
Chondroitin Sulfate N-acetylgalactosaminyltransferase-2 Contributes to the Replication of Infectious Bursal Disease Virus via Interaction with the Capsid Protein VP2

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Plasmids

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences(5'-3') | Usage | Products Size |
|---|---|---|---|
| GtVP2-U | CCGAATTCATGACAAACCTGCAAGAT | Construction for pGtVP2 | 1335 bp |
| GtVP2-L | CCGGTACCTTATGCTCCTGCAATCTTCAG | ||
| Bait-VP2-U |  ATGACAAACCTGCAAGAT ATGACAAACCTGCAAGAT | Construction for pDONR221-GtVP2 | 1365 bp |
| Bait-VP2-L |  TTATGCTCCTGCAATCTTCAG TTATGCTCCTGCAATCTTCAG | ||
| Gal -U | CAGAATTCATGAGAATGCCCAGAAGAGGCT | Construction for pCSGalNAcT2 | 1635 bp |
| Gal -L | ACGGTACCATTAACCCACTGTTTCGCTGTT | ||
| ∆Gal -U | CAGAATTCATGCCACAGACAGATAGTAAT | Construction for p∆CSGalNAcT2 | 1524 bp |
| ∆Gal -L | ACGGTACCATTAACCCACTGTTTCGCTGTT | ||
| β-actin-U | CTGTGCCCATCTATGAAGGCTA | Quantification for β-actin | 138 bp |
| β-actin-L | ATTTCTCTCTCGGCTGTGGTG | ||
| Gal-F | AAGAGCCCCAAGGTCAATGG | Quantification for CSGalNAcT2 | 138 bp |
| Gal-R | GACGCCGTATTCACTAGGCA |
2.3. Virus Infection and Titration
2.4. Viral RNA Isolation and Real-Time RT-PCR Analysis
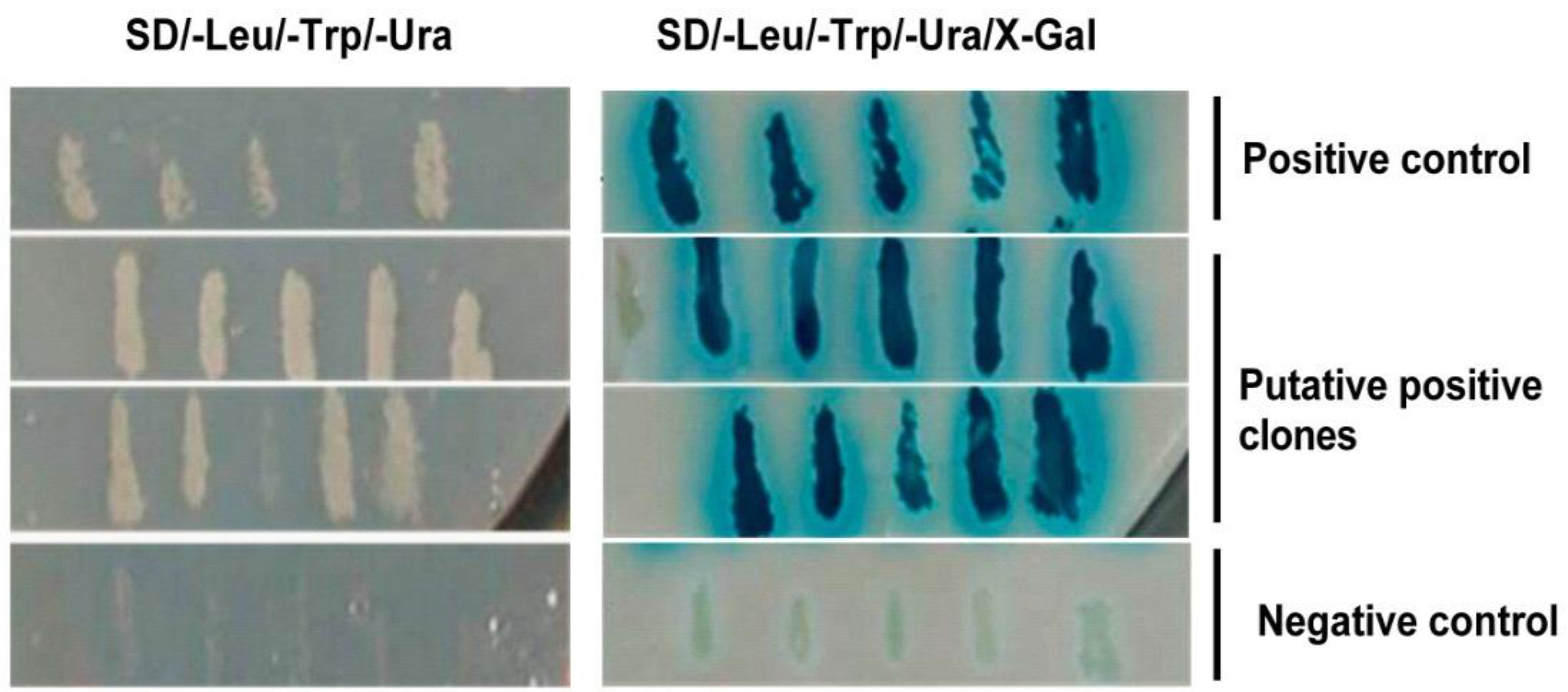
2.5. Yeast Two-Hybrid Screening
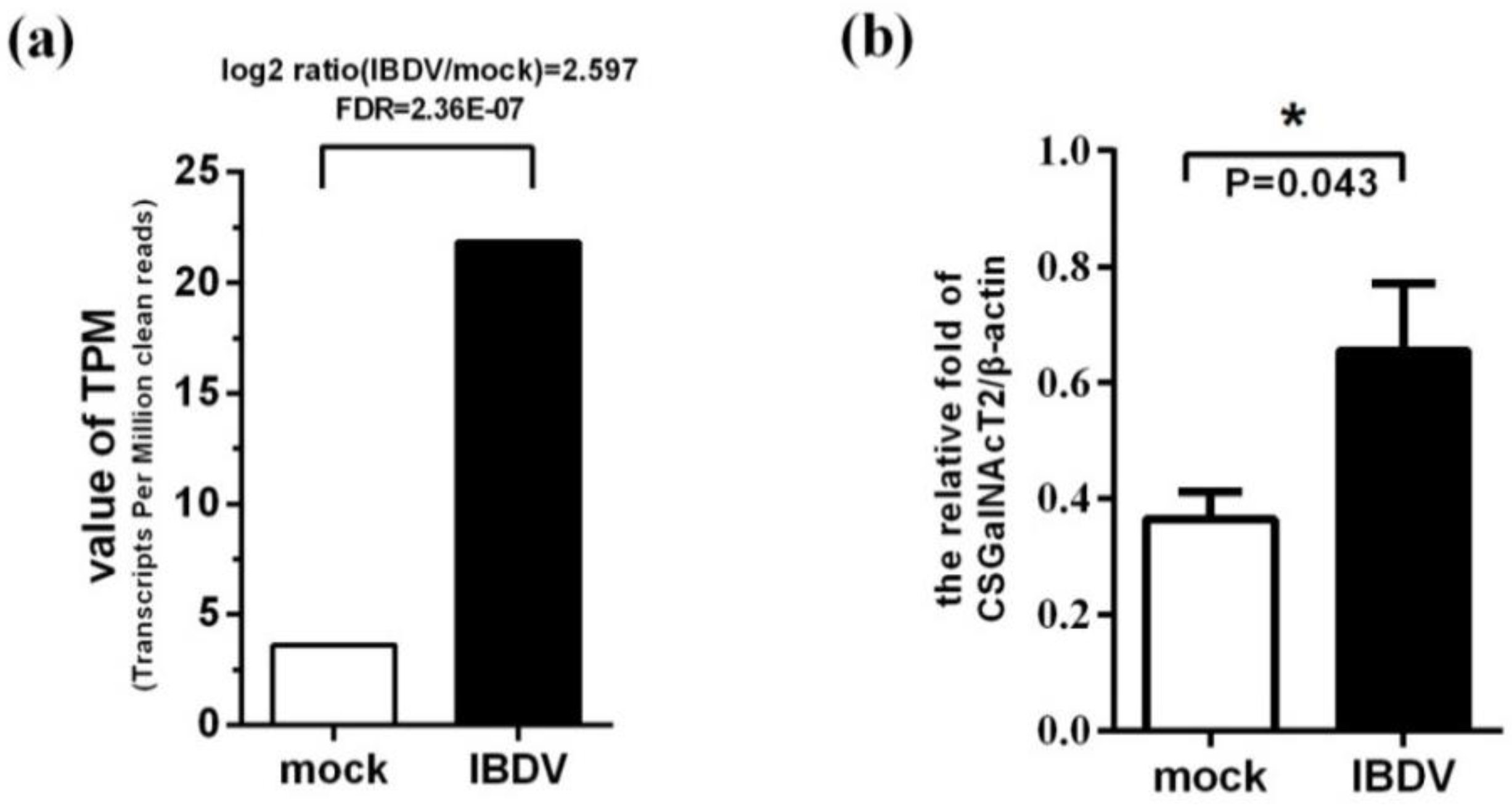
2.6. Analysis of Differentially Expressed Genes
2.7. Validation of CSGalNAcT2 Gene Expression
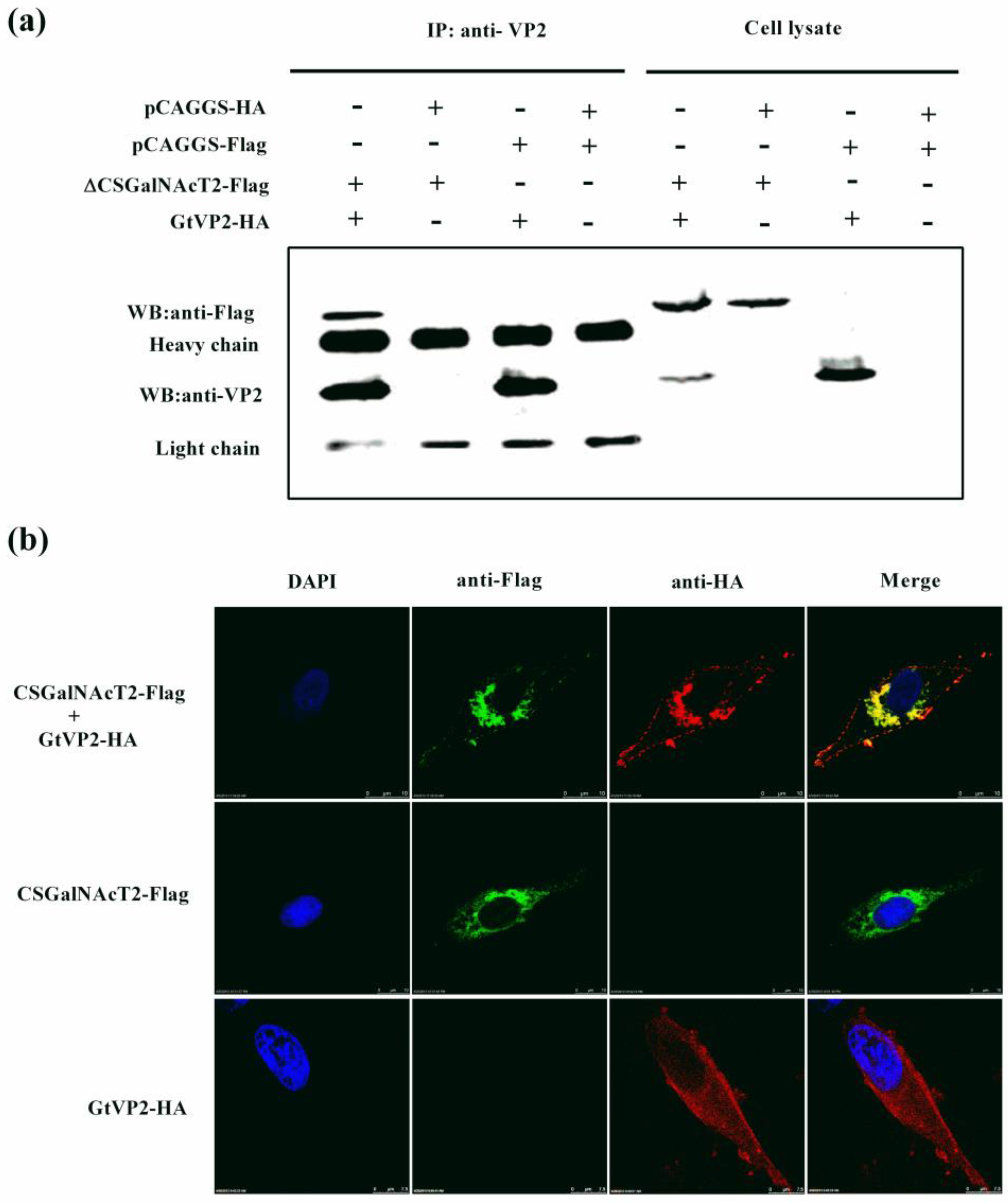
2.8. Co-Immunoprecipitation and Western Blot Analysis
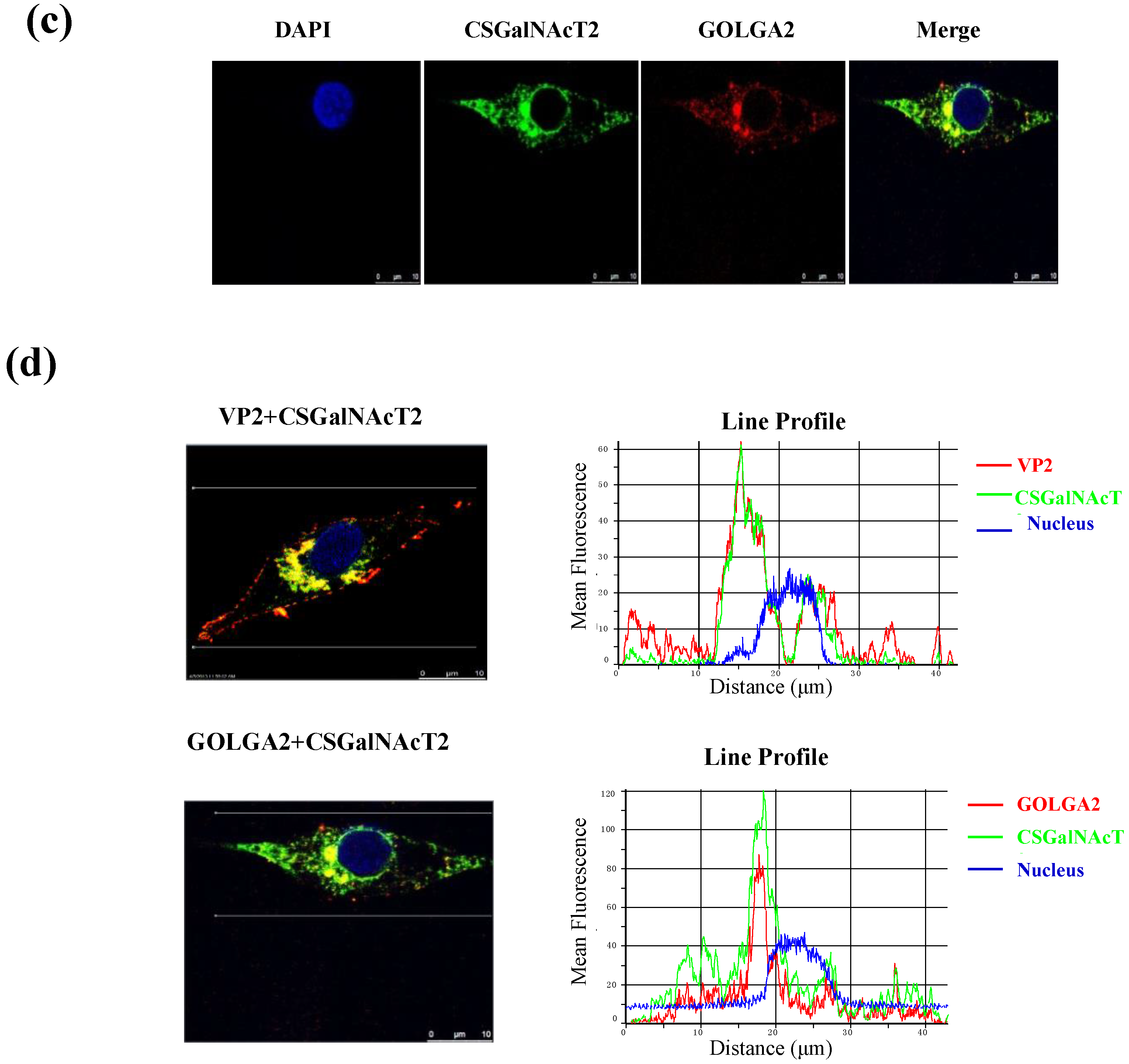
2.9. Confocal Laser Scanning Microscopy Assay
2.10. CSGalNAcT2 Overexpression Assay
2.11. RNA Interference of CSGalNAcT2
| Name | Sequences (5'-3') | Orientation | Position (bp) |
|---|---|---|---|
| 1# siRNA | CCAAAGAGCAAGCAUCCAATT | sense | 341–359 |
| UUGGAUGCUUGCUCUUUGGTT | antisense | 341–359 | |
| 2# siRNA | GCGAGGUCCUGAUGUUCUUTT | sense | 1070–1088 |
| AAGAACAUCAGGACCUCGCTT | antisense | 1070–1088 | |
| 3# siRNA | GCAUGUGUAUCCAGUCCAATT | sense | 1493–1511 |
| UUGGACUGGAUACACAUGCTT | antisense | 1493–1511 | |
| NC siRNA | UUCUCCGAACGUGUCACGUTT | sense | no |
| ACGUGACACGUUCGGAGAATT | antisense | no |
2.12. Brefeldin A Assay
2.13. Statistical Analysis
3. Results
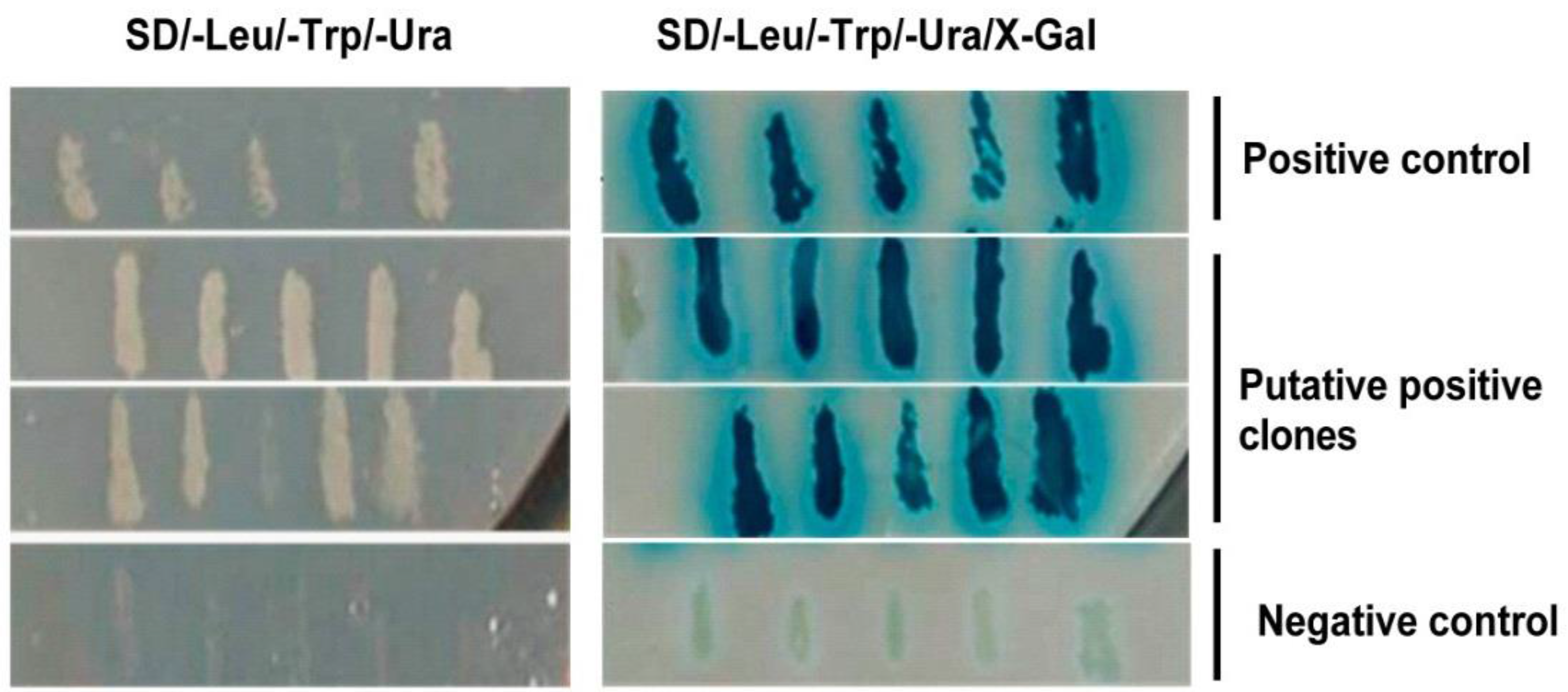
3.1. Host Cell Proteins Interact with VP2 of IBDV
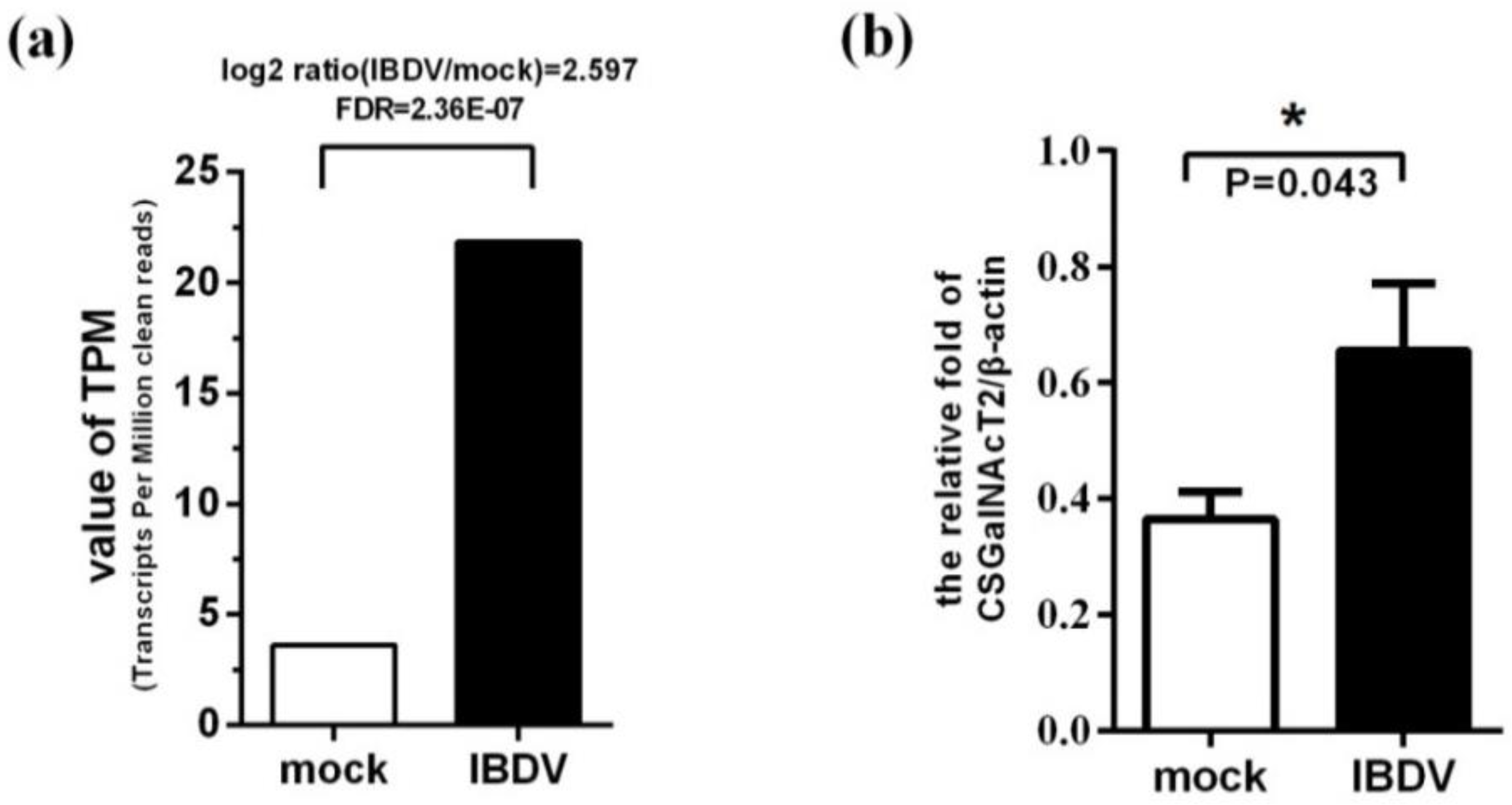
3.2. Expression of the Gene Encoding CSGalNAcT2 is Up-Regulated during IBDV Infection
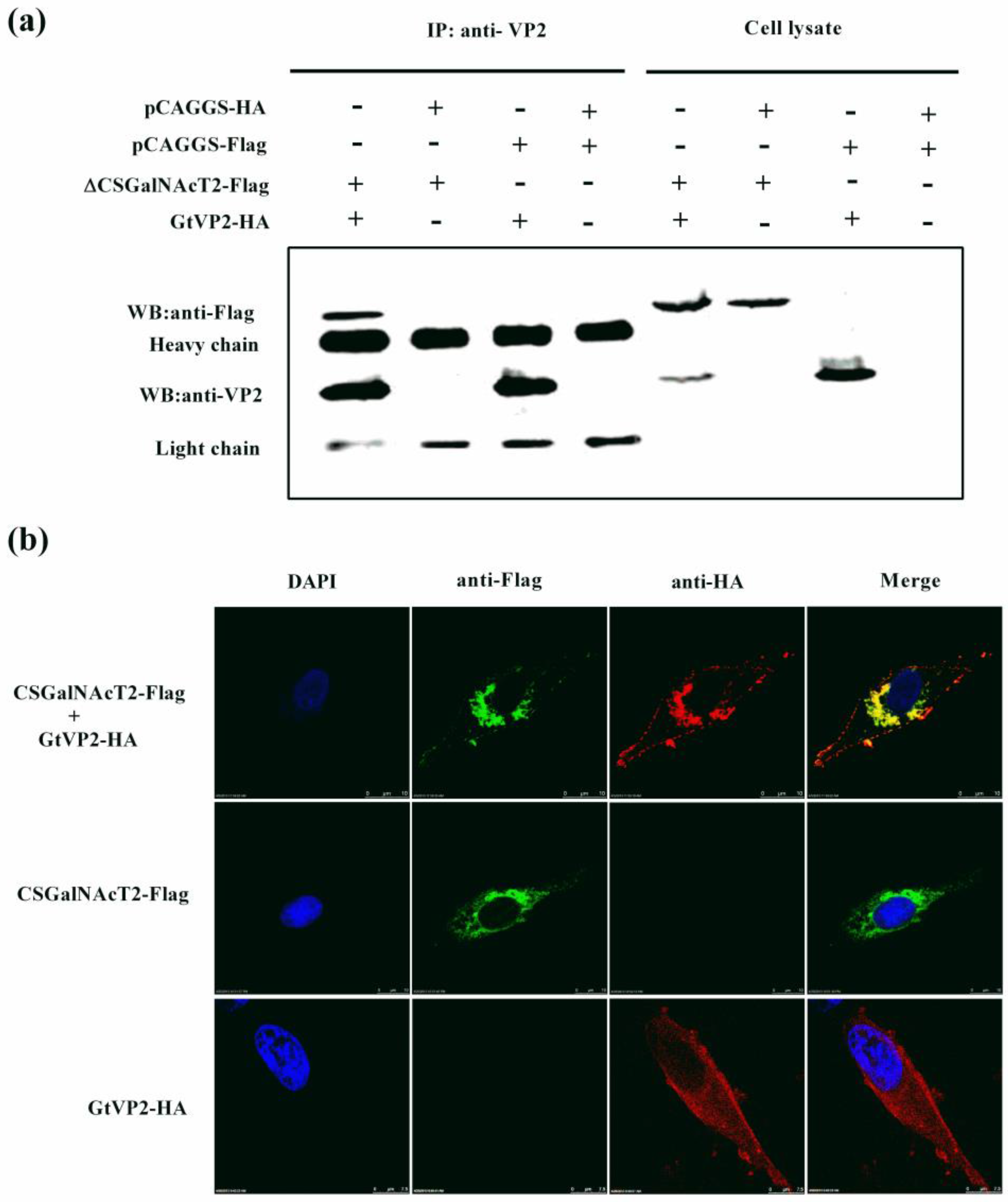
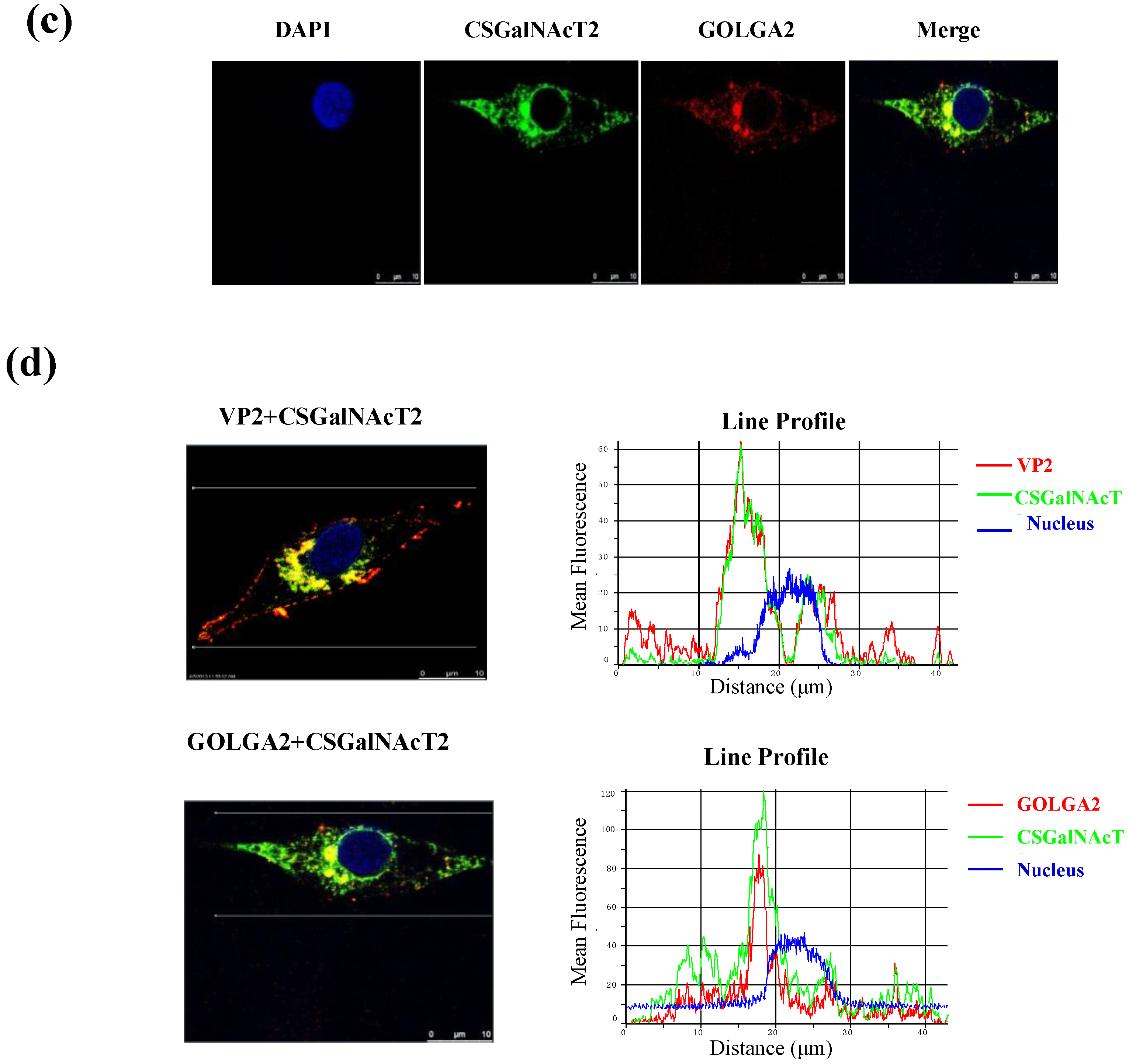
3.3. VP2 of IBDV Interacts with Host Cell Protein CSGalNAcT2 in the Golgi Apparatus
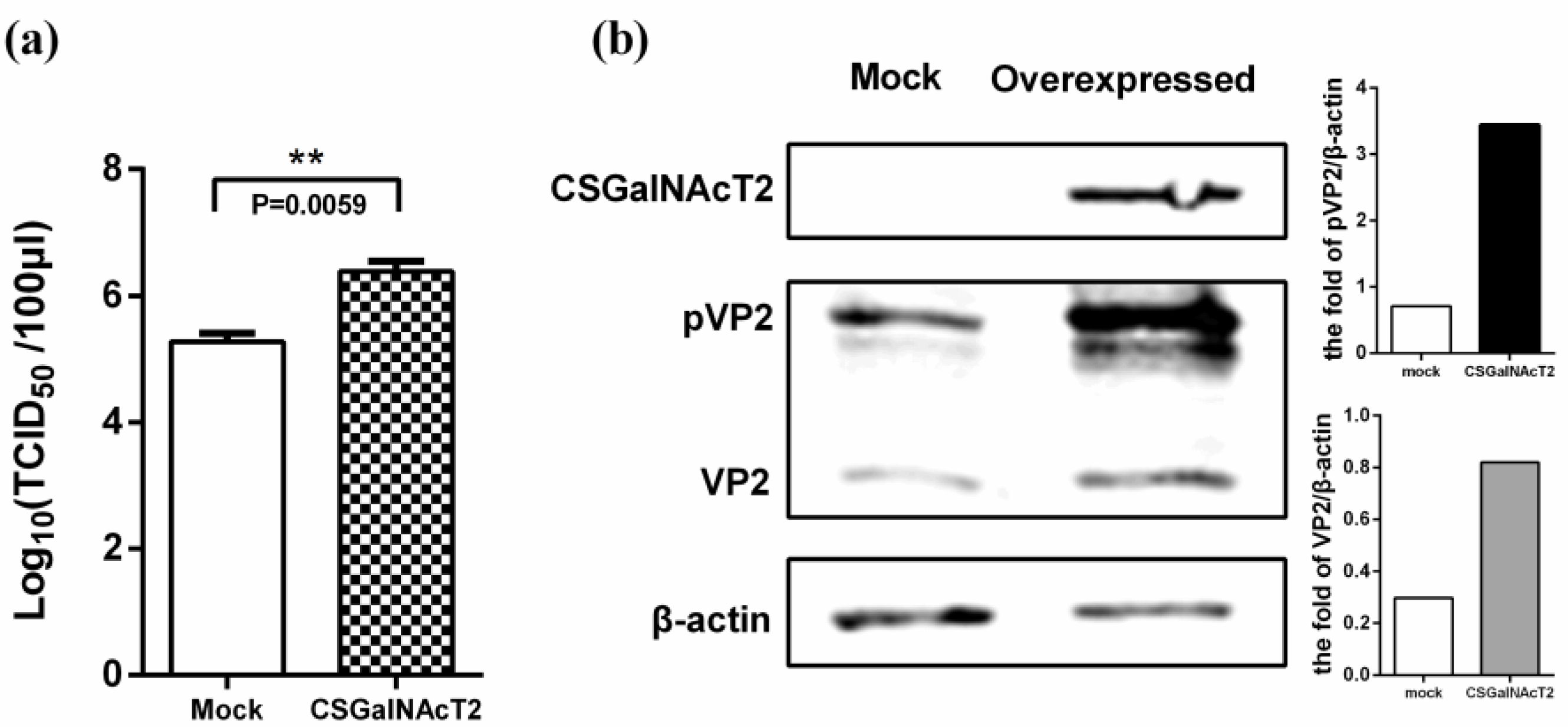
3.4. Overexpression of CSGalNAcT2 Promotes IBDV Replication
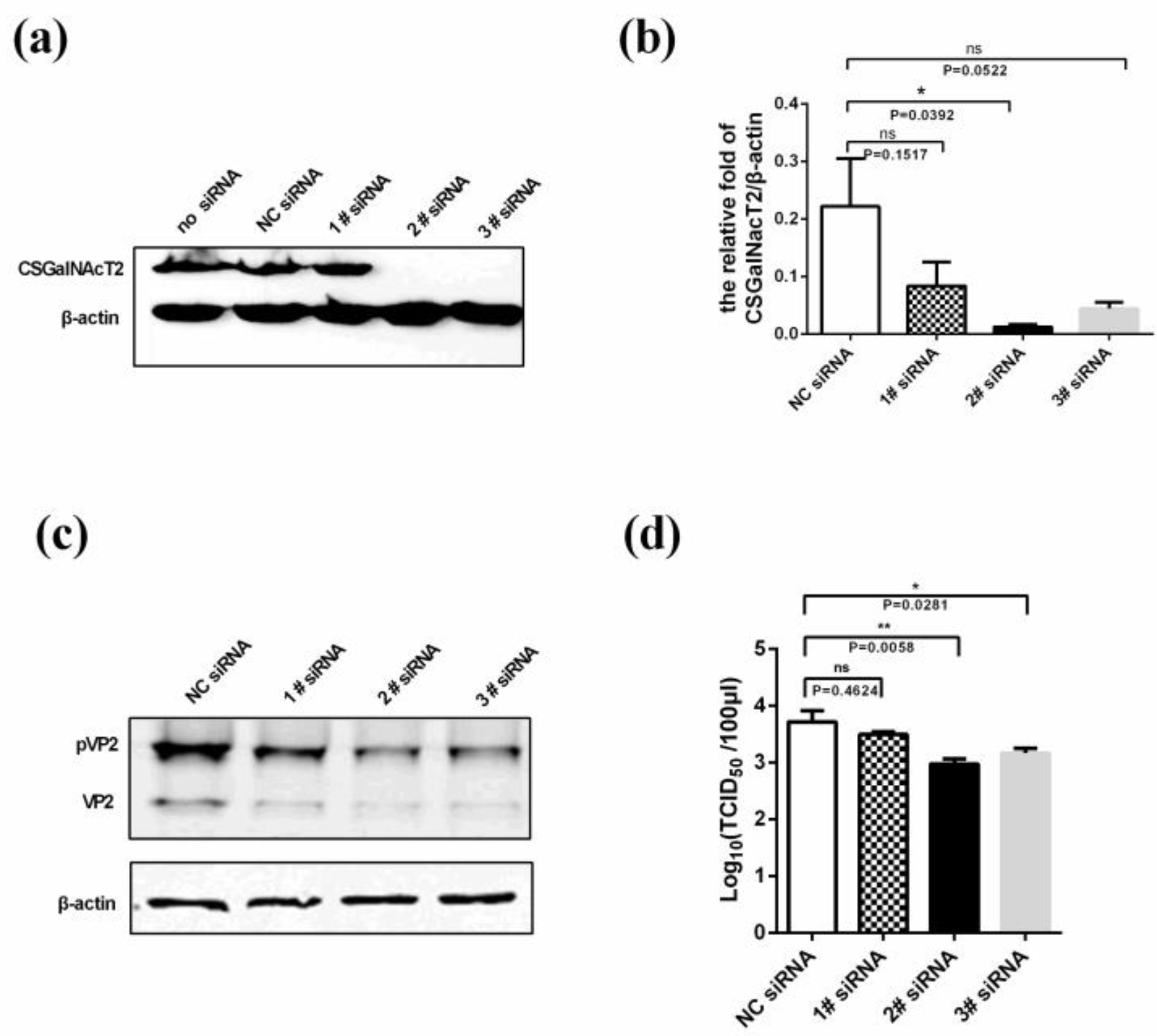
3.5. Knockdown of CSGalNAcT2 Inhibits IBDV Replication
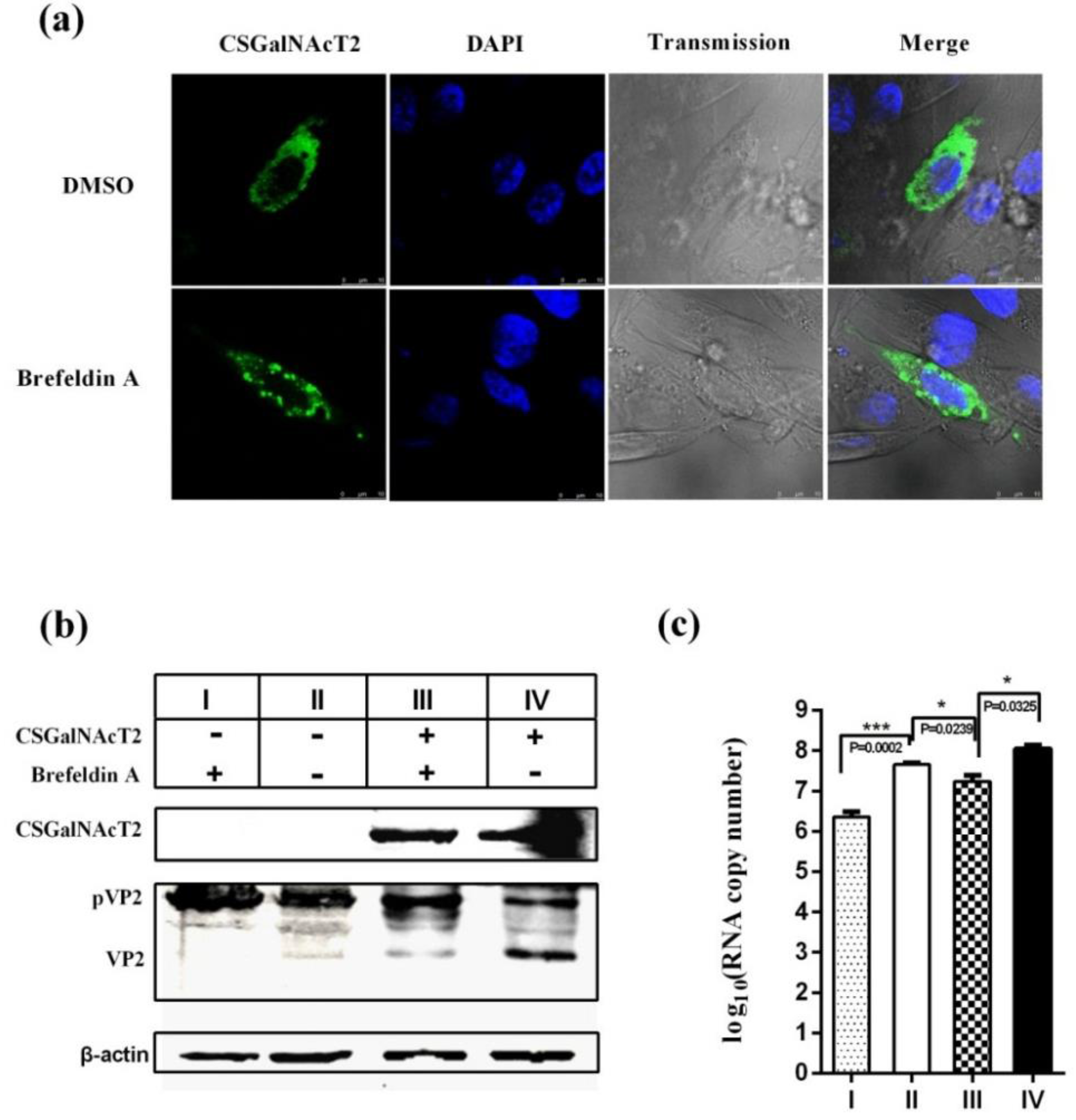
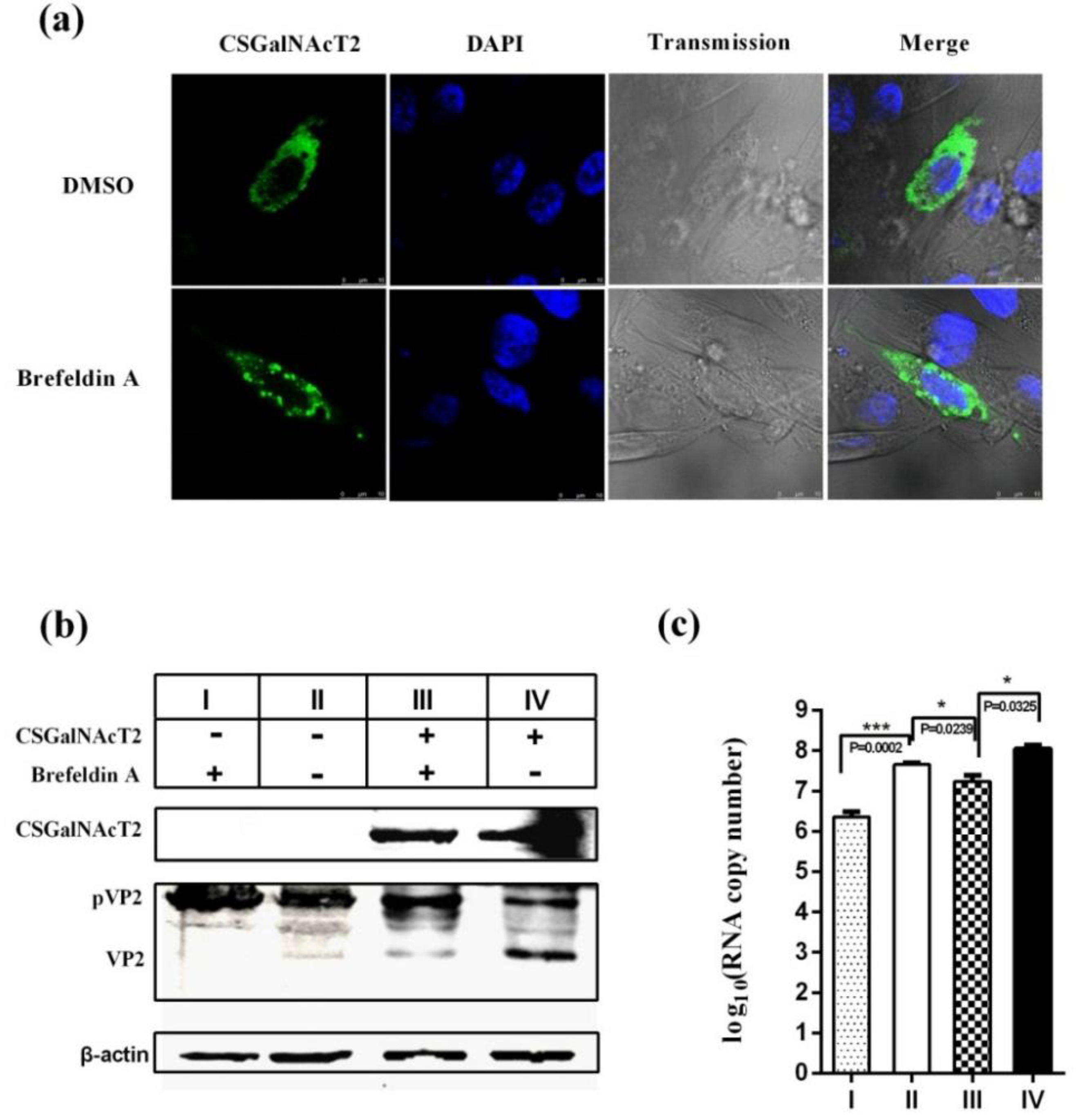
3.6. The Integrity of the Golgi Apparatus is Involved in the CSGalNAcT2 Effect on IBDV Replication
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cosgrove, A.S. An apparently new disease of chickens: Avian nephrosis. Avian Dis. 1962, 6, 385–389. [Google Scholar]
- Kibenge, F.S.; Dhillon, A.S.; Russell, R.G. Biochemistry and immunology of infectious bursal disease virus. J. Gen. Virol. 1988, 69, 1757–1775. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, M.M.; Kibenge, F.S. Infectious bursal disease virus: A review of molecular basis for variations in antigenicity and virulence. Can. J. Vet. Res. 1997, 61, 81–88. [Google Scholar] [PubMed]
- Sharma, J.M.; Kim, I.J.; Rautenschlein, S.; Yeh, H.Y. Infectious bursal disease virus of chickens: Pathogenesis and immunosuppression. Dev. Comp. Immunol. 2000, 24, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Müller, H.; Islam, M.R.; Raue, R. Research on infectious bursal disease—The past, the present and the future. Vet. Microbiol. 2003, 97, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Lasher, H.N.; Shane, S.M. Infectious bursal disease. World Poultry Sci. J. 1994, 50, 133–166. [Google Scholar] [CrossRef]
- Müller, H.; Scholtissek, C.; Becht, H. The genome of infectious bursal disease virus consists of two segments of double-stranded RNA. J. Virol. 1979, 31, 584–589. [Google Scholar] [PubMed]
- Dobos, P.; Hill, B.J.; Hallett, R.; Kells, D.T.; Becht, H.; Teninges, D. Biophysical and biochemical characterization of five animal viruses with bisegmented double-stranded RNA genomes. J. Virol. 1979, 32, 593–605. [Google Scholar] [PubMed]
- Sánchez, A.B.; Rodriguez, J.F. Proteolytic processing in infectious bursal disease virus: Identification of the polyprotein cleavage sites by site-directed mutagenesis. Virology 1999, 262, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Hudson, P.J.; McKern, N.M.; Power, B.E.; Azad, A.A. Genomic structure of the large RNA segment of infectious bursal disease virus. Nucleic Acids Res. 1986, 14, 5001–5012. [Google Scholar] [CrossRef] [PubMed]
- Jagadish, M.N.; Staton, V.J.; Hudson, P.J.; Azad, A.A. Birnavirus precursor polyprotein is processed in Escherichia coli by its own virus-encoded polypeptide. J. Virol. 1988, 62, 1084–1087. [Google Scholar] [PubMed]
- Da Costa, B.; Chevalier, C.; Henry, C.; Huet, J.C.; Petit, S.; Lepault, J.; Boot, H.; Delmas, B. The capsid of infectious bursal disease virus contains several small peptides arising from the maturation process of pVP2. J. Virol. 2002, 76, 2393–2402. [Google Scholar]
- Chevalier, C.; Lepault, J.; Erk, I.; Da Costa, B.; Delmas, B. The maturation process of pVP2requires assembly of infectious bursal disease virus capsids. J. Virol. 2002, 76, 2384–2392. [Google Scholar] [CrossRef] [PubMed]
- Mundt, E.; Köllner, B.; Kretzschmar, D. VP5 of infectious bursal disease virus is not essential for viral replication in cell culture. J. Virol. 1997, 71, 5647–5651. [Google Scholar] [PubMed]
- Mundt, E.; Beyer, J.; Muller, H. Identification of a novel viral protein in infectious bursal disease virus-infected cells. J. Gen. Virol. 1995, 76, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Saugar, I.; Luque, D.; Ona, A.; Rodriguez, J.F.; Carrascosa, J.L.; Trus, B.L.; Caston, J.R. Structural polymorphism of the major capsid protein of a double-stranded RNA virus: An amphipathic alpha helix as a molecular switch. Structure 2005, 13, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Coulibaly, F.; Chevalier, C.; Gutsche, I.; Pous, J.; Navaza, J.; Bressanelli, S.; Delmas, B.; Rey, F.A. The birnavirus crystal structure reveals structural relationships among icosahedral viruses. Cell 2005, 120, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Vakharia, V.N.; He, J.; Ahamed, B.; Snyder, D.B. Molecular basis of antigenic variation in infectious bursal disease virus. Virus Res. 1994, 31, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, D.J.; Sommer-Wagner, S.E. Amino acids contributing to antigenic drift in the infectious bursal disease birnavirus (IBDV). Virology 2011, 409, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Zhang, L.; Chen, Y.; Gao, L.; Wu, G.; Qin, L.; Wang, Y.; Ren, X.; Gao, Y.; Gao, H.; et al. Mutations of residues 249 and 256 in VP2 are involved in the replication and virulence of infectious bursal disease virus. PLoS One 2013, 8, e70982. [Google Scholar] [CrossRef] [PubMed]
- Brandt, M.; Yao, K.; Liu, M.; Heckert, R.A.; Vakharia, V.N. Molecular determinants of virulence, cell tropism, and pathogenic phenotype of infectious bursal disease virus. J. Virol. 2001, 75, 11974–11982. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.L.; Cao, Y.; Yu, T.; Mo, C.W. Adaptation of very virulent infectious bursal disease virus to chicken embryonic fibroblasts by site-directed mutagenesis of residues 279 and 284 of viral coat protein VP2. J. Virol. 1999, 73, 2854–2862. [Google Scholar] [PubMed]
- Qi, X.; Gao, H.; Gao, Y.; Qin, L.; Wang, Y.; Gao, L.; Wang, X. Naturally occurring mutations at residues 253 and 284 in VP2 contribute to the cell tropism and virulence of very virulent infectious bursal disease virus. Antivir. Res. 2009, 84, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, A.A.; de Haas, N.; Zeyda, I.; Mundt, E. Alteration of amino acids in VP2 of very virulent infectious bursal disease virus results in tissue culture adaptation and attenuation in chickens. J. Gen. Virol. 2002, 83, 121–129. [Google Scholar]
- Novoa, R.R.; Calderita, G.; Cabezas, P.; Elliott, R.M.; Risco, C. Key golgi factors for structural and functional maturation of bunyamwera virus. J. Virol. 2005, 79, 10852–10863. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, Y.; Li, X.; Li, X.; Cao, H.; Zheng, S.J. Critical roles of glucocorticoid-induced leucine zipper in infectious bursal disease virus (IBDV)-induced suppression of type I interferon expression and enhancement of IBDV growth in host cells via interaction with VP4. J. Virol. 2013, 87, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Hou, L.; Zhu, S.; Wang, J.; Zhou, J.; Liu, J. Infectious bursal disease virus activates the phosphatidylinositol 3-kinase (PI3K)/akt signaling pathway by interaction of VP5 protein with the p85α subunit of PI3K. Virology 2011, 417, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, Y.; Xue, Y.; Li, X.; Cao, H.; Zheng, S.J. Critical role for voltage-dependent anion channel 2 in infectious bursal disease virus-induced apoptosis in host cells via interaction with VP5. J. Virol. 2012, 86, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Zeng, X.W.; Gao, H.L.; Fu, C.Y.; Wei, P. Changes in VP2 gene during the attenuation of very virulent infectious bursal disease virus strain Gx isolated in china. Avian Dis. 2004, 48, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Niwa, H.; Yamamura, K.; Miyazaki, J. Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene 1991, 108, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Wang, Y.; Qi, X.; Gao, H.; Gao, Y.; Lin, H.; Song, X.; Pei, L.; Wang, X. Comparative study of the replication of infectious bursal disease virus in DF-1 cell line and chicken embryo fibroblasts evaluated by a new real-timeRT-PCR. J. Virol. Methods 2009, 157, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Audic, S.; Claverie, J.M. The significance of digital gene expression profiles. Genome Res. 1997, 7, 986–995. [Google Scholar] [PubMed]
- Cheng, Y.; Wang, X.; Gao, Y.; Gao, H.; Fu, C.; Jian, Z.; Qi, X.; Lu, G. Preparation and primary analysis of antigenic epitopes to monoclonal antibodies against Gt strain of chicken infectious bursal disease virus. Chin. J. Prevent. Vet. Med. 2006, 6, 687–700. [Google Scholar]
- Klausner, R.D. Brefeldin a: Insights into the control of membrane traffic and organelle structure. J. Cell Biol. 1992, 116, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Dinter, A.; Berger, E.G. Golgi-disturbing agents. Histochem. Cell Biol. 1998, 109, 571–590. [Google Scholar] [CrossRef] [PubMed]
- Duchez, S.; Pascal, V.; Cogné, N.; Jayat-Vignoles, C.; Julien, R.; Cogné, M. Glycotranscriptome study reveals an enzymatic switch modulating glycosaminoglycan synthesis during B-cell development and activation. Eur. J. Immunol. 2011, 41, 3632–3644. [Google Scholar] [CrossRef] [PubMed]
- Corse, E.; Machamer, C.E. Infectious bronchitis virus e protein is targeted to the Golgi complex and directs release of virus-like particles. J. Virol. 2000, 74, 4319–4326. [Google Scholar] [CrossRef] [PubMed]
- Bugarcic, A.; Taylor, J.A. Rotavirus nonstructural glycoprotein NSP4 is secreted from the apical surfaces of polarized epithelial cells. J. Virol. 2006, 80, 12343–12349. [Google Scholar] [CrossRef] [PubMed]
- Risco, C.; Carrascosa, J.L.; Frey, T.K. Structural maturation of rubella virus in the Golgi complex. Virology 2003, 312, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Fontana, J.; López-Montero, N.; Elliott, R.M.; Fernández, J.J.; Risco, C. The unique architecture of bunyamwera virus factories around theGolgi complex. Cell. Microbiol. 2008, 10, 2012–2028. [Google Scholar] [CrossRef] [PubMed]
- Weiland, E.; Weiland, F. Autoantibodies against Golgi apparatus induced by arteriviruses. Cell. Mol. Biol. (Noisy-le-Grand, France) 2002, 48, 279–284. [Google Scholar]
- Mayinger, P. Signaling at the Golgi. CSH Perspect. Biol. 2011, 3, a005314. [Google Scholar]
- Delgui, L.R.; Rodriguez, J.F.; Colombo, M.I. The endosomal pathway and the Golgi complex are involved in the infectious bursal disease virus life cycle. J. Virol. 2013, 87, 8993–9007. [Google Scholar] [CrossRef] [PubMed]
- Ona, A.; Luque, D.; Abaitua, F.; Maraver, A.; Caston, J.R.; Rodriguez, J.F. The c-terminal domain of the pVP2 precursor is essential for the interaction between VP2 and VP3, the capsid polypeptides of infectious bursal disease virus. Virology 2004, 322, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Hjalmarsson, A.; Carlemalm, E.; Everitt, E. Infectious pancreatic necrosis virus: Identification of a VP3-containing ribonucleoprotein core structure and evidence for O-linked glycosylation of the capsid protein VP2. J. Virol. 1999, 73, 3484–3490. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Ren, X.; Chen, Y.; Gao, Y.; Wang, N.; Lu, Z.; Gao, L.; Qin, L.; Wang, Y.; Gao, H.; et al. Chondroitin Sulfate N-acetylgalactosaminyltransferase-2 Contributes to the Replication of Infectious Bursal Disease Virus via Interaction with the Capsid Protein VP2. Viruses 2015, 7, 1474-1491. https://doi.org/10.3390/v7031474
Zhang L, Ren X, Chen Y, Gao Y, Wang N, Lu Z, Gao L, Qin L, Wang Y, Gao H, et al. Chondroitin Sulfate N-acetylgalactosaminyltransferase-2 Contributes to the Replication of Infectious Bursal Disease Virus via Interaction with the Capsid Protein VP2. Viruses. 2015; 7(3):1474-1491. https://doi.org/10.3390/v7031474
Chicago/Turabian StyleZhang, Lizhou, Xiangang Ren, Yuming Chen, Yulong Gao, Nian Wang, Zhen Lu, Li Gao, Liting Qin, Yongqiang Wang, Honglei Gao, and et al. 2015. "Chondroitin Sulfate N-acetylgalactosaminyltransferase-2 Contributes to the Replication of Infectious Bursal Disease Virus via Interaction with the Capsid Protein VP2" Viruses 7, no. 3: 1474-1491. https://doi.org/10.3390/v7031474
APA StyleZhang, L., Ren, X., Chen, Y., Gao, Y., Wang, N., Lu, Z., Gao, L., Qin, L., Wang, Y., Gao, H., Li, K., Jiang, L., Cui, H., Liu, C., Zhang, Y., Qi, X., & Wang, X. (2015). Chondroitin Sulfate N-acetylgalactosaminyltransferase-2 Contributes to the Replication of Infectious Bursal Disease Virus via Interaction with the Capsid Protein VP2. Viruses, 7(3), 1474-1491. https://doi.org/10.3390/v7031474