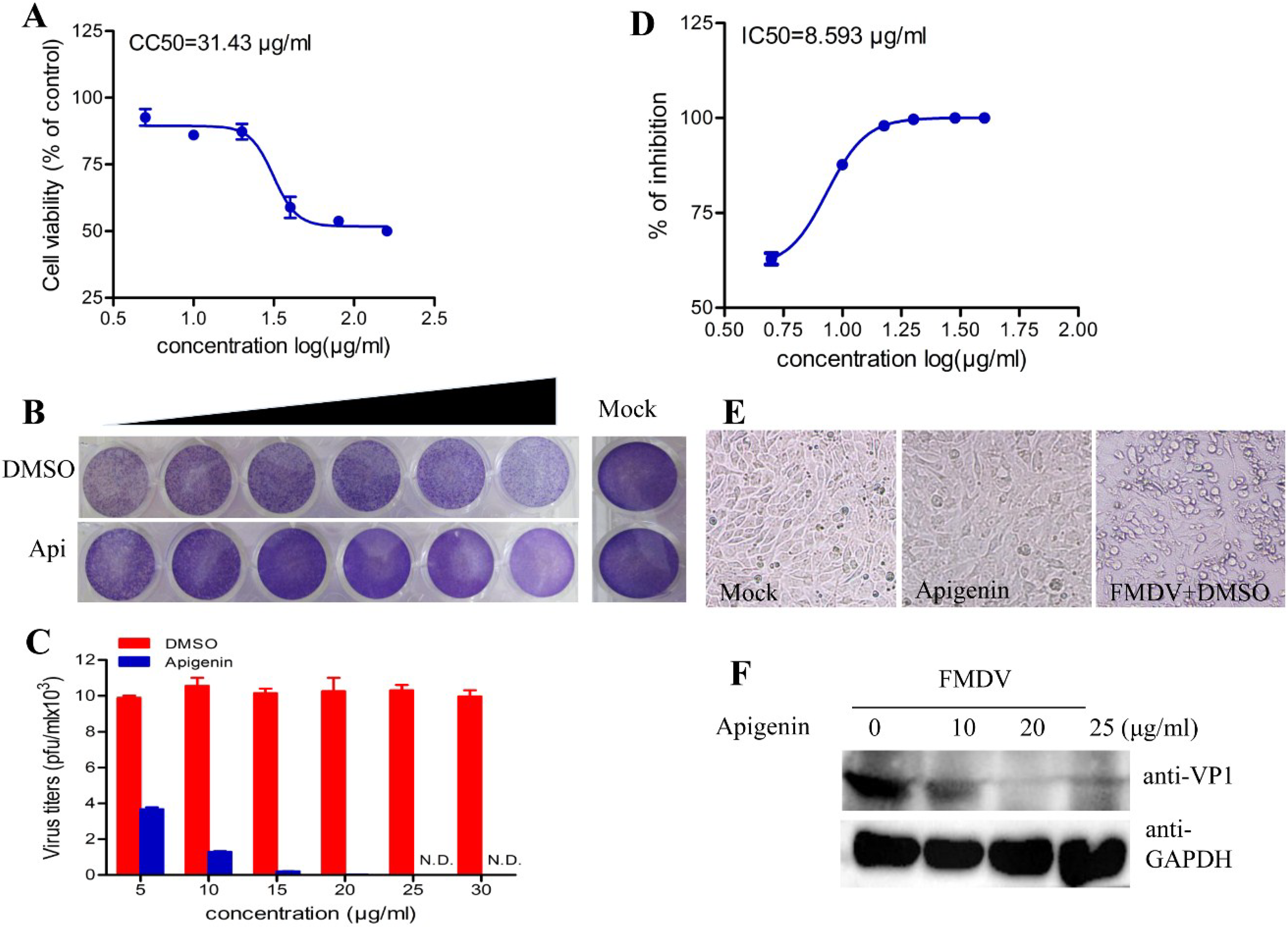
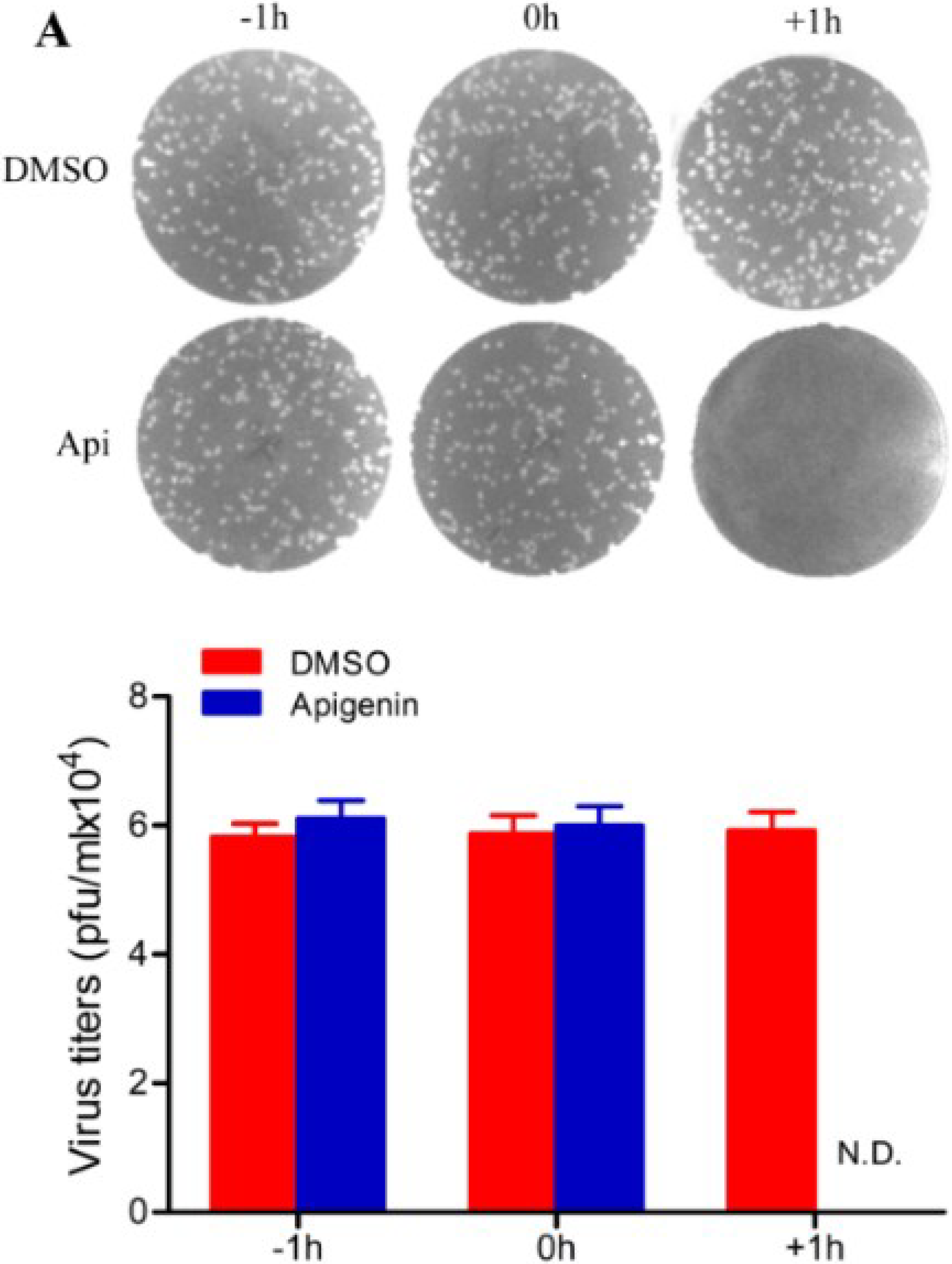
3.2. Apigenin Inhibits FMDV Infection during Post-Entry
3.3. Apigenin Suppresses FMDV IRES Activity
4. Discussion
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Grubman, M.J.; Baxt, B. Foot-and-Mouth Disease. Clin. Microbiol. Rev. 2004, 17, 465–493. [Google Scholar] [PubMed]
- DiMarchi, R.; Brooke, G.; Gale, C.; Cracknell, V.; Doel, T.; Mowat, N. Protection of cattle against foot-and-mouth disease by a synthetic peptide. Science 1986, 232, 639–641. [Google Scholar] [CrossRef] [PubMed]
- Domingo, E.; Escarmı, C.; Ruiz-jarabo, C.M.; Carrillo, E.; Nu, J.I.; Sobrino, F. Evolution of foot-and-mouth disease virus. Virus Res. 2003, 91, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Francis, M.J.; Hastings, G.Z.; Clarke, B.E.; Brown, A.L.; Beddell, C.R.; Rowlands, D.J. Neutralizing antibodies to all seven serotypes of foot-and-mouth disease virus elicited by synthetic peptides. Immunology 1990, 69, 171–176. [Google Scholar] [PubMed]
- Kuhn, R.; Luz, N.; Beck, E. Functional analysis of the internal translation initiation site of foot-and-mouth disease virus. J Virol. 1990, 64, 4625–4631. [Google Scholar]
- Belsham, G.J.; Brangwyn, J.K. A region of the 5’ noncoding region of foot-and-mouth disease virus RNA directs efficient internal initiation of protein synthesis within cells: Involvement with the role of L protease in translational control. J. Virol. 1990, 64, 5389–5395. [Google Scholar] [PubMed]
- De Quinto, S.L.; Lafuente, E.; Martínez-Salas, E. IRES interaction with translation initiation factors: Functional characterization of novel RNA contacts with eIF3, eIF4B, and eIF4GII. RNA 2001, 7, 1213–1226. [Google Scholar]
- De Quinto, S.L.; Martínez-Salas, E. Interaction of the eIF4G initiation factor with the aphthovirus IRES is essential for internal translation initiation in vivo. RNA 2000, 6, 1380–1392. [Google Scholar]
- Ochs, K.; Saleh, L.; Bassili, G.; Sonntag, V.H.; Zeller, A.; Niepmann, M. Interaction of Translation Initiation Factor eIF4B with the Poliovirus Internal Ribosome Entry Site. J. Virol. 2002, 76, 2113–2122. [Google Scholar] [CrossRef] [PubMed]
- Rust, C.; Ochs, K.; Niepmann, M. Translation Initiation Factor eIF4B Interacts with a Picornavirus Internal Ribosome Entry Site in both 48S and 80S Initiation Complexes Independently of Initiator AUG Location. J. Virol. 1999, 73, 7505–7514. [Google Scholar] [PubMed]
- Martínez-Salas, E.; Sáiz, J.C.; Dávila, M.; Belsham, G.J.; Domingo, E. A single nucleotide substitution in the internal ribosome entry site of foot-and-mouth disease virus leads to enhanced cap-independent translation in vivo. J. Virol. 1993, 67, 3748–3755. [Google Scholar] [PubMed]
- Golde, W.T.; Pacheco, J.M.; Duque, H.; Doel, T.; Penfold, B.; Ferman, G.S.; Gregg, D.R.; Rodriguez, L.L. Vaccination against foot-and-mouth disease virus confers complete clinical protection in 7 days and partial protection in 4 days: Use in emergency outbreak response. Vaccine 2005, 23, 5775–5782. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, J.M.; Brum, M.C.S.; Moraes, M.P.; Golde, W.T.; Grubman, M.J. Rapid protection of cattle from direct challenge with foot-and-mouth disease virus (FMDV) by a single inoculation with an adenovirus-vectored FMDV subunit vaccine. Virology 2005, 337, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Moraes, M.; Mayr, G.; Mason, P.; Grubman, M. Early protection against homologous challenge after a single dose of replication-defective human adenovirus type 5 expressing capsid proteins of foot-and-mouth disease virus (FMDV) strain A. Vaccine 2002, 20, 1631–1639. [Google Scholar] [CrossRef] [PubMed]
- Randall, R.E.; Goodbourn, S. Interferons and viruses: an interplay between induction, signalling, antiviral responses and virus countermeasures. J. Gen. Virol. 2008, 89, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Schoggins, J.W.; Rice, C.M. Interferon-stimulated genes and their antiviral effector functions. Curr. Opin. Virol. 2011, 1, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Chinsangaram, J.; Piccone, M.E.; Grubman, M.J. Ability of Foot-and-Mouth Disease Virus to Form Plaques in Cell Culture Is Associated with Suppression of Alpha/Beta Interferon. J. Virol. 1999, 73, 9891–9898. [Google Scholar] [PubMed]
- Diaz-San Segundo, F.; Moraes, M.P.; de Los Santos, T.; Dias, C.C.A.; Grubman, M.J. Interferon-induced protection against foot-and-mouth disease virus infection correlates with enhanced tissue-specific innate immune cell infiltration and interferon-stimulated gene expression. J. Virol. 2010, 84, 2063–2077. [Google Scholar] [CrossRef] [PubMed]
- Chinsangaram, J.; Koster, M.; Grubman, M.J. Inhibition of L-Deleted Foot-and-Mouth Disease Virus Replication by Alpha/Beta Interferon Involves Double-Stranded RNA-Dependent Protein Kinase. J. Virol. 2001, 75, 5498–5503. [Google Scholar] [CrossRef] [PubMed]
- Inhibitor, T.P. Antiviral Effects of a Thiol Protease Inhibitor Foot-and-Mouth Disease Virus on. J. Virol. 1992, 66, 7168–7175. [Google Scholar] [PubMed]
- Guarné, A.; Hampoelz, B.; Glaser, W.; Carpena, X.; Tormo, J.; Fita, I.; Skern, T. Structural and biochemical features distinguish the foot-and-mouth disease virus leader proteinase from other papain-like enzymes. J. Mol. Biol. 2000, 302, 1227–1240. [Google Scholar] [CrossRef] [PubMed]
- Kronovetr, J.; Skern, T. Foot-and-mouth disease virus leader proteinase: A papain-like enzyme requiring an acidic environment in the active site. FEBS 2002, 528, 58–62. [Google Scholar] [CrossRef]
- Lefebvre, D.J.; de Vleeschauwer, A.R.; Goris, N.; van Borm, S.; Chimirri, A.; Monforte, A.M.; Valdazo-Gonzalez, B.; King, D.P.; Neyts, J.; de Clercq, K. A thiazepino[4,5-a]benzimidazole derivative hampers the RNA replication of Eurasian serotypes of foot-and-mouth disease virus. Biochem. Biophys. Res. Commun. 2014, 455, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.A.; Kasum, C.M. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Annu. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Gupta, S. Molecular targets for apigenin-induced cell cycle arrest and apoptosis in prostate cancer cell xenograft. Mol. Cancer Ther. 2006, 5, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Qiu, M.; Chen, D.; Zheng, N.; Jin, Y.; Wu, Z. Apigenin inhibits enterovirus 71 replication through suppressing viral IRES activity and modulating cellular JNK pathway. Antivir. Res. 2014, 109, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, G.; Gurley, E.C.; Zhou, H. Flavonoid apigenin inhibits lipopolysaccharide-induced inflammatory response through multiple mechanisms in macrophages. PLoS ONE 2014, 9, e107072. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Qiao, H.; Lv, Y.; Wang, J.; Chen, X.; Hou, Y.; Tan, R.; Li, E. Apigenin Inhibits Enterovirus-71 Infection by Disrupting Viral RNA Association with trans-Acting Factors. PLoS ONE 2014, 9, e110429. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Y.-T.; Xiao, L.; Zhu, L.; Wang, Q.; Yan, T. Anti-Inflammatory effects of apigenin in lipopolysaccharide-induced inflammatory in acute lung injury by suppressing COX-2 and NF-κB pathway. Inflammation 2014, 37, 2085–2090. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Qian, P.; Wu, Q.; Liu, S.; Fan, W.; Zhang, K.; Wang, R.; Zhang, H.; Chen, H.; Li, X. Swine interferon-induced transmembrane protein, sIFITM3, inhibits foot-and-mouth disease virus infection in vitro and in vivo. Antivir. Res. 2014, 109, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Li, B.; Mills, D.M.; Panchal, R.G.; Cardinale, S.C.; Butler, M.M.; Peet, N.P.; Majgier-Baranowska, H.; Williams, J.D.; Patel, I.; et al. Identification of a small-molecule entry inhibitor for filoviruses. J. Virol. 2011, 85, 3106–3119. [Google Scholar] [CrossRef] [PubMed]
- Johari, J.; Kianmehr, A.; Mustafa, M.R.; Abubakar, S.; Zandi, K. Antiviral activity of baicalein and quercetin against the Japanese encephalitis virus. Int. J. Mol. Sci. 2012, 13, 16785–16795. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Gaynor, R.B. Therapeutic potential of inhibition of the NF-kappaB pathway in the treatment of inflammation and cancer. J. Clin. Invest. 2001, 107, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.T.; Lamb, A.J. Recent advances in understanding the antibacterial properties of flavonoids. Int. J. Antimicrob. Agents 2011, 38, 99–107. [Google Scholar] [CrossRef]
- Friedman, M. Overview of antibacterial, antitoxin, antiviral, and antifungal activities of tea flavonoids and teas. Mol. Nutr. Food Res. 2007, 51, 116–134. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Schuier, M.; Sies, H.; Illek, B.; Fischer, H. Biochemical and Molecular Actions of Nutrients Transport across T84 Human Colon Epithelia. J. Nutr. 2005, 135, 2320–2325. [Google Scholar] [PubMed]
- Hour, M.J.; Huang, S.H.; Chang, C.Y.; Lin, Y.K.; Wang, C.Y.; Chang, Y.S.; Lin, C.W. Baicalein, ethyl acetate, and chloroform extracts of Scutellaria baicalensis inhibit the neuraminidase activity of pandemic 2009 H1N1 and seasonal influenza A viruses. Evid.-Based Complement. Altern. Med. 2013. [Google Scholar] [CrossRef]
- Lyu, S.-Y.; Rhim, J.-Y.; Park, W.-B. Antiherpetic activities of flavonoids against herpes simplex virus type 1 (HSV-1) and type 2 (HSV-2) in vitro. Arch. Pharm. Res. 2005, 28, 1293–1301. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, T.; Du, J.; Cui, S.; Yang, F.; Jin, Q. Anti-Enterovirus 71 Effects of Chrysin and Its Phosphate Ester. PLoS ONE 2014, 9, e89668. [Google Scholar] [CrossRef] [PubMed]
- Ahmed-belkacem, A.; Brillet, R.; Ahnou, N.; Hernandez, E.; Pallier, C.; Pawlotsky, J.; Navacelles, D. Inhibition of RNA binding to hepatitis C virus RNA-dependent RNA polymerase : A new mechanism for antiviral intervention. Nucleic Acids Res. 2014, 42, 9399–9409. [Google Scholar]
- Gravina, H.D.; Tafuri, N.F.; Silva Júnior, A.; Fietto, J.L.R.; Oliveira, T.T.; Diaz, M.A.N.; Almeida, M.R. In vitro assessment of the antiviral potential of trans-cinnamic acid, quercetin and morin against equid herpesvirus. Res. Vet. Sci. 2011, 91, e158–e162. [Google Scholar]
- Dou, J.; Chen, L.; Xu, G.; Zhang, L.; Zhou, H.; Wang, H.; Su, Z.; Ke, M.; Guo, Q.; Zhou, C. Effects of baicalein on Sendai virus in vivo are linked to serum baicalin and its inhibition of hemagglutinin-neuraminidase. Arch. Virol. 2011, 156, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wu, Z.; Du, J.; Hu, Y.; Liu, L.; Yang, F.; Jin, Q. Anti-Japanese-encephalitis-viral effects of kaempferol and daidzin and their RNA-binding characteristics. PLoS ONE 2012, 7, e30259. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, E.; Teoh, B.-T.; Sam, S.-S.; Lani, R.; Hassandarvish, P.; Chik, Z.; Yueh, A.; Abubakar, S.; Zandi, K. Baicalin, a metabolite of baicalein with antiviral activity against dengue virus. Sci. Rep. 2014, 4, 5452. [Google Scholar] [CrossRef] [PubMed]
- Shibata, C.; Ohno, M.; Otsuka, M.; Kishikawa, T.; Goto, K.; Muroyama, R.; Kato, N.; Yoshikawa, T.; Takata, A.; Koike, K. The flavonoid apigenin inhibits hepatitis C virus replication by decreasing mature microRNA122 levels. Virology 2014, 462–463, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Critchfield, J.W.; Butera, S.T.; Folks, T.M. Inhibition of HIV activation in latently infected cells by flavonoid compounds. AIDS Res. Hum. Retrovir. 1996, 12, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Kanerva, A.; Raki, M.; Ranki, T.; Särkioja, M.; Koponen, J.; Desmond, R.A.; Helin, A.; Stenman, U.-H.; Isoniemi, H.; Höckerstedt, K.; et al. Chlorpromazine and apigenin reduce adenovirus replication and decrease replication associated toxicity. J. Gene Med. 2007, 9, 3–9. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, S.; Fan, W.; Qian, P.; Zhang, D.; Wei, Y.; Chen, H.; Li, X. Apigenin Restricts FMDV Infection and Inhibits Viral IRES Driven Translational Activity. Viruses 2015, 7, 1613-1626. https://doi.org/10.3390/v7041613
Qian S, Fan W, Qian P, Zhang D, Wei Y, Chen H, Li X. Apigenin Restricts FMDV Infection and Inhibits Viral IRES Driven Translational Activity. Viruses. 2015; 7(4):1613-1626. https://doi.org/10.3390/v7041613
Chicago/Turabian StyleQian, Suhong, Wenchun Fan, Ping Qian, Dong Zhang, Yurong Wei, Huanchun Chen, and Xiangmin Li. 2015. "Apigenin Restricts FMDV Infection and Inhibits Viral IRES Driven Translational Activity" Viruses 7, no. 4: 1613-1626. https://doi.org/10.3390/v7041613
APA StyleQian, S., Fan, W., Qian, P., Zhang, D., Wei, Y., Chen, H., & Li, X. (2015). Apigenin Restricts FMDV Infection and Inhibits Viral IRES Driven Translational Activity. Viruses, 7(4), 1613-1626. https://doi.org/10.3390/v7041613